
Ground Cover—Biomass Functions for Early-Seral Vegetation



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites Description
2.2. Study Design
2.3. Model Description
2.4. Model Fitting and Evaluation
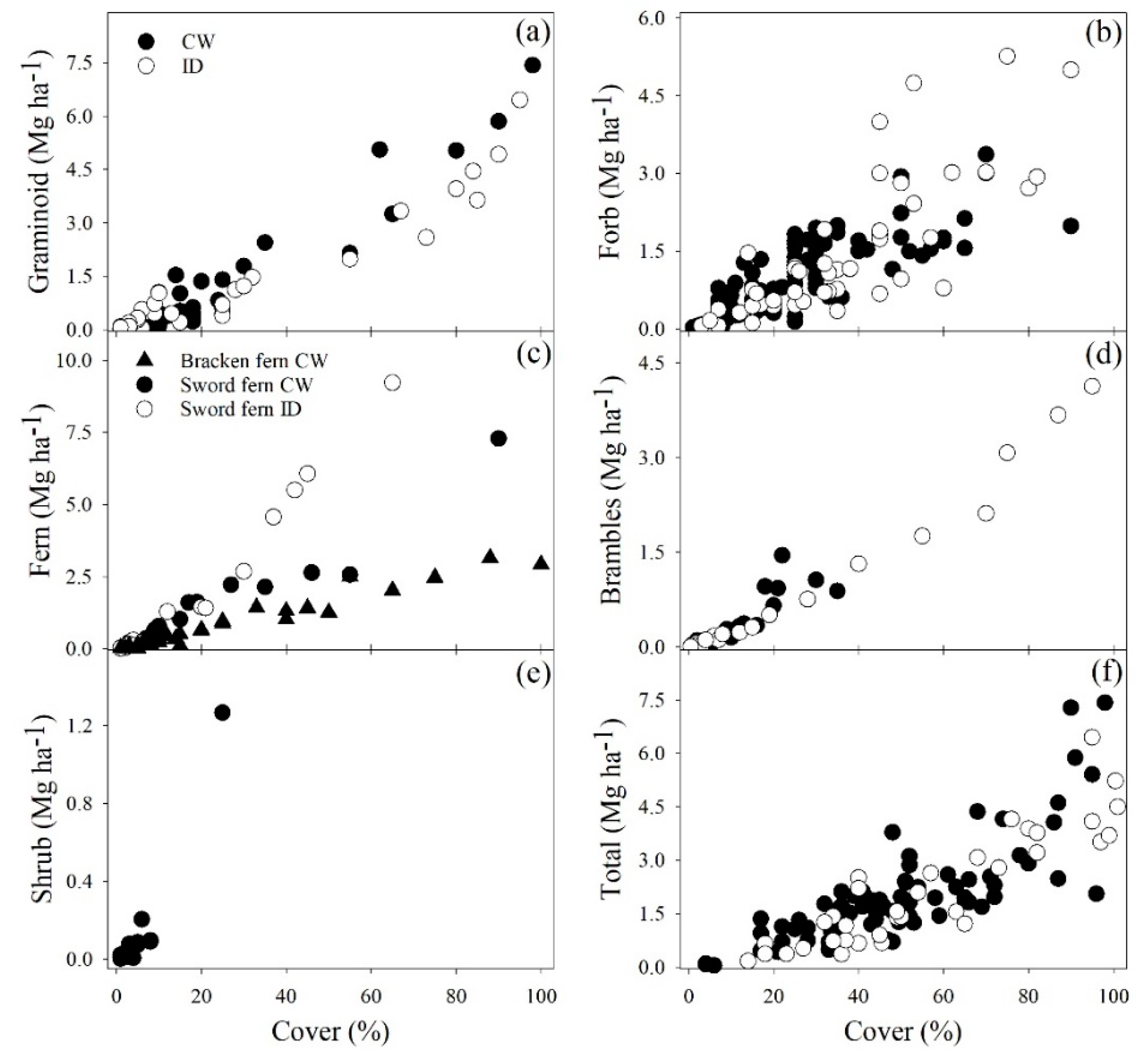
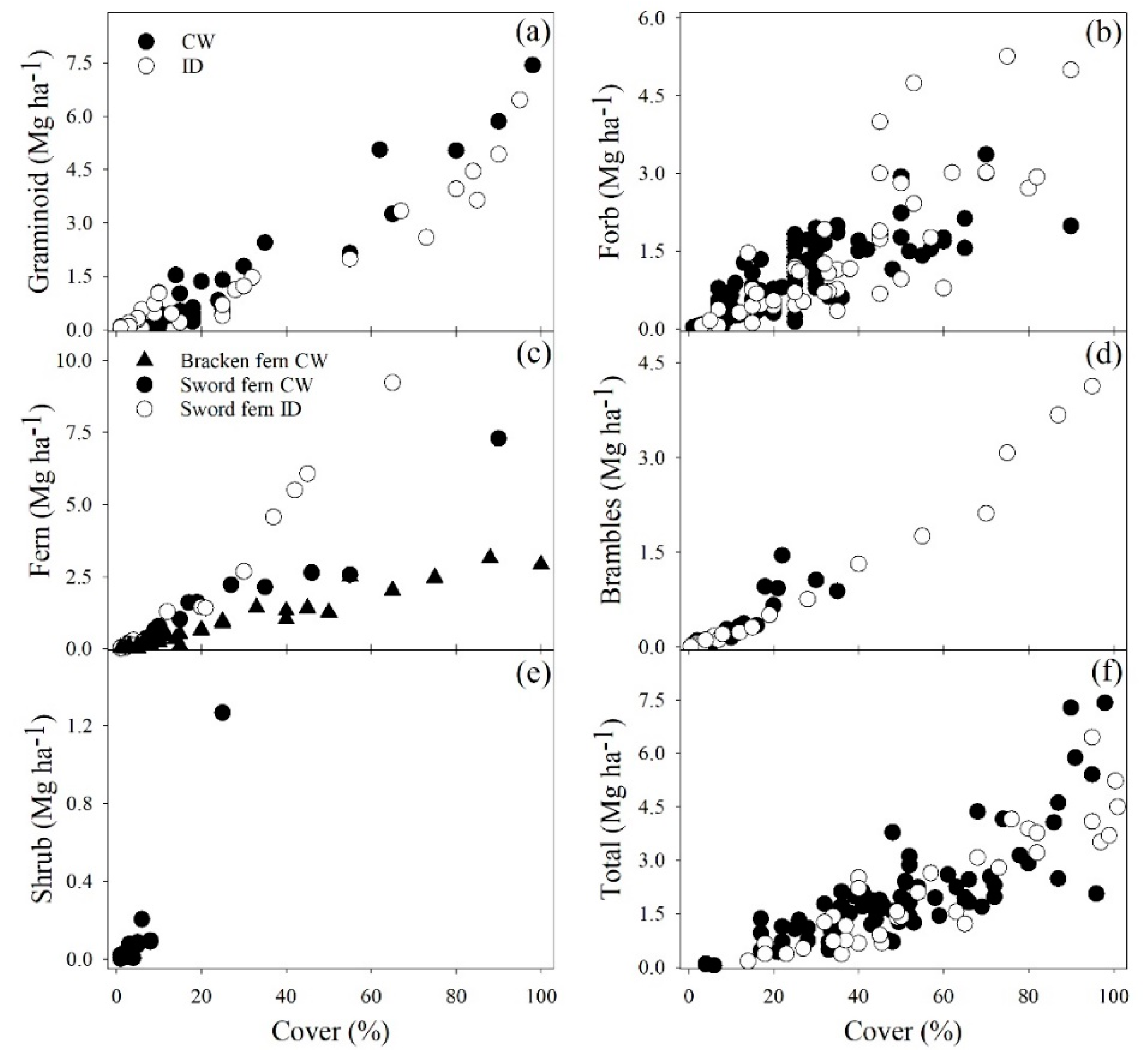
3. Results
3.1. Model Selection
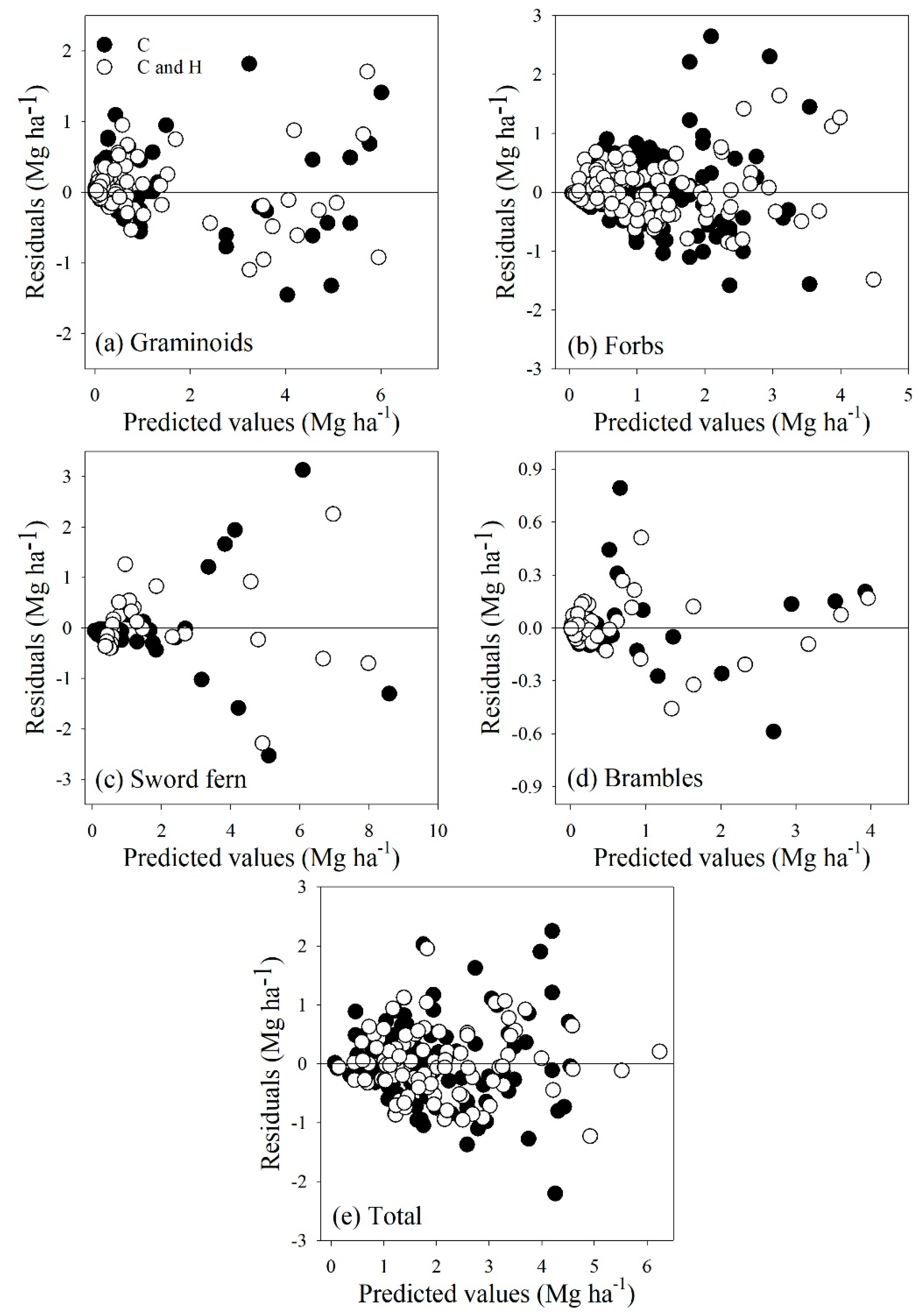
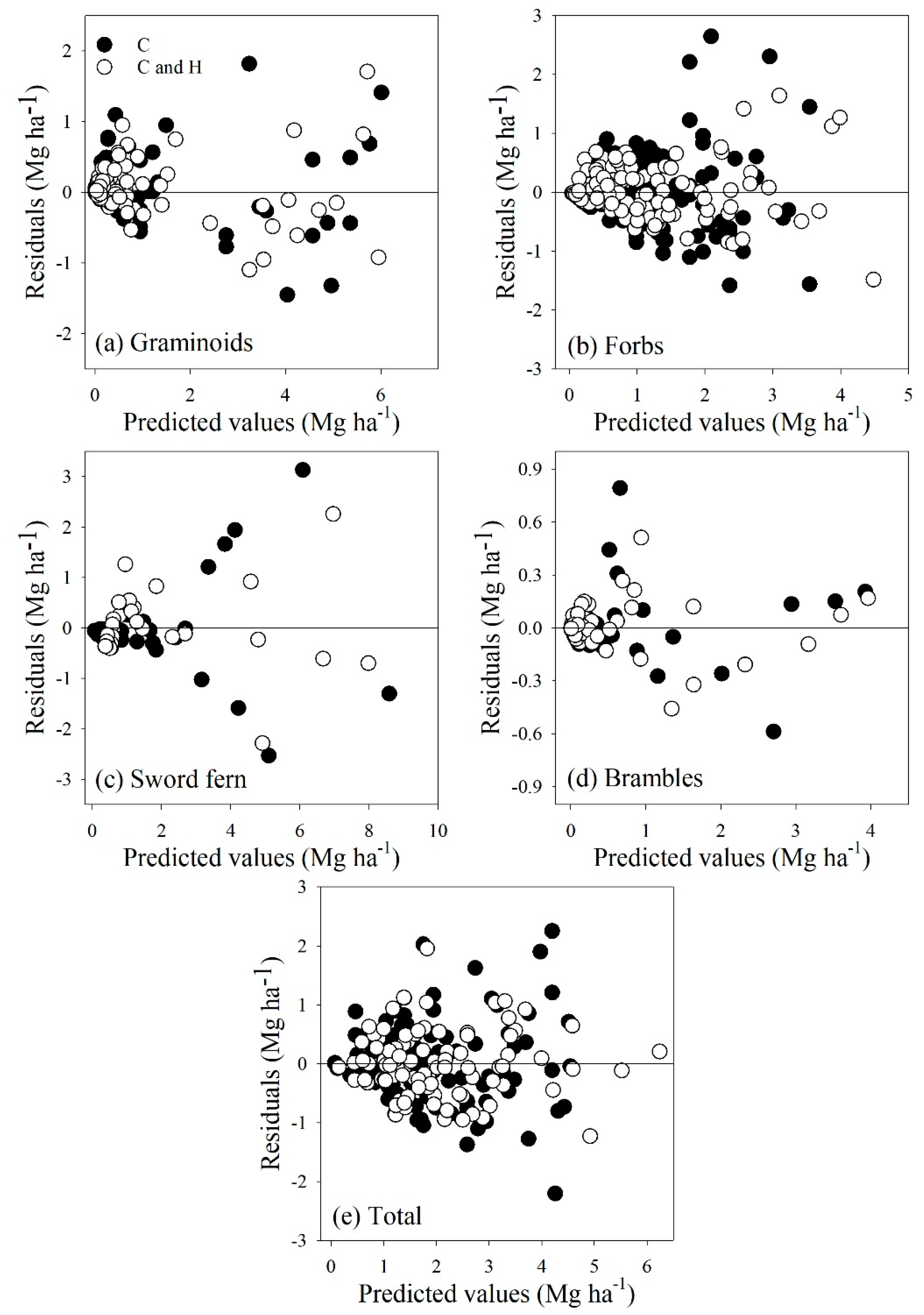
3.2. Model Fitting
3.3. Model Evaluation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ji, L.; Wylie, B.K.; Nossov, D.R.; Peterson, B.; Waldrop, M.P.; McFarland, J.W.; Rover, J.; Hollingsworth, T.N. Estimating aboveground biomass in interior Alaska with Landsat data and field measurements. Int. J. Appl. Earth Obs. Geoinf. 2012, 18, 451–461. [Google Scholar] [CrossRef]
- Gonzalez-Benecke, C.; Samuelson, L.; Stokes, T.; Cropper, W.; Martin, T.; Johnsen, K. Understory plant biomass dynamics of prescribed burned Pinus palustris stands. For. Ecol. Manag. 2015, 344, 84–94. [Google Scholar] [CrossRef]
- García, M.; Riaño, D.; Chuvieco, E.; Danson, F. Estimating biomass carbon stocks for a Mediterranean forest in central Spain using LiDAR height and intensity data. Remote Sens. Environ. 2010, 114, 816–830. [Google Scholar] [CrossRef]
- Eisfelder, C.; Klein, I.; Bekkuliyeva, A.; Kuenzer, C.; Buchroithner, M.F.; Dech, S. Above-ground biomass estimation based on NPP time-series—A novel approach for biomass estimation in semi-arid Kazakhstan. Ecol. Indic. 2017, 72, 13–22. [Google Scholar] [CrossRef]
- Gonzalez-Benecke, C.A.; Gezan, S.A.; Albaugh, T.J.; Allen, H.; Burkhart, H.E.; Fox, T.R.; Jokela, E.J.; Maier, C.A.; Martin, T.A.; Rubilar, R.; et al. Local and general above-stump biomass functions for loblolly pine and slash pine trees. For. Ecol. Manag. 2014, 334, 254–276. [Google Scholar] [CrossRef]
- Yan, P.; Xiao, C.; Xu, L.; Yu, G.; Li, A.; Piao, S.; He, N. Biomass EeTerrestrial Eco-systems: Insights into the Nation’s Sustainable Energy Supply. Renew. Sustain. Energy Rev. 2020, 127, 1–10. [Google Scholar] [CrossRef]
- Garroutte, E.L.; Hansen, A.J.; Lawrence, R.L. Using NDVI and EVI to Map Spa-tiotemporal Variation in the Biomass and Quality of Forage for Migratory Elk in the Greater Yellowstone Ecosystem. Remote Sens. 2016, 8, 404. [Google Scholar]
