
Microarthropods Living on the Endemic Tree Zelkova abelicea (Ulmaceae) with Particular Attention to Collembola Diversity


 , , ,
, , ,  , ,
, ,
Abstract
1. Introduction
2. Methods
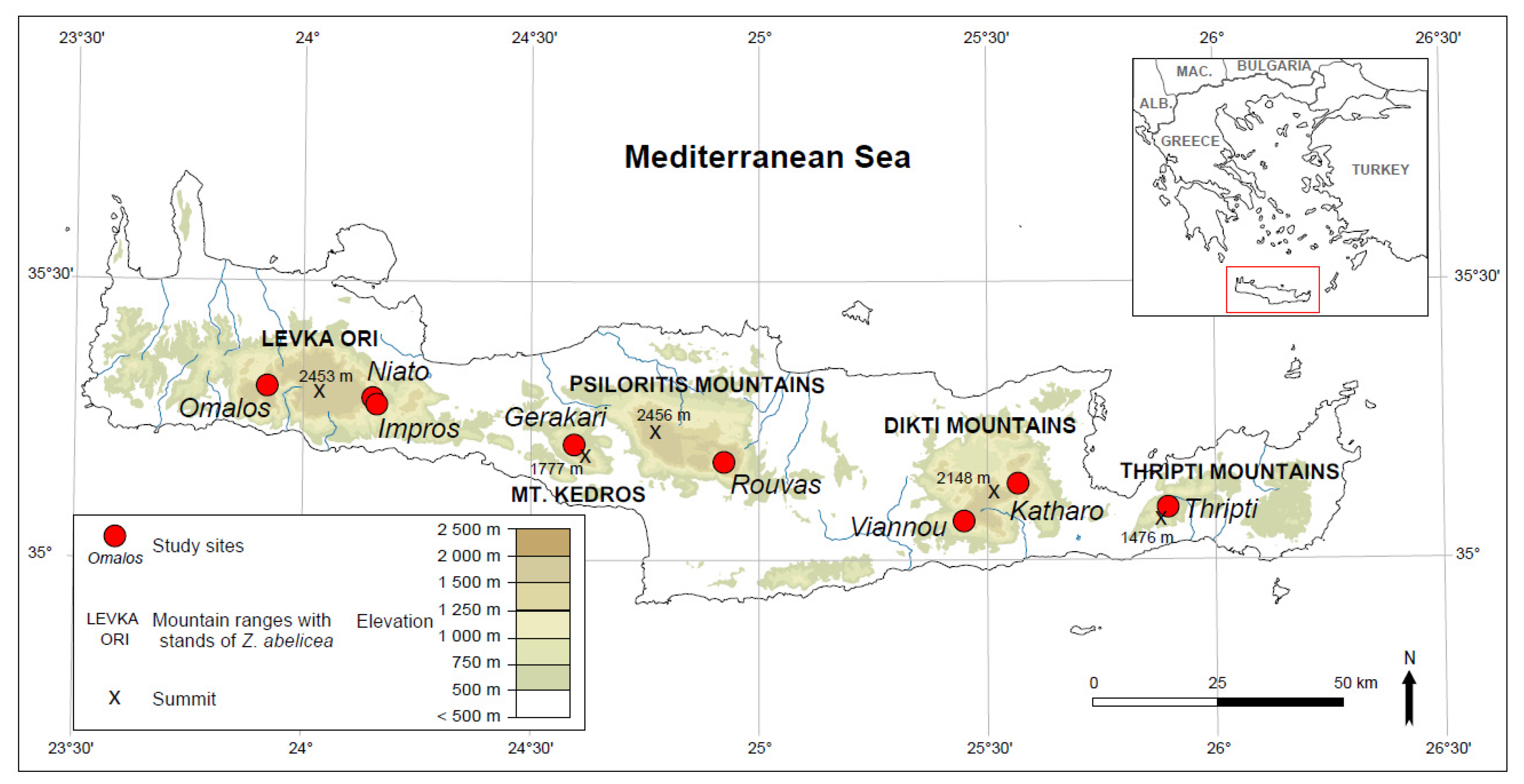
2.1. Field Studies
- Omalos, Levka Ori (Latitude 35, 31901; Longitude 23, 91871), Altitude—1160 m a.s.l., topology: Slope, microhabitat: Bark of arborescent trees, date—21 May 2019, Coll. D. Ghosn;
- Niato, Levka Ori (35, 287527; 24, 145503), 1215 m a.s.l., doline, branches of dwarfed individuals, 21 May 2019, Coll. D. Ghosn;
- Impros, Levka Ori (35, 270546; 24, 15315), 1175 m a.s.l., slope, bark of arborescent trees, 21 May 2019, Coll. D. Ghosn;
- Gerakari, Mt. Kedros (35, 194829; 24, 606713), 1255 m a.s.l., slope, bark of arborescent trees, 11 October 2018, Coll. D.J. Gwiazdowicz;
- Rouvas, Psiloritis Mountains, (35, 164333; 24, 922794), 1320 m a.s.l., slope, bark of arborescent trees, 10 October 2018, Coll. D.J. Gwiazdowicz;
- Viannou, Dikti Mountains, (35, 064291; 25, 469778), 1320 m a.s.l., slope, bark of arborescent trees, 9 October 2018, Coll. D.J. Gwiazdowicz;
- Katharo, Dikti Mountains, (35, 148004; 25, 567558), 1160 m a.s.l., slope, bark of arborescent trees, 9 October 2018, Coll. D.J. Gwiazdowicz;
- Thripti, Thripti Mountains, (35, 080588; 25, 887408), 1150 m a.s.l., doline, branches of dwarfed individuals, 14 May 2019, Coll. D. Ghosn.
