
Composition and Spatial Variation of Germinable Seed Bank in Burned Nothofagus pumilio Forests in Patagonia Argentina

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Seed Viability
2.3. Experimental Design and Sampling
2.4. Seed Bank
2.5. Data Analysis
2.5.1. Nothofagus pumilio Germination Capacity
2.5.2. Soil Seed Bank Response
2.5.3. Comparison between Above-Ground Vegetation and Soil Seed Bank Composition
3. Results
3.1. Nothofagus pumilio Germination Capacity
3.2. Emergence from the Soil Seed Bank
3.2.1. Emergence of Tree Species
3.2.2. Emergence of Dwarf Shrub-Shrub Species
3.2.3. Emergence of Herbaceous and Grass Species
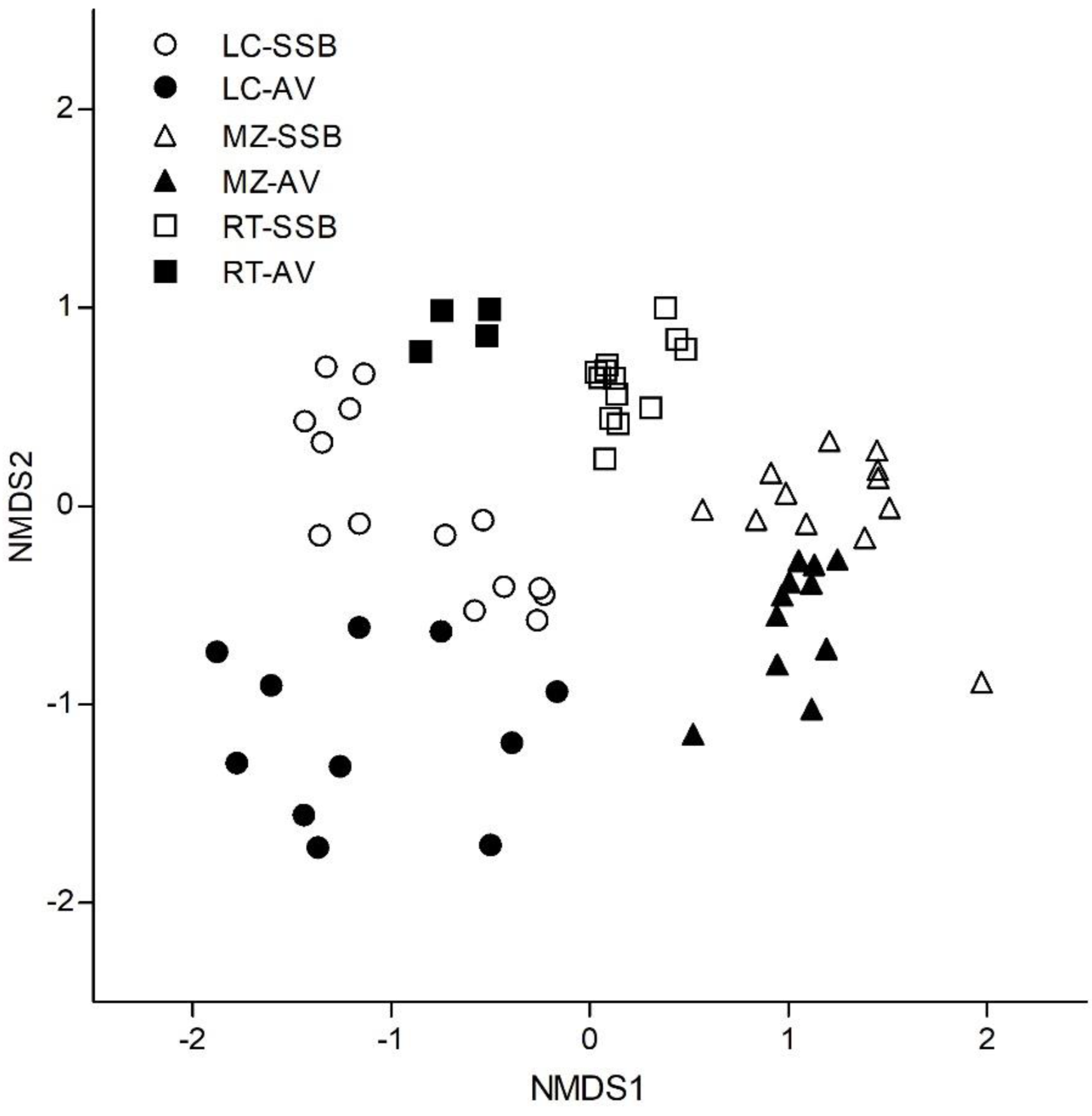
3.3. Above-Ground Vegetation and Soil Seed Bank Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pickett, S.; McDonnell, M. Seed bank dynamics in temperate deciduous forest. In Ecology of Soil Seed Banks; Leck, M., Parker, V., Simpson, R., Eds.; Academic Press Inc.: San Diego, CA, USA, 1989; pp. 123–147. [Google Scholar]
- Donoso Zegers, C. Bosques Templados de Chile y Argentina. Variación, Estructura y Dinámica, 1st ed.; Editorial Universitaria SA: Valdivia, Chile, 1993; p. 484. [Google Scholar]
- Veblen, T.T.; Donoso Zegers, C.; Kitzberger, T.; Rebertus, A.J. Ecology of southern Chilean and southern Argentinean Nothofagus forests. In The Ecology and Biogeography of Nothofagus Forests; Veblen, T., Hill, R.S., Read, J., Eds.; Yale University Press: New Haven, CT, USA; London, UK, 1996; pp. 293–353. [Google Scholar]
