
Biscogniauxia Charcoal Canker—A New Potential Threat for Mid-European Forests as an Effect of Climate Change

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fieldwork
2.2. Molecular Analysis
2.2.1. Preparation of Isolates for Molecular Analysis and DNA Extraction
2.2.2. PCR Assays
2.2.3. Sequencing and Data Analysis
2.3. Meteorological and Hydrological Data Analysis
3. Results
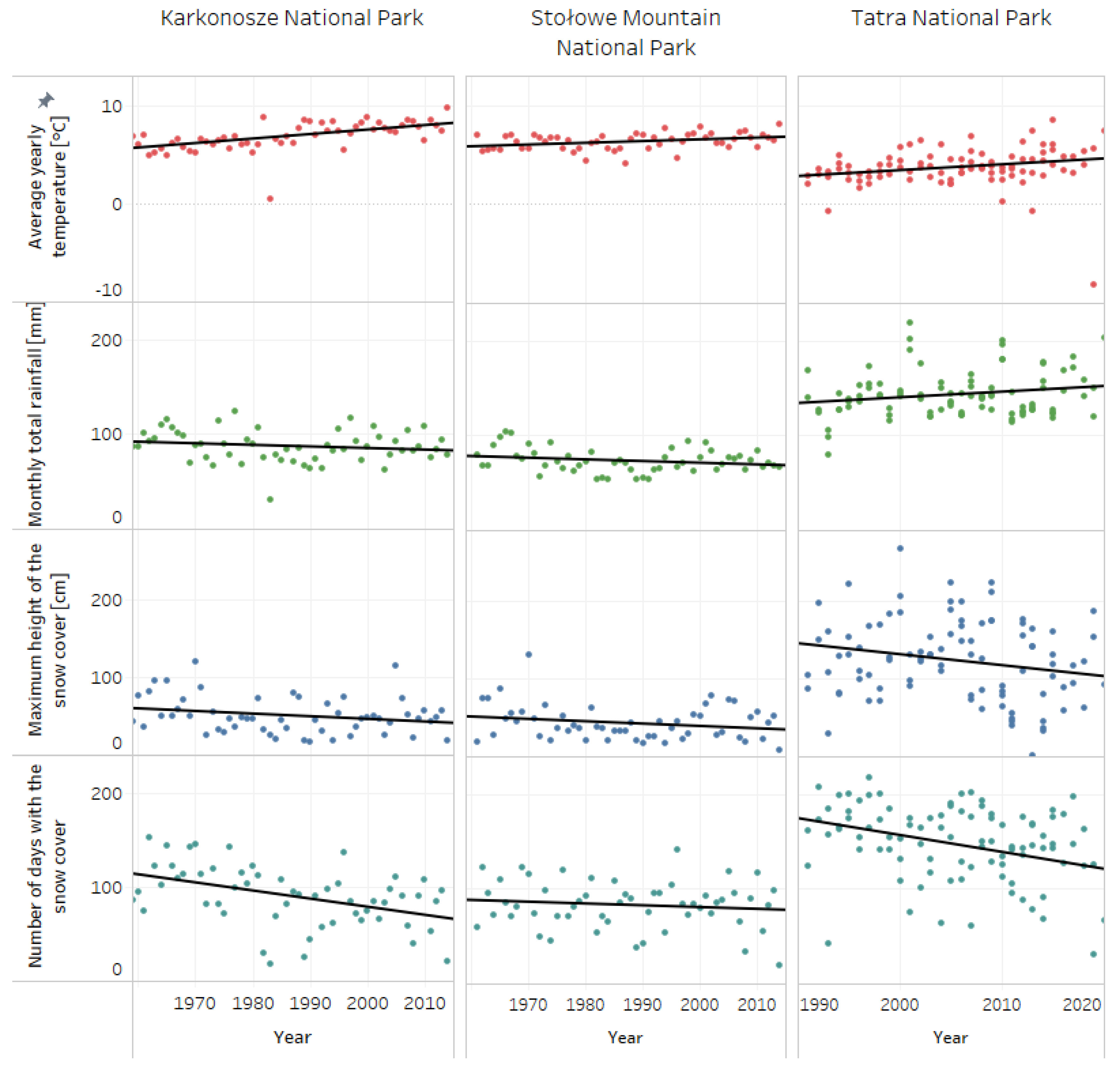