- Moreau, S.; Toan, T.L. Biomass quantification of Andean wetland Forages Using ERS Satellite SAR Data for Optimizing Livestock Management. Remote Sens. Environ. 2003, 84, 477–492. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies and Vegetation Processes; John Wiley and Sons: Chichester, UK, 1979. [Google Scholar]
- Catchpole, W.R.; Wheeler, C.J. Estimating plant biomass: A review of techniques. Austral Ecol. 1992, 17, 121–131. [Google Scholar] [CrossRef]
- Jonasson, S. Evaluation of the Point Intercept Method for the Estimation of Plant Biomass. Oikos 1988, 52, 101. [Google Scholar] [CrossRef]
- Somogyi, Z.; Cienciala, E.; Mäkipää, R.; Muukkonen, P.; Lehtonen, A.; Weiss, P. Indirect methods of large-scale forest biomass estimation. Eur. J. For. Res. 2006, 126, 197–207. [Google Scholar] [CrossRef]
- Dubayah, R.O.; Sheldon, S.L.; Clark, D.B.; Hofton, M.A.; Blair, J.B.; Hurtt, G.C.; Chazdon, R. Estimation of tropical forest height and biomass dynamics using lidar remote sensing at La Selva, Costa Rica. J. Geophys. Res. Space Phys. 2010, 115. [Google Scholar] [CrossRef]
- Kronseder, K.; Ballhorn, U.; Böhm, V.; Siegert, F. Above ground biomass estimation across forest types at different degradation levels in Central Kalimantan using LiDAR data. Int. J. Appl. Earth Obs. Geoinf. 2012, 18, 37–48. [Google Scholar] [CrossRef]
- Thenkabail, P.S.; Stucky, N.; Griscom, B.W.; Ashton, M.S.; Diels, J.; Van Der Meer, B.; Enclona, E. Biomass estimations and carbon stock calculations in the oil palm plantations of African derived savannas using IKONOS data. Int. J. Remote Sens. 2004, 25, 5447–5472. [Google Scholar] [CrossRef]
- Bendig, J.; Yu, K.; Aasen, H.; Bolten, A.; Bennertz, S.; Broscheit, J.; Gnyp, M.L.; Bareth, G. Combining UAV-based plant height from crop surface models, visible, and near infrared vegetation indices for biomass monitoring in barley. Int. J. Appl. Earth Obs. Geoinf. 2015, 39, 79–87. [Google Scholar] [CrossRef]
- Eisfelder, C.; Kuenzer, C.; Dech, S. Derivation of biomass information for semi-arid areas using remote-sensing data. Int. J. Remote Sens. 2011, 33, 2937–2984. [Google Scholar] [CrossRef]
- Yarie, J. The Role of Understory Vegetation in the Nutrient Cycle of Forest-ed Ecosystems in the Mountain Hemlock Biogeoclimatic Zone. Ecology 1980, 61, 1498–1514. [Google Scholar] [CrossRef] [Green Version]
- Rottgermann, M.; Steinlein, T.; Beyschlag, W.; Dietz, H. Linear relationships between aboveground biomass and plant cover in low open herbaceous vegetation. J. Veg. Sci. 2000, 11, 145–148. [Google Scholar] [CrossRef]