- Little, S. Effects of fire on temperate forests: Northeastern United States. In Fire and Ecosystems; Kozlowski, T., Ahlgren, C., Eds.; Academic Press: New York, NY, USA, 1974; pp. 225–250. [Google Scholar]
- Keeley, J.E. Fire intensity, fire severity and burn severity: A brief review and suggested usage. Int. J. Wildland Fire 2009, 18, 116–126. [Google Scholar] [CrossRef]
- Ferrandis, P.; Herranz, J.; Martínez-Sánchez, J. The role of soil seed bank in the early stages of plant recovery after fire in a Pinus pinaster forest in SE Spain. Int. J. Wildland Fire 1996, 6, 31–35. [Google Scholar] [CrossRef]
- Falk, D.A.; van Mantgem, P.J.; Keeley, J.E.; Gregg, R.M.; Guiterman, C.H.; Tepley, A.J.; Young, D.J.; Marshall, L.A. Mechanisms of forest resilience. Forest Ecol. Manag. 2022, 512, 120–129. [Google Scholar] [CrossRef]
- Veblen, T.T.; Kitzberger, T.; Raffaele, E.; Mermoz, M.; González, M.E.; Sibold, J.S.; Holz, A. The historical range of variability of fires in the Andean–Patagonian Nothofagus forest region. Int. J. Wildland Fire 2008, 17, 724–741. [Google Scholar] [CrossRef]
- CIEFAP; SAyDS. Monitoreo de la Superficie de Bosque Nativo de la República Argentina. Región Forestal Bosque Andino Patagónico; Centro de Investigación y Extensión Forestal Andino Patagónico-Secretaría de Ambiente y Desarrollo Sustentable: Buenos Aires, Argentina, 2019; p. 86. Available online: https://www.argentina.gob.ar/sites/default/files/monitoreo_de_la_superficie_de_bosque_nativo_bap_2_de_octubre_2019.pdf (accessed on 10 June 2022).