2.2. Laboratory Procedures
2.3. Statistical Analyses
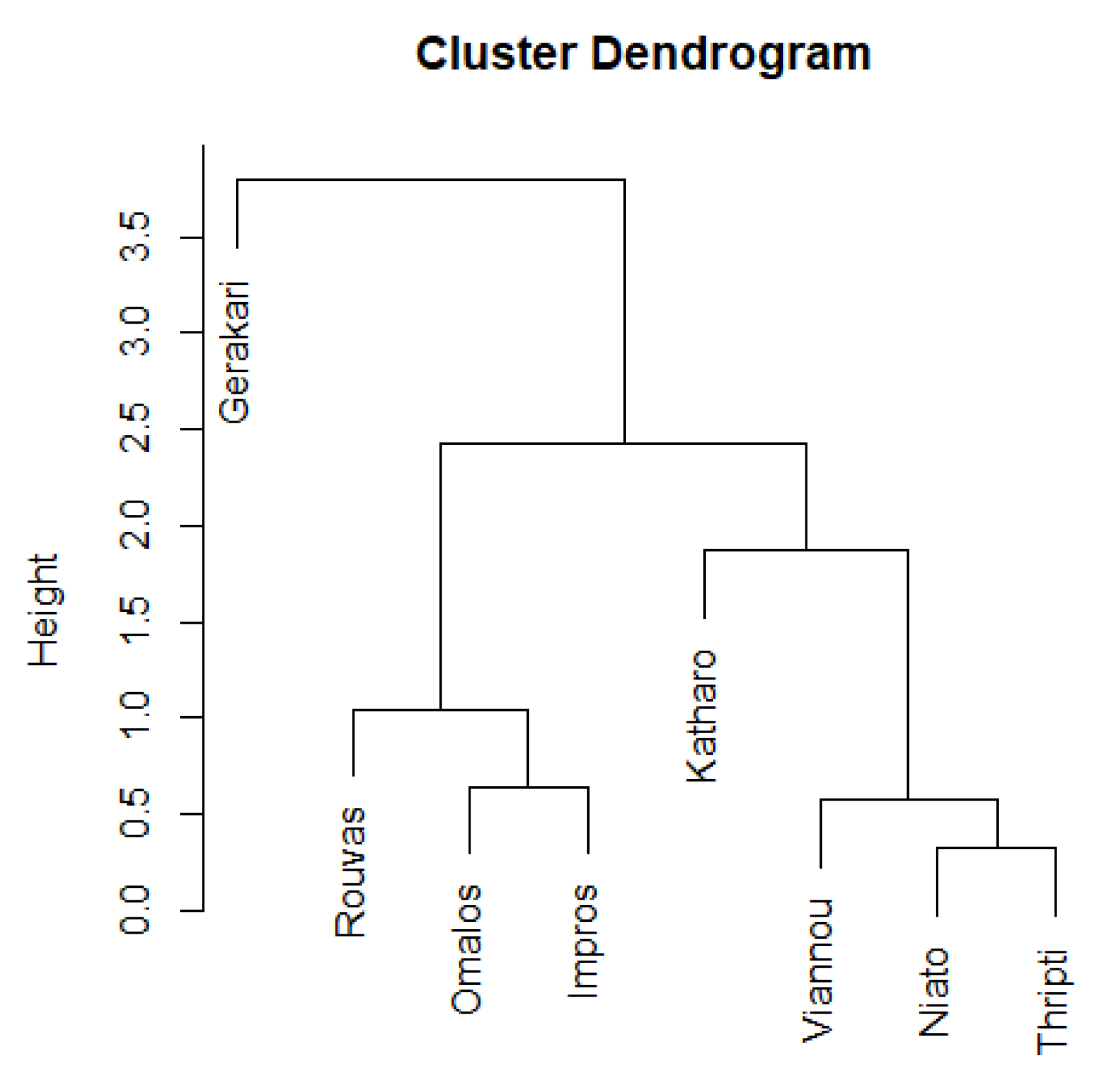
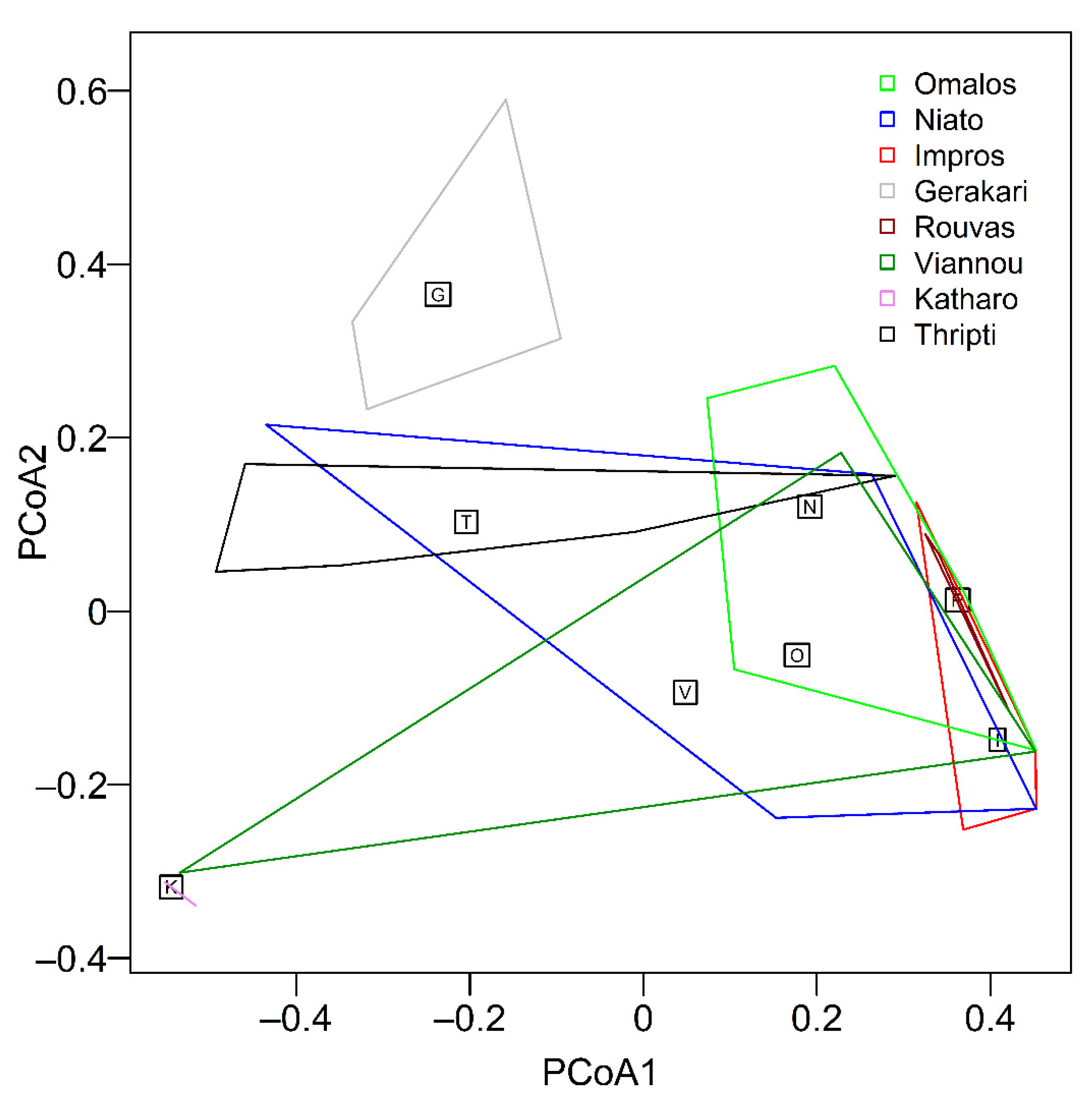
3. Results
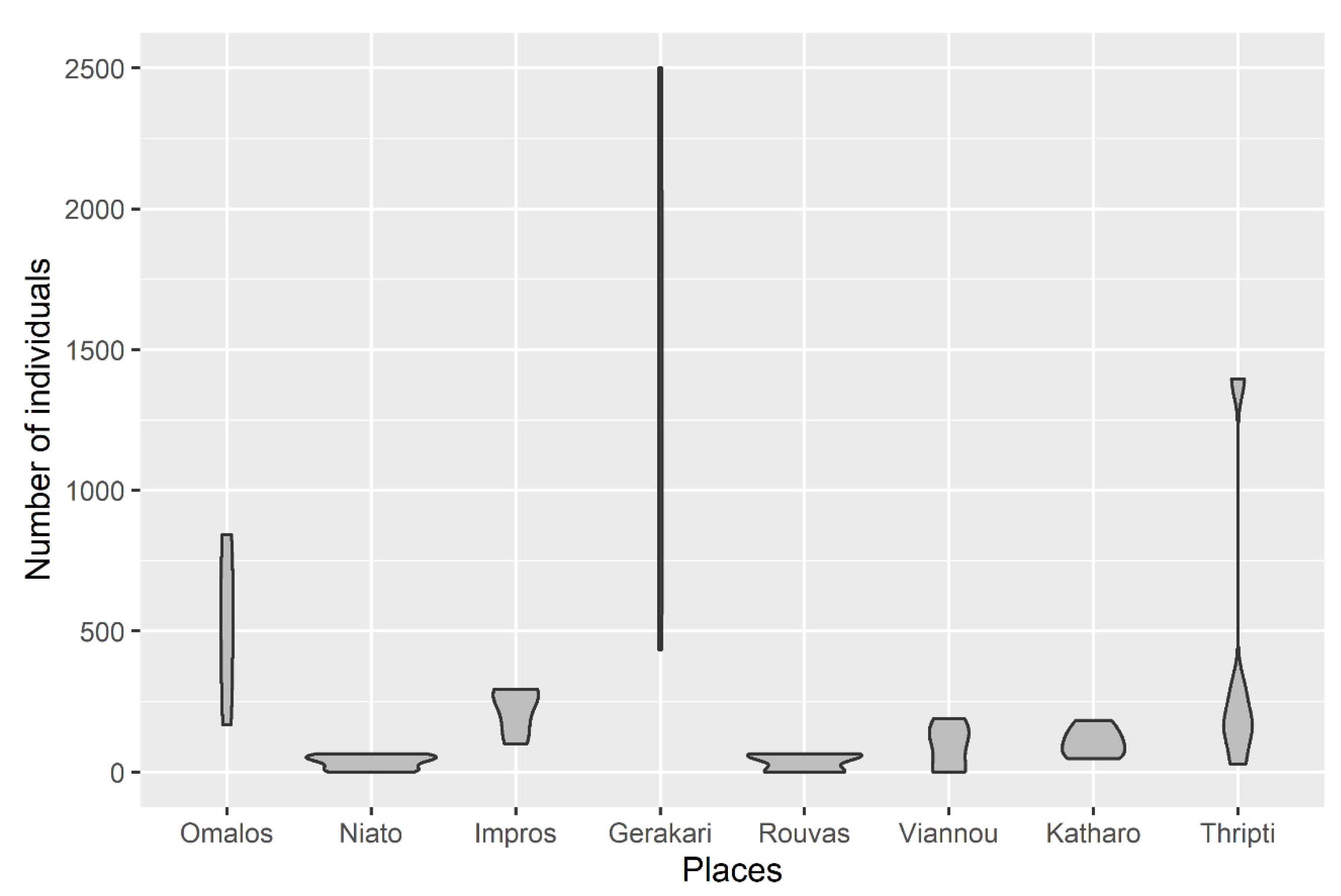
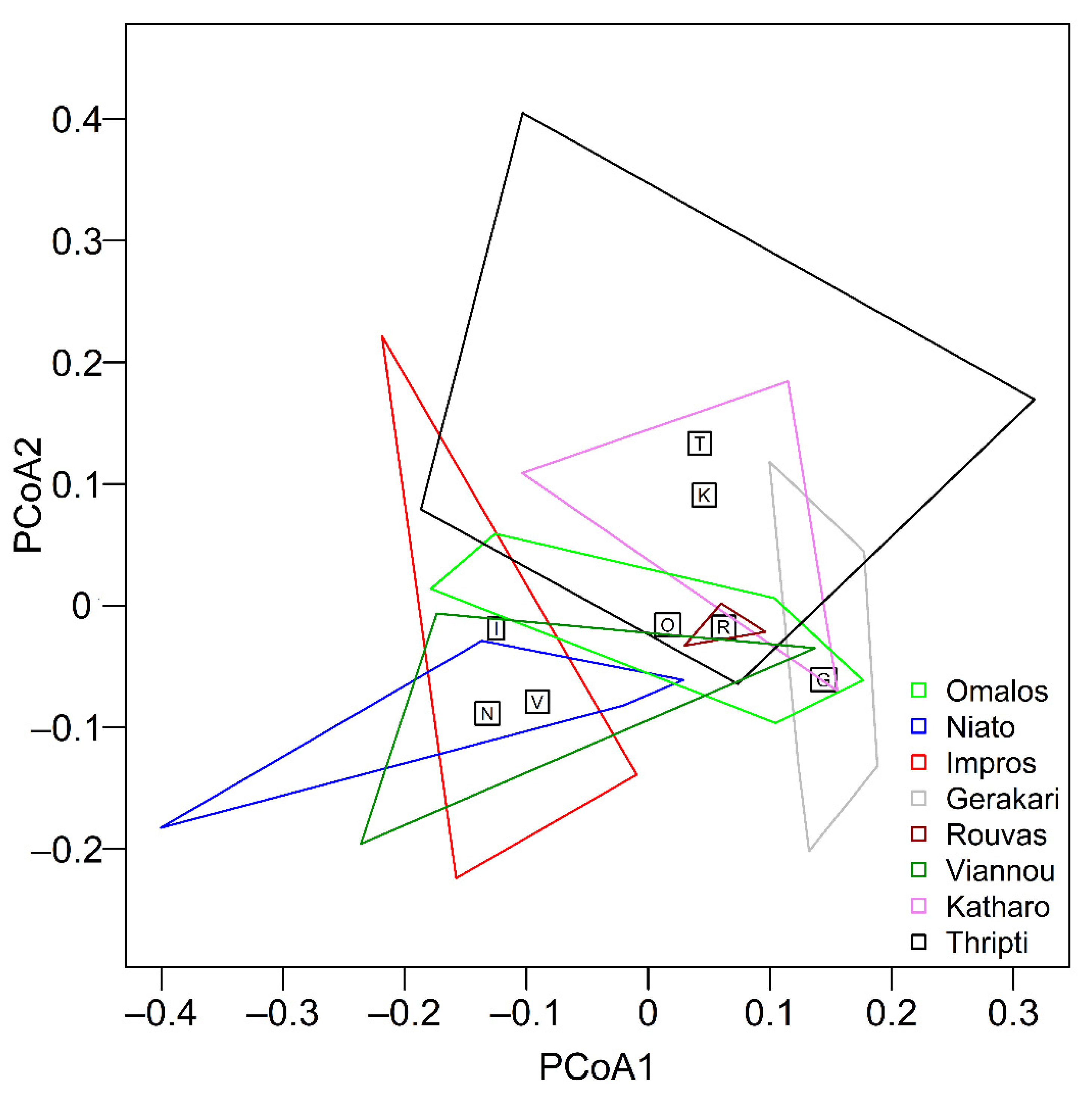
3.1. Diversity of Microarthropod Assemblages in Separate Zelkova abelicea Localities
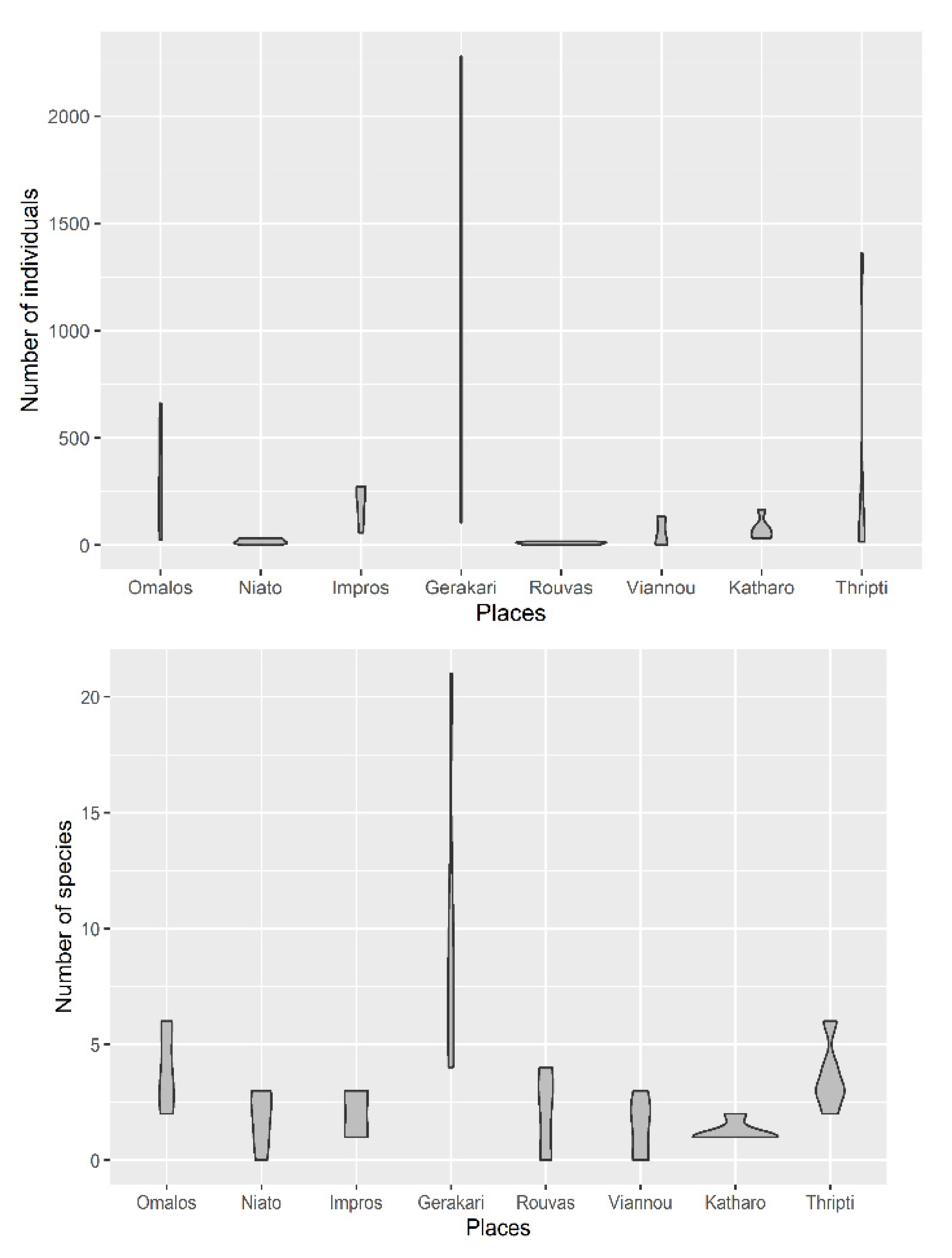
3.2. Diversity of Collembola Communities on Zelkova abelicea Trees
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hampe, A.; Jump, A.S. Climate relicts: Past, present and future. An. Rev. Ecol. Evol. Syst. 2011, 42, 313–333. [Google Scholar] [CrossRef]
- Milne, R.I.; Abbott, R.J. The origin and evolution of Tertiary relict floras. Adv. Bot. Res. 2002, 38, 281–314. [Google Scholar]
- Kozlowski, G.; Gratzfeld, J. Zelkova—An Ancient Tree. Global Status and Conservation Action; Natural History Museum: Fribourg, Switzerland, 2013; pp. 1–60. [Google Scholar]
- Rejžek, M.; Svátek, M.; Šebesta, J.; Adolt, R.; Maděra, P.; Matula, R. Loss of a single tree species will lead to an overall decline in plant diversity: Effect of Dracaena cinnabari Balf. f. on the vegetation of Socotra Island. Biol. Conserv. 2016, 196, 165–172. [Google Scholar] [CrossRef]
- Maděra, P.; Habrová, H.; Šenfeldr, M.; Kholová, I.; Lvončík, S.; Ehrenbergerová, L.; Roth, M.; Nadezhdina, N.; Němec, P.; Rosenthal, J.; et al. Growth dynamics of endemic Dracaena cinnabari Balf of Socotra Island suggest essential elements for a conservation strategy. Biologia 2019, 74, 339–349. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Laurance, W.F. The ecology, distribution, conservation and management of large old trees. Biol. Rev. 2017, 92, 1434–1458. [Google Scholar] [CrossRef] [PubMed]
- Kraus, D.; Bütler, R.; Krumm, F.; Lachat, T.; Larrieu, L.; Mergner, U.; Paillet, Y.; Schuck, A.; Winter, S. Catalogue of Tree Microhabitats: Reference Field List; European Forest Institute: Freiburg, Germany, 2016; pp. 1–16. [Google Scholar]
- Larrieu, L.; Paillet, Y.; Winter, S.; Bütler, R.; Kraus, D.; Krumm, F.; Lachat, T.; Michel, A.K.; Regnery, B.; Vanderkerkhove, K. Tree related microhabitats in temperate and Mediterranean European forests: A hierarchical typology for inventory standardization. Ecol. Indic. 2018, 84, 194–207. [Google Scholar] [CrossRef]
- Mai, D.H. Palaeofloristic changes in Europe and the confirmation of the Arctotertiary-Palaeotropical geofloral concept. Rev. Palaeobot. Palynol. 1991, 68, 29–36. [Google Scholar] [CrossRef]
