

Evidence of Pheromone Use in a Fulgorid, Spotted Lanternfly



{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Whole Body Extract
2.3. Behavioral Bioassays
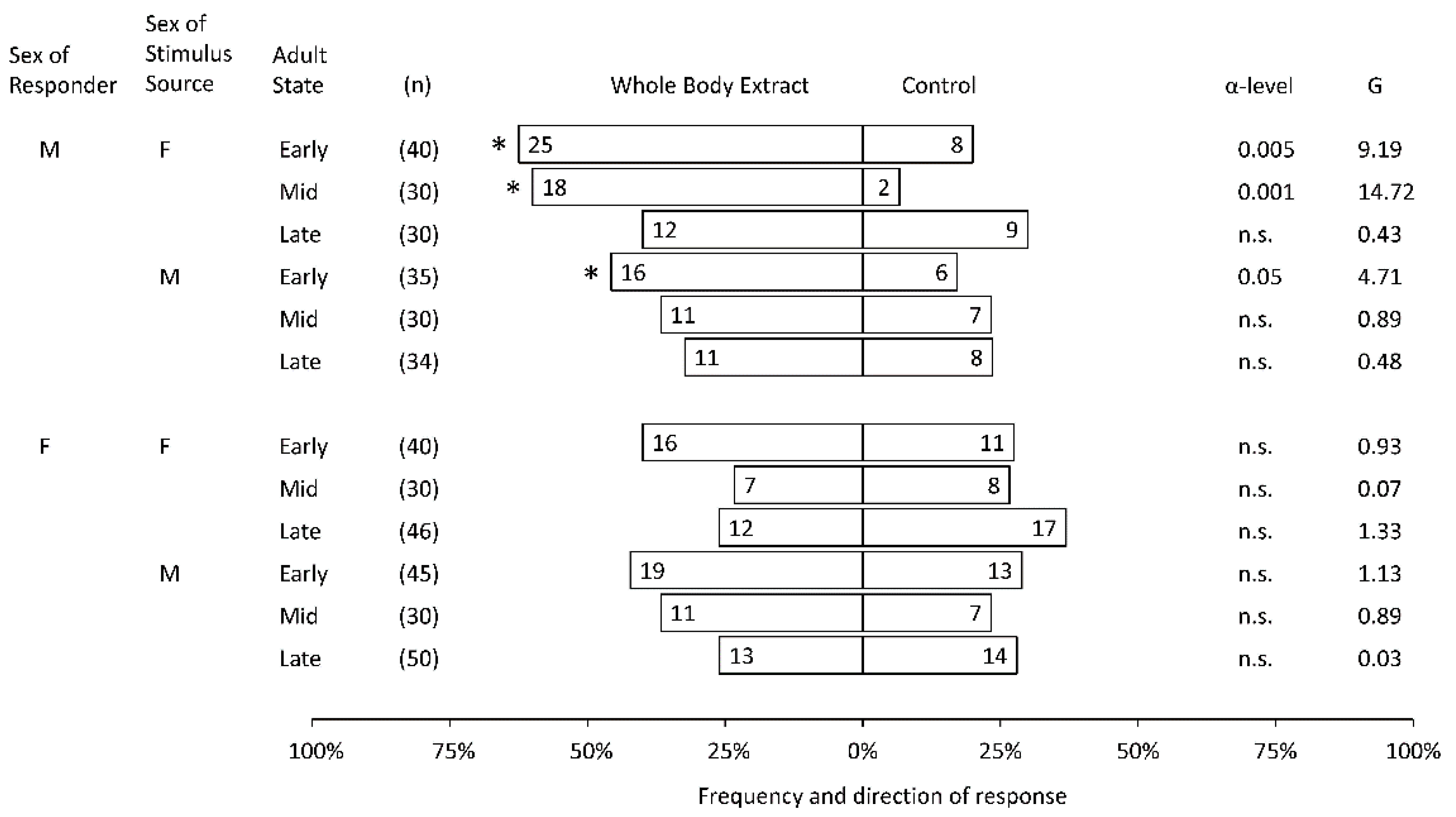
3. Results
Sex-Specific Responses to Whole Body Extracts
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, D.-H.; Park, Y.-L.; Leskey, T.C. A review of biology and management of Lycorma delicatula (Hemiptera: Fulgoridae), an emerging global invasive species. J. Asia-Pac. Entomol. 2019, 22, 589–596. [Google Scholar] [CrossRef]
- Shorey, H.H. Behavioral responses to insect pheromones. Annu. Rev. Entomol. 1973, 18, 349–380. [Google Scholar] [CrossRef]
- Ichikawa, T. Mutual communication by substrate vibrations in the mating behavior of planthoppers (Homoptera: Delphacidae). Appl. Entomol. Zool. 1976, 11, 8–21. [Google Scholar] [CrossRef][Green Version]
- Ichikawa, T.; Ishii, S. Mating signal of the brown planthopper, Nilaparvata lugens Stål (Homoptera: Delphacidae): Vibration of the substrate. Appl. Entomol. Zool. 1974, 9, 196–198. [Google Scholar] [CrossRef]
- Claridge, M.F.; Morgan, J.C.; Moulds, M.S. Substrate-transmitted acoustic signals of the primitive cicada, Tettigarcta crinita Distant (Hemiptera Cicadoidea, Tettigarctidae). J. Nat. Hist. 1999, 33, 1831–1834. [Google Scholar] [CrossRef]
- Tishechkin, D.Y. New data on vibrational calling signals of Fulgoroidea (Homoptera: Auchenorrhyncha) from the Asian part of Palaearctic with new records of three species of Cixiidae. Russ. Entomol. J. 2016, 25, 307–322. [Google Scholar] [CrossRef]
- Tishechkin, D.Y. On the similarity of temporal pattern of vibrational calling signals in different species of Fulgoroidea (Homoptera: Auchenorrhyncha). Russ. Entomol. J. 2008, 17, 349–357. [Google Scholar]
- Tishechkin, D.Y. Vibrational communication in Cercopoidea and Fulgoroidea (Homoptera: Cicadina) with notes on classification of higher taxa. Russ. Entomol. J. 2003, 12, 127–181. [Google Scholar]
- Virant-Doberlet, M.; Žežlina, I. Vibrational communication of Metcalfa pruinosa (Hemiptera: Fulgoroidea: Flatidae). Ann. Entomol. Soc. Am. 2007, 100, 73–82. [Google Scholar] [CrossRef]
- Davranoglou, L.-R.; Cicirello, A.; Taylor, G.K.; Mortimer, B. Planthopper bugs use a fast, cyclic elastic recoil mechanism for effective vibrational communication at small body size. PLoS Biol. 2019, 17, e3000155. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liang, A.-P. Identification of a self-regulatory pheromone system that controls nymph aggregation behavior of rice spittlebug Callitettix versicolor. Front. Zool. 2015, 12, 10. [Google Scholar] [CrossRef]
