
Does the Reaction of Inflorescences and Flowers of the Invasive Prunus serotina Ehrh. to Various Herbicides Give Hope for Elimination of This Species from Polish Forests?

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Study Area
3. Materials and Methods
4. Statistical Analysis
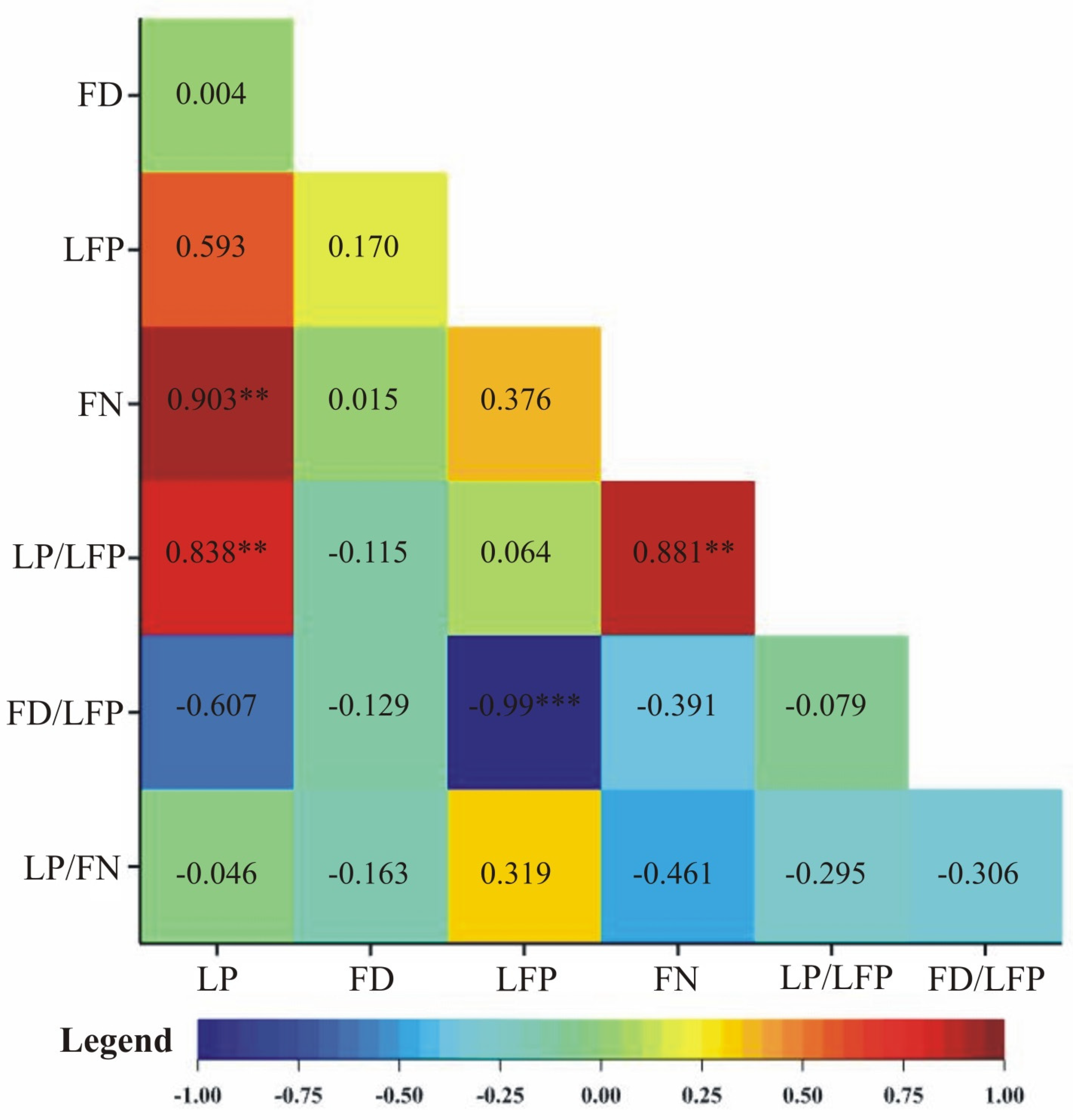
5. Results
6. Discussion
7. Conclusions
- This study revealed morphological differences among the seven (out of 9) tested features of P. serotina inflorescences and flowers collected from 10 control trees compared to the 29 trees treated with six various herbicides. Two features—stamen viability (SV) and pistil number (PN) were practically the same for all the trees investigated. The only exception was four flowers from tree no. 29 (treated with Logo), for which two pistils were found instead of one.
- A new aspect of the current research was a long-term analysis of the herbicide impact on flower and inflorescence development. The investigated P. serotina trees were treated with six types of chemicals at the beginning of April 2020, and the generative material for the study was harvested at full flowering in June 2021. Thus our experiment lasted over a year. As a rule, in other investigations with herbicides, their effectiveness is checked within a short period of at most several weeks after application, where inhibition of growth and plant development is visible after only a few days. The doses of chemicals used are intended to cause plant death. Therefore it is not possible to check the side effects of herbicides on plant development in the longer term. Thus, our research supplements knowledge on the herbicide influence after a relatively long time after its application.