- Kozlowski, G.; Frey, D.; Fazan, L.; Egli, B.; Pirintsos, S. Zelkova abelicea. The IUCN Red List of Threatened Species. 2012. e.T30319A17701987. Available online: https://www.fr.ch/sites/default/files/2018-12/Kozlowski_et_al._2012a.pdf (accessed on 28 October 2021). [CrossRef]
- Garfì, G.; Pasta, S.; Fazan, L.; Kozlowski, G. Zelkova sicula. The IUCN Red List of Threatened Species. 2017. e.T61678A86134112. Available online: https://www.zelkova.ch/sites/default/files/Garfi%20et%20al.%202017%20IUCN%20assessment%20Zelkova%20sicula.pdf (accessed on 28 October 2021). [CrossRef]
- Barbagallo, S. Zelkovaphis trinacriae, a new Eriosomatine aphid genus and species living on Zelkova in Sicily (Rhynchota: Aphididae). Boll. Zool. Agr. E Bachic. Ser. II 2002, 34, 281–301. [Google Scholar]
- Mazzeo, G.; Nucifora, S.; Russo, A.; Suma, P.; Longo, S. An updated list of the scale insect (Hemiptera Coccomorpha) fauna of some Sicilian endemic plants. Redia 2016, 99, 201–206. [Google Scholar]
- Campo, G.; Mazzeo, G.; Nucifora, S.; Perrotta, G.; Sidoti, A.; Bella, S. Insects and fungi on the relict Zelkova sicula (Rosales, Ulmaceae) in Sicily (Italy): New records and known species in a short review. Redia 2018, 101, 23–30. [Google Scholar] [CrossRef]
- Hsin-Ting, Y.; Chi-Feng, L.; Shipher, W.; Li-Cheng, S.; Ya-Nan, W.; Jui-Chu, Y. Investigation of Insect Feeding Guilds on Zelkova serrata (Ulmaceae) in Wanlong Plantation, Pintung County. J. Exp. For. Natl. Taiwan Univ. 2013, 27, 251–262. [Google Scholar] [CrossRef]
- Ohsawa, M. Life Cycle, Ecological Characteristics, and Control of Trachys yanoi (Coleoptera: Buprestidae), an Important Pest of Zelkova serrata. Insects 2017, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Gol, A.; Sadeghi-Namaghi, H.; de Lillo, E. Two new species of eriophyoid mites (Acari: Trombidiformes: Eriophyoidea) on Zelkova carpinifolia (Ulmaceae) from Iran. Syst. Appl. Acarol. 2018, 23, 393–400. [Google Scholar] [CrossRef]
- Fazan, L.; Stoffel, M.; Frey, D.J.; Pirintsos, S.; Kozlowski, G. Small does not mean young: Age estimation of severely browsed trees in anthropogenic Mediterranean landscapes. Biol. Conserv. 2012, 153, 97–100. [Google Scholar] [CrossRef]
- Kozlowski, G.; Frey, D.; Fazan, L.; Egli, B.; Bétrisey, S.; Gratzfeld, J.; Garfì, G.; Pirintsos, S. The Tertiary relict tree Zelkova abelicea (Ulmaceae): Distribution, population structure and conservation status on Crete. Oryx 2014, 48, 80–87. [Google Scholar] [CrossRef]
- Rackham, O.; Moody, J. The Making of the Cretan Landscape; Manchester University Press: Manchester, UK, 1996; pp. 1–200. [Google Scholar]
- Bauer, E.M.; Bergmeier, E. The mountain woodlands of western Crete—Plant communities, forest goods, grazing impact and conservation. Phytocoenologia 2011, 41, 73–115. [Google Scholar] [CrossRef]