- Derstine, N.T.; Meier, L.; Canlas, I.; Murman, K.; Cannon, S.; Carrillo, D.; Wallace, M.; Cooperband, M.F. Plant volatiles help mediate host plant selection and attraction of the spotted lanternfly, Lycorma delicatula (Hemiptera: Fulgoridae): A generalist with a preferred host. Environ. Entomol. 2020, 49, 1049–1062. [Google Scholar] [CrossRef] [PubMed]
- Cooperband, M.F.; Wickham, J.; Cleary, K.; Spichiger, S.-E.; Zhang, L.; Baker, J.; Canlas, I.; Derstine, N.; Carrillo, D. Discovery of three kairomones in relation to trap and lure development for spotted lanternfly (Hemiptera: Fulgoridae). J. Econ. Entomol. 2019, 112, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Nixon, L.J.; Leach, H.; Barnes, C.; Urban, J.; Kirkpatrick, D.M.; Ludwick, D.C.; Short, B.; Pfeiffer, D.G.; Leskey, T.C. Development of Behaviorally Based Monitoring and Biosurveillance Tools for the Invasive Spotted Lanternfly (Hemiptera: Fulgoridae). Environ. Entomol. 2020, 49, 1117–1126. [Google Scholar] [CrossRef]
- Cooperband, M.; Meier, L.; Wickham, J.; Murman, K.; Cannon, S.; Abreu, L. Tree effects: Understanding what makes “hot” trees hot to spotted lanternfly. In Otis Laboratory 2018 Annual Report; Trepanowski, N., Vieira, K., Heller, S., Booth, E., Eds.; United States Department of Agriculture: Buzzards Bay, MA, USA, 2019; pp. 62–63. [Google Scholar]
- Faal, H.; Meier, L.R.; Canlas, I.J.; Murman, K.; Wallace, M.S.; Carrillo, D.; Cooperband, M.F. Volatiles from male honeydew excretions attract conspecific male spotted lanternflies, Lycorma delicatula (Hemiptera: Fulgoridae). Front. Insect Sci. 2022, 2, 982965. [Google Scholar] [CrossRef]
- Urban, J.M. Perspective: Shedding light on spotted lanternfly impacts in the USA. Pest Manag. Sci. 2020, 76, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Borges, M.; Jepson, P.C.; Howse, P.E. Long-range mate location and close-range courtship behaviour of the green stink bug, Nezara viridula and its mediation by sex pheromones. Entomol. Exp. Appl. 1987, 44, 205–212. [Google Scholar] [CrossRef]
- Čokl, A.; McBrien, H.L.; Millar, J.G. Comparison of substrate-borne vibrational signals of two stink bug species, Acrosternum hilare and Nezara viridula (Heteroptera: Pentatomidae). Ann. Entomol. Soc. Am. 2001, 94, 471–479. [Google Scholar] [CrossRef]
- Miklas, N.; Lasnier, T.; Renou, M. Male bugs modulate pheromone emission in response to vibratory signals from conspecifics. J. Chem. Ecol. 2003, 29, 561–574. [Google Scholar] [CrossRef] [PubMed]
- Zgonik, V.; Čokl, A. The role of signals of different modalities in initiating vibratory communication in Nezara viridula. Open Life Sci. 2014, 9, 200–211. [Google Scholar] [CrossRef]
- Čokl, A.; Laumam, R.A.; Stritih, N. Substrate-borne vibratory communication. In Stink Bugs; Čokl, A., Borges, M., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 125–164. [Google Scholar]
