
Analysis of the Alternative Splicing Events of Exogenous δ-Aminolevulinic Acid under NaCl Stress in Wild Jujube Seedlings

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.1.1. Culture Conditions
2.1.2. Treatment of the Experimental Materials
2.2. Measurement of the Contents of Chlorophyll and Its Intermediates
2.3. RNA Extraction, Library Construction and Sequencing
2.4. Measurement of the Expression of Genes Related to Chlorophyll Biosynthesis
2.5. Statistical Analysis
2.5.1. Identification of Alternative Splicing Events
2.5.2. Functional Annotation and Enrichment Analysis of the Alternative Splice Genes
3. Results
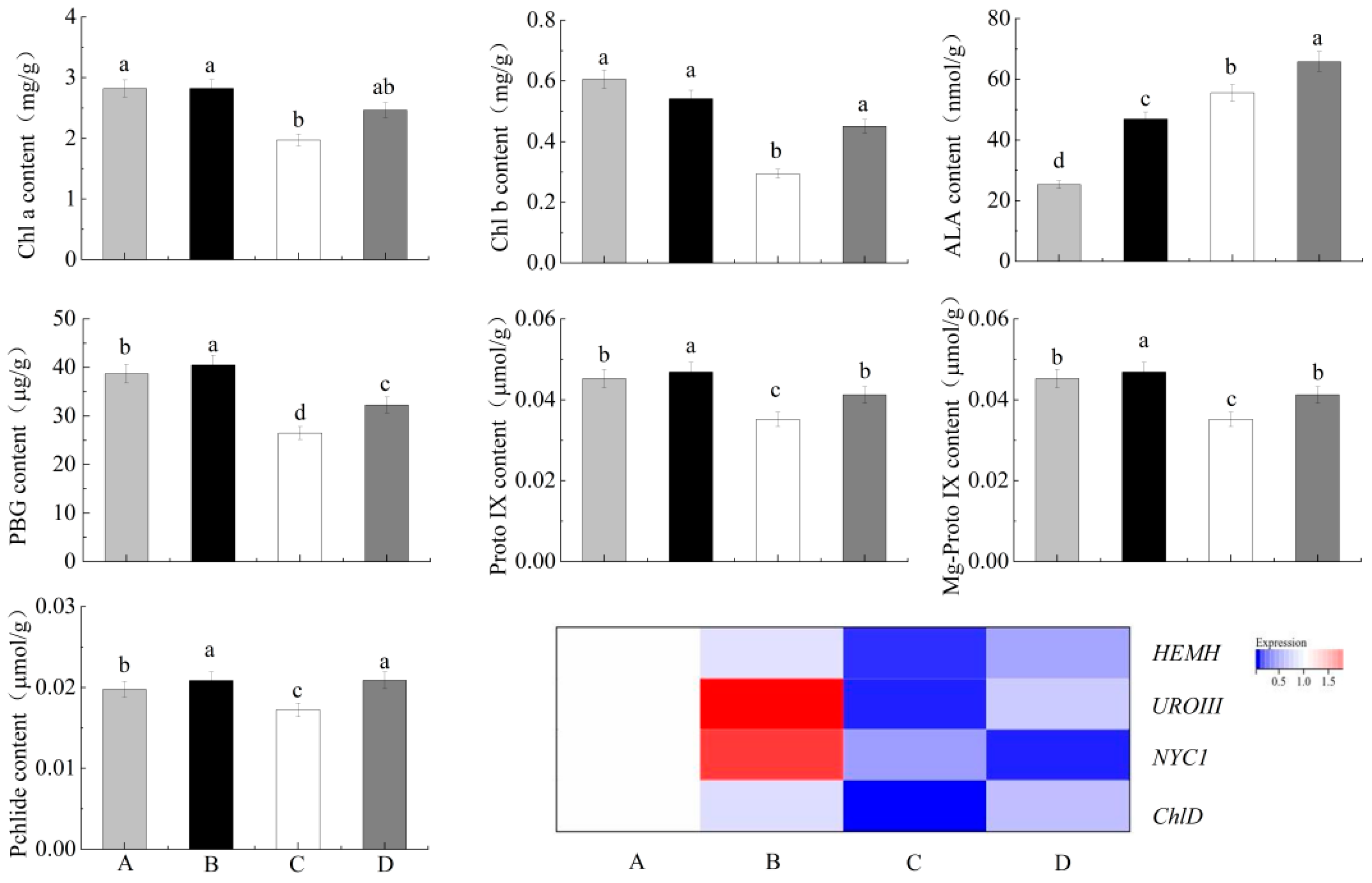
3.1. Effect of Exogenous ALA on the Chlorophyll Content of Jujube Seedlings under NaCl Stress
3.2. Sequencing Quality Analysis
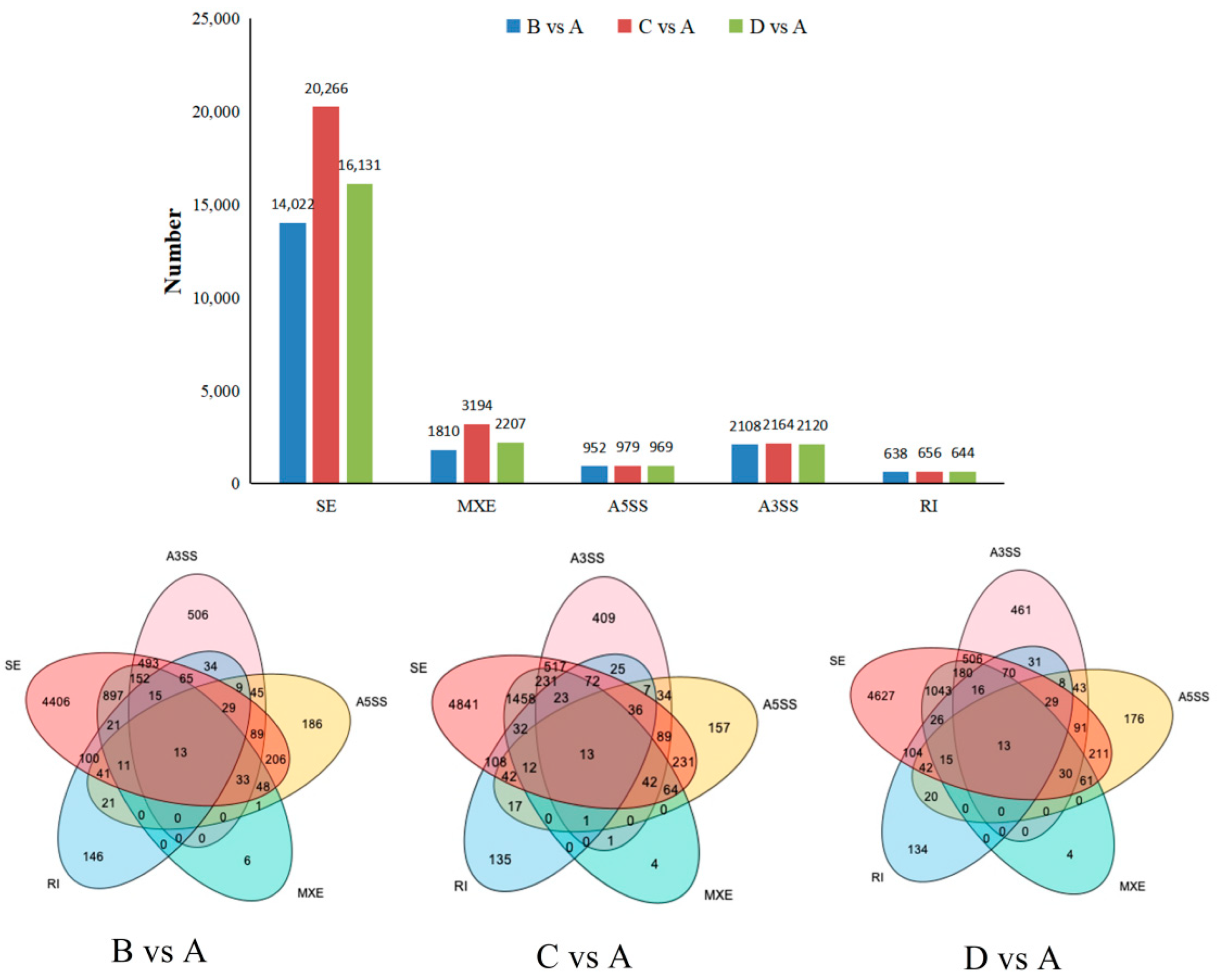
3.3. Identification of Alternative Splicing Event Types
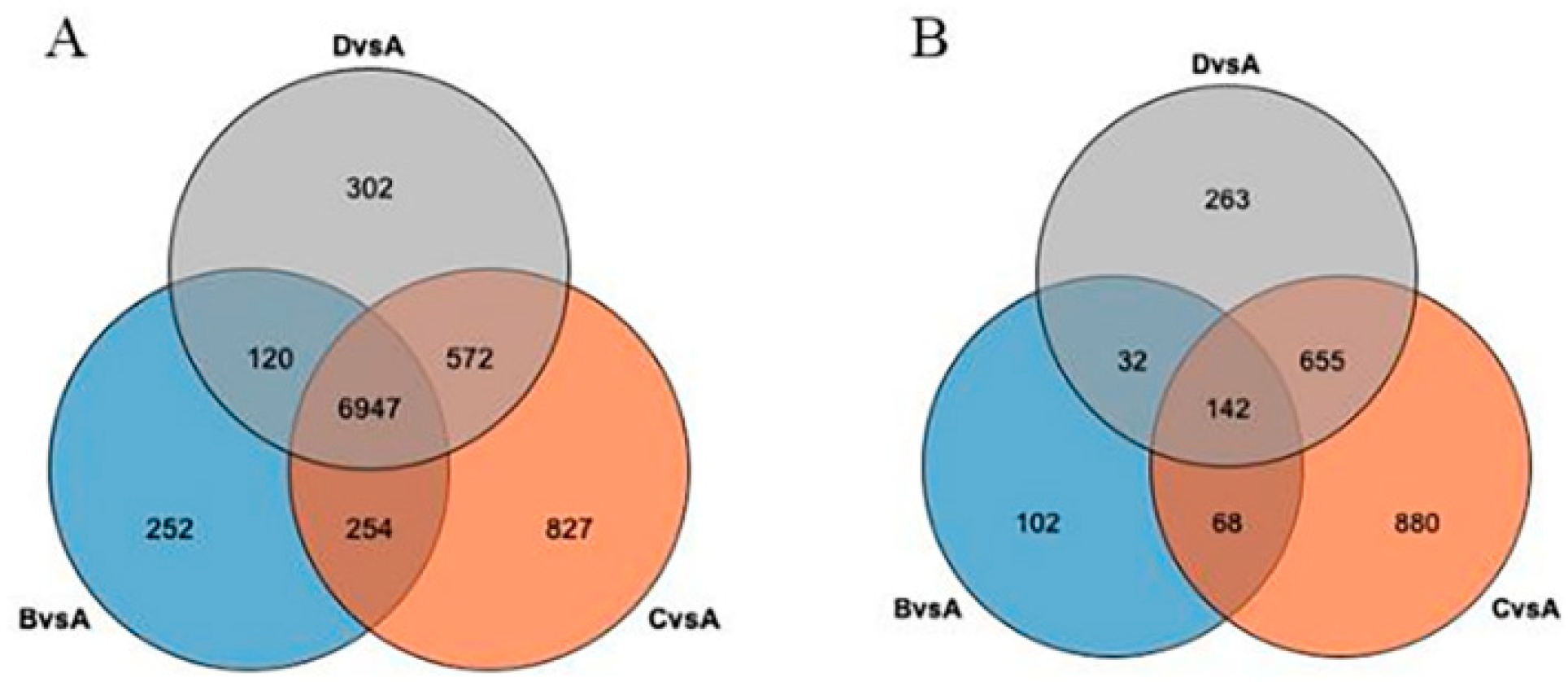
3.4. Analysis of the Number of Alternative Splicing Genes Common to and Specific to the CK and Each Treatment Comparison Group
3.5. GO Annotation and KEGG Functional Enrichment Analysis of the Alternative Splice Genes of the CK and Each Treatment Comparison Group of Acid Jujube
3.5.1. GO Annotation and KEGG Functional Enrichment Analysis of Differentially Alternative Splice Genes Specific to the CK and ALA Comparison Groups
3.5.2. GO Annotation and KEGG Functional Enrichment Analyses of Differentially Alternative Spliced Genes Specific to the CK Versus NaCl Comparison Group