- Stathakis, T.I. Predatory Mites of the Family Phytoseiidae (Acari: Mesostigmata) on Native Plants of Crete. Master’s Thesis, Agricultural University of Athens, Athina, Greece, 2011; pp. 1–138, (In Greek with English abstract). [Google Scholar]
- Hilszczanski, J. New data on the occurrence of Stephanids (Hymenoptera: Stephanidae) in Turkey and Greece. Opole Sci. Soc. Nat. J. 2011, 44, 192–196. [Google Scholar]
- Christe, C.; Kozlowski, G.; Frey, D.; Bétrisey, S.; Maharramova, E.; Garfì, G.; Pirintsos, S.; Naciri, Y. Footprints of past intensive diversification and structuring in the genus Zelkova (Ulmaceae) in south-western Eurasia. J. Biogeogr. 2014, 41, 1081–1093. [Google Scholar] [CrossRef]
- Gisin, H. Collembolenfauna Europas; Muséum D’histoire Naturelle: Geneva, Switzerland, 1960; pp. 1–312. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of this Group of Insects Tribe: Orchesellini; PWN: Krakow, Poland, 1960; pp. 1–151. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of this Group of Insects Tribe: Entomobryini; PWN: Krakow, Poland, 1963; pp. 1–126. [Google Scholar]
- Massoud, Z. Monographie des Neanuridae, Collemboles Poduromorphes à pièces buccales modifiées. In Biologie de l’Amérique Australe; Delamare Deboutteville, C., Rapoport, E.H., Eds.; CNRS: Paris, France, 1967; pp. 7–399. [Google Scholar]
- Ellis, W.N. Autumn fauna of Collembola from Central Crete. Tijdschrif. Entomol. 1976, 119, 221–236. [Google Scholar]
- Kaprus’, I.J.; Weiner, W.M. Two interesting species of Onychiurinae (Collembola) from Ukraine and some remarks on Allaphorura franzi (STACH, 1946). Acta Zool. Cracov. 1994, 37, 59–64. [Google Scholar]
- Jordana, R.; Arbea, J.I.; Simón, C.; Luciáñez, M.J. Collembola, Poduromorpha. In Fauna Ibérica; CSIC: Madrid, Spain, 1997; pp. 1–807. [Google Scholar]
- Pomorski, R.J.; Skarżyński, D. A redescription of Metaphorura denisi Simon Benito, 1985 (Collembola, Onychiuridae, Tullbergiinae). Genus 1997, 8, 489–496. [Google Scholar]
- Simon Benito, J.C.; Deharveng, L. Une nouvelle Friesea du department du Var (Collembola, Neanuridae). Bull. Soc. Entomol. Fran. 1997, 102, 453–455. [Google Scholar] [CrossRef]
- Fjellberg, A. The Collembola of Fennoscandia and Denmark. Part 1 Poduromorpha. Fauna Entomol. Scand. 1998, 35, 1–184. [Google Scholar]
- Fjellberg, A. The Collembola of Fennoscandia and Denmark. Part II: Entomobryomorpha and Symphypleona. Fauna Entomol. Scand. 2007, 42, 1–265. [Google Scholar]
- Pomorski, R.J. Onychiurinae of Poland (Collembola: Onychiuridae). Genus 1998, 9, 1–201. [Google Scholar]