- Soroker, V.; Talebaev, S.; Harari, A.R.; Wesley, S.D. The role of chemical cues in host and mate location in the pear psylla Cacopsylla bidens (Homoptera: Psyllidae). J. Insect Behav. 2004, 17, 613–626. [Google Scholar] [CrossRef]
- Wenninger, E.J.; Stelinski, L.L.; Hall, D.G. Behavioral evidence for a female-produced sex attractant in Diaphorina citri. Entomol. Exp. Appl. 2008, 128, 450–459. [Google Scholar] [CrossRef]
- Guédot, C.; Horton, D.R.; Landolt, P.J. Sex attraction in Bactericera cockerelli (Hemiptera: Triozidae). Environ. Entomol. 2010, 39, 1302–1308. [Google Scholar] [CrossRef] [PubMed]
- Guédot, C.; Millar, J.G.; Horton, D.R.; Landolt, P.J. Identification of a sex attractant pheromone for male winterform pear psylla, Cacopsylla pyricola. J. Chem. Ecol. 2009, 35, 1437–1447. [Google Scholar] [CrossRef]
- Moghbeli Gharaei, A.; Ziaaddini, M.; Jalali, M.; Michaud, J. Sex-specific responses of Asian citrus psyllid to volatiles of conspecific and host-plant origin. J. Appl. Entomol. 2014, 138, 500–509. [Google Scholar] [CrossRef]
- Stockton, D.G.; Martini, X.; Stelinski, L.L. Male psyllids differentially learn in the context of copulation. Insects 2017, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Čokl, A.; Žunič-Kosi, A.; Laumann, R.A. Stink bug communication with multimodal signals transmitted through air and substrate. Emerg. Sci. J. 2019, 3, 407–424. [Google Scholar] [CrossRef]
- Blassioli-Moraes, M.C.; Magalhaes, D.M.; ČOkl, A.; Laumann, R.A.; Da Silva, J.P.; Silva, C.C.A.; Borges, M. Vibrational communication and mating behaviour of Dichelops melacanthus (H emiptera: P entatomidae) recorded from loudspeaker membranes and plants. Physiol. Entomol. 2014, 39, 1–11. [Google Scholar] [CrossRef]
- Borges, M.; Moraes, M.C.B.; Peixoto, M.F.; Pires, C.S.S.; Sujii, E.R.; Laumann, R.A. Monitoring the Neotropical brown stink bug Euschistus heros (F.)(Hemiptera: Pentatomidae) with pheromone-baited traps in soybean fields. J. Appl. Entomol. 2011, 135, 68–80. [Google Scholar] [CrossRef]
- Polajnar, J.; Čokl, A. The effect of vibratory disturbance on sexual behaviour of the southern green stink bug Nezara viridula (Heteroptera, Pentatomidae). Open Life Sci. 2008, 3, 189–197. [Google Scholar] [CrossRef]
- Claridge, M.F. Vibratory Signals Produced by Heteroptera—Pentatomorpha and Cimicomorpha: Matija Gogala. In Insect Sounds and Communication; CRC Press: Boca Raton, FL, USA, 2005; pp. 293–314. [Google Scholar]
- Rohde, B.B.; Cooperband, M.F.; Canlas, I.; Mankin, R.W. Evidence of receptivity to vibroacoustic stimuli in the spotted lanternfly. J. Econ. Entomol. 2022. [Google Scholar]
- Cooperband, M.F.; Murman, K. Responses of adult spotted lanternflies to artificial aggregations composed of all males or females. Front. Insect Sci. 2022, 2, 981832. [Google Scholar] [CrossRef]