3.1. Meteorological Data
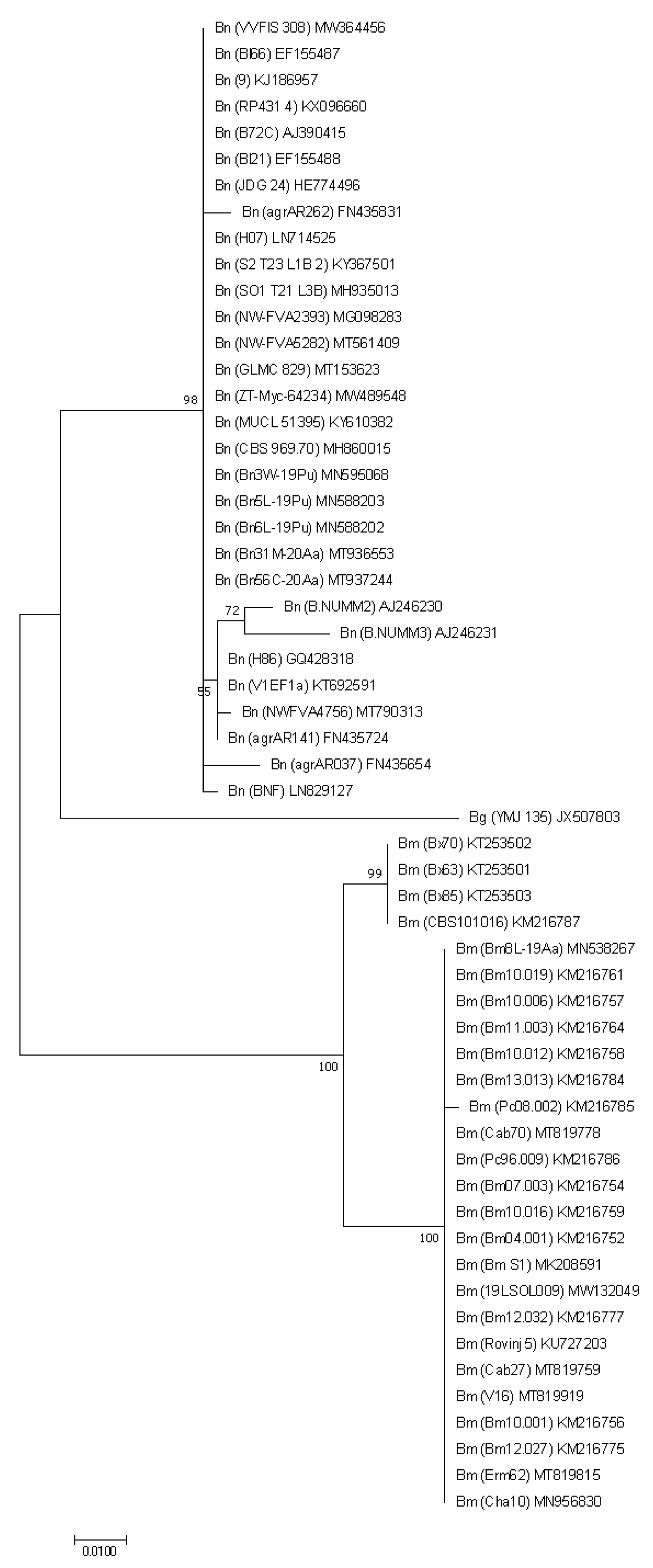
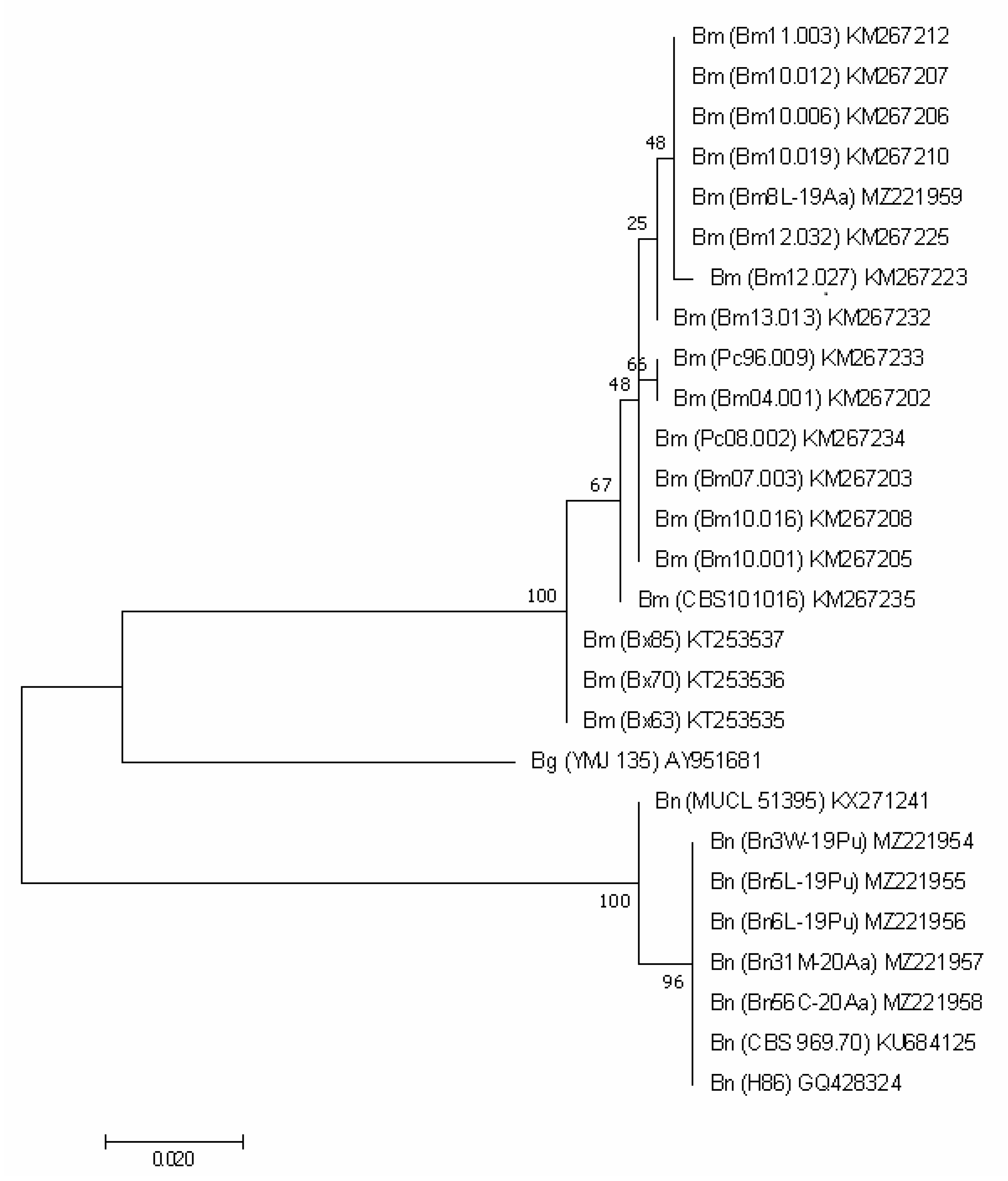
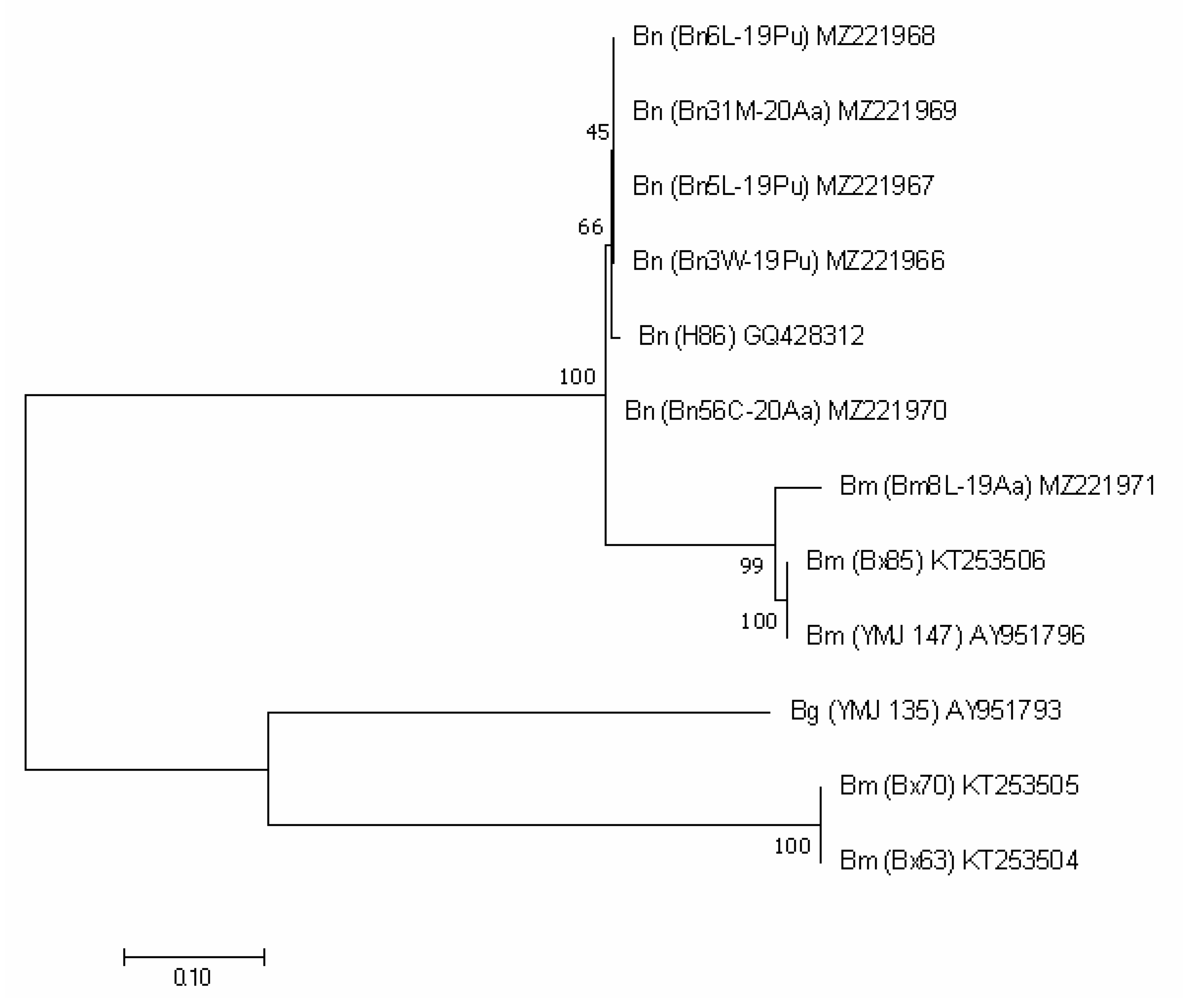
3.2. Molecular Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics
References
- Ragazzi, A.; Dellavalle Fedi, I.; Mesturino, L. The oak decline: A new problem in Italy. Eur. J. For. Pathol. 1989, 19, 105–110. [Google Scholar] [CrossRef]
- Granata, G.; Sidoti, A. Biscogniauxia nummularia: Pathogenic agent of a beech decline. For. Pathol. 2004, 34, 363–367. [Google Scholar] [CrossRef]
- Nugent, L.K.; Sihanonth, P.; Thienhirun, S.; Whalley, A.J.S. Biscogniauxia: A genus of latent in-vaders. Mycologist 2005, 19, 40–43. [Google Scholar] [CrossRef]
- Luchi, N.; Capretti, P.; Feducci, M.; Vannini, A.; Ceccarelli, B.; Vettraino, A.M. Latent infection of Biscogniauxia nummularia in Fagus sylvatica: A possible bioindicator of beech health conditions. iForest 2015, 9, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Henriques, J.; Nóbrega, F.; Sousa, E.; Lima, A. Morphological and genetic diversity of Biscogniauxia mediterranea associated to Quercus suber in the Mediterranean Basin. Rev. Ciênc. Agrár. 2015, 38, 166–175. Available online: http://hdl.handle.net/10400.5/10652 (accessed on 29 November 2021).