- Bretfeld, G. Symphypleona. In Synopses on Palaearctic Collembola; Staatliches Museum für Naturkunde Görlitz, Germany, 1999; pp. 1–318.
- Carapelli, A.; Frati, F.; Fanciulli, P.P.; Dallai, R. Taxonomic revision of 14 southwestern European species of Isotomurus (Collembola, Isotomidae), with description of four new species and the designation of the neotype for I. palustris. Zool. Scr. 2001, 3, 115–143. [Google Scholar] [CrossRef]
- Potapov, M. Isotomidae. In Synopses on Palaearctic Collembola; Dunger, W., Ed.; Staatliches Museum für Naturkunde Görlitz: Görlitz, Germany, 2001; pp. 1–603. [Google Scholar]
- Thibaud, J.M.; Schulz, H.J.; Assalino, M.M.D.G. Hypogastruridae. In Synopses on Palaearctic Collembola; Dunger, W., Ed.; Staatliches Museum für Naturkunde Görlitz: Görlitz, Germany, 2004; pp. 1–287. [Google Scholar]
- Gioia Cipola, N.; Arbea, J.; Baquero, E.; Jordana, R.; Wellington de Mmorais, J.; Cavalcante Bbellini, B. The survey of Seira Lubbock, 1870 (Collembola, Entomobryidae, Seirinae) from Iberian Peninsula and Canary Islands, including three new species. Zootaxa 2018, 4458, 1–66. [Google Scholar] [CrossRef]
- Skarżyński, D.; Piwnik, A.; Porco, D. Integrating morphology and DNA barcodes for species delimitation within the species complex Xenylla maritima (Collembola: Hypogastruridae). Arthr. Sys. Phylog. 2018, 76, 31–43. [Google Scholar]
- Lafooraki, E.Y.; Hajizadeh, J.; Antipova, M.; Kremenitsa, A.; Shayanmehr, M.; Potapov, M.; Hosseini, R. Vertagopus (Collembola, Isotomidae) of Iran and Caucasus. Zootaxa 2020, 4786, 574–582. [Google Scholar] [CrossRef]
- Anderberg, M.R. Cluster Analysis for Applications; Academic Press: New York, NY, USA, 1973; ISBN 9781483191393. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd English ed.; Elsevier Science BV: Amsterdam, The Netherlands, 2012; ISBN 9780444538697. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecology 1999, 69, 1–24. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Pielou, E.C. The Usefulness of Ecological Models: A Stock-Taking. Quart. Rev. Biol. 1981, 56, 17–31. [Google Scholar] [CrossRef]
- Odum, E.P. Fundamentals of Ecology, 3th ed.; Toppan Compani Ltd.: London, UK, 1971; pp. 1–574. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; ISBN 3900051070. Available online: http://www.R-project.org/ (accessed on 28 October 2021).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; et al. Vegan: Community Ecology Package. R Package Version 2.10. 2020. Available online: https://cran.ism.ac.jp/web/packages/vegan/vegan.pdf (accessed on 28 October 2021).