3.5.3. Analysis of GO Annotation and KEGG Functional Enrichment of Differentially Spliced Genes Specific to the CK vs. NaCl + ALA Comparative Group
3.6. Annotation Information of the Genes Related to the Chlorophyll Metabolism Pathways
3.7. Expression of Chlorophyll-Related Metabolites and Genes of Enzymes in the Leaves of Jujube Seedlings under NaCl Stress for 72 h by Exogenous ALA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, M.J. Advances in taxonomy of jujube (Literature Review). Acta Hortic. Sin. 1999, 5, 302–308. [Google Scholar]
- Hua, Y.; Xu, X.X.; Guo, S.; Xie, H.; Yan, H.; Ma, X.F.; Niu, Y.; Duan, J.A. Wild jujube (Ziziphus jujuba var. spinosa): A review of its phytonutrients, health benefits, metabolism, and applications. J. Agric. Food Chem. 2022, 70, 7871–7886. [Google Scholar] [CrossRef]
- Porra, R.J. Recent progress in porphyrin and chlorophyll biosynthesis. J. Photochem. Photobiol. 1997, 65, 492–516. [Google Scholar] [CrossRef]
- Castelfranco, P.A.; Beale, S.I. Chlorophyll biosynthesis: Recent advances and areas of current interest. Annu. Rev. Plant Physiol. 1983, 34, 241–276. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Li, J.; Dawuda, M.M.; Ali, B.; Wu, Y.; Yu, J.; Tang, Z.; Lyu, J.; Xiao, X.; et al. Exogenous application of 5-aminolevulinic acid promotes coloration and improves the quality of tomato fruit by regulating carotenoid metabolism. Front. Plant Sci. 2021, 12, 683868. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Zhao, B.L. Effects of exogenous ALA on leaf photosynthetic characteristics and fruit quality of Crimson seedless grape. Acta Bot. Sin. 2018, 38, 493–500. [Google Scholar]
- Zhao, B.L.; Liu, P.; Zhang, X. Effects of 5-aminolevulinic acid on photosynthetic and growth characteristics of grape leaves under salt stress. Northern Hortic. 2016, 14, 1–6. [Google Scholar]
- An, Y.Y.; Zhang, L.Y.; Feng, F.F.; Tian, F.; Li, J.; Wang, L.J. Effect of 5-aminolevulinic acid on weak light tolerance of apple leaves. Acta Bot. Sin. 2016, 36, 987–995. [Google Scholar]
- Gao, J.J.; Feng, F.F.; Duan, C.H.; Li, J.H.; Shi, Z.X.; Gao, F.Y.; Wang, L.J. Effects of ALA on the photosynthetic performance of apple leaves and fruit quality. J. Fruit Sci. 2013, 30, 944–951. [Google Scholar]
- Werneke, J.M.; Chatfield, J.M.; Ogren, W.L. Alternative mRNA splicing generates the two ribulose bisphosphate carboxylase/oxygenase activase polypeptides in spinach and Arabidopsis. Plant Cell 1989, 1, 815–825. [Google Scholar]
- Reddy, A.S.; Marquez, Y.; Kalyna, M.; Barta, A. Complexity of the alternative splicing landscape in plants. Plant Cell 2013, 25, 3657–3683. [Google Scholar] [PubMed]
- Huang, L.; Li, Q.Y.; Wei, S.G.; Lai, J.; Dai, S.D.; Zhang, Q.F.; Zeng, H.L.; Liu, J.; Ye, P.S. Identification and difference analysis of the alternative splicing event in the hermaphroditic flowers and male flowers of Asparagus officinalis. Acta Hortic. Sin. 2019, 46, 1503–1518. [Google Scholar]