- Donoso Zegers, C. Variación natural en especies de Nothofagus en Chile. Bosque 1987, 8, 85–97. [Google Scholar] [CrossRef]
- Bava, J.O.; Lencinas, J.D.; Haag, A. Determinación De La Materia Prima Disponible Para Proyectos de Inversión Forestales En Cuencas De La Provincia Del Chubut; Consejo Federal de Inversiones, Provincia del Chubut: Esquel, Argentina, 2006; p. 151. [Google Scholar]
- Mohr Bell, D. Superficies Afectadas por Incendios en la Región Bosque Andino Patagónico Durante los Veranos de 2013–2014 y 2014–2015—Report (Unpublished work) Nodo Regional Bosque Andino Patagónico (SAyDS–CIEFAP). Unpublished Work. 2015; 12. [Google Scholar]
- Dimitri, M. La Región de los Bosques Andino-Patagónicos; Ediciones INTA: Buenos Aires, Argentina, 1983; p. 381. [Google Scholar]
- Thompson, K.; Grime, J.P. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. J. Ecol. 1979, 67, 893–921. [Google Scholar] [CrossRef]
- Rusch, V. Estudio sobre la regeneración de la lenga (Nothofagus pumilio) en la cuenca del Río Manso Superior, Río Negro. In Informe Final Beca de Iniciación; CONICET: Buenos Aires, Argentina, 1987; p. 113. [Google Scholar]
- Cuevas, J.G.; Arroyo, M.T. Ausencia de banco de semillas persistente en Nothofagus pumilio (Fagaceae) en Tierra del Fuego, Chile. Rev. Chil. Hist. Nat. 1999, 72, 73–82. [Google Scholar]
- Urretavizcaya, M.F.; Contardi, L.; Oyharçabal, M.F.; Pasquini, M. Calidad de semillas de especies nativas del bosque andino patagónico de la provincia de Chubut y su importancia para la producción de plantines. Rev. Fac. Agron. Univ. Nac. La Plata 2016, 115, 9–18. [Google Scholar]
- González, M.; Donoso Zegers, C.; Ovalle, P.; Martínez-Pastur, G. Nothofagus pumilio (Poep. et Endl.) Krasser. Lenga, roble blanco, leñar, roble de Tierra del Fuego. In Las Especies Arbóreas de los Bosques Templados de Chile y Argentina; Donoso Zegers, C., Ed.; Marisa Cuneo Ediciones: Valdivia, Chile, 2006; pp. 486–500. [Google Scholar]
- Rodrigo, A.; Arnan, X.; Retana, J. Relevance of soil seed bank and seed rain to immediate seed supply after a large wildfire. Int. J. Wildland Fire 2012, 21, 449–458. [Google Scholar] [CrossRef]
- Rusch, V. Determinación de las transiciones de estado en bosques de lenga (Nothofagus pumilio). In Informe Final Beca de Avance; CONICET: Buenos Aires, Argentina, 1989; p. 76. [Google Scholar]
- Kitzberger, T.; Raffaele, E.; Heinemann, K.; Mazzarino, M.J. Effects of fire severity in a north Patagonian subalpine forest. J. Veg. Sci. 2005, 16, 5–12. [Google Scholar] [CrossRef]
- Varela, S.A.; Gobbi, M.E.; Laos, F. Banco de semillas de un bosque quemado de Nothofagus pumilio: Efecto de la aplicación de compost de biosólidos. Ecol. Austral 2006, 16, 63–78. [Google Scholar]
- Quinteros, C.P.; Bava, J.O.; López Bernal, P.M.; Gobbi, M.E.; Defossé, G.E. Competition effects of grazing-modified herbaceous vegetation on growth, survival and water relations of lenga (Nothofagus pumilio) seedlings in a temperate forest of Patagonia, Argentina. Agroforest Syst. 2017, 91, 597–611. [Google Scholar] [CrossRef]
- Urretavizcaya, M.F. Propiedades del suelo en bosques quemados de Austrocedrus chilensis en Patagonia, Argentina. Bosque 2010, 31, 140–149. [Google Scholar] [CrossRef]
- Urretavizcaya, M.F.; Peri, P.L.; Monelos, L.; Arriola, H.; Oyharçabal, M.F.; Contardi, L.T.; Muñoz, M.; Sepúlveda, E.; Defossé, G.E. Condiciones de suelo y vegetación en tres bosques quemados de Nothofagus pumilio en Argentina y experiencias para su restauración activa. Ecol. Austral 2018, 28, 383–399. [Google Scholar] [CrossRef]
- Mutch, L.S.; Swetnam, T.W. Effects of fire severity and climate on ring-width growth of giant sequoia after fire. In Proceedings of the Symposium on Fire in Wilderness and Park Management: Past Lessons and Future Opportunities; Brown, J.K., Mutch, R.W., Spoon, C.W., Wakimoto, R.H., Eds.; Gen. Tech. Rep. INT-GTR-320; US Department of Agriculture, Forest Service: Ogden, UT, USA, 1995. [Google Scholar]
- Karger, D.N.; Zimmermann, N.E. Climatologies at High Resolution for the Earth Land Surface Areas CHELSA V1. 2: Technical Specification; Swiss Federal Research Institute WSL: Birmensdorf, Switzerland, 2019.