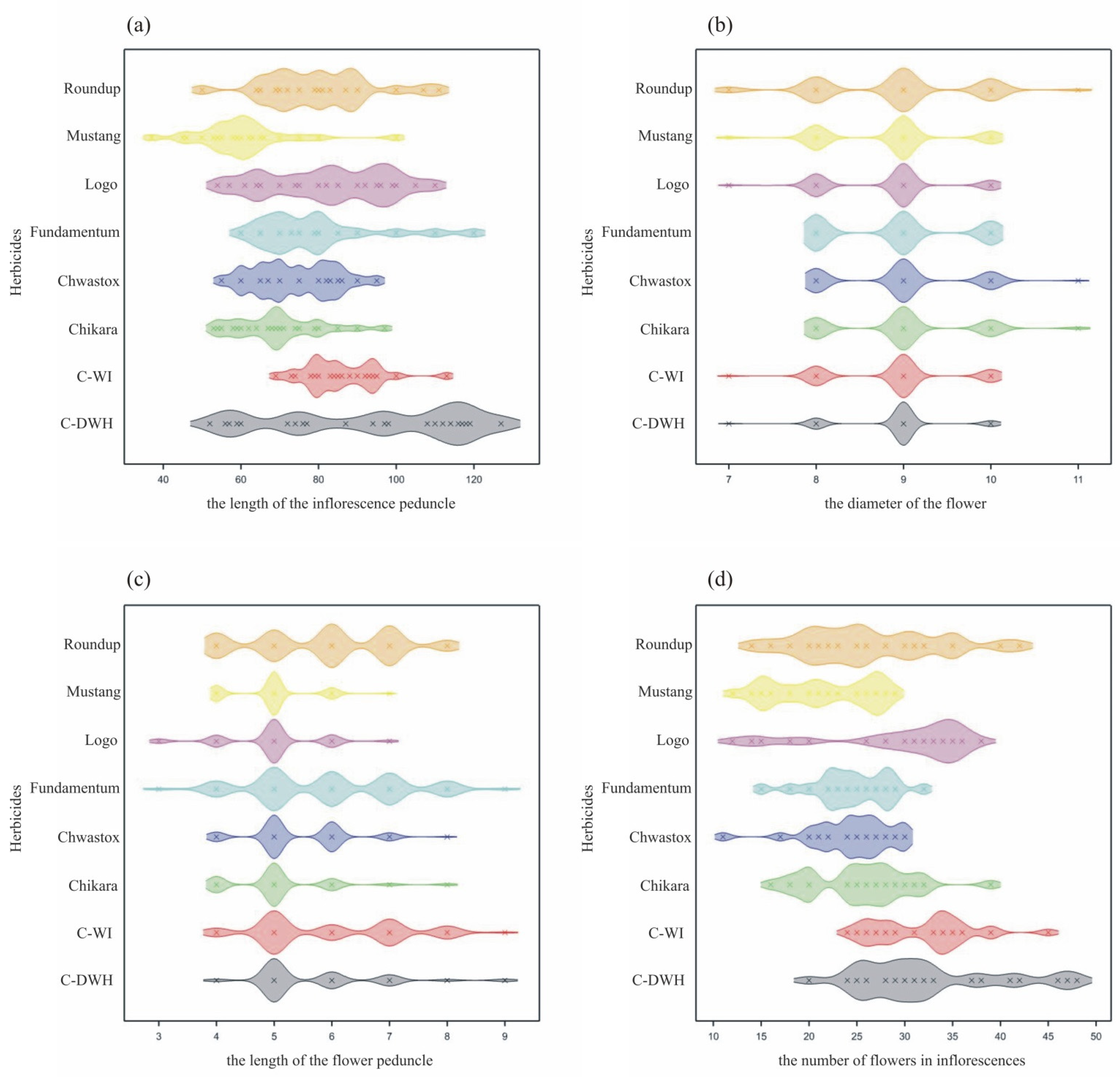
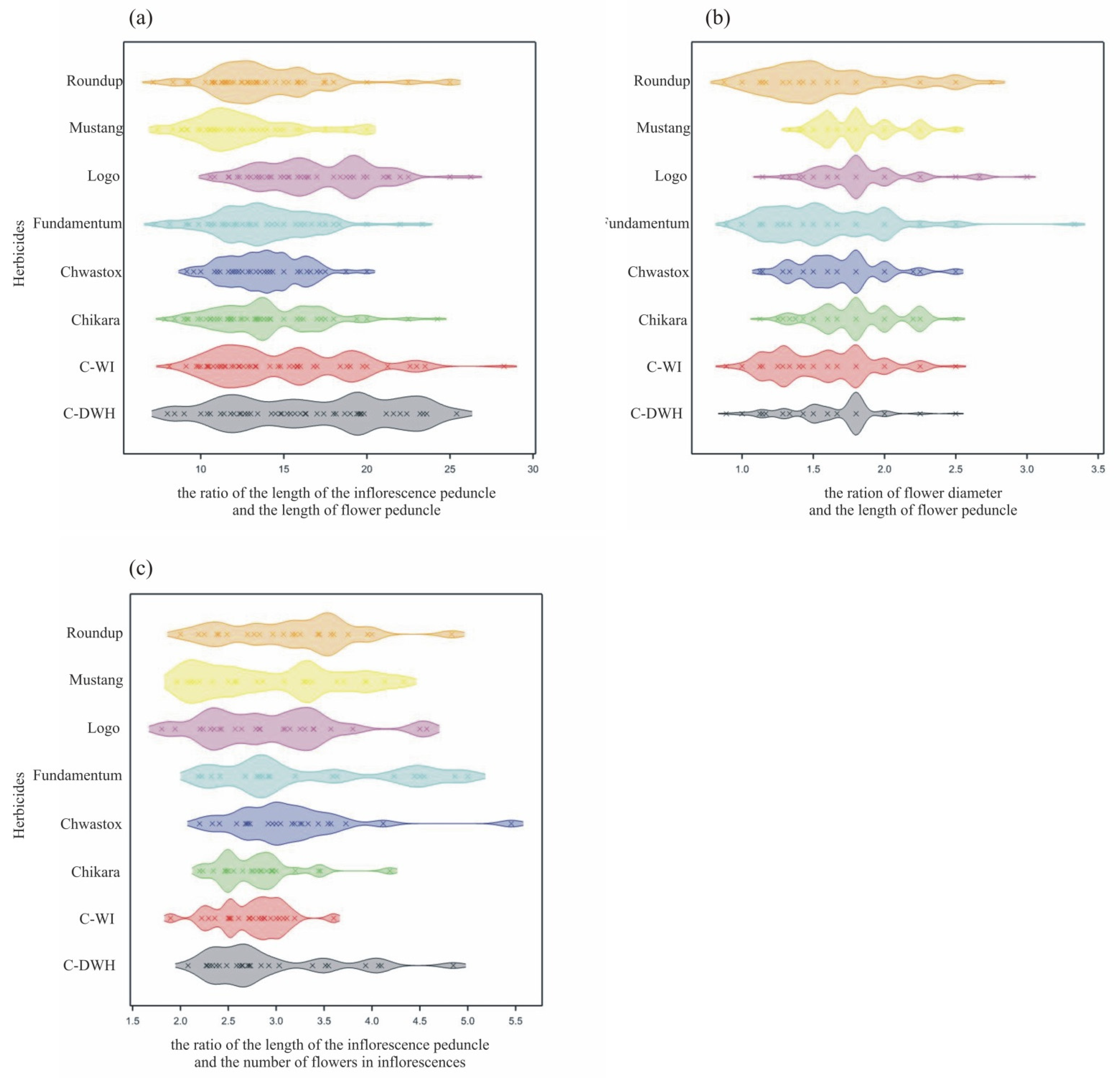
- The largest ranges of values of the studied features were observed in inflorescences and flowers collected from the control trees (C-WI, C-DWH). The average values of these features were similar and most often higher than the values for trees treated with herbicides.