- Guevara, J.; Gonnet, J.; Estevez, O. Biomass estimation for native perennial grasses in the plain of Mendoza, Argentina. J. Arid. Environ. 2002, 50, 613–619. [Google Scholar] [CrossRef]
- Heinrichs, S.; Bernhardt-Römermann, M.; Schmidt, W. The estimation of aboveground biomass and nutrient pools of understorey plants in closed Norway spruce forests and on clearcuts. Eur. J. For. Res. 2010, 129, 613–624. [Google Scholar] [CrossRef] [Green Version]
- Hermy, M. Accuracy of Visual Cover Assessments in Predicting Standing Crop and Environmental Correlation in Deciduous Forests. Vegetatio 1988, 75, 57–64. [Google Scholar] [CrossRef]
- Chiarucci, A.; Wilson, J.B.; Anderson, B.J.; De Dominicis, V. Cover versus biomass as an estimate of species abundance: Does it make a difference to the conclusions? J. Veg. Sci. 1999, 10, 35–42. [Google Scholar] [CrossRef]
- Muukkonen, P.; Mäkipää, R.; Laiho, R.; Minkkinen, K.; Vasander, H.; Finér, L. Relationship between Biomass and Percentage Cover in Understorey Vegeta-tion of Boreal Coniferous Forests. Silva Fenn. 2006, 40, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Gilliam, F.S.; Turrill, N.L. Herbaceous Layer Cover and Biomass in a Young Versus a Mature Stand of a Central Appalachian Hardwood Forest. Bull. Torrey Bot. Club 1993, 120, 445. [Google Scholar] [CrossRef]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley and Sons: New York, NY, USA, 1974. [Google Scholar]
- Axmanová, I.; Tichý, L.; Fajmonová, Z.; Hájková, P.; Hettenbergerová, E.; Li, C.-F.; Merunková, K.; Nejezchlebová, M.; Otýpková, Z.; Vymazalová, M.; et al. Estimation of herbaceous biomass from species composition and cover. Appl. Veg. Sci. 2012, 15, 580–589. [Google Scholar] [CrossRef]
- Ferrari, L. Relationships between Plant Biomass and Cover in the Ground Cover Layer of Longleaf Pine Forests at Fort Benning, GA. Master’s Thesis, Auburn University, Auburn, AL, USA, 14 December 2013. [Google Scholar]
- Flombaum, P.; Sala, O. A non-destructive and rapid method to estimate biomass and aboveground net primary production in arid environments. J. Arid. Environ. 2007, 69, 352–358. [Google Scholar] [CrossRef]
- Johnson, P.S.; Johnson, C.L.; West, N.E. Estimation of Phytomass for Ungrazed Crested Wheatgrass Plants Using Allometric Equations. J. Range Manag. 1988, 41, 421. [Google Scholar] [CrossRef]