- Yangui, I.; Zouaoui Boutiti, M.; Vettraino, A.M.; Bruni, N.; Vannini, A.; Ben Jamaâ, M.L.; Boussaid, M.; Messaoud, C. Biscogniauxia mediterranea associated with cork oak (Quercus suber) in Tunisia: Relationships between phenotypic variation, genetic diversity and ecological factors. Fungal Ecol. 2019, 41, 224–233. [Google Scholar] [CrossRef]
- Luchi, N.; Capretti, P.; Vettraino, A.M.; Vannini, A.; Pinzani, P.; Pazzagli, M. Early detection of Biscogniauxia nummularia in symptomless European beech (Fagus sylvatica L.) by TaqMan™real-time PCR. Lett. Appl. Microbiol. 2006, 43, 33–38. [Google Scholar] [CrossRef] [PubMed]
- La Porta, N.; Capretti, P.; Thomsen, I.M.; Kasanen, R.; Hietala, A.M.; Von Weissenberg, K. Forest pathogens with higher damage potential due to climate change in Europe. Can. J. Plant Pathol. 2008, 30, 177–195. [Google Scholar] [CrossRef]
- Zabalgogeazcoa, I.; Pedro, J.; Canals, R.M. Biscogniauxia nummularia infecting beech (Fagus sylvatica) trees and sympatric plants of the sedge Carex brevicollis. For. Pathol. 2015, 45, 346–348. [Google Scholar] [CrossRef]
- Paoletti, E.; Goggioli, V.; Maresi, G. Reperti diB. nummularia su faggio in Italia. [Biscogniauxia nummularia on beech in Italy]. Micol. Ital. 1996, 1, 27–35. (In Italian) [Google Scholar]
- Moriondo, F.; Mugnai, L. A case of group dying of beech in Italy. In Proceedings of the 8th Congress of the Mediterranean Phytopathological Union, Agadir, Morocco, 28 October–3 November 1990; pp. 121–122. [Google Scholar]
- Granata, G.; Whalley, A.J.S. Decline of beech associated to Biscogniauxia nummularia in Italy. Petria 1994, 4, 111–116. [Google Scholar]
- Petrini, L.; Petrini, O. Xylariaceous fungi as endophytes. Sydowia 1985, 38, 216–234. [Google Scholar]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fungal Biol. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Osono, T.; Tateno, O.; Masuya, H. Diversity and ubiquity of xylariaceous endophytes in live and dead leaves of temperate forest trees. Mycoscience 2013, 54, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Lakatos, F.; Molnár, M. Mass Mortality of Beech (Fagus sylvatica L.) in South-West Hungary. Acta Silv. Lign. Hung. 2009, 5, 75–82. [Google Scholar]
- Zíbarová, L.; Kout, J. Xylariaceous pyrenomycetes from Bohemia: Species of Biscogniauxia and Hypoxylon new to the Czech Republic, and notes on other rare species. Czech. Mycol. 2017, 69, 77–108. [Google Scholar] [CrossRef]
- Vujanovic, V.; Kim, S.H.; Latinovic, J.; Latinovic, N. Natural Fungicolous Regulators of Biscogniauxia destructiva sp. nov. That Causes Beech Bark Tarcrust in Southern European (Fagus sylvatica) Forests. Microorganisms 2020, 8, 1999. [Google Scholar] [CrossRef]
- Ju, Y.M.; Rogers, J.D.; San Martín, F.; Granmo, A. The genus Biscogniauxia. Mycotaxon 1998, 66, 1–98. [Google Scholar]
- Patejuk, K.; Baturo-Cieśniewska, A.; Kaczmarek-Pieńczewska, A.; Pusz, W. Mycobiota of peat-bog pine (Pinus × rhaetica) needles in the Stołowe Mountains National Park, Poland. Nova Hedwigia 2021, 112, 253–265. [Google Scholar] [CrossRef]
- Pusz, W.; Kaczmarek, A.; Baturo-Cieśniewska, A. Ocena zdrowotności jodły w Karkonoskim Parku Narodowym. Annual Report of Project Funded by Forest Fund; Karkonoski National Park: Jelenia Góra, Poland, 2020. [Google Scholar]
- Pusz, W.; Baturo-Cieśniewska, A.; Kaczmarek-Pieńczewska, A.; Zwijacz-Kozica, T.; Patejuk, K. The mycobiota of needles and shoots of silver fir (Abies alba Mill.) with symptoms of Herpotrichia needle browning in the Tatra Mts. (Poland). Ann. For. Res. 2020, 63, 45–56. [Google Scholar] [CrossRef]
- Vannini, A.; Mazzaglia, A.; Anselmi, N. Use of random amplified polymorphic DNA (RAPD) for detection of genetic variation and proof of the heterothallic mating system in Hypoxylon mediterraneum. Eur. J. Plant Pathol. 1999, 29, 209–218. [Google Scholar] [CrossRef]
