
Suppression of Ganoderma boninense Using Benzoic Acid: Impact on Cellular Ultrastructure and Anatomical Changes in Oil Palm Wood

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Screening of Benzoic Acid against the Growth of G. boninense Using “Poison Food” Technique
2.2. Morphological and Ultrastructural Alterations in the Mycelium of G. boninense
2.2.1. Scanning Electron Microscopy (SEM)
2.2.2. High-Resolution Transmission Electron Microscopy (HR-TEM)
2.3. Flow Cytometry
2.3.1. Membrane Depolarization Assay
2.3.2. Propidium Iodide Influx Assay
2.4. Biodegradation of Oil Palm Wood Blocks
2.5. Scanning Electron Microscopy (SEM) Analysis
2.6. Production of Ligninolytic and Hydrolytic Enzymes by G. boninense
2.6.1. Lignolytic Enzymes Assay
2.6.2. Laccase Enzyme Assay
2.6.3. Lignin Peroxidase Enzyme Assay
2.6.4. Manganese Peroxidase Enzyme Assay
2.6.5. Cellulase, Xylanase, and Amylase Enzymes Assays
2.7. Statistical Analysis
3. Results
3.1. Growth of G. boninense on the Media Incorporated with Benzoic Acid
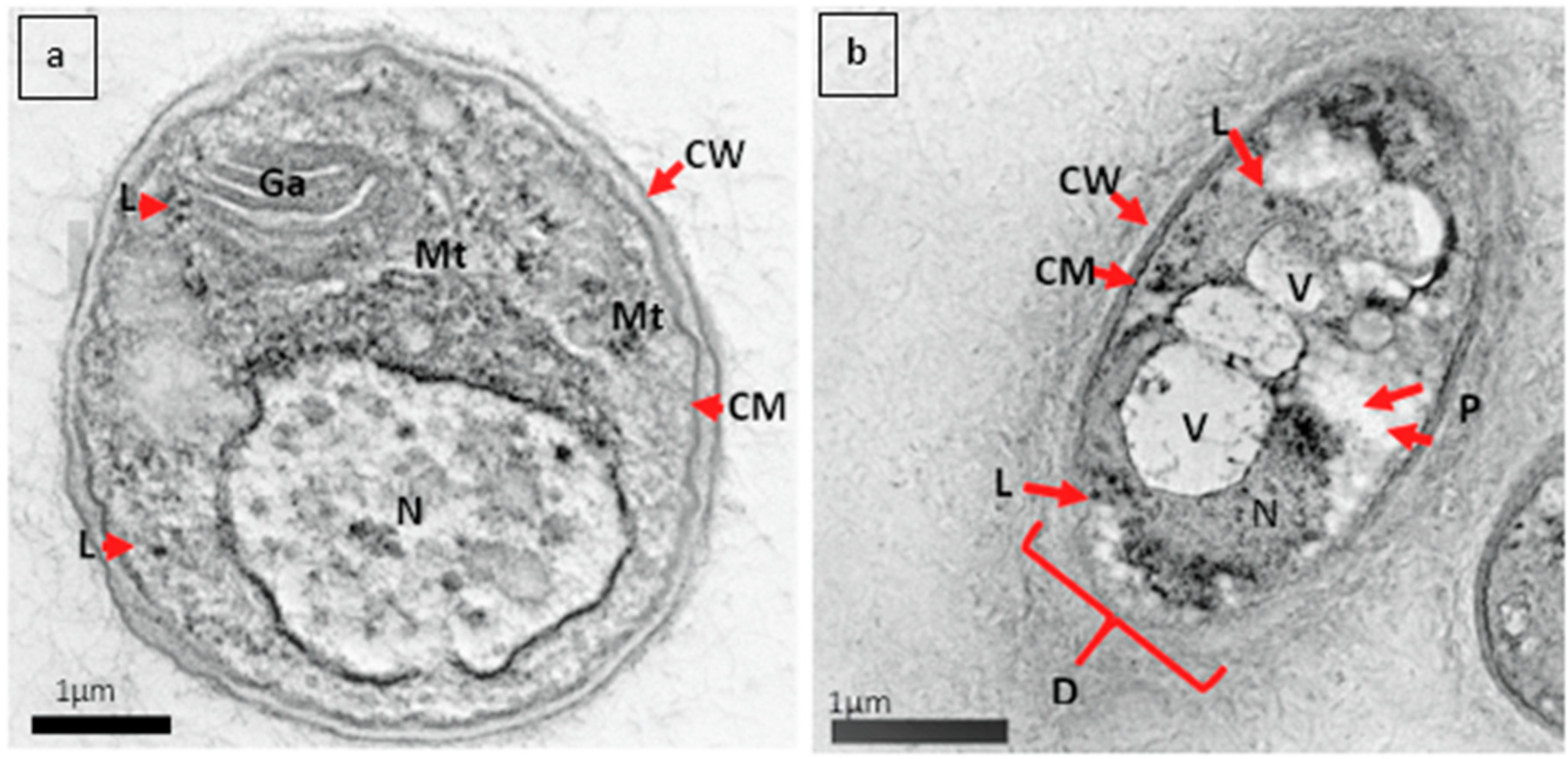
3.2. Alteration in the Ultrastructure of G. boninense
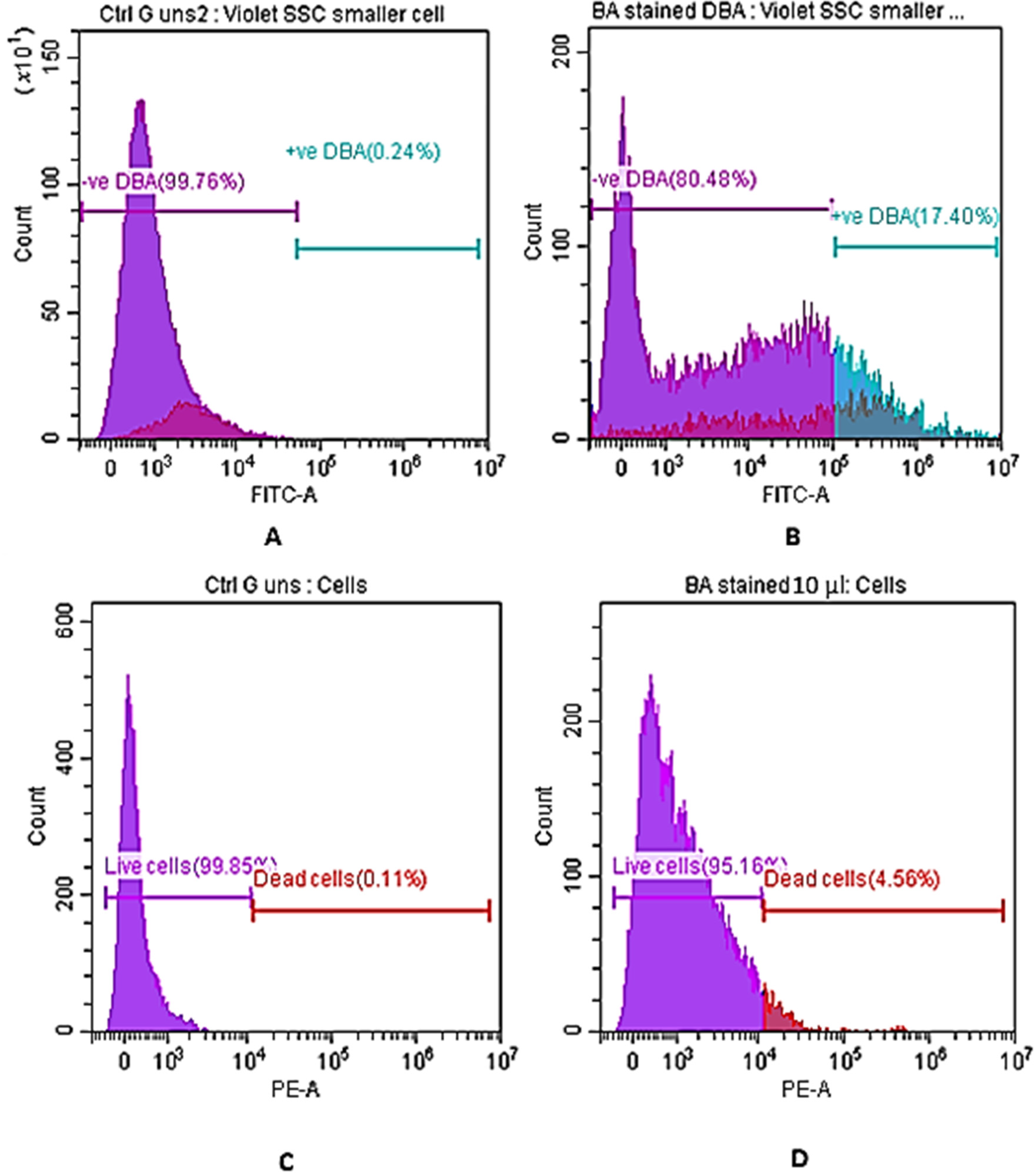
3.3. Cell Membrane Integrity and Depolarization
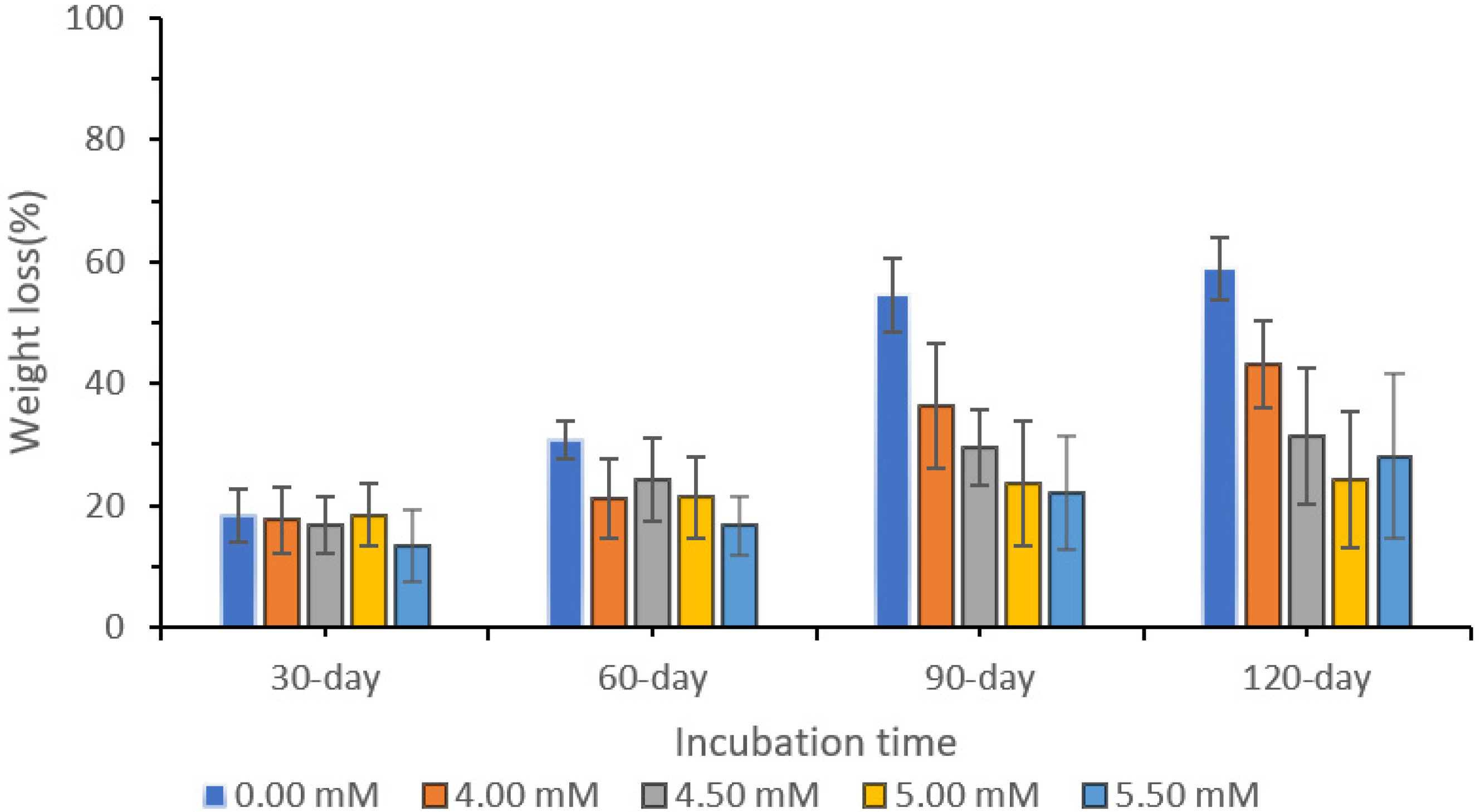
3.4. Biodegradation of Oil Palm Woodblock
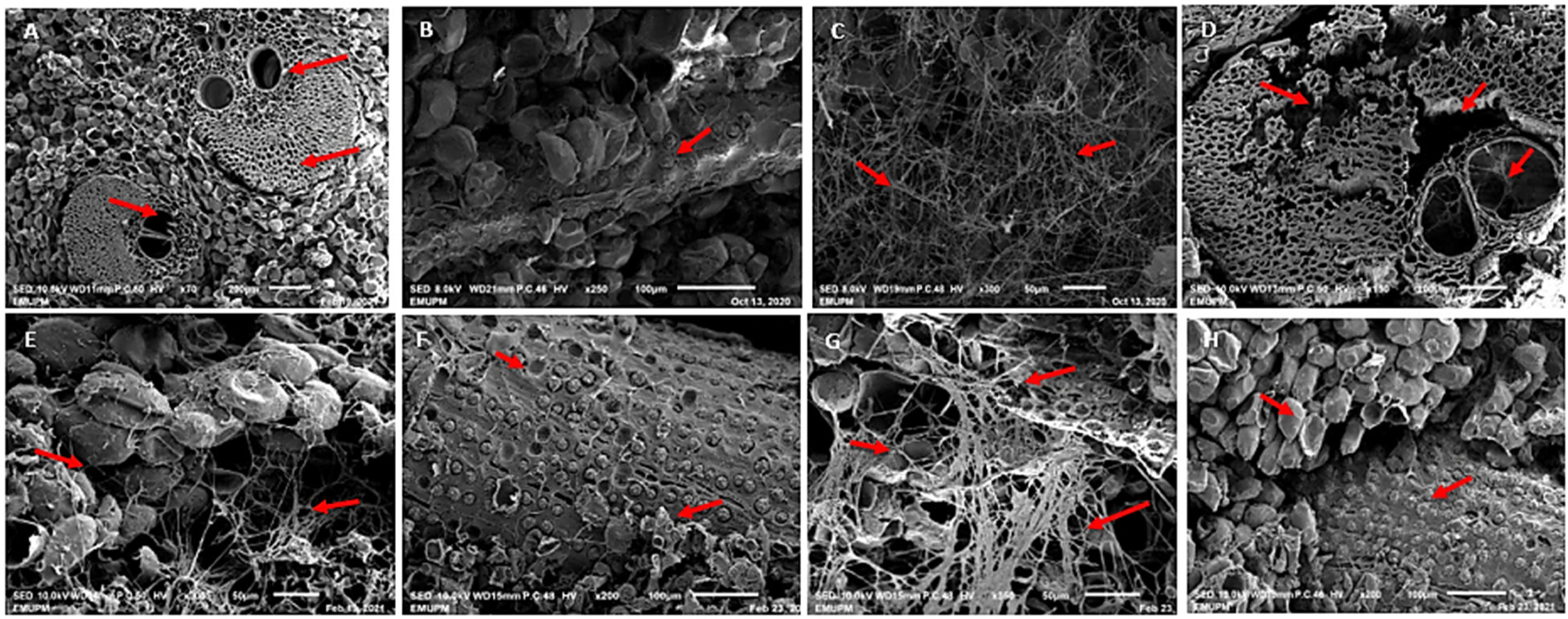
3.5. Anatomical Changes in Oil Palm Wood Structure
3.6. Ligninolytic Enzymes Activity of G. boninense
3.7. Hydrolytic Enzymes Activity of G. boninense
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- USDA Foreign Agricultural Services. Indonesia Oilseeds and Products Annual 2019; Global Agricultural Inforrmation Network. 2019; 16p, GAIN Report Number:ID1903. Available online: https://www.fas.usda.gov/data/indonesia-oilseeds-and-products-annual-3 (accessed on 24 July 2020).