- Guevara, C.A.; Gonzalez-Benecke, C.A.; Wightman, M.G. Competition and Site Interactions Experiment Tier I.; Western Hemlock and Douglas-Fir (Vegeta-tion Dynamics, Soil Moisture, Seedling Growth, and Survival). Veg. Manag. Res. Coop. Annu. Rep. 2019, 1–21. [Google Scholar]
- Web Soil Survey, Soil Survey Staff. Web Soil Survey: Soil Data Mart. USDA-NCRS. Available online: https://websoilsurvey.sc.egov.usda.gov/ (accessed on 20 October 2018).
- Wang, T.; Hamann, A.; Spittlehouse, D.L.; Murdock, T.Q. ClimateWNA—High-Resolution Spatial Climate Data for Western North America. J. Appl. Meteorol. Clim. 2012, 51, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Samuelson, L.J.; Stokes, T.A. Developing Tools for Ecological Forestry and Carbon Management in Longleaf Pine. In Field Methods and Protocols; Auburn University: Auburn, AL, USA, 2011. [Google Scholar]
- Seber, G.A.; Wild, C.J. Nonlinear Regression; Wiley: Hoboken, NJ, USA, 2003. [Google Scholar]
- Myers, R.H. Classical and Modern Regression with Applications, 2nd ed.; Duxbury: Pacific Grove, CA, USA, 2000. [Google Scholar]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Neter, J.; Kutner, M.H.; Nachtsheim, C.J.; Wasserman, W. Applied Linear Statistical Models, 4th ed.; Irwin: Toronto, ON, Canada, 1996. [Google Scholar]
- Paul, K.I.; Roxburgh, S.H.; Chave, J.; England, J.; Zerihun, A.; Specht, A.; Lewis, T.; Bennett, L.; Baker, T.G.; Adams, M.; et al. Testing the generality of above-ground biomass allometry across plant functional types at the continent scale. Glob. Chang. Biol. 2016, 22, 2106–2124. [Google Scholar] [CrossRef] [PubMed]
- Paul, K.I.; Larmour, J.; Specht, A.; Zerihun, A.; Ritson, P.; Roxburgh, S.H.; Sochacki, S.; Lewis, T.; Barton, C.V.; England, J.R.; et al. Testing the generality of below-ground biomass allometry across plant functional types. For. Ecol. Manag. 2018, 432, 102–114. [Google Scholar] [CrossRef]
- Blujdea, V.; Pilli, R.; Dutca, I.; Ciuvat, L.; Abrudan, I. Allometric biomass equations for young broadleaved trees in plantations in Romania. For. Ecol. Manag. 2012, 264, 172–184. [Google Scholar] [CrossRef]
- Landuyt, D.; Maes, S.L.; Depauw, L.; Ampoorter, E.; Blondeel, H.; Perring, M.; Brūmelis, G.; Brunet, J.; Decocq, G.; Ouden, J.D.; et al. Drivers of above-ground understorey biomass and nutrient stocks in temperate deciduous forests. J. Ecol. 2019, 108, 982–997. [Google Scholar] [CrossRef]