- López Bernal, P.M.; Urretavizcaya, M.F.; Defossé, G.E. Seedling Dynamics in an Environmental Gradient of Andean Patagonia, Argentina. In From Seed Germination to Young Plants: Ecology, Growth and Environmental Influences; Busso, C., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2012; pp. 189–210. [Google Scholar]
- Correa, M. Flora Patagónica I-VIII; Instituto Nacional de Tecnología Agropecuaria: Buenos Aires, Argentina, 1969–1988. [Google Scholar]
- Zuloaga, F.; Morrone, O.; Belgrano, M. Flora deldono Sur. Catálogo de las Plantas Vasculares. 2013. Available online: http://www.darwin.edu.ar/Proyectos/FloraArgentina/fa.htm (accessed on 1 February 2022).
- van der Valk, A.; Pederson, R. Seed banks and the management and restoration of natural vegetation. In Ecology of soil seed banks; Leck, M., Parker, V., Simpson, R., Eds.; Academic Press: San Diego, CA, USA, 1989; pp. 329–346. [Google Scholar]
- ISTA. International Rules for Seed Testing; International Seed Testing Association: Zürich, Switzerland, 1999. [Google Scholar]
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK, 1977. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–145. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference. R package version 1.43.17. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 10 June 2022).
- Team, R.C. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.r-project.org/ (accessed on 10 June 2022).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D. Community Ecology Package. R package version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 10 June 2022).
- Cavallero, L. Heterogeneidad Ambiental y Dispersión de Semillas en Comunidades de Distinta Edad Post-Fuego del Noroeste de Patagonia; Universidad Nacional del Comahue: Bariloche, Argentina, 2012. [Google Scholar]
- Pickett, S.; Cadenasso, M.; Jones, C. Generation of heterogeneity by organisms: Creation, maintenance and transformation. In The Ecological Consequences of Environmental Heterogeneity; Hutchings, M., John, E., Stewart, J., Eds.; Blacwell Science: London, UK, 2000; pp. 33–52. [Google Scholar]
- Bakker, J.; Poschlod, P.; Strykstra, R.; Bekker, R.; Thompson, K. Seed banks and seed dispersal: Important topics in restoration ecology. Acta Bot. Neerl. 1996, 45, 461–490. [Google Scholar] [CrossRef]
- Cellini, J.M. Estructura y Regeneración Bajo Distintas Propuestas de Manejo de Bosques de Nothofagus Pumilio (Poepp. Et. Endl.) Krasser en Tierra del Fuego, Argentina; Universidad Nacional de La Plata: La Plata, Argentina, 2010. [Google Scholar]
- Rusch, V. Principales limitantes para la regeneración de la lenga en la zona NE de su área de distribución. In Proceedings of the Seminario de manejo forestal de la lenga y aspectos ecológicos relacionados, Esquel, Argentina, 23–24 June 1992; pp. 61–73. [Google Scholar]
- Bava, J.O. Aportes Ecológicos y Silviculturales a la Transformación de Bosques Virgenes de Lenga (Nothofagus Pumilio (Poepp. et Endl.) Krasser en Bosques Manejados en Sector Argentino de Tierra del Fuego; CIEFAP, Centro de Investigación y Extensión Forestal Andino Patagónico: Esquel, Argentina, 1999; p. 138. [Google Scholar]