- Laloum, T.; Martín, G.; Duque, P. Alternative splicing control of abiotic stress responses. Trends Plant Sci. 2018, 23, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.Y.; William, D.; Sandhu, D. Isolation and characterization of Salt Overly Sensitive family genes in spinach. Physiol. Plant. 2021, 171, 520–532. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Zhang, Y.; Yu, J.J.; Zhang, H.; Wang, L.Y.; Wang, S.N.; Guo, S.Y.; Miao, Y.C.; Chen, S.X.; Li, Y.; et al. NaCl-responsive ROS scavenging and energy supply in alkaligrass callus revealed from proteomic analysis. BMC Genom. 2019, 20, 990. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Y.; Xie, Q.J.; Tang, F.F.; Wu, J.; Dong, W.; Wang, C.F.; Gao, C.Q. The ThSOS3 Gene improves the salt tolerance of transgenic Tamarix hispida and Arabidopsis thaliana. Front. Plant Sci. 2021, 11, 597480. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Zhang, X.Y.; Su, H.; Liu, X.F.; Wang, Y.F.; Hong, G.J. Genome-wide analysis of PYL-PP2C-SnRK2s family in Camellia sinensis. Bioengineered 2020, 11, 103–115. [Google Scholar] [CrossRef]
- Ke, Y.Y.; Liu, Y.; Zhao, Y.C.; Han, Z.; Zhang, J.; Yu, C.M. Advances in physiological and molecular mechanisms of salt stress in garden plants. Mol. PBA 2022, 1–10. Available online: https://kns.cnki.net/kcms/detail/46.1068.S.20220106.1038.002.html (accessed on 27 October 2022).