- Goldberg, D.E. Simplifying the Study of Competition at the Individual Plant Level: Consequences of Distinguishing between Competitive Effect and Response for Forest Vegetation Management. N. Z. J. For. Sci. 1996, 26, 19–38. [Google Scholar]

{kind=link}
{kind=link}
| CW | ID | |||
|---|---|---|---|---|
| 2017 | 2018 | 2018 | 2019 | |
| Rainy days | 27 | 39 | 6 | 14 |
| Total Rain (mm) | 53.8 | 29.9 | 37.0 | 39.3 |
| T max (°C) | 19.1 | 19.1 | 27.2 | 26.3 |
| T mean (°C) | 14.6 | 14.6 | 18.4 | 18.2 |
| RH (%) | 74.7 | 81.1 | 54.8 | 57.3 |
| VPD max (kPa) | 0.95 | 0.80 | 2.60 | 2.31 |
| VPD mean (kPa) | 0.56 | 0.39 | 1.42 | 1.28 |
| Site | Vegetation Type | Cover (%) | Height (cm) | Biomass (Mg ha−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | Mean | Min | Max | n | Mean | Min | Max | Mean | Min | Max | ||
| CW | Bracken fern | 32 | 26.8 | 1 | 100 | 19 | 49.6 | 12 | 100 | 0.8 | 0.01 | 3.1 |
| Sword fern | 18 | 20.4 | 2 | 90 | 18 | 39.3 | 20 | 72 | 1.4 | 0.04 | 7.3 | |
| Forbs | 90 | 27.4 | 1 | 90 | 65 | 32.5 | 5 | 150 | 1.1 | 0.03 | 3.4 | |
| Graminoids | 76 | 14.1 | 1 | 98 | 54 | 28.8 | 5 | 148 | 0.7 | 0.002 | 7.4 | |
| Brambles | 47 | 7.4 | 1 | 35 | 36 | 17.9 | 4 | 60 | 0.2 | 0.001 | 1.5 | |
| Shrubs | 18 | 4.6 | 1 | 25 | 16 | 23.1 | 9 | 50 | 0.1 | 0.002 | 1.3 | |
| ID | Sword fern | 19 | 16.7 | 1 | 65 | 19 | 37.6 | 5 | 68 | 1.8 | 0.01 | 9.2 |
| Forbs | 47 | 35.6 | 3 | 90 | 47 | 28.6 | 5 | 80 | 1.4 | 0.04 | 5.3 | |
| Graminoids | 29 | 30.7 | 1 | 95 | 29 | 39.9 | 7 | 86 | 1.4 | 0.02 | 6.5 | |
| Brambles | 30 | 18.2 | 1 | 95 | 30 | 21.5 | 5 | 50 | 0.6 | 0.001 | 4.1 | |
| Pooled | Sword fern | 37 | 18.5 | 1 | 90 | 37 | 38.4 | 5 | 72 | 1.6 | 0.01 | 9.2 |
| Forbs | 137 | 30.2 | 1 | 90 | 112 | 30.9 | 5 | 150 | 1.2 | 0.03 | 5.3 | |
| Graminoids | 105 | 18.7 | 1 | 98 | 83 | 32.7 | 5 | 148 | 0.9 | 0.002 | 7.4 | |
| Brambles | 77 | 11.6 | 1 | 95 | 66 | 19.5 | 4 | 60 | 0.4 | 0.001 | 4.1 | |
| Site | Growth Habit | Model | Parameter | Parameter Estimate | SE | R2 | RMSE | CV |
|---|---|---|---|---|---|---|---|---|
| CW | Bracken fern | a | 0.030351 | 0.007426 | 0.972 | 0.21 | 24.4 | |
| b | 1.016627 | 0.058688 | ||||||
| Sword fern | a | 0.039499 | 0.017128 | 0.958 | 0.44 | 32.3 | ||
| b | 1.139870 | 0.104380 | ||||||
| Forbs | a | 0.093137 | 0.025081 | 0.881 | 0.44 | 41.4 | ||
| b | 0.751169 | 0.072756 | ||||||