- Stewart-Jones, A.; Poppy, G.M. Comparison of glass vessels and plastic bags for enclosing living plant parts for headspace analysis. J. Chem. Ecol. 2006, 32, 845–864. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Sokal, R.R. Statistical Tables; Macmillan: New York, NY, USA, 1995. [Google Scholar]
- Horton, D.R.; Guédot, C.; Landolt, P.J. Attraction of male summerform pear psylla to volatiles from female pear psylla: Effects of female age, mating status, and presence of host plant. Can. Entomol. 2008, 140, 184–191. [Google Scholar] [CrossRef]
- Horton, D.R.; Landolt, P.J. Attraction of male pear psylla, Cacopsylla pyricola, to female-infested pear shoots. Entomol. Exp. Appl. 2007, 123, 177–183. [Google Scholar] [CrossRef]
- Guédot, C.; Horton, D.R.; Landolt, P.J.; Munyaneza, J.E. Effect of mating on sex attraction in B actericera cockerelli with evidence of refractoriness. Entomol. Exp. Appl. 2013, 149, 27–35. [Google Scholar] [CrossRef]
- Lanier, G.N.; Qi, Y.-T.; West, J.R.; Park, S.C.; Webster, F.X.; Silverstein, R.M. Identification of the sex pheromone of three Matsucoccus pine bast scales. J. Chem. Ecol. 1989, 15, 1645–1659. [Google Scholar] [CrossRef]
- Tashiro, H.; Chambers, D.L. Reproduction in the California red scale, Aonidiella aurantii (Homoptera: Diaspididae). I. Discovery and extraction of a female sex pheromone. Ann. Entomol. Soc. Am. 1967, 60, 1166–1170. [Google Scholar] [CrossRef]
- Bakthavatsalam, N. Semiochemicals in Mealybugs. In Mealybugs and their Management in Agricultural and Horticultural Crops; Springer: Berlin/Heidelberg, Germany, 2016; pp. 173–198. [Google Scholar]
- Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M.; Hardie, J. The chemical ecology of aphids. Annu. Rev. Entomol. 1992, 37, 67–90. [Google Scholar] [CrossRef]
- Boullis, A.; Verheggen, F.J. Chemical ecology of aphids (Hemiptera: Aphididae). Biol. Ecol. Aphids 2016, 171, 171–198. [Google Scholar] [CrossRef]
- Blassioli-Moraes, M.C.; Khrimian, A.; Michereff, M.F.F.; Magalhães, D.M.; Hickel, E.; de Freitas, T.F.S.; Barrigossi, J.A.F.; Laumann, R.A.; Silva, A.T.; Guggilapu, S.D. Male-produced sex pheromone of Tibraca limbativentris revisited: Absolute configurations of zingiberenol stereoisomers and their influence on chemotaxis behavior of conspecific females. J. Chem. Ecol. 2020, 46, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Borges, M.; Birkett, M.; Aldrich, J.R.; Oliver, J.E.; Chiba, M.; Murata, Y.; Laumann, R.A.; Barrigossi, J.A.; Pickett, J.A.; Moraes, M.C.B. Sex attractant pheromone from the rice stalk stink bug, Tibraca limbativentris Stal. J. Chem. Ecol. 2006, 32, 2749–2761. [Google Scholar] [CrossRef] [PubMed]