- Kong, J.; Gong, J.M.; Zhang, Z.G.; Zhang, J.S.; Chen, S.Y. A new AOX homologous gene OsIM1 from rice (Oryza sativa) with an alternative splicing mechanism under salt stress. Theor. Appl. Genet. 2003, 107, 326–331. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, S.; Zhang, Y.; Wang, X.; Li, D.; Li, Q.; Yue, M.; Li, Q.; Zhang, Y.E.; Xu, Y.; et al. Arabidopsis floral initiator SKB1 confers high salt tolerance by regulating transcription and pre-mRNA splicing through altering histone H4R3 and small nuclear ribonucleoprotein LSM4 methylation. Plant Cell 2011, 23, 396–411. [Google Scholar] [CrossRef]
- Palusa, S.G.; Ali, G.S.; Reddy, A.S. Alternative splicing of pre-mRNAs of Arabidopsis serine/arginine-rich proteins: Regulation by hormones and stresses. Plant J. 2007, 49, 1091–1107. [Google Scholar] [CrossRef]
- Feng, Y.L.; Xiong, Y.; Zhang, J.; Yuan, J.L.; Cai, A.S.; Ma, C. Role of alternative splicing in plant development and abiotic stress responses. J. Nucl. Agric. Sci. 2020, 34, 62–70. [Google Scholar]
- Li, F.F. Alleviation effect of exogenous 5-aminolevulinic acid on NaCl stress in wild jujube seeds and seedlings. Master’s Thesis, Shihezi University, Shihezi, China, 2017. [Google Scholar]
- Chang, X.Y.; Sun, J.L.; Zhao, B.L. Effects of exogenous ALA on photosynthetic characteristics and membrane lipid peroxidation of Ziziphus jujube seedlings under NaCl stress. Xinjiang Agric. Sci. 2019, 56, 1635–1644. [Google Scholar]
- Zhang, X.Z. Determination of chlorophyll content in plants—Acetone ethanol mixture method. Liaoning Agric. Sci. 1986, 3, 26–28. [Google Scholar]
- Wu, Y.; Jin, X.; Liao, W.; Hu, L.; Dawuda, M.M.; Zhao, X.; Tang, Z.; Gong, T.; Yu, J. 5-Aminolevulinic acid (ALA) alleviated salinity stress in cucumber seedlings by enhancing chlorophyll synthesis pathway. Front. Plant Sci. 2018, 9, 635. [Google Scholar] [CrossRef] [PubMed]
- Hodgins, R.R.; Van Huystee, R.B. Rapid simultaneous estimation of protoporphyrin and Mg-porphyrins in higher plants. J. Plant Physiol. 1986, 125, 311–323. [Google Scholar] [CrossRef]
- Bogorad, L. Porphyrin synthesis. Methods in Enzymology. 1962, 5, 885–895. [Google Scholar]
- Ma, Y.P.; Bai, L.Y.; Xu, W.R.; Cao, B.; Zhao, S.M.; Song, L.H.; Liu, J.J. Differential analysis of fruit transcriptome in grafted and root-tiller propagated plants of Lingwu long jujube. Mol. Plant Breed. 2021, 19, 5297–5306. [Google Scholar] [CrossRef]
- Ding, D.; Wang, H.B.; Zheng, L.J. Physiological response and comprehensive evaluation of different tea Chrysanthemun varieties under NaCl stress. Plant Physiol. J. 2021, 57, 692–702. [Google Scholar]
- Abrar, M.M.; Saqib, M.; Abbas, G.; Atiq-ur-Rahman, M.; Mustafa, A.; Shah, S.A.A.; Mehmood, K.; Maitlo, A.A.; ul-Hassan, M.; Sun, N.; et al. Evaluating the contribution of growth, physiological, and ionic components towards salinity and drought stress tolerance in Jatropha curcas. Plants 2020, 9, 1574. [Google Scholar] [CrossRef]
- Abrar, M.M.; Sohail, M.; Saqib, M.; Akhtar, J.; Abbas, G.; Wahab, H.A.; Mumtaz, M.Z.; Mehmood, K.; Memon, M.S.; Sun, N.; et al. Interactive salinity and water stress severely reduced the growth, stress tolerance, and physiological responses of guava (Psidium guajava L.). Sci. Rep. 2022, 2, 18952. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.X.; Guo, Y.F.; Zhang, M. Effects of exogenous 5-aminolevulinic acid on seed germination and seedling growth of maize under Salt Stress. J. Agric. 2021, 11, 7–12. [Google Scholar]