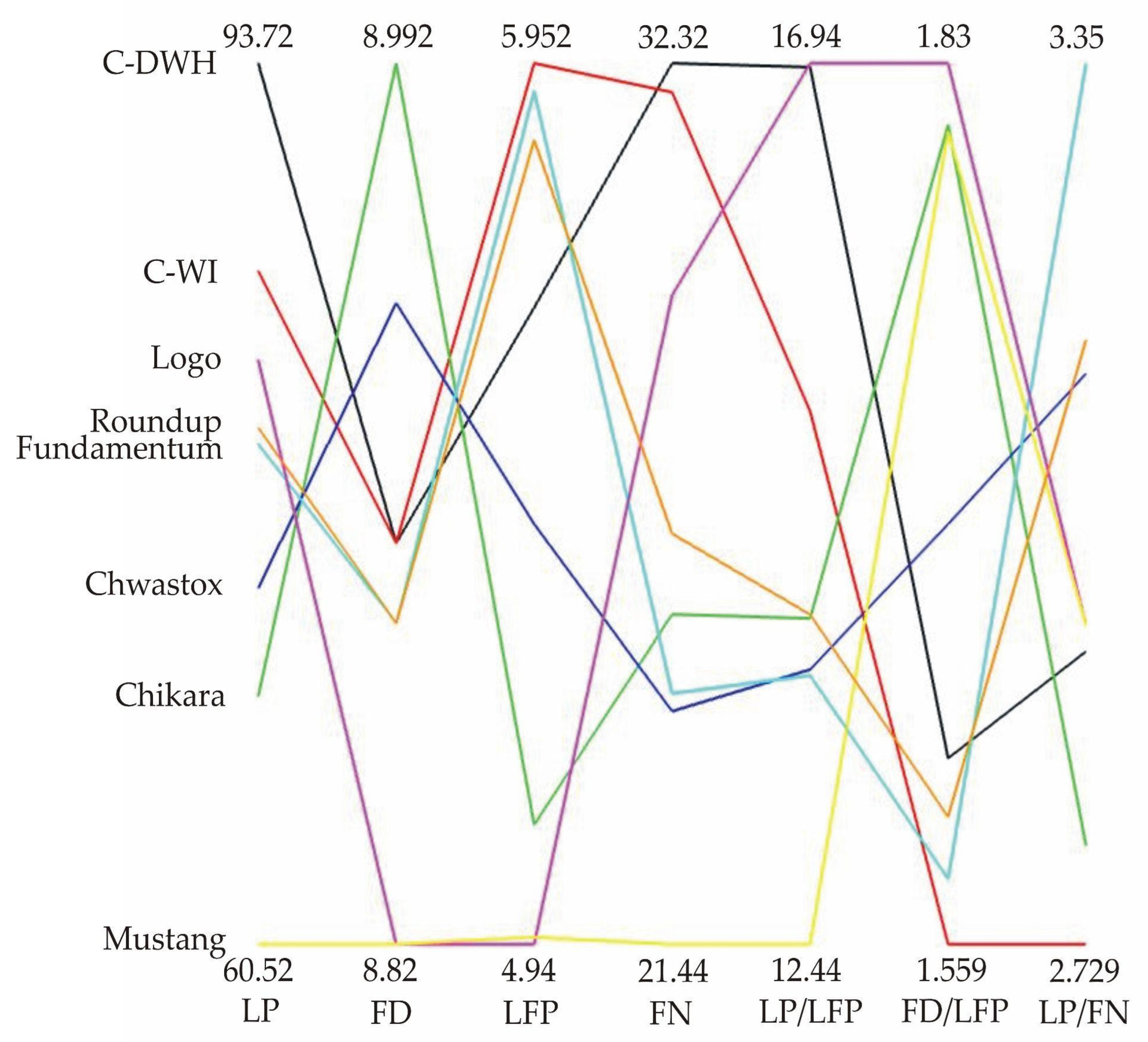
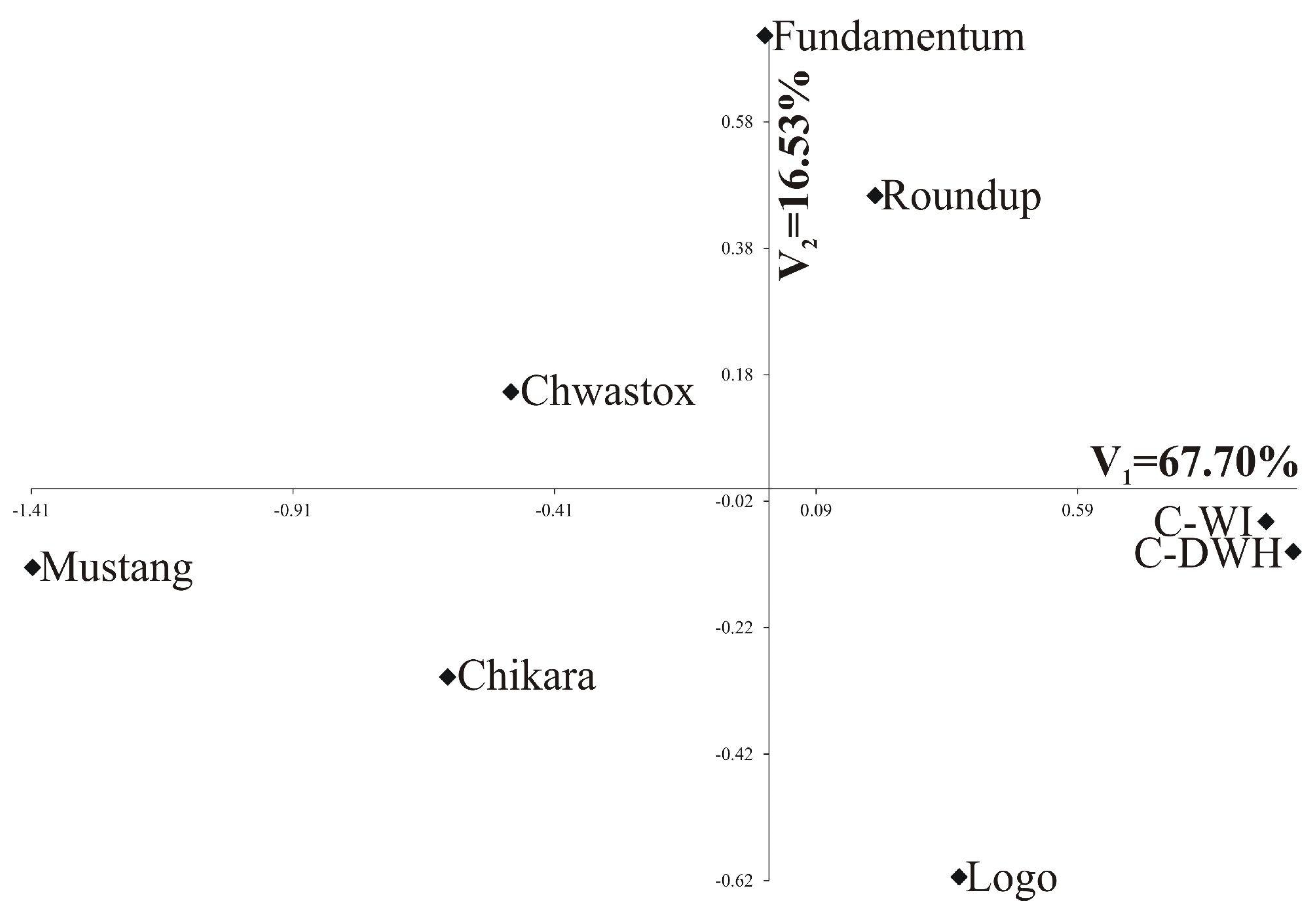
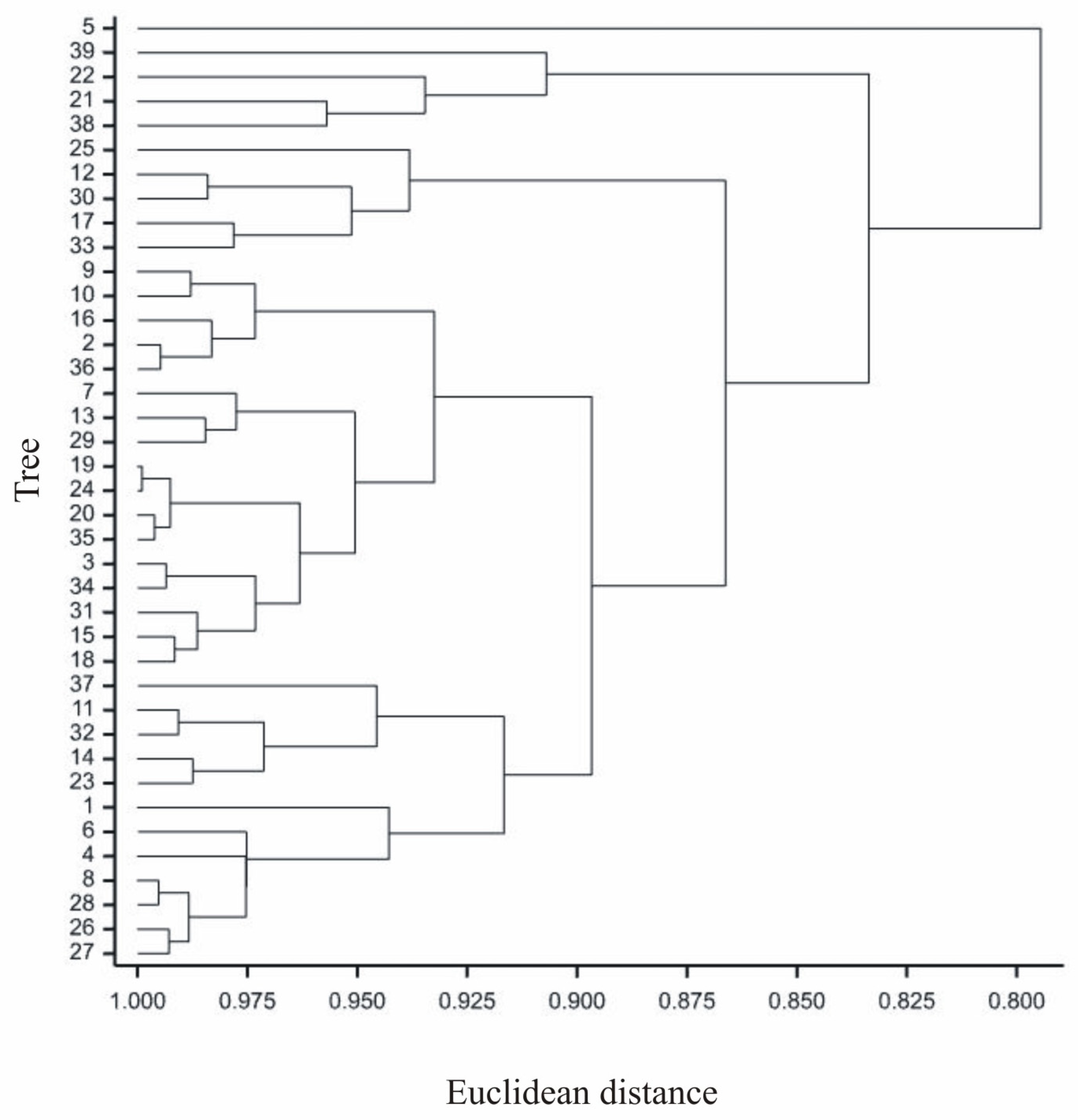
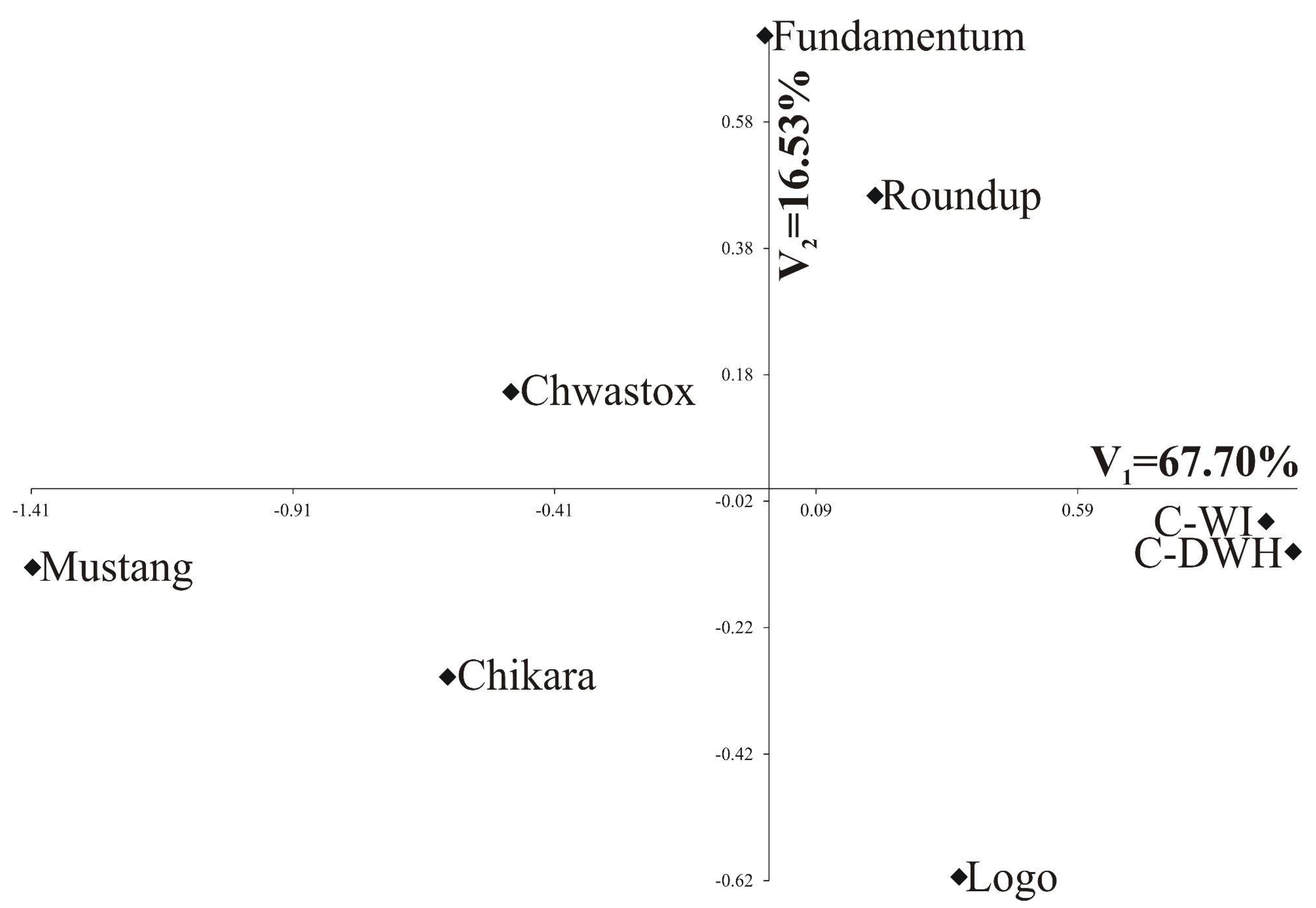
- The greatest variability in the features of inflorescences and flowers was found for C-DWH. The values of the features after the application of Fundamentum slightly differed from those obtained for the other herbicides, being the closest to Roundup. Similar features of the examined generative organs were observed in the trees treated with Logo and Roundup, as well as those treated with Chwastox, Chikara and Mustang (see Figure 5).