- Eaton, E.; Caudullo, G.; Oliveira, S.; de Rigo, D. Quercus robur and Quercus petraea in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; p. e01c6df+. [Google Scholar]
- Ragazzi, A.; Ginetti, B.; Moricca, S. First report of Biscogniauxia mediterranea on English ash in Italy. Plant Dis. 2012, 96, 1694. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.-J.; Lasch, P.; Eggers, J.; van der Maaten-Theunissen, M.; et al. Climate change and European forests: What do we know, what are the uncertainties, and what are theimplications for forest management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef] [Green Version]
- Hendry, S.J.; Boddy, L.; Lonsdale, D. Abiotic variables effect differentialexpression of latent infections in beech (Fagus sylvatica). New Phytol. 2002, 155, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Dulamsuren, C.; Hauck, M.; Kopp, G.; Ruff, M.; Leuschner, C. European beech responds to climate change with growth decline at lower, and growth increase at higher elevations in the center of its distribution range (SW Germany). Trees 2016, 31, 673–686. [Google Scholar] [CrossRef]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance in major European temperate foresttrees: Climatic drivers and levels of variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, J.; Hauck, M.; Dulamsuren, C.; Leuschner, C. Climate warming-related growth decline affectsFagus sylvatica, but not other broad-leaved tree species in Central Europeanmixed forests. Ecosystems 2015, 18, 560–572. [Google Scholar] [CrossRef]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage; Springer: New York, NY, USA, 2009; 520p. [Google Scholar] [CrossRef]
- Watanabe, T. Pictorial Atlas of Soil and Seed Fungi: Morphologies of Cultured Fungi and Key to Species, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2010; 426p. [Google Scholar] [CrossRef]
- Ellis, M.B.; Ellis, J.P. Microfungi on Land Plants an Identification Handbook; Richmond Publishing: Slough, UK, 1997. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Baturo-Cieśniewska, A.; Pusz, W.; Patejuk, K. Problems, limitations, and challenges in species identification of Ascomycota members on the basis of ITS regions. Acta Mycol. 2020, 55, 5512. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- BLAST Database. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome (accessed on 11 August 2021).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Institute of Meteorology and Water Management–National Research Institute. Available online: https://danepubliczne.imgw.pl/ (accessed on 22 December 2021).
- Chlebicki, A. Some overlooked and rare Xylariaceous fungi from Poland. Polish Bot. J. 2008, 53, 71–80. [Google Scholar]
- Mułenko, W.; Majewski, T.; Ruszkiewicz-Michalska, M. A preliminary checklist of micromycetes in Poland. In Szafer Institute of Botany; Polish Academy of Sciences: Kraków, Poland, 2008. [Google Scholar]
- Houston Durrant, T.; de Rigo, D.; Caudullo, G. Fagus sylvatica and other beeches in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; p. e012b90+. [Google Scholar]
- Herbich, J. (Ed.) Lasy i Bory. Poradniki Ochrony Siedlisk i Gatunków Natura 2000—Podręcznik Metodyczny; Ministerstwo Środowiska: Warszawa, Poland, 2004; Volume 5, p. 344. [Google Scholar]
- European Commission DG Environment. Interpretation manual of European Union habitats—EUR 28. Nature ENV B.3. 2013. Available online: https://ec.europa.eu/environment/nature/legislation/habitatsdirective/docs/Int_Manual_EU28.pdf (accessed on 5 October 2021).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | No of Isolate | Host | Place of Isolation | Part of the Plant | References |
|---|---|---|---|---|---|