- Xiang, L.X.; Hu, L.P.; Meng, S. Effect of spraying spermidine on chlorophyll biosynthesis and metabolism of tomato under high temperature stress. Acta Bot. Sin. 2020, 40, 846–851. [Google Scholar]
- Li, L.J. Alleviating Effect and Regulation Mechanism of Exogenous Spermidine (SPD) on Drought Stress in Maize. Master’s Thesis, Northeast Agricultural University, Harbin, China, 2019. [Google Scholar]
- Jin, X.Q. Exogenousγ- Aminobutyric Acid Regulates the Metabolism of Reactive Oxygen Species and Chlorophyll to Enhance the Tolerance of Muskmelon Seedlings to Saline Alkali Stress. Master’s Thesis, Northwest University of Agriculture and Forestry Science and Technology, Xianyang, China, 2019. [Google Scholar]
- Lee, H.J.; Eom, S.H.; Lee, J.H. Genome-wide analysis of alternative splicing events during response to drought stress in tomato (Solanum lycopersicum L.). J. Hortic. Sci. Biotechnol. 2020, 95, 286–293. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, M.Y.; Tang, M.Q. Analysis on the difference of alternative shear in transcriptome of two cotton varieties under high temperature stress. Jiangsu Agric. Sci. 2022, 1–11. Available online: https://kns.cnki.net/kcms/detail/32.1214.S.20220706.1830.002.html (accessed on 27 October 2022).
- Wen, J.J.; Yu, K.H.; Liu, Z.S.; Feng, M.; Peng, H.R.; Ni, Z.F.; Yao, Y.Y.; Hu, Z.R.; Xin, M.M.; Sun, Q.X. Analysis on the mechanism of alternative splicing gene tahsfa6e involved in heat tolerance regulation in wheat. In Proceedings of the 19th Annual Academic Meeting of China Crop Society, Wuhan, China, 8 November 2020; p. 68. [Google Scholar]
- Li, S.; Yamada, M.; Han, X.; Ohler, U.; Benfey, P.N. High-resolution expression map of the Arabidopsis root reveals alternative splicing and lincRNA regulation. Dev. Cell 2016, 39, 508–522. [Google Scholar] [CrossRef]
- Huang, L.; Lai, J.; Wei, S.G. Analysis of antioxidant enzyme activity and alternative splicing of related genes in amphoteric and male flowers of Asparagus officinalis. Plant Physiol. 2019, 55, 1231–1238. [Google Scholar]
- Wai, W.; Jia, Y.L.; Wu, S. Full length transcriptome analysis of jujube fruit under water stress. Acta Agric. Boreali-Sin. 2020, 35, 63–71. [Google Scholar]
- Liu, L. Regulation Mechanism of Plant Hormones on Rice Seed Germination and Seedling Root Growth under Salt Stress. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2018. [Google Scholar]
- Pitann, B.; Zörb, C.; Mühling, K.H. Comparative proteome analysis of maize (Zea mays L.) expansins under salinity. J. Plant Nutr. Soil Sci. 2009, 172, 75–77. [Google Scholar] [CrossRef]
- Skorupa, M.; Gołębiewski, M.; Domagalski, K.; Kurnik, K.; Abu Nahia, K.; Złoch, M.; Tretyn, A.; Tyburski, J. Transcriptomic profiling of the salt stress response in excised leaves of the halophyte Beta vulgaris ssp. maritima. Plant Sci. 2016, 243, 56–70. [Google Scholar] [CrossRef]
- Wang, L.J.; Jiang, W.B.; Liu, H. Promotion by 5-aminolevulinic acid of germination of pakchoi (Brassica campestris ssp. chinensis var. communis Tsen et Lee) seeds under salt stress. J. Integr. Plant Biol. 2005, 47, 8. [Google Scholar]