- This study contributes to the search for effective methods of combating the invasive black cherry in forests. The use of six different herbicides—with different active substance contents—to control P. serotina proves to be effective. One year after their application a reduction was observed in the size of inflorescences, as well as the number of flowers. This effect was obtained for all the herbicides used. It may indicate that already a small amount of the active substance is sufficient to reduce the size of inflorescences and the number of flowers in P. serotina. There is no doubt that a continuation of the research in the years following herbicide application will yield further valuable results.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Marquis, D.A. Prunus serotina Ehrh. Black Cherry. Silv. N. Am. 1990, 2, 594–604. [Google Scholar]
- Starfinger, U. NOBANIS–Invasive Alien Species Fact Sheet–Prunus serotina. Available online: https://www.nobanis.org/globalassets/speciesinfo/p/prunus-serotina/prunus-serotina.pdf (accessed on 24 September 2021).
- Fresnedo-Ramírez, J.; Segura, S.; Muratalla-Lúa, A. Morphovariability of Capulín (Prunus serotina Ehrh.) in the Central-Western Region of Mexico from a Plant Genetic Resources Perspective. Genet. Resour. Crop Evol. 2011, 58, 481–495. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, F.A.; Segura, S.; Fresnedo-Ramirez, J. Morphological Variation in Black Cherry (Prunus serotina Ehrh.) Associated with Environmental Conditions in Mexico and the United States. Genet. Resour. Crop Evol. 2018, 65, 2151–2168. [Google Scholar] [CrossRef]
- McVaugh, R. A Revision of the North American Black Cherries (Prunus serotina Ehrh., and Relatives). Brittonia 1951, 7, 279–315. [Google Scholar] [CrossRef]
- Munro, D.B.; Munro, G.A. The Biology of Canadian Weeds. 51. Prunus virginiana L. and P. serotina Ehrh. Can. J. Plant Sci. 1982, 61, 977–992. [Google Scholar]
- Prunus serotina Ehrh. | Plants of the World Online | Kew Science. Available online: http://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:730262-1 (accessed on 24 September 2021).
- Vanhellemont, M. Present and Future Population Dynamics of Prunus serotina in Forests in Its Introduced Range. Ph.D. Thesis, Ghent University, Ghent, Belgium, 2009. [Google Scholar]
- Rohrer, J.R. Prunus (Rosaceae). In Flora of North America; Editorial Committee, Ed.; Flora of North America North of Mexico; Oxford University Press: New York, NY, USA, 2014; Volume 9, pp. 363–365. [Google Scholar]
- Segura, S.; Guzmán-Díaz, F.; López-Upton, J.; Mathuriau, C.; López-Medina, J. Distribution of Prunus serotina Ehrh. in North America and Its Invasion in Europe. J. Geosci. Environ. Prot. 2018, 6, 111–124. (In Polish) [Google Scholar] [CrossRef] [Green Version]
- Tokarska-Guzik, B.; Dajdok, Z.; Zajc, M.; Zajc, A.; Urbisz, A.; Danielewicz, W.; Hodyski, C. Rośliny Obcego Pochodzenia w Polsce Ze Szczególnym Uwzględnieniem Gatunków Inwazyjnych; Generalna Dyrekcja Ochrony Środowiska: Warszawa, Poland, 2012. (In Polish)
- Dominik, T. Przyczynek Do Znajomości Wartości Hodowlanych Czeremchy Amerykańskiej. Sylwan 1947, 91, 123–132. (In Polish) [Google Scholar]
- Danielewicz, W. Rozsiedlenie czeremchy amerykańskiej [Prunus serotina Ehrh.] na terenie Nadleśnictwa Doświadczalnego Zielonka. Pr. Kom. Nauk Rol. Kom. Nauk Leśnych Pozn. Tow. Przyj. Nauk 1994, 78, 35–42. (In Polish) [Google Scholar]
- Halarewicz, A. Odnawianie się Czeremchy Amerykańskiej (Prunus serotina Ehrh.) Na Siedliskach Borowych. Sylwan 2011, 155, 530–534. (In Polish) [Google Scholar]
- Bijak, S.; Czajkowski, M.; Ludwisiak, Ł. Occurrence of Black Cherry (Prunus serotina Ehrh.) in the State Forests in Poland. For. Res. Pap. 2014, 75, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Danielewicz, W.; Wiatrowska, B. Inwazyjne Gatunki Drzew i Krzewów w Lasach Polski. Peckiana 2014, 9, 59–67. (In Polish) [Google Scholar]
- Dyderski, M.K.; Jagodziński, A.M. Wkraczanie Czeremchy Amerykańskiej Padus serotina (Ehrh.) Borkh. Do Olsów i Łęgów Olszowo-Jesionowych. Acta Sci. Pol. Silvarum Colendarum Ratio Ind. Lignaria 2015, 14, 103–113. (In Polish) [Google Scholar] [CrossRef]
- Halarewicz, A.; Otręba, A.; Danielewicz, W. Karta Informacyjna Gatunku. Padus serotina: Czeremcha Amerykańska. Available online: http://projekty.gdos.gov.pl/files/artykuly/127077/Padus-serotina_czeremcha-amerykanska_KG_WWW_icon.pdf (accessed on 21 September 2021).