- Martínez Pastur, G.; Peri, P.; Fernández, C.; Staffieri, G.; Rodríguez, D. Desarrollo de la regeneración a lo largo del ciclo del manejo forestal de un bosque de Nothofagus pumilio: 1. Incidencia de la cobertura y el aprovechamiento o cosecha. Bosque 1999, 20, 39–46. [Google Scholar] [CrossRef]
- Bertolin, M.L.; Urretavizcaya, M.; Defossé, G. Fire emissions and car-bon uptake in severely burned lenga beech (Nothofagus pumilio) forests of Patagonia, Argentina. Fire Ecol. 2015, 11, 32–54. [Google Scholar] [CrossRef]
- Walker, L.R.; del Moral, R. Lessons from primary succession for restoration of severely damaged habitats. Appl. Veg. Sci. 2009, 12, 55–67. [Google Scholar] [CrossRef]
- Cubiña, A.; Aide, T.M. The effect of distance from forest edge on seed rain and soil seed bank in a tropical pasture 1. Biotropica 2001, 33, 260–267. [Google Scholar] [CrossRef]
- Lee, P. The impact of burn intensity from wildfires on seed and vegetative banks, and emergent understory in aspen-dominated boreal forests. Can. J. Bot. 2004, 82, 1468–1480. [Google Scholar] [CrossRef]
- de Paz, M.; Raffaele, E. Cattle change plant reproductive phenology, promoting community changes in a post-fire Nothofagus forest in northern Patagonia, Argentina. J. Plant Ecol. 2013, 6, 459–467. [Google Scholar] [CrossRef][Green Version]
- White, P.S.; Pickett, S. Natural disturbance and patch dynamics: An introduction. In Natural Disturbance and Patch Dynamics; Pickett, S., White, P.S., Eds.; Academic Press: San Diego, CA, USA, 1985; pp. 3–13. [Google Scholar]
- SSB; CIEFAP; INTA. Programa Integral de Manejo y Restauración de las Grandes Áreas Afectadas por los Incendios Forestales de la Temporada 2014–2015 en la Provincia de Chubut Subsecretaría de Bosques de Chubut; Centro de Investigación y Extensión Forestal Andino Patagónico, Instituto Nacional de Tecnologia Agropecuearia: Esquel, Argentina, 2015; p. 144. [Google Scholar]
- Williams, A.; Orellana, I.; Bertoldi, G.; von Müller, A.; Roveta, R.; Urretavizcaya, M. Análisis del banco de semillas post-incendio en “Las Horquetas”, Cholila, Chubut. In Proceedings of the IV Jornadas Patagónicas de Biología, III Jornadas Patagónicas de Ciencias Ambientales y VI Jornadas Estudiantiles de Ciencias Biológicas, Trelew, Argentina, 19–21 September 2018. [Google Scholar]
- Grandin, U.; Rydin, H. Attributes of the seed bank after a century of primary succession on islands in Lake Hjälmaren, Sweden. J. Ecol. 1998, 86, 293–303. [Google Scholar] [CrossRef]
- Armstrong, G.W. A stochastic characterisation of the natural disturbance regime of the boreal mixedwood forest with implications for sustainable forest management. Can. J. For. Res. 1999, 29, 424–433. [Google Scholar] [CrossRef]
- Hopfensperger, K.N. A review of similarity between seed bank and standing vegetation across ecosystems. Oikos 2007, 116, 1438–1448. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LC | MZ | RT | ||||||
|---|---|---|---|---|---|---|---|---|
| Genus and Species | SSB | AV | SSB | AV | SSB | AV | Habit | Status |
| Acaena sp. | X | X | X | X | X | H | N/E | |