- Pardo, J.M. Biotechnology of water and salinity stress tolerance. Curr. Opin. Biotech. 2010, 21, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Yu, X.D.; Cai, Z.P.; Wu, F.H.; Luo, J.J.; Zheng, L.T.; Chu, W.Q. An overview of chlorophyll biosynthesis in higher plants. Mol. PBA 2019, 17, 6013–6019. [Google Scholar]
- Li, Y.X.; Zhu, G.C. Research progress on the mechanism of plant leaf color variation. Agric. Technol. 2022, 42, 1–3. [Google Scholar]
- Zhang, L.; Xu, Z.R.; Hu, X.H.; Hu, L.P.; Zhou, Z.R.; Pan, X.H. Effects of foliar spermidine spraying on the growth and chlorophyll synthesis precursor content of tomato seedlings under saline alkali stress. Acta Bot. Sin. 2015, 35, 125–130. [Google Scholar]
- Huang, J.; Su, J.; Zhou, P.; Zhang, Q.; Zhang, M. Regulation characteristics of chlorophyll metabolism in leaves of Lycium chinense var. aureus during regreening period. J. Northeast. For. Univ. 2021, 49, 51–54. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer | Reverse Primer | |
|---|---|---|
| HEMH | TAATTCCGCTTCGCCGCTCATC | AGGCTGAACATCGTCCAGAGTCTC |
| UROIII | CTGTGCCTTCTTGTCCGCTACTTC | CTTGCCATTCTTCCCACGCTCTC |
| ChlD | GCAGAGAAGAGTGGTGATGTTGGTC | GCCTTGGTGTATCAGCAGTAGTAGC |
| NYC1 | GCTGTTTTGGGAGGTGTTGGTTTG | GCCAGTACAACTCCAGTGCTCATC |
| 18SrRNA | CAAAGCAAGCCTACGCTCTGT | CTATGAAATACGAATGCCCCC |
| Sample | Clean Base (G) | Error Rate (%) | Percentage of Bases with Phred Values Greater than 20 and 30 to the Total Number of Bases Q20 (%) Q30 (%) | Number of G/C Bases as a Percentage of the Total Number of Bases GC Content (%) | |
|---|---|---|---|---|---|
| A-1 | 11.46 | 0.03 | 97.46 | 93.14 | 41.75 |
| A-2 | 13.65 | 0.03 | 97.85 | 94.01 | 44.05 |
| A-3 | 11.99 | 0.03 | 97.66 | 93.54 | 41.58 |
| B-1 | 11.81 | 0.03 | 97.58 | 93.34 | 43.28 |
| B-2 | 12.93 | 0.03 | 97.81 | 93.98 | 43.75 |
| B-3 | 12.24 | 0.03 | 97.84 | 94.08 | 42.42 |
| C-1 | 11.69 | 0.03 | 97.55 | 93.46 | 41.78 |
| C-2 | 12.92 | 0.03 | 97.72 | 93.78 | 42.79 |
| C-3 | 11.34 | 0.03 | 97.75 | 93.85 | 42.83 |
| D-1 | 11.98 | 0.03 | 98.04 | 94.52 | 42.31 |
| D-2 | 13.07 | 0.03 | 97.91 | 94.16 | 43.02 |
| D-3 | 11.84 | 0.03 | 97.43 | 93.25 | 41.41 |
| Sample | Total Reads | Total Mapped | Mapping Rate (%) | Multiple Mapped | % of Reads Mapped to Multiple Loci | Uniquely Mapped | % of Reads Mapped to Unique Loci |
|---|---|---|---|---|---|---|---|
| A-1 | 38,214,548 | 34,835,669 | 91.16 | 5,052,402 | 13.22 | 29,783,267 | 77.94 |
| A-2 | 45,512,838 | 41,786,926 | 91.81 | 5,851,882 | 12.86 | 35,935,044 | 78.96 |
| A-3 | 39,967,417 | 36,688,376 | 91.80 | 5,145,188 | 12.87 | 31,543,188 | 78.92 |
| B-1 | 39,356,003 | 36,031,996 | 91.55 | 5,277,867 | 13.41 | 30,754,129 | 78.14 |
| B-2 | 43,088,377 | 39,568,358 | 91.83 | 5,752,816 | 13.35 | 33,815,542 | 78.48 |
| B-3 | 40,790,163 | 36,912,375 | 90.49 | 5,412,930 | 13.27 | 31,499,445 | 77.22 |
| C-1 | 38,979,318 | 35,374,366 | 90.75 | 5,239,216 | 13.44 | 30,135,150 | 77.31 |
| C-2 | 43,072,244 | 38,672,215 | 89.78 | 5,750,831 | 13.35 | 32,921,384 | 76.43 |
| C-3 | 37,799,291 | 34,009,911 | 89.97 | 5,153,749 | 13.63 | 28,856,162 | 76.34 |
| D-1 | 39,941,654 | 36,855,317 | 92.27 | 5,272,635 | 13.20 | 31,582,682 | 79.07 |
| D-2 | 39,454,369 | 35,729,624 | 90.56 | 5,248,463 | 13.30 | 30,481,161 | 77.26 |
| D-3 | 43,555,391 | 39,851,171 | 91.50 | 5,856,798 | 13.45 | 33,994,373 | 78.05 |
| GO Number | GO Description | Gene Number | p- Value | GO Category | Pathway Name | Gene Number | p- Value |
|---|---|---|---|---|---|---|---|