- Matuszkiewicz, J.M. Zespoły Leśne Polsk; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 2007. (In Polish) [Google Scholar]
- Mędrzycki, P.; Otręba, A.; Pabjanek, P.; Brys, K.; Panufnik-Medrzycka, D.; Gwiazda, S.; Anczarska, J. Badania Rozmieszczenia i Liczebności Drzewiastych Inwazyjnych Gatunków Obcych (IGO) a Czynna Ochrona Zagrożonych Ekosystemów Leśnych w Kampinoskim Parku Narodowym. Studia Mater. Cent. Edukac. Przyr.-Leśnej 2007, 9, 245–256. (In Polish) [Google Scholar]
- Namura-Ochalska, A. Walka z Czeremchą Amerykańską Padus serotina (Ehrh.) Borkh.Ocena Skuteczności Wybranych Metod w Kampinoskim Parku Narodowym. Stud. Mater. CEPL W Rogowie 2012, 33, 190–200. (In Polish) [Google Scholar]
- Biereżnoj-Bazille, U.; Werpachowski, C. Inwazyjne gatunki roślin w ekosystemach Biebrzańskiego Parku Narodowego—pierwsze próby zwalczania. In Zwalczanie Inwazyjnych Gatunków Roślin Obcego Pochodzenia—Dobre i złe Doświadczenia; Krzysztofiak, L., Krzysztofiak, A., Eds.; Stowarzyszenie “Człowiek i Przyroda”: Krzywe, Poland, 2015; pp. 11–25. (In Polish) [Google Scholar]
- Otręba, A. Czeremcha amerykańska Padus serotina (Ehrh.) Borkh. In Metody Zwalczania Obcych Gatunków Roślin Występujących na Terenie Puszczy Kampinoskiej; Obidziński, A., Kołaczkowska, E., Otręba, A., Eds.; Wydawnictwo BioDar: Izabelin, Kraków, Poland, 2016; pp. 73–88. (In Polish) [Google Scholar]
- Starfinger, U.; Kowarik, I.; Rode, M.; Schepker, H. From Desirable Ornamental Plant to Pest to Accepted Addition to the Flora?–The Perception of an Alien Tree Species through the Centuries. Biol. Invasions 2003, 5, 323–335. [Google Scholar] [CrossRef]
- Van Den Meersschaut, D.; Lust, N. Comparison of Mechanical, Biological and Chemical Methods for Controlling Black Cherry (Prunus serotina) in Flanders (Belgium). Silva Gandav. 1997, 62, 90–109. [Google Scholar] [CrossRef] [Green Version]
- Csiszár, Á.; Korda, M. Summary of invasive plant control experiments. In Practical Experiences in Invasive Alien Plant Control; Csiszár, A., Korda, M., Eds.; Duna–Ipoly National Park Directorate: Budapest, Hungary, 2015; pp. 203–235. [Google Scholar]
- Demeter, L.; Lesku, B. Eradication of invasive alien plants in the southern Nyírség area of the Hortobágy National Park Directorate. In Practical Experiences in Invasive Alien Plant Control; Csiszár, A., Korda, M., Eds.; Duna–Ipoly National Park Directorate: Budapest, Hungary, 2015; pp. 75–80. [Google Scholar]
- Carpenter, D.J.; Mathiassen, S.K.; Boutin, C.; Strandberg, B.; Casey, C.S.; Damgaard, C. Effects of Herbicides on Flowering. Environ. Toxicol. Chem. 2020, 39, 1244–1256. [Google Scholar] [CrossRef]
- Korzeniewicz, R. Experiments on the Effectiveness of Herbicides Not Included in the List of “Recommended Plant Protection Products for Forestry Use” to Combat P. serotina. In Development of Methods of Combating the Black Cherry in Pine Stands; Korzeniewicz, R., Ed.; Research Project Implementation Report NR OR.271.3.13.2017; Generalna Dyrekcja Lasów Państwowych: Warszawa, Poland, 2020. (In Polish) [Google Scholar]
- Wang, X.; Gong, J.; Li, Q.; Wang, J.; Ma, Y.; Zhang, X.; Chang, Z.; Wen, J.; Zhao, L. Floral Organogenesis of Prunus laurocerasus and P. serotina and Its Significance for the Systematics of the Genus and Androecium Diversity in Rosaceae. Botany 2019, 97, 71–84. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Bocianowski, J.; Szulc, P.; Nowosad, K. Parallel Coordinate Plots of Maize Traits under Different Magnesium Applications. J. Integr. Agric. 2015, 14, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Rencher, A.C. Interpretation of Canonical Discriminant Functions, Canonical Variates, and Principal Components. Am. Stat. 1992, 46, 217–225. [Google Scholar] [CrossRef]
- Seidler-Łożykowska, K.; Bocianowski, J. Evaluation of Variability of Morphological Traits of Selected Caraway (Carum carvi L.) Genotypes. Ind. Crops Prod. 2012, 35, 140–145. [Google Scholar] [CrossRef]
- Mahalanobis, P.C. On the Generalized Distance in Statistics. Proc. Natl. Acad. Sci. India A 1936, 2, 49–55. [Google Scholar]
- Skomra, U.; Bocianowski, J.; Agacka, M. Agro-Morphological Differentiation between European Hop (Humulus lupulus L.) Cultivars in Relation to Their Origin. J. Food Agric. Environ. 2013, 11, 1123–1128. [Google Scholar]