| B. nummularia (Bull.) Kuntze | Bn3W-19Pu Bn5L-19Pu Bn6L-19Pu | Pinus × rhaetica Brügger | Great Peat Bog of Batorów—Stołowe Mountains National Park (SW Poland) | fresh needles | [20] |
| B. nummularia (Bull.) Kuntze | Bn31M-20Aa Bn56C-20Aa | Abies alba Mill. | Karkonosze National Park (SW Poland) | fresh needles | [21] |
| B. mediterranea (De Not.) Kuntze | Bm8L-19Aa | Abies alba Mill. | Tatra National Park (S Poland) | shoots | [22] |
| Target | Primer Name and Sequence 5′→ 3′ | Annealing Conditions | Reference |
|---|---|---|---|
| ITS | ITS1: TCCGTAGGTGAACCTGCGG ITS4: TCCTCCGCTTATTGATATGC | 55 °C/45 s | [37] |
| TEF1 | EF1-728F: CATCGAGAAGTTCGAGAAGG EF1-986R: TACTTGAAGGAACCCTTACC | 54 °C/1 min | [38] |
| TUB2 | Bt2a: GGTAACCAAATCGGTGCTGCTTTC Bt2b: ACCCTCAGTGTAGTGACCCTTGGC | 58 °C/1 min | [39] |
| ACT | ACT-512F: ATGTGCAAGGCCGGTTTCGC ACT-783R: TACGAGTCCTTCTGGCCCAT | 60 °C TD-PCR * | [38] |
| Collection Symbol | Locality | Host | GenBank Accession Number | |||
|---|---|---|---|---|---|---|
| ITS | TEF1 | TUB2 | ACT | |||
| Biscogniauxia mediterranea (De Not.) Kuntze | ||||||
| Bm8L-19Aa | Poland | Abies alba Mill. | MN5382671 | MZ221965 | MZ221959 | MZ221971 |
| Bm04.001 | Portugal | Quercus suber L. | KM216752 | KM216788 | KM267202 | / 2 |
| Bm07.003 | Portugal | Quercus suber L. | KM216754 | KM216790 | KM267203 | / |
| Bm10.019 | Portugal | Quercus suber L. | KM216761 | KM216797 | KM267210 | / |
| Bm10.001 | Portugal | Quercus suber L. | KM216756 | KM216792 | KM267205 | / |
| Bm10.006 | Portugal | Quercus suber L. | KM216757 | KM216793 | KM267206 | / |
| Bm10.012 | Portugal | Quercus rotundifolia Lam. | KM216758 | KM216794 | KM267207 | / |
| Bm11.003 | Portugal | Quercus suber L. | KM216764 | KM216800 | KM267212 | / |
| Bm12.027 | Portugal | Quercus robur L. | KM216775 | KM216811 | KM267223 | / |
| Bm12.032 | Portugal | Quercus suber L. | KM216777 | KM216813 | KM267225 | / |
| Pc08.002 | Portugal | Quercus suber L. | KM216785 | KM216822 | KM267234 | / |
| Pc96.009 | Portugal | Quercus suber L. | KM216786 | KM216821 | KM267233 | / |
| Erm62 | Portugal | Quercus suber L. | MT819815 | / | / | / |
| Cab27 | Portugal | Quercus suber L. | MT819759 | / | / | / |
| Cab70 | Portugal | Quercus suber L. | MT819778 | / | / | / |
| Cha10 | Portugal | Prunus dulcis (Mill.) D.A.Webb | MN956830 | / | / | / |
| V16 | Portugal | Quercus suber L. | MT819919 | / | / | / |
| Bm10.016 | Italy, Sardinia | Quercus suber L. | KM216759 | KM216795 | KM267208 | / |
| Bx63 | Italy | Quercus pubescens Willd. | KT253501 | KT253535 | KT253504 | |
| Bx70 | Italy | Quercus pubescens Willd. | KT253502 | / | KT253536 | KT253505 |
| Bx85 | Italy | Quercus pubescens Willd. | KT253503 | / | KT253537 | KT253506 |
| Bm_S1 | Croatia | Quercus ilex L. | MK208591 | / | / | / |
| Rovinj 5 | Croatia | Quercus ilex L. | KU727203 | / | / | / |
| 19LSOL009 | France | Lycium hybrid cultivar L. | MW132049 | / | / | / |
| YMJ 147 | France | / | / | / | // 3 | AY951796 |
| CBS101016 | Netherlands | Quercus robur L. | KM216787 | KM216823 | KM267235 | / |
| Bm13.013 | Spain | Quercus suber L. | KM216784 | KM216820 | KM267232 | / |
| Biscogniauxia nummularia (Bull.) Kuntze | ||||||
| Bn3W-19Pu | Poland | Pinus mugo subsp. uncinata (Ramond) Domin | MN595068 | MZ221960 | MZ221954 | MZ221966 |
| Bn5L-19Pu | Poland | Pinus mugo subsp. uncinata (Ramond) Domin | MN588203 | MZ221961 | MZ221955 | MZ221967 |
| Bn6L-19Pu | Poland | Pinus mugo subsp. uncinata (Ramond) Domin | MN588202 | MZ221962 | MZ221956 | MZ221968 |
| Bn31M-20Aa | Poland | Abies alba Mill. | MT936553 | MZ221963 | MZ221957 | MZ221969 |
| Bn56C-20Aa | Poland | Abies alba Mill. | MT937244 | MZ221964 | MZ221958 | MZ221970 |
| GLMC 829 | Germany | Prunus avium L. | MT153623 | / | / | / |
| NWFVA4756 | Germany | Pinus sylvestris L. | MT790313 | / | / | / |