- Simon Benito, J.C. Colembolos de suelos de sabinar en la provincia de Guadalajara. Eos 1985, 61, 293–318. [Google Scholar]
- Schulz, H.-J.; Lymberakis, P. First contribution to the knowledge of the Collembola fauna of the White Mountains (Lefká Óri) in West Crete (Insecta, Collembola). Senckenberg. Biol. 2006, 86, 229–234. [Google Scholar]
- Schulz, H.-J. Second contribution to the knowledge of the springtail fauna of the White Mountains (Lefká Óri) in West Crete (Insecta: Collembola). Mitt. Int. Entomol. Ver. Frankfurt a.M. 2007, 32, 175–183. [Google Scholar]
- Schulz, H.-J. A contribution to the Collembola fauna of Crete (Insecta: Collembola). Mitt. Int. Entomol. Ver. 2010, 35, 101–110. [Google Scholar]
- Schulz, H.-J.; Burkhardt, U. Interessante Collembolennachweise aus Ostkreta (Hexapoda, Collembola). Linz. Biolog. Beitr. 2012, 44, 1669–1676. [Google Scholar]
- Arbea, J.I. Revisión de las especies del grupo de Folsomides angularis (Axelson, 1905), con descripción de dos nuevas especies para la fauna europea y del norte de África (Collembola, Isotomidae). Arq. Entomol. 2015, 13, 413–428. [Google Scholar]
- Groombridge, B.; Jenkins, M.D. World Atlas of Biodiversity; UNEP World Conservation Monitoring Centre; University of California Press: Berkeley, CA, USA, 2002; pp. 1–340. [Google Scholar]
- Martinez, J.J.I.; Raz, R.; Mgocheki, N. Differential spatial distribution of arthropods under epiphytic lichens on trees. J. Insect Biodivers. 2014, 2, 1–7. [Google Scholar] [CrossRef][Green Version]
- Marcelino, J.; Borges, P.A.V.; Borges, I.; Pereira, E.; Santos, V.; Soares, A.O. Standardised arthropod (Arthropoda) inventory across natural and anthropogenic impacted habitats in the Azores archipelago. Biodivers. Dat. J. 2021, 9, e62157. [Google Scholar] [CrossRef] [PubMed]
- Legakis, A.; Kypriotakis, Z. A biogeographical analysis of the island of Crete, Greece. J. Biogeogr. 1994, 21, 441–445. [Google Scholar] [CrossRef]
- Jahn, R.; Schönfelder, P. Exkursionsflora für Kreta; Ulmer: Stuttgart, Germany, 1995. [Google Scholar]
- Poulakakis, N.; Lymberakis, P.; Antoniou, A.; Chalkia, D.; Zouros, E.; Mylonas, M.; Valakos, E. Molecular phylogeny and biogeography of the wall-lizard Podarcis erhardii (Squamata: Lacertidae). Mol. Phylogen. Evol. 2003, 28, 38–46. [Google Scholar] [CrossRef]
- Chatzaki, M.; Arnedo, M.A. Taxonomic revision of the epigean representatives of the spider subfamily Harpacteinae (Araneae: Dysderidae) on the island of Crete. Zootaxa 2006, 1169, 1–32. [Google Scholar] [CrossRef]
- Bergmeier, E. New floristic records, confirmations and other phytogeographical notes from Crete (Greece). Willdenowia 2011, 41, 167–177. [Google Scholar] [CrossRef]
- Trigas, P.; Panitsa, M.; Tsiftsis, S. Elevational gradient of vascular plant species richness and endemism in Crete—The effect of post-isolation mountain uplift on a continental island system. PLoS ONE 2013, 8, e59425. [Google Scholar] [CrossRef]
- Assing, V. Monography of the Staphylinidae of Crete (Greece). Part, I. Diversity and endemisms (Insecta: Coleoptera). Contr. Entomol. 2019, 69, 197–239. [Google Scholar] [CrossRef]
- Mitsainas, G.; Vohralik, V.; Kennerley, R. Crocidura zimmermanni. The IUCN Red List of Threatened Species. 2019. e.T5588A90818329. Available online: https://www.iucnredlist.org/species/5588/90818329 (accessed on 28 October 2021). [CrossRef]
- Gwiazdowicz, D.J. Sejoidea, Antennophoroidea, Celaenopsoidea, Microgynioidea (Acari, Mesostigmata) of Poland; Bogucki Wydawnictwo Naukowe: Poznan, Poland, 2010; pp. 1–142. [Google Scholar]
- Goedecke, F.; Bergmeier, E. Ecology and potential distribution of the Cretan endemic tree species Zelkova abelicea. J. Medit. Ecol. 2018, 16, 15–26. [Google Scholar]
- Varouchakis, E.A.; Corzo, G.A.; Karatzas, G.P.; Kotsopoulou, A. Spatio-temporal analysis of annual rainfall in Crete, Greece. Act. Geophysic. 2018, 66, 319–328. [Google Scholar] [CrossRef]
- Agou, V.D.; Varouchakis, E.A.; Hristopulos, D.T. Geostatistical analysis of precipitation in the island of Crete (Greece) based on a sparse monitoring network. Environ. Monit. Asses. 2019, 191, 353. [Google Scholar] [CrossRef] [PubMed]
- Ellis, W.N. The spring fauna of Collembola (Insecta) from Rhodos, with descriptions of some new taxa. Beaufortia 1974, 22, 105–152. [Google Scholar]
- Kaprus’, I.J.; Shrubovych, J.J.; Tarashchuk, M.V. Catalogue of the Collembola and Protura of Ukraine; SNHMU, NASU: L’viv, Ukraine, 2006; pp. 1–164. [Google Scholar]
- Fazan, L.; Gwiazdowicz, D.J.; Fragnière, Y.; Faltynowicz, W.; Ghosn, D.; Remoundou, I.; Rusinska, A.; Urbanski, P.; Pasta, S.; Garfi, G.; et al. Factors influencing the diversity and distribution of epiphytic lichens and bryophytes on the relict tree Zelkova abelicea (Lam.) Boiss. (Ulmaceae). Lichenologist 2022. in print. [Google Scholar]
- Pflug, A.; Wolters, V. Influence of drought and litter age on Collembola communities. Eur. J. Soil Biol. 2001, 37, 305–308. [Google Scholar] [CrossRef]