- VSN International. GenStat for Windows, 18th ed.; VSN International: Hemel Hempstead, UK, 2015. [Google Scholar]
- Guzmán, F.A.; Segura, S.; Aradhya, M.; Potter, D. Evaluation of the Genetic Structure Present in Natural Populations of Four Subspecies of Black Cherry (Prunus serotina Ehrh.) from North America Using SSR Markers. Sci. Hortic. 2018, 232, 206–215. [Google Scholar] [CrossRef]
- Boutin, C.; Strandberg, B.; Carpenter, D.; Mathiassen, S.K.; Thomas, P.J. Herbicide Impact on Non-Target Plant Reproduction: What Are the Toxicological and Ecological Implications? Environ. Pollut. 2014, 185, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Morales-Flores, F.; King-Diaz, B.; Aguilar, M.I.; Lotina-Hansen, B. Weeds and their mechanisms of resistance to herbicide. In Handbook on Herbicides: Biological Activity, Classification and Health Environmental Implications; Kobajashi, D., Watanabe, E., Eds.; Global Agriculture Development; Nova Science Publishers, Inc.: New York, NY, USA, 2014; pp. 29–54. [Google Scholar]
- Yasuor, H.; Riov, J.; Rubin, B. Glyphosate-Induced Male Sterility in Glyphosate-Resistant Cotton (Gossypium hirsutum L.) Is Associated with Inhibition of Anther Dehiscence and Reduced Pollen Viability. Crop Prot. 2007, 26, 363–369. [Google Scholar] [CrossRef]
- Zhou, Q.; Liu, W.; Zhang, Y.; Liu, K.K. Action Mechanisms of Acetolactate Synthase-Inhibiting Herbicides. Pestic. Biochem. Physiol. 2007, 89, 89–96. [Google Scholar] [CrossRef]
- Yu, C.; Dong, J.; Hu, S.; Xu, A. Exposure to Trace Amounts of Sulfonylurea Herbicide Tribenuron-Methyl Causes Male Sterility in 17 Species or Subspecies of Cruciferous Plants. BMC Plant Biol. 2017, 17, 95. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Lian, J.; Gong, Q.; Ren, L.-S.; Huang, Z.; Xu, A.-X.; Dong, J.-G. Sublethal Application of Various Sulfonylurea and Imidazolinone Herbicides Favors Outcrossing and Hybrid Seed Production in Oilseed Rape. BMC Plant Biol. 2020, 20, 69. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Herbicide—Trade Name | Abbreviation | Herbicide Concentration | Active Substance Content | Manufacturer |
|---|---|---|---|---|
| Chikara 25 WG | Chikara | 1.96% | Flazasulfuron (a compound from the group of sulfonylurea derivatives)—25%. | Belchim Crop. Protection Belgium Brussels |
| Chwastox turbo 340 sl | Chwastox | 25% | MCPA (4-chloro-2-methylphenoxy) acetic acid (a compound from the group of phenoxy carboxylic acids)—(25.9%); Dicamba (a compound from the group of benzoic acid derivatives)—(3.4%). | CIECH Sarzyna Poland Warsaw |
| Fundamentum 700 wg + Asystent | Fundamentum | 0.29% Fundamentum 700 wg 0.1% Asystent | Tribenuron-methyl (a compound from the group of sulfonylurea derivatives)—(40.0%). Metsulfuron-methyl (a compound from the group of sulfonylurea derivatives)—(13.5%); Florasulam (a compound from the triazolopyrimidine group)—(16.5%). | Innvigo Poland Warsaw |
| Logo 310 WG + Mero 842 EC | Logo | 1.47% Logo 310 WG 0.1% Mero 842 EC | Foramsulfuron (a compound from the group of sulfonylurea derivatives)—(30.0%); Iodosulfuron-methyl sodium (a compound from the group of sulfonylurea derivatives)—(1.0%). | Bayer SAS France Lyon |
| Mustang Forte 195 SE | Mustang | 10% | Florasulam (a compound from the triazolopyrimidine group—(0.47%); Aminopyralid (a compound from the pyridine group—(0.94%); 2,4-D—a compound from the phenoxy acid group—(17%). | Corteva Agriscience™ U.S. Indianapolis |
| Roundup Flex 480 FL | Roundup | 50% | Glyphosate (a compound from the group of aminophosphonates in the form of potassium salt)—(35.75%). | Monsanto Europe S.A./N.V Belgium Diegem |
| Features | Herbicides | C-DWH | C-WI | Chikara | Chwastox | Fundamentum | Logo | Mustang | Roundup | LSD0.05 |
|---|---|---|---|---|---|---|---|---|---|---|
| LP | Mean | 93.72a | 85.88b | 69.84e | 73.92de | 79.35cd | 82.52bc | 60.52f | 79.96bcd | 6.473 |
| Min-Max | 52–127 | 69–113 | 53–97 | 55–95 | 60–120 | 54–110 | 37–100 | 50–111 | ||
| s.d. | 24.34 | 9.55 | 10.88 | 10.33 | 15.19 | 15.49 | 12.09 | 13.7 | ||
| cv | 25.97 | 11.12 | 15.58 | 13.97 | 19.14 | 18.77 | 19.98 | 17.13 | ||
| FD | Mean | 8.896a | 8.896a | 8.992a | 8.944a | 8.88a | 8.816a | 8.816a | 8.88a | 0.316 |