| Graminoids | a | 0.013234 | 0.003293 | 0.954 | 0.33 | 48.3 | ||
| b | 1.366124 | 0.057796 | ||||||
| Brambles | a | 1.095629 | 0.066066 | 0.924 | 0.11 | 51.2 | ||
| b | 70.709855 | 30.877161 | ||||||
| c | 0.285186 | 0.032782 | ||||||
| Shrubs | a | 0.003051 | 0.001236 | 0.983 | 0.04 | 35.0 | ||
| b | 1.872408 | 0.127451 | ||||||
| Total | a | 0.024005 | 0.012096 | 0.896 | 0.70 | 37.5 | ||
| b | 1.119684 | 0.121707 | ||||||
| ID | Sword fern | a | 9.965688 | 0.373641 | 0.994 | 0.23 | 12.8 | |
| b | 45.279774 | 6.867032 | ||||||
| c | 0.096193 | 0.005173 | ||||||
| Forbs | a | 0.040084 | 0.012863 | 0.838 | 0.62 | 52.0 | ||
| b | 0.996012 | 0.082203 | ||||||
| Graminoids | a | 663,843,166 | 7.28 × 1012 | 0.972 | 0.38 | 26.2 | ||
| b | 1,932,462,172 | 2.12 × 1013 | ||||||
| c | 0.030093 | 0.001911 | ||||||
| Brambles | a | 0.015168 | 0.002712 | 0.971 | 0.15 | 39.7 | ||
| b | 1.220071 | 0.042044 | ||||||
| Total | a | 0.012402 | 0.004927 | 0.908 | 0.70 | 36.2 | ||
| b | 1.279081 | 0.093410 | ||||||
| Pooled | Sword fern | a | 0.023319 | 0.005501 | 0.992 | 0.29 | 15.8 | |
| b | 1.440794 | 0.060425 | ||||||
| Forbs | a | 0.014729 | 0.010594 | 0.829 | 0.81 | 56.2 | ||
| b | 1.266369 | 0.175836 | ||||||
| Graminoids | a | 0.005639 | 0.004692 | 0.960 | 0.45 | 31.2 | ||
| b | 1.502171 | 0.189706 | ||||||
| Brambles | a | 0.005966 | 0.001728 | 0.992 | 0.11 | 18.4 | ||
| b | 1.432098 | 0.066262 | ||||||
| Total | a | 0.004544 | 0.003300 | 0.931 | 0.70 | 32.6 | ||
| b | 1.509432 | 0.164803 |
| Site | Growth Habit | Model | Parameter | Parameter Estimate | SE | R2 | RMSE | CV |
|---|---|---|---|---|---|---|---|---|
| CW | Bracken fern | a | 0.050765 | 0.030871 | 0.971 | 0.26 | 31.0 | |
| b | 1.141709 | 0.140055 | ||||||
| c | −0.250187 | 0.202383 | ||||||
| Sword fern | a | 0.013492 | 0.005656 | 0.979 | 0.32 | 23.1 | ||
| b | 0.907455 | 0.078963 | ||||||
| c | 0.511214 | 0.137416 | ||||||
| Forb | a | 0.043841 | 0.012741 | 0.920 | 0.36 | 33.8 | ||
| b | 0.765630 | 0.073362 | ||||||
| c | 0.224945 | 0.050597 | ||||||
| Graminoid | a | 0.012240 | 0.003973 | 0.945 | 0.38 | 56.3 | ||
| b | 1.299804 | 0.095254 | ||||||
| c | 0.085844 | 0.07546 | ||||||
| Bramble | a | 1.132271 | 0.078635 | 0.923 | 0.12 | 33.0 | ||
| b | 15.110096 | 4.215749 | ||||||
| c | 0.520423 | 0.07042 | ||||||
| Shrub | a | 0.000120887 | 0.000045629 | 0.994 | 0.01 | 10.8 | ||