- Rusek, J. Microhabitats of Collembola (Insecta: Entognatha) in beech and spruce forests and their influence on biodiversity. Eur. J. Soil Biol. 2001, 37, 237–244. [Google Scholar] [CrossRef]
- Mehrabi, Z.; Slade, E.M.; Solis, A.; Mann, D.J. The importance of microhabitat for biodiversity sampling. PLoS ONE 2014, 9, e114015. [Google Scholar] [CrossRef]
- Benito, J.C.S.; Sanchez, M.J.L. Ecology of soil springtails (Collembola, Insecta) from pine woods and Rhododendron shrublands in the Central and Eastern Pyrenees (North Spain). Pedobiologia 2000, 44, 430–441. [Google Scholar] [CrossRef]
- Rossetti, I.; Bagella, S.; Cappai, C.; Caria, M.C.; Lai, R.; Roggero, P.P.; Martins da Silva, P.; Sousa, J.P.; Querner, P.; Seddaiu, G. Isolated cork oak trees affect soil properties and biodiversity in a Mediterranean wooded grassland. Agri. Ecosyst. Environ. 2015, 202, 203–216. [Google Scholar] [CrossRef]
- Welti, E.; Helzer, C.; Joern, A. Impacts of plant diversity on arthropod communities and plant-herbivore network architecture. Ecosphere 2017, 8, e01983. [Google Scholar] [CrossRef]
- Dombos, M. Collembola of loess grassland. Effects of grazing and landscape on community composition. Soil Biol. Biochem. 2001, 33, 2037–2045. [Google Scholar] [CrossRef]
- Sousa, J.P.; Vingada, J.V.; Barrocas, H.; da Cama, M.M. Effects of introduced exotic tree species on Collembola communities: The importance of management techniques. Pedobiologia 1997, 41, 145–153. [Google Scholar]
- Sousa, J.P.; da Gama, M.M.; Pinto, C.; Keating, A.; Calhôa, F.; Lemos, M.; Castro, C.; Luz, T.; Leitao, P.; Dias, S. Effects of land-use on Collembola diversity patterns in a Mediterranean landscape. Pedobiologia 2004, 48, 609–622. [Google Scholar] [CrossRef]
- Lauga-Reyrel, F.; Deconchat, M. Diversity within the Collembola community in fragmented coppice forests in south-western France. Eur. J. Soil Biol. 1999, 35, 177–187. [Google Scholar] [CrossRef]
- Alvarez, T.; Frampton, G.K.; Goulson, D. Epigeic Collembola in winter wheat under organic, integrated and conventional farm management regimes. Agr. Ecosys. Environ. 2001, 83, 95–110. [Google Scholar] [CrossRef]
- Blaum, N.; Seymour, C.; Rossmanith, E.; Schwager, M.; Jeltsch, F. Changes in arthropod diversity along a land use driven gradient of shrub cover in savanna rangelands: Identification of suitable indicators. Biodivers. Conserv. 2009, 18, 1187–1199. [Google Scholar] [CrossRef]
- Ganser, D.; Denmead, L.H.; Clough, Y.; Buchori, D.; Tscharntke, T. Local and landscape drivers of arthropod diversity and decomposition processes in oil palm leaf axils. Agri. For. Entomol. 2017, 19, 60–69. [Google Scholar] [CrossRef]
- Garcia-Tejero, S.; Taboada, A.; Tarrega, R.; Salgado, J.M. Land use changes and ground dwelling beetle conservation in extensive grazing dehesa systems of north-west Spaine. Biol. Conserv. 2013, 161, 58–66. [Google Scholar] [CrossRef]
- Meloni, F.; Civieta, B.F.; Zaragoza, J.A.; Moraza, M.L.; Bautista, S. Vegetation pattern modulates ground arthropod diversity in semi-arid Mediterranean steppes. Insects 2020, 11, 59. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Systematics/Mountains | Levka Ori | Kedros | Psiloritis | Dikti | Thripti | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Group of Arthropods/ Localities | Omalos | Niato | Impros | Gerakari | Rouvas | Viannou | Katharo | Thripti | Mean ± SE (min, max) Number of Specimens per Tree | |
| Arachnida | Araneae | 109 | 16 | 10 | 15 | 12 | 40 | 2 | 8 | 5.3 ± 1.6 (0, 52) |
| Pseudoscorpiones | 2 | 0 | 4 | 5 | 0 | 0 | 0 | 9 | 0.5 ± 0.2 (0, 6) | |
| Acari | 593 | 44 | 55 | 1248 | 45 | 61 | 48 | 143 | 55.9 ± 16.9 (0, 427) | |
| Myriapoda | 7 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0.3 ± 0.2 (0, 5) | |
| Collembola | 1521 | 84 | 951 | 5325 | 40 | 283 | 355 | 1726 | 258.0 ± 76.1 (0, 2279) | |
| Insecta | Coleoptera | 2 | 0 | 0 | 22 | 1 | 8 | 1 | 17 | 1.3 ± 0.5 (0, 15) |
| Dermaptera | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0.1 ± 0.1 (0, 3) | |
| Diptera | 0 | 0 | 0 | 8 | 0 | 0 | 5 | 1 | 0.4 ± 0.2 (0, 8) | |
| Entognatha | 12 | 0 | 20 | 25 | 0 | 0 | 3 | 15 | 1.9 ± 0.8 (0, 20) | |
| Hemiptera | 4 | 0 | 0 | 4 | 2 | 1 | 1 | 7 | 0.5 ± 0.2 (0, 3) | |
| Heteroptera | 0 | 0 | 0 | 3 | 1 | 0 | 1 | 9 | 0.4 ± 0.2 (0, 9) | |
| Hymenoptera | 190 | 0 | 25 | 27 | 0 | 0 | 2 | 1 | 6.1 ± 3.3 (0, 102) | |
| Lepidoptera | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0.05 ± 0.03 (0, 1) | |
| Psocoptera | 122 | 13 | 18 | 53 | 65 | 37 | 36 | 78 | 10.6 ± 2.1 (0, 51) | |
| Rhaphidioptera | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0.03 ± 0.03 (0, 1) | |
| Thysanoptera | 13 | 15 | 16 | 20 | 12 | 13 | 54 | 20 | 4.1 ± 1.0 (0, 30) | |
| TOTAL | 2575 | 173 | 1104 | 6760 | 178 | 443 | 509 | 2034 | 345.3 ± 86.8 (0, 2279) | |
| Mean ± SE (min, max) number of specimens per tree | 515.0 ± 120.9 (167, 844) | 34.6 ± 12.35 (0, 64) | 220.8 ± 38.0 (99, 294) | 1352.0 ± 402.3 (435, 2501) | 35.6 ± 14.7 (0, 64) | 88.6 ± 38.1 (0, 189) | 107.8 ± 24.3 (47, 183) | 408.0 ± 250.4 (29, 1396) | ||