| Acaena splendens | X | X | H | N/E | ||||
| Anemone multifida | X | X | H | N/E | ||||
| Austrocedrus chilensis | X | T | N/E | |||||
| Azorella prolifera | X | X | S | N/E | ||||
| Baccharis magellanica | X | X | S | N/E | ||||
| Baccharis sp. | X | X | X | S | N/E | |||
| Berberis microphylla | X | X | X | X | X | S | N/E | |
| Berberis serrato-dentata | X | X | S | N/E | ||||
| Cerastium sp. | X | X | H | A/Ex | ||||
| Cirsium vulgare | X | X | H | A/Ex | ||||
| Draba verna. | X | X | H | A/Ex | ||||
| Erigeron sp. | X | H | N/E | |||||
| Galium aparine | X | X | H | A/Ex | ||||
| Gaulteria mucronata | X | S | N/E | |||||
| Hieracium glaucifolium | X | X | H | N/E | ||||
| Leucheria sp. | X | X | X | H | N/E | |||
| Pinnasa bergii | X | H | N/E | |||||
| Maytenus chubutensis | X | S | N/E | |||||
| Muehlenbeckia hastulata | X | X | H | N/E | ||||
| Nothofagus pumilio | X | X | X | X | T | N/E | ||
| Osmorhiza berteroi | X | X | X | H | N/E | |||
| Phacelia secunda | X | X | H | N/E | ||||
| Poa sp. | X | G | N/E | |||||
| Prunella vulgaris | X | H | A/Ex | |||||
| Ribes cucullatum | X | X | X | S | N/E | |||
| Rumex acetosella | X | X | X | H | A/Ex | |||
| Schinus patagonicus | X | X | S | N/E | ||||
| Senecio neaei | X | H | N/E | |||||
| Senecio trifurcatus | X | X | H | N/E | ||||
| Sisyrinchium sp. | X | H | N/E | |||||
| Stellaria media | X | H | A/Ex | |||||
| Pappostipa sp. | X | X | G | A/Ex | ||||
| Taraxacum officinale | X | X | X | H | A/Ex | |||
| Vicia magellanica | X | X | H | N/E | ||||
| Viola maculata | X | X | X | X | H | N/E | ||
| Sp 01 | X | H | ||||||
| Sp 02 | X | H | ||||||
| Sp 03 | X | H | ||||||
| Sp 04 | X | X | H | |||||
| Sp 05 | X | H | ||||||
| Sp 06 | X | H | ||||||
| Sp 07 | X | H | ||||||
| Sp 08 | X | H | ||||||
| Sp 09 | X | H | ||||||
| Sp 10 | X | X | H | |||||
| Sp 11 | X | H | ||||||
| Sp 12 | X | H | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urretavizcaya, M.F.; Albarracín, V.; Orellana, I.; Rago, M.M.; López Bernal, P.; Monelos, L.; Peri, P.L. Composition and Spatial Variation of Germinable Seed Bank in Burned Nothofagus pumilio Forests in Patagonia Argentina. Forests 2022, 13, 1902. https://doi.org/10.3390/f13111902
Urretavizcaya MF, Albarracín V, Orellana I, Rago MM, López Bernal P, Monelos L, Peri PL. Composition and Spatial Variation of Germinable Seed Bank in Burned Nothofagus pumilio Forests in Patagonia Argentina. Forests. 2022; 13(11):1902. https://doi.org/10.3390/f13111902
Chicago/Turabian StyleUrretavizcaya, M. Florencia, Viviana Albarracín, Ivonne Orellana, M. Melisa Rago, Pablo López Bernal, Lucas Monelos, and Pablo Luis Peri. 2022. "Composition and Spatial Variation of Germinable Seed Bank in Burned Nothofagus pumilio Forests in Patagonia Argentina" Forests 13, no. 11: 1902. https://doi.org/10.3390/f13111902
APA StyleUrretavizcaya, M. F., Albarracín, V., Orellana, I., Rago, M. M., López Bernal, P., Monelos, L., & Peri, P. L. (2022). Composition and Spatial Variation of Germinable Seed Bank in Burned Nothofagus pumilio Forests in Patagonia Argentina. Forests, 13(11), 1902. https://doi.org/10.3390/f13111902