| Min-Max | 7–10 | 7–10 | 8–11 | 8–11 | 8–10 | 7–10 | 7–10 | 7–11 | ||
| s.d. | 0.551 | 0.6699 | 0.7459 | 0.7218 | 0.7559 | 0.6523 | 0.7 | 0.8762 | ||
| cv | 6.19 | 7.53 | 8.30 | 8.07 | 8.51 | 7.40 | 7.94 | 9.87 | ||
| LFP | Mean | 5.672ab | 5.952a | 5.08cd | 5.424bc | 5.92a | 4.944d | 4.952d | 5.864ab | 0.443 |
| Min-Max | 4–9 | 4–9 | 4–8 | 4–8 | 3–9 | 3–7 | 4–7 | 4–8 | ||
| s.d. | 1.0532 | 1.2369 | 0.8481 | 0.8256 | 1.2608 | 0.8063 | 0.6458 | 1.1937 | ||
| cv | 18.57 | 20.78 | 16.69 | 15.22 | 21.30 | 16.31 | 13.04 | 20.36 | ||
| FN | Mean | 32.32a | 31.96ab | 25.48c | 24.28c | 24.5c | 29.44b | 21.4d | 26.48c | 2.627 |
| Min-Max | 20–48 | 24–45 | 16–39 | 11–30 | 15–32 | 12–38 | 12–29 | 14–42 | ||
| s.d. | 7.463 | 5.052 | 5.253 | 4.302 | 4.014 | 7.498 | 5.205 | 7.14 | ||
| cv | 23.09 | 15.81 | 20.62 | 17.72 | 16.38 | 25.47 | 24.32 | 26.96 | ||
| LP/LFP | Mean | 16.92a | 15.16b | 14.1b | 13.84bc | 13.81bc | 16.94a | 12.44c | 14.12b | 1.535 |
| Min-Max | 8–25.4 | 8.11–28.25 | 7.86–24.25 | 9.17–20 | 7.22–23.33 | 10.57–26.25 | 7.4–20 | 7.14–25 | ||
| s.d. | 4.899 | 4.008 | 3.116 | 2.292 | 2.921 | 3.248 | 3.035 | 3.484 | ||
| cv | 28.95 | 26.44 | 22.10 | 16.56 | 21.15 | 19.17 | 24.40 | 24.67 | ||
| FD/LFP | Mean | 1.616b | 1.559b | 1.811a | 1.688ab | 1.579b | 1.83a | 1.809a | 1.598b | 0.149 |
| Min-Max | 0.89–2.5 | 0.89–2.5 | 1.13–2.5 | 1.13–2.5 | 0.89–3.33 | 1.14–3 | 1.33–2.5 | 0.88–2.75 | ||
| s.d. | 0.2834 | 0.339 | 0.2948 | 0.2985 | 0.4115 | 0.3214 | 0.2709 | 0.4479 | ||
| cv | 17.54 | 21.74 | 16.28 | 17.68 | 26.06 | 17.56 | 14.98 | 28.03 | ||
| LP/FN | Mean | 2.935bc | 2.729c | 2.798c | 3.131ab | 3.35a | 2.953bc | 2.953bc | 3.155ab | 0.289 |
| Min-Max | 2.08–4.85 | 1.9–3.6 | 2.2–4.19 | 2.2–5.45 | 2.19–5 | 1.81–4.57 | 1.96–4.33 | 2–4.83 | ||
| s.d. | 0.693 | 0.3661 | 0.4322 | 0.6498 | 0.9095 | 0.6959 | 0.7219 | 0.6582 | ||
| cv | 23.61 | 13.42 | 15.45 | 20.75 | 27.15 | 23.57 | 24.45 | 20.86 |
| Feature | First Canonical Variable | Second Canonical Variable |
|---|---|---|
| LP | 0.977 *** | 0.036 |
| FD | −0.007 | 0.114 |
| LFP | 0.616 | 0.743 * |
| FN | 0.941 *** | −0.312 |
| LP/LFP | 0.811 * | −0.471 |
| FD/LFP | −0.62 | −0.724 * |
| LP/FN | −0.16 | 0.768 * |
| Percentage of explained multivariate variability | 67.70% | 16.53% |
| Feature | First Canonical Variable | Second Canonical Variable |
|---|---|---|
| The length of the inflorescence peduncle (LP) | 0.998 *** | −0.044 |
| Flower diameter (FD) | −0.081 | −0.364 * |
| The length of the flower peduncle (LFP) | 0.268 | 0.354 * |
| The number of flowers in inflorescences (FN) | 0.611 *** | −0.761 *** |
| LP/LFP | 0.736 *** | −0.282 |
| FD/LFP | −0.280 | −0.385 * |
| LP/FN | 0.256 | 0.899 *** |
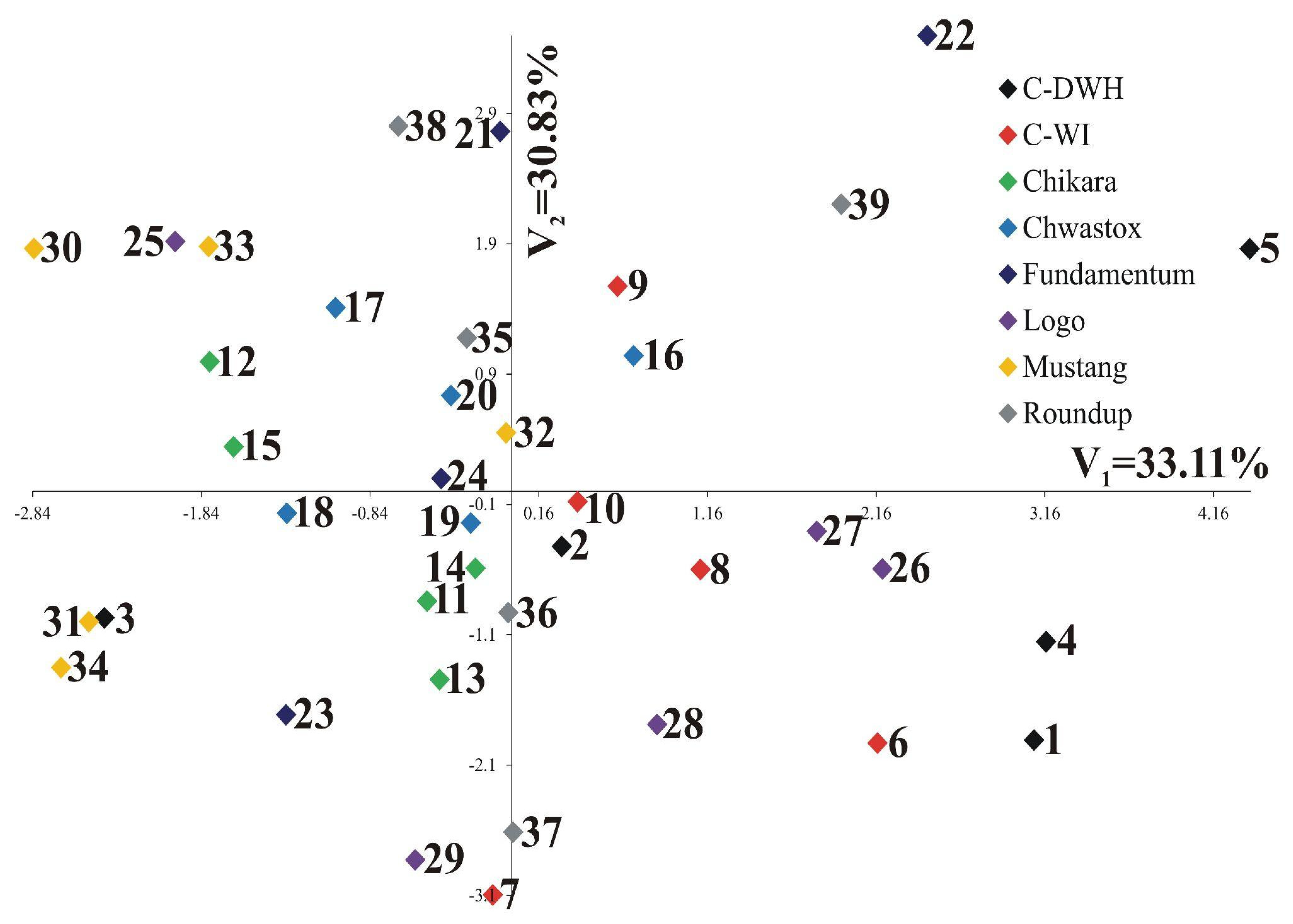
| Percentage of explained multivariate variability | 33.11 | 30.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wrońska-Pilarek, D.; Maciejewska-Rutkowska, I.; Bocianowski, J.; Korzeniewicz, R.; Lechowicz, K.; Hauke-Kowalska, M. Does the Reaction of Inflorescences and Flowers of the Invasive Prunus serotina Ehrh. to Various Herbicides Give Hope for Elimination of This Species from Polish Forests? Forests 2022, 13, 21. https://doi.org/10.3390/f13010021
Wrońska-Pilarek D, Maciejewska-Rutkowska I, Bocianowski J, Korzeniewicz R, Lechowicz K, Hauke-Kowalska M. Does the Reaction of Inflorescences and Flowers of the Invasive Prunus serotina Ehrh. to Various Herbicides Give Hope for Elimination of This Species from Polish Forests? Forests. 2022; 13(1):21. https://doi.org/10.3390/f13010021
Chicago/Turabian StyleWrońska-Pilarek, Dorota, Irmina Maciejewska-Rutkowska, Jan Bocianowski, Robert Korzeniewicz, Kacper Lechowicz, and Maria Hauke-Kowalska. 2022. "Does the Reaction of Inflorescences and Flowers of the Invasive Prunus serotina Ehrh. to Various Herbicides Give Hope for Elimination of This Species from Polish Forests?" Forests 13, no. 1: 21. https://doi.org/10.3390/f13010021
APA StyleWrońska-Pilarek, D., Maciejewska-Rutkowska, I., Bocianowski, J., Korzeniewicz, R., Lechowicz, K., & Hauke-Kowalska, M. (2022). Does the Reaction of Inflorescences and Flowers of the Invasive Prunus serotina Ehrh. to Various Herbicides Give Hope for Elimination of This Species from Polish Forests? Forests, 13(1), 21. https://doi.org/10.3390/f13010021