| Mountain | Levka Ori | Kedros | Psiloritis | Dikti | Thripti | ||||
|---|---|---|---|---|---|---|---|---|---|
| Site | Omalos | Niato | Impros | Gerakari | Rouvas | Viannou | Katharo | Thripti | |
| 1. | Hypogastrura cf. gisini ** | 2 | |||||||
| 2. | Xenylla sp. 1 (maritima complex) ** | 1436 | 40 | 933 | 25 | 139 | 58 | ||
| 3. | Xenylla sp. 2 (maritima complex) ** | 38 | 24 | 8 | 3233 | 6 | 55 | ||
| 4. | Xenylla maritima | 1 | 15 | 1759 | 132 | 354 | 1583 | ||
| 5. | Protanura sp. ** | 1 | |||||||
| 6. | Deutonura sp. ** | 2 | |||||||
| 7. | Endonura sp. ** | 1 | |||||||
| 8. | Friesea cf. cassagnaui ** | 1 | |||||||
| 9. | Friesea sp. ** | 4 | |||||||
| 10. | Pseudachorutella sp. ** | 2 | |||||||
| 11. | Protaphorura aurantiaca | 4 | |||||||
| 12. | Thalassaphorura franzi * | 2 | 1 | ||||||
| 13. | Metaphorura affinis | 1 | |||||||
| 14. | Anurophorinae 1 * | 2 | 1 | ||||||
| 15. | Anurophorinae 2 * | 1 | |||||||
| 16. | Folsomia ksenemani | 2 | 1 | ||||||
| 17. | Folsomia quadrioculata * | 36 | |||||||
| 18. | Hemisotoma pontica | 6 | |||||||
| 19. | Isotoma sp. | 2 | |||||||
| 20. | Isotomurus fucicolus * | 3 | |||||||
| 21. | Uzelia cf. kuehnelti ** | 8 | 4 | 7 | 5 | 7 | 1 | 10 | |
| 22. | Vertagopus arboreus * | 6 | |||||||
| 23. | Vertagopus cf. persicus ** | 72 | |||||||
| 24. | Entomobrya handschini | 25 | |||||||
| 25. | Entomobrya multifasciata | 4 | 1 | 77 | 3 | 5 | |||
| 26. | Lepidocyrtus lanuginosus | 1 | |||||||
| 27. | Lepidocyrtus cf. lignorum | 3 | |||||||
| 28. | Lepidocyrtus sp. 1 | 2 | |||||||
| 29. | Lepidocyrtus sp. 2 | 2 | |||||||
| 30. | Orchesella taurica * | 1 | 96 | ||||||
| 31. | Pseudosinella octopunctata | 6 | 1 | ||||||
| 32. | Seira ferrarii * | 5 | 4 | 14 | |||||
| 33. | Sminthurinus alpinus bisetosus | 1 | |||||||
| TOTAL SPECIES | 9 | 5 | 5 | 27 | 4 | 6 | 2 | 7 | |
| TOTAL SPECIMENS | 1521 | 84 | 951 | 5325 | 40 | 283 | 355 | 1726 | |
| Average number of species | 4.00 ± 0.84 (2, 6) | 1.80 ± 0.58 (0, 3) | 2.00 ± 0.45 (1, 3) | 9.60 ± 3.11 (4, 21) | 2.00 ± 0.84 (0, 4) | 1.4 ± 0.6 (0, 3) | 1.20 ± 0.20 (1, 2) | 3.60 ± 0.68 (2, 6) | |
| Average number of specimens | 304.2 ± 117.4 (24, 661) | 16.8 ± 6.04 (0, 33) | 190.2 ± 41.38 (57, 273) | 1065 ± 392.4 (105, 2279) | 8 ± 3.51 (0, 18) | 56.6 ± 31.34 (0, 133) | 77 ± 24.06 (32, 165) | 346.4 ± 256.2 (16, 1359) | |
| Simpson Index | 0.18 ± 0.11 (0.01, 0.61) | 0.37 ± 0.13 (0.0, 0.53) | 0.05 ± 0.02 (0.0, 0.11) | 0.47 ± 0.12 (0.07, 0.8) | 0.52 ± 0.05 (0.43, 0.57) | 0.23 ± 0.21) (0.01, 0.65) | 0.0 ± 0.002 (0.0, 0.2) | 0.32 ± 0.12 (0.04, 0.77) | |
| Pielou’s Index | 0.25 ± 0.13 (0.03, 0.72) | 0.82 ± 0.05 (0.75, 0.92) | 0.19 ± 0.04 (0.11, 0.23) | 0.44 ± 0.09 (0.13, 0.7) | 0.77 ± 0.04 (0.69, 0.83) | 0.37 ± 0.30 (0.06, 0.98) | 0.09 ± NA (0.09, 0.09) | 0.48 ± 0.14 (0.11, 0.88) | |
| Dominance Index | 0.82 ± 0,11 (0.39, 0.99) | 0.63 ± 0,13 (0.47, 1.00)) | 0.95 ± 0.02 (0.89, 1.00) | 0.53 ± 0.12 (0.2, 0.93) | 0.48 ± 0.05 (0.43, 0.57) | 0.77 ± 0.21 (0.35, 0.99) | 1.0 ± 0.004 (0.98, 1.00) | 0.68 ± 0.12 (0.23, 0.96) | |
| Stat | p Value | |
|---|---|---|
| Group Gerakari | ||
| Orchesella taurica Stach, 1960 | 0.809 | 0.0020 |
| Lepidocyrtus cf. lignorum (Fabricius, 1793) | 0.742 | 0.0120 |
| Vertagopus cf. persicus Potapov, Yoosefi & Shayanmehr, 2020 | 0.717 | 0.0143 |
| Group Gerakari + Rouvas | ||
| Entomobrya multifasciata (Tullberg, 1871) | 0.632 | 0.0150 |
| Group Katharo + Thripti | ||
| Xenylla maritima Tullberg, 1869 | 0.682 | 0.0037 |
| Group Gerakari + Niato + Thripti | ||
| Xenylla sp. 2 (maritima complex) | 0.621 | 0.0160 |
| Group Impros + Niato + Omalos + Rouvas + Viannou | ||
| Xenylla sp. 1 (maritima complex) | 0.743 | 0.0003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gwiazdowicz, D.J.; Skarżyński, D.; Fazan, L.; Fragnière, Y.; Ghosn, D.; Kozlowski, G.; Kuźmiński, R.; Remoundou, I.; Zawieja, B. Microarthropods Living on the Endemic Tree Zelkova abelicea (Ulmaceae) with Particular Attention to Collembola Diversity. Forests 2022, 13, 195. https://doi.org/10.3390/f13020195
Gwiazdowicz DJ, Skarżyński D, Fazan L, Fragnière Y, Ghosn D, Kozlowski G, Kuźmiński R, Remoundou I, Zawieja B. Microarthropods Living on the Endemic Tree Zelkova abelicea (Ulmaceae) with Particular Attention to Collembola Diversity. Forests. 2022; 13(2):195. https://doi.org/10.3390/f13020195
Chicago/Turabian StyleGwiazdowicz, Dariusz J., Dariusz Skarżyński, Laurence Fazan, Yann Fragnière, Dany Ghosn, Gregor Kozlowski, Robert Kuźmiński, Ilektra Remoundou, and Bogna Zawieja. 2022. "Microarthropods Living on the Endemic Tree Zelkova abelicea (Ulmaceae) with Particular Attention to Collembola Diversity" Forests 13, no. 2: 195. https://doi.org/10.3390/f13020195
APA StyleGwiazdowicz, D. J., Skarżyński, D., Fazan, L., Fragnière, Y., Ghosn, D., Kozlowski, G., Kuźmiński, R., Remoundou, I., & Zawieja, B. (2022). Microarthropods Living on the Endemic Tree Zelkova abelicea (Ulmaceae) with Particular Attention to Collembola Diversity. Forests, 13(2), 195. https://doi.org/10.3390/f13020195