- Moraes, M.C.B.; Laumann, R.A.; Cokl, A.; Borges, M. Vibratory signals of four Neotropical stink bug species. Physiol. Entomol. 2005, 30, 175–188. [Google Scholar] [CrossRef]
- Borges, M.; Millar, J.G.; Laumann, R.A.; Moraes, M.C.B. A male-produced sex pheromone from the neotropical redbanded stink bug, Piezodorus guildinii (W.). J. Chem. Ecol. 2007, 33, 1235–1248. [Google Scholar] [CrossRef]
- Baker, T.C.; Smyers, E.; Urban, J.; Meng, Z.; Damadaram, K.P.; Myrick, A.J.; Cooperband, M.; Domingue, M. Progression of seasonal activities of adults of the spotted lanternfly, Lycorma delicatula, during the 2017 season of mass flight dispersal behavior in eastern Pennsylvania. J. Asia-Pac. Entomol. 2019, 22, 705–713. [Google Scholar] [CrossRef]
- Baker, T.; Myrick, A.; Wolfin, M.; Wang, Y. Visual Responses of Flight-Dispersing Spotted Lanternflies, Lycorma delicatula toward a Tall Vertical Silhouette in a Vineyard. J. Insect Behav. 2021, 34, 49–60. [Google Scholar] [CrossRef]
- Cardé, R.T.; Baker, T.C. Sexual communication with pheromones. In Chemical Ecology of Insects; Bell, W.J., Cardé, R.T., Eds.; Springer: London, UK, 1984; pp. 355–383. [Google Scholar]
- Cardé, R.T. Defining attraction and aggregation pheromones: Teleological versus functional perspectives. J. Chem. Ecol. 2014, 40, 519–520. [Google Scholar] [CrossRef]
- Mason, R.T.; Fales, H.M.; Jones, T.H.; O’Brien, L.B.; Taylor, T.W.; Hogue, C.L.; Blum, M.S. Characterization of fulgorid waxes (Homoptera: Fulgoridae: Insecta). Insect Biochem. 1989, 19, 737–740. [Google Scholar] [CrossRef]
- O’Brien, L.B.; Wilson, S.W. Planthopper systematics and external morphology. In The Leafhoppers and Planthoppers; Nault, L.R., Rodriguez, J.G., Eds.; John Wiley & Sons: New York, NY, USA, 1985; pp. 61–102. [Google Scholar]
- Cho, S.-R.; Lee, J.-E.; Jeong, J.-W.; Yang, J.-O.; Yoon, C.; Kim, G.-H. Comparison of cuticular hydrocarbons of different developmental stages of the spot clothing wax cicada, Lycorma delicatula (Hemiptera: Fulgoridae). Korean J. Appl. Entomol. 2011, 50, 185–194. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faal, H.; Cooperband, M.F.; Canlas, I.; Carrillo, D. Evidence of Pheromone Use in a Fulgorid, Spotted Lanternfly. Forests 2022, 13, 1639. https://doi.org/10.3390/f13101639
Faal H, Cooperband MF, Canlas I, Carrillo D. Evidence of Pheromone Use in a Fulgorid, Spotted Lanternfly. Forests. 2022; 13(10):1639. https://doi.org/10.3390/f13101639
Chicago/Turabian StyleFaal, Hajar, Miriam F. Cooperband, Isaiah Canlas, and Daniel Carrillo. 2022. "Evidence of Pheromone Use in a Fulgorid, Spotted Lanternfly" Forests 13, no. 10: 1639. https://doi.org/10.3390/f13101639
APA StyleFaal, H., Cooperband, M. F., Canlas, I., & Carrillo, D. (2022). Evidence of Pheromone Use in a Fulgorid, Spotted Lanternfly. Forests, 13(10), 1639. https://doi.org/10.3390/f13101639