| b | 1.255475 | 0.038010 | ||||||
| c | 1.333292 | 0.111233 | ||||||
| Total | a | 0.029662 | 0.012457 | 0.941 | 0.55 | 29.3 | ||
| b | 0.949564 | 0.110260 | ||||||
| c | 0.241096 | 0.054032 | ||||||
| ID | Sword fern | a | 0.004689 | 0.002191 | 0.993 | 0.21 | 11.7 | |
| b | 1.436381 | 0.048306 | ||||||
| c | 0.407966 | 0.105510 | ||||||
| Forb | a | 0.001469 | 0.000615 | 0.971 | 0.34 | 23.2 | ||
| b | 1.021933 | 0.076051 | ||||||
| c | 0.918408 | 0.072488 | ||||||
| Graminoid | a | 0.001849 | 0.000762 | 0.989 | 0.24 | 16.5 | ||
| b | 0.906781 | 0.064931 | ||||||
| c | 0.919863 | 0.107180 | ||||||
| Bramble | a | 0.002623 | 0.000539 | 0.994 | 0.07 | 10.7 | ||
| b | 1.278391 | 0.038965 | ||||||
| c | 0.400863 | 0.054239 | ||||||
| Total | a | 0.006148 | 0.003314 | 0.958 | 0.55 | 25.6 | ||
| b | 1.307160 | 0.129175 | ||||||
| c | 0.225179 | 0.045990 | ||||||
| Pooled | Sword fern | a | 7.990414 | 0.588840 | 0.932 | 0.71 | 44.0 | |
| b | 20.531989 | 5.062166 | ||||||
| c | 0.151918 | 0.015650 | ||||||
| Forb | a | 0.011322 | 0.003521 | 0.911 | 0.48 | 39.8 | ||
| b | 0.958610 | 0.067395 | ||||||
| c | 0.418276 | 0.049445 | ||||||
| Graminoid | a | 0.008294 | 0.002681 | 0.954 | 0.40 | 44.9 | ||
| b | 1.098820 | 0.075305 | ||||||
| c | 0.352346 | 0.057777 | ||||||
| Bramble | a | 0.004285 | 0.001250 | 0.982 | 0.12 | 33.5 | ||
| b | 1.082031 | 0.038280 | ||||||
| c | 0.499640 | 0.081455 | ||||||
| Total | a | 0.016582 | 0.005320 | 0.946 | 0.56 | 28.9 | ||
| b | 1.089033 | 0.081624 | ||||||
| c | 0.225714 | 0.035750 |
| Site | Growth Habit | Model | Explanatory Variable (s) | RMSE Bias | |||||
|---|---|---|---|---|---|---|---|---|---|
| CW | Bracken fern | Cover | 0.85 | 0.87 | 0.15 | (17.3) | −0.02 | (−1.9) | |
| Cover and Height | 1.17 | 1.18 | 0.21 | (18.1) | −0.01 | (−0.9) | |||
| Sword fern | Cover | 1.37 | 1.33 | 0.34 | (24.9) | 0.05 | (3.4) | ||
| Cover and Height | 1.37 | 1.38 | 0.26 | (18.9) | −0.01 | (−0.6) | |||
| Forbs | Cover | 1.07 | 1.07 | 0.42 | (39.0) | −0.01 | (−0.7) | ||
| Cover and Height | 1.05 | 1.05 | 0.33 | (31.8) | 0.00 | (0.0) | |||
| Graminoids | Cover | 0.68 | 0.67 | 0.25 | (36.5) | 0.01 | (1.4) | ||
| Cover and Height | 0.83 | 0.81 | 0.27 | (32.1) | 0.02 | (2.7) | |||
| Brambles | Cover | 0.21 | 0.20 | 0.28 | (133.5) | 0.00 | (2.1) | ||
| C∙H | 0.25 | 0.26 | 0.11 | (44.5) | 0.00 | (−1.5) | |||
| Shrubs | Cover | 0.11 | 0.11 | 0.03 | (23.8) | 0.01 | (5.4) | ||