- Uzir, M. Department of Statistics Malaysia: Press Release Malaysia Economic Performance Fourth Quarter 2018; Department of Statistics Malaysia: Putrajaya, Malaysia, 2019; pp. 1–5. [Google Scholar]
- Purnomo, H.; Okarda, B.; Dermawan, A.; Ilham, Q.P.; Pacheco, P.; Nurfatriani, F.; Suhendang, E. Reconciling oil palm economic development and environmental conservation in Indonesia: A value chain dynamic approach. For. Policy Econ. 2020, 111, 102089. [Google Scholar] [CrossRef]
- MPOB. Malaysian Oil Palm Statistics 2017, 37th ed.; MPOB: Bangi, Malaysia, 2018; 205p. [Google Scholar]
- Nossal, K.R.; Stubbs, R. Malaysia: An Emerging Middle Power? In Niche Diplomacy. Studies in Diplomacy; Cooper, A.F., Ed.; Palgrave Macmillan: London, UK, 1997. [Google Scholar]
- DPJ. Tree Crop Estate Statistics of Indonesia 2018–2020; Secretariate of Directorate General of Estate: Jakarta, Indonesia, 2019; 82p. [Google Scholar]
- Statistik, B.P. Statistical Yearbook of Indonesia; BPS-Statistics: Jakarta, Indonesia, 2021; 758p. [Google Scholar]
- Koh, L.P.; Wilcove, D.S. Is oil palm agriculture really destroying tropical biodiversity? Conserv. Lett. 2008, 1, 60–64. [Google Scholar] [CrossRef]
- USDA. Malaysia: Stagnating Palm Oil Yields Impede Growth. In Commodity Intelligence Report; USDA-FAS, Office of Global Analysis Report; USDA: Washington, DC, USA, 2012. [Google Scholar]
- Corley, R.H.V.; Tinker, P.B. The Oil Palm, 5th ed.; Wiley Balack-Well: Hoboken, NJ, USA, 2016; 674p. [Google Scholar]
- Cooper, R.M.; Flood, J.; Rees, R.W. Ganoderma boninense in oil palm plantations: Current thinking on epidemiology, resistance and pathology. Planter 2011, 87, 515–526. [Google Scholar]
- Idris, A.S.; Rajinder, S.; Madihah, A.Z.; Wahid, M.B. Multiplex PCR-DNA Kit for Early Detection and Identification of Ganoderma Species in Oil Palm; MPOB Information Series; Malaysian Palm Oil Board: Kuala Lumpur, Malaysia, 2010; pp. 1–3. [Google Scholar]
- Surendran, A.; Siddiqui, Y.; Ahmad, K.; Fernanda, R. Deciphering the Physicochemical and Microscopical Changes in Ganoderma boninense-Infected Oil Palm Woodblocks under the Influence of Phenolic Compounds. Plants 2021, 10, 1797. [Google Scholar] [CrossRef]
- Mohammed, C.; Rimbawanto, A.; Page, D. Management of basidiomycete root-and stem-rot diseases in oil palm, rubber and tropical hardwood plantation crops. For. Pathol. 2014, 44, 428–446. [Google Scholar] [CrossRef]
- Susanto, A.; Sudharto, P.; Purba, R. Enhancing biological control of basal stem rot disease (Ganoderma boninense) in oil palm plantations. Mycopathologia 2005, 159, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Assis, K.; Chong, K.P.; Idris, A.S.; Ho, C.M. Economic loss due to Ganoderma disease in oil palm. Int. J. Soc. Behav. Educ. Econ. Bus. Ind. Eng. 2016, 10, 604–608. [Google Scholar]
- Olaniyi, O.N.; Szulczyk, K.R. Estimating the economic damage and treatment cost of basal stem rot striking the Malaysian oil palms. For. Policy Econ. 2020, 116, 102163. [Google Scholar] [CrossRef]
- Midot, F.; Lau, S.Y.L.; Wong, W.C.; Tung, H.J.; Yap, M.L.; Lo, M.L.; Jee, M.S.; Dom, S.P.; Melling, L. Genetic diversity and demographic history of Ganoderma boninense in oil palm plantations of Sarawak, Malaysia inferred from ITS regions. Microorganisms 2019, 7, 464. [Google Scholar] [CrossRef] [Green Version]
- Roslan, A.; Idris, A.S. Economic Impact of Ganoderma Incidence on Malaysian Oil Palm Plantation—A Case Study in Johor. Oil Palm Ind. Econ. J. 2012, 12, 24–30. [Google Scholar]