| GO: 0004674 | Protein serine/threonine kinase activity | 16 | 0.0019 | Molecular Function | Lipoic acid metabolism | 1 | 0.0125 |
| GO: 0005886 | Plasma membrane | 35 | 0.0265 | Cellular components | DNA replication | 2 | 0.0203 |
| GO: 0007178 | Transmembrane receptor protein serine/threonine kinase signaling pathway | 7 | 0.0045 | Biological processes | Fatty acid biosynthesis | 2 | 0.0272 |
| GO Number | GO Description | Gene Number | p- Value | GO Category | Pathway Name | Gene Number | p- Value |
|---|---|---|---|---|---|---|---|
| GO: 0016779 | Nucleotidyl transferase activity | 12 | 0.0015 | Molecular Function | Spliceosome | 20 | 0.0019 |
| GO: 0016772 | Transferase activity of transferring phosphorus-containing groups | 63 | 0.0048 | Molecular Function | RNA transfer | 20 | 0.002 |
| GO: 0044444 | Cytoplasmic fraction | 283 | 0.0000003 | Cellular components | Genetic Information Processing | 100 | 0.0022 |
| GO: 0005789 | Endoplasmic reticulum membrane | 29 | 0.0000068 | Cellular components | Translation | 48 | 0.0034 |
| GO: 1901564 | Organic nitrogen complex metabolic process | 186 | 0.0000002 | Biological processes | Transcription | 24 | 0.0138 |
| GO Number | GO Description | Gene Number | p- Value | GO Category | Pathway Name | Gene Number | p- Value |
|---|---|---|---|---|---|---|---|
| GO: 0016740 | Transferase activity | 37 | 0.0197 | Molecular Function | (GPI)-anchored biosynthesis | 4 | 0.0005 |
| GO: 0044436 | Vesicle fraction | 9 | 0.0015 | Cellular components | Polysaccharide synthesis and metabolism | 5 | 0.0256 |
| GO: 0009535 | Chloroplast vesicle-like membranes | 7 | 0.0067 | Cellular components | Porphyrin and chlorophyll metabolic processes | 3 | 0.0261 |
| GO: 0006302 | Double-strand break repair | 9 | 0.0015 | Biological processes | Phosphatidylinositol signaling system | 4 | 0.0283 |
| Gene ID | Gene Annotation | Abbreviations | AS Event Types |
|---|---|---|---|
| LOC107412374 | ferrochelatase-2 | HEMH | MXE, SE |
| LOC107410856 | magnesium-chelatase subunit ChlD | ChlD | SE |
| LOC107420915 | uroporphyrinogen-III synthase | UROIII | MXE, SE, A3SS |
| LOC107431821 | probable chlorophyll(ide) b reductase NYC1 | NYC1 | SE |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, C.; Liu, Z.; Chang, X.; Zhang, Z.; Shi, W.; Zhang, Z.; Zhao, B.; Sun, J. Analysis of the Alternative Splicing Events of Exogenous δ-Aminolevulinic Acid under NaCl Stress in Wild Jujube Seedlings. Forests 2022, 13, 2076. https://doi.org/10.3390/f13122076
Zhu C, Liu Z, Chang X, Zhang Z, Shi W, Zhang Z, Zhao B, Sun J. Analysis of the Alternative Splicing Events of Exogenous δ-Aminolevulinic Acid under NaCl Stress in Wild Jujube Seedlings. Forests. 2022; 13(12):2076. https://doi.org/10.3390/f13122076
Chicago/Turabian StyleZhu, Chunmei, Zhiyu Liu, Xinyi Chang, Zhijun Zhang, Wenchao Shi, Zhongrong Zhang, Baolong Zhao, and Junli Sun. 2022. "Analysis of the Alternative Splicing Events of Exogenous δ-Aminolevulinic Acid under NaCl Stress in Wild Jujube Seedlings" Forests 13, no. 12: 2076. https://doi.org/10.3390/f13122076
APA StyleZhu, C., Liu, Z., Chang, X., Zhang, Z., Shi, W., Zhang, Z., Zhao, B., & Sun, J. (2022). Analysis of the Alternative Splicing Events of Exogenous δ-Aminolevulinic Acid under NaCl Stress in Wild Jujube Seedlings. Forests, 13(12), 2076. https://doi.org/10.3390/f13122076