| NW-FVA5282 | Germany | Fagus sylvatica L. | MT561409 | / | / | / |
| NW-FVA2393 | Germany | Pinus sylvestris L. | MG098283 | / | / | / |
| agrAR262 | Germany | Viscum album subsp. austriacum (Wiesb.) Volim. | FN435831 | / | / | / |
| agrAR037 | Germany | Viscum album subsp. austriacum (Wiesb.) Volim. | FN435654 | / | / | / |
| agrAR141 | Germany | Viscum album subsp. austriacum (Wiesb.) Volim. | FN435724 | / | / | / |
| RP431_4 | Germany | endophyte on plant | KX096660 | / | / | / |
| BI21 | Germany | Fagus sylvatica L. | EF155488 | / | / | / |
| BI66 | Germany | / | EF155487 | / | / | / |
| B.NUMM2 | Italy | / | AJ246230 | / | / | / |
| B.NUMM3 | Italy | / | AJ246231 | / | / | / |
| S2_T23_L1B_2 | Italy | Fraxinus ornus L. | KY367501 | / | / | / |
| MUCL 51395 | France | / | KY610382 | / | KX271241 | |
| V1EF1a | France | wheat | KT692591 | / | / | / |
| ZT-Myc-64234 | Switzerland | Fagus sylvatica L. | MW489548 | / | / | / |
| SO1_T21_L3B | Switzerland | Fraxinus ornus L. | MH935013 | / | / | / |
| JDG 24 | Slovenia | Fraxinus excelsior L. | HE774496 | / | / | / |
| 9 | Slovenia | olive tree | KJ186957 | / | / | / |
| B72C | Spain | / | AJ390415 | / | / | / |
| BNF | Spain | Fagus sylvatica L. | LN829127 | / | / | / |
| H07 | Czech Republic | / | LN714525 | / | / | / |
| VVFIS_308 | Hungary | Vitis vinifera L. | MW364456 | / | / | / |
| H86 | Slovakia | Salix alba L. | GQ428318 | / | GQ428324 | GQ428312 |
| CBS 969.70 | United Kingdom | Fagus sylvatica L. | MH860015 | / | KU684125 | / |
| Biscogniauxia granmoi Lar.N. Vassiljeva | ||||||
| YMJ 135 | Austria | Prunus padus L. | JX507803 | / | AY951681 | AY951793 |
| Biscogniauxia petrensis Z.F. Zhang, F. Liu and L. Cai | ||||||
| LC5697 | China | rock | // | KX855215 | // | // |
| Karkonosze National Park | Stołowe Mountain National Park | Tatra National Park | ||
|---|---|---|---|---|
| Average yearly temperature | p-value | <0.0001 | 0.0141009 | 0.031045 |
| R-squared | 0.349813 | 0.110399 | 0.049553 | |
| Equation: | x = 0.0457126y + −83.9245 | x = 0.0175113y + −28.4773 | x = 0.0591253y + −114.823 | |
| Monthly total rainfall | p-value | 0.147077 | 0.108164 | 0.0582271 |
| R-squared | 0.0336043 | 0.0488743 | 0.0384444 | |
| Equation: | x = −0.166531y + 418.273 | x = −0.176304y + 423.724 | x = 0.597223y + −1054.27 | |
| Maximum height of snow cover | p-value | 0.0343486 | 0.121498 | 0.0406373 |
| R-squared | 0.0702087 | 0.0454902 | 0.0447694 | |
| Equation: | x = −0.33734y + 720.542 | x = −0.302306y + 643.111 | x = −1.40737y + 2945.36 | |
| Number of days with snow cover | p-value | <0.0001 | 0.390762 | 0.000853 |
| R-squared | 0.243498 | 0.0141997 | 0.114448 | |
| Equation: | x = −0.861401y + 1801.51 | x = −0.188565y + 457.828 | x = −1.78431y + 3725.48 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patejuk, K.; Baturo-Cieśniewska, A.; Pusz, W.; Kaczmarek-Pieńczewska, A. Biscogniauxia Charcoal Canker—A New Potential Threat for Mid-European Forests as an Effect of Climate Change. Forests 2022, 13, 89. https://doi.org/10.3390/f13010089
Patejuk K, Baturo-Cieśniewska A, Pusz W, Kaczmarek-Pieńczewska A. Biscogniauxia Charcoal Canker—A New Potential Threat for Mid-European Forests as an Effect of Climate Change. Forests. 2022; 13(1):89. https://doi.org/10.3390/f13010089
Chicago/Turabian StylePatejuk, Katarzyna, Anna Baturo-Cieśniewska, Wojciech Pusz, and Agata Kaczmarek-Pieńczewska. 2022. "Biscogniauxia Charcoal Canker—A New Potential Threat for Mid-European Forests as an Effect of Climate Change" Forests 13, no. 1: 89. https://doi.org/10.3390/f13010089
APA StylePatejuk, K., Baturo-Cieśniewska, A., Pusz, W., & Kaczmarek-Pieńczewska, A. (2022). Biscogniauxia Charcoal Canker—A New Potential Threat for Mid-European Forests as an Effect of Climate Change. Forests, 13(1), 89. https://doi.org/10.3390/f13010089