- Kamu, A.; Mun, H.C.; Phin, C.K.; Seman, I.A. Identifying the early visible symptoms of the ganoderma-infected oil palms: A case study on the infected palms which collapsed within twelve months after disease census. ASM Sci. J. 2018, 11, 156–163. [Google Scholar]
- Najmie, M.; Khalid, K.; Sidek, A.; Jusoh, M. Density and ultrasonic characterization of oil palm trunk infected by Ganoderma boninense disease. Meas. Sci. Rev. 2011, 11, 160–164. [Google Scholar] [CrossRef] [Green Version]
- Durmaz, S.; Özgenç, Ö.; Boyaci, I.H.; YildIz, Ü.C.; Erişir, E. Examination of the chemical changes in spruce wood degraded by brown-rot fungi using FT-IR and FT-Raman spectroscopy. Vib. Spectrosc. 2016, 85, 202–207. [Google Scholar] [CrossRef]
- Rahmati, Y.; Nourmohammadi, K.; Naghdi, R.; Kartoolinejad, D. Effect of fungal degradation on physicochemical properties of exploited stumps of oriental beech over a 25-year felling period and the obtained Kraft pulp properties. J. For. Sci. 2019, 65, 96–105. [Google Scholar] [CrossRef]
- Lundell, T.K.; Mäkelä, M.R.; de Vries, R.P.; Hildén, K.S. Chapter Eleven—Genomics, Lifestyles and Future Prospects of Wood-Decay and Litter-Decomposing Basidiomycota. In Advances in Botanical Research; Martin, F.M., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 70, pp. 329–370. [Google Scholar]
- Koyani, R.D.; Bhatt, I.M.; Patel, H.R.; Vasava, A.M.; Rajput, K.S. Evaluation of Schizophyllum commune Fr. potential for biodegradation of lignin: A light microscopic analysis. Wood Mater. Sci. Eng. 2016, 11, 46–56. [Google Scholar] [CrossRef]
- Erwin Takemoto, S.; Hwang, W.J.; Takeuchi, M.; Itoh, T.; Imamura, Y. Anatomical characterization of decayed wood in standing light red meranti and identification of the fungi isolated from the decayed area. J. Wood Sci. 2008, 54, 233–241. [Google Scholar] [CrossRef]
- Rees, R.W.; Flood, J.; Hasan, Y.; Potter, U.; Cooper, R.M. Basal stem rot of oil palm (Elaeis guineensis); Mode of root infection and lower stem invasion by Ganoderma boninense. Plant Pathol. 2009, 58, 982–989. [Google Scholar] [CrossRef]
- Surendran, A.; Siddiqui, Y.; Ali, N.S.; Manickam, S. Inhibition and kinetic studies of cellulose- and hemicellulose-degrading enzymes of Ganoderma boninense by naturally occurring phenolic compounds. J. Appl. Microbiol. 2018, 124, 1544–1555. [Google Scholar] [CrossRef]
- Surendran, A.; Siddiqui, Y.; Saud, H.M.; Ali, N.S.; Manickam, S. Inhibition and kinetic studies of lignin degrading enzymes of Ganoderma boninense by naturally occurring phenolic compounds. J. Appl. Microbiol. 2018, 125, 876–887. [Google Scholar] [CrossRef]
- Bucher, V.V.C.; Hyde, K.D.; Pointing, S.B.; Reddy, C.A. Production of wood decay enzymes, mass loss and lignin solubilization in wood by marine ascomycetes and their anamorphs. Fungal Divers. 2004, 15, 1–14. [Google Scholar]
- Siddiqui, Y.; Surendran, A.; Paterson, R.R.M.; Ali, A.; Ahmad, K. Current strategies and perspectives in detection and control of basal stem rot of oil palm. Saudi J. Biol. Sci. 2021, 28, 2840–2849. [Google Scholar] [CrossRef]
- Williams, M.; Senaratna, T.; Dixon, K.; Sivasithamparam, K. Benzoic acid induces tolerance to biotic stress caused by Phytophthora cinnamomi in Banksia attenuata. Plant Growth Regul. 2003, 41, 89–91. [Google Scholar] [CrossRef]
- Zeece, M. Chapter Seven—Food additives. In Introduction to the Chemistry of Food; Zeece, M., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 251–311. [Google Scholar]
- Del Olmo, A.; Calzada, J.; Nuñez, M. Benzoic acid and its