| Cover and Height | 0.13 | 0.12 | 0.01 | (7.5) | 0.00 | (2.9) | |||
| Total | Cover | 1.86 | 1.85 | 0.66 | (35.3) | 0.01 | (0.5) | ||
| Cover and Height | 1.98 | 1.98 | 0.46 | (23.3) | 0.00 | (0.2) | |||
| ID | Sword fern | Cover | 1.82 | 1.87 | 1.34 | (73.5) | −0.05 | (−2.6) | |
| Cover and Height | 1.82 | 1.81 | 0.15 | (8.0) | 0.01 | (0.7) | |||
| Forbs | Cover | 1.45 | 1.44 | 0.78 | (53.6) | 0.00 | (0.3) | ||
| Cover and Height | 1.45 | 1.41 | 0.31 | (21.4) | 0.04 | (2.7) | |||
| Graminoids | Cover | 1.44 | 1.45 | 0.32 | (22.2) | −0.02 | (−1.3) | ||
| Cover and Height | 1.44 | 1.40 | 0.21 | (14.3) | 0.04 | (2.8) | |||
| Brambles | Cover | 0.63 | 0.61 | 0.09 | (14.2) | 0.01 | (2.4) | ||
| Cover and Height | 0.63 | 0.61 | 0.05 | (8.6) | 0.01 | (1.8) | |||
| Total | Cover | 2.14 | 2.14 | 0.58 | (27.2) | 0.00 | (0.1) | ||
| Cover and Height | 2.14 | 2.15 | 1.37 | (63.8) | −0.01 | (−0.5) | |||
| Pooled | Sword fern | Cover | 1.60 | 1.67 | 0.73 | (45.2) | −0.06 | (−3.9) | |
| C∙H | 1.60 | 1.65 | 0.54 | (33.7) | −0.04 | (−2.6) | |||
| Forbs | Cover | 1.20 | 1.19 | 0.58 | (48.5) | 0.00 | (0.3) | ||
| Cover and Height | 1.22 | 1.20 | 0.45 | (37.2) | 0.02 | (1.5) | |||
| Graminoids | Cover | 0.89 | 0.85 | 0.39 | (43.9) | 0.04 | (4.1) | ||
| Cover and Height | 1.04 | 1.00 | 0.35 | (33.6) | 0.04 | (3.7) | |||
| Brambles | Cover | 0.37 | 0.37 | 0.10 | (27.0) | 0.00 | (−0.8) | ||
| Cover and Height | 0.42 | 0.42 | 0.10 | (23.8) | 0.00 | (1.0) | |||
| Total | Cover | 1.95 | 1.94 | 0.67 | (34.4) | 0.01 | (0.7) | ||
| Cover and Height | 2.04 | 2.04 | 1.12 | (54.9) | 0.00 | (0.1) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guevara, C.; Gonzalez-Benecke, C.; Wightman, M. Ground Cover—Biomass Functions for Early-Seral Vegetation. Forests 2021, 12, 1272. https://doi.org/10.3390/f12091272
Guevara C, Gonzalez-Benecke C, Wightman M. Ground Cover—Biomass Functions for Early-Seral Vegetation. Forests. 2021; 12(9):1272. https://doi.org/10.3390/f12091272
Chicago/Turabian StyleGuevara, Claudio, Carlos Gonzalez-Benecke, and Maxwell Wightman. 2021. "Ground Cover—Biomass Functions for Early-Seral Vegetation" Forests 12, no. 9: 1272. https://doi.org/10.3390/f12091272
APA StyleGuevara, C., Gonzalez-Benecke, C., & Wightman, M. (2021). Ground Cover—Biomass Functions for Early-Seral Vegetation. Forests, 12(9), 1272. https://doi.org/10.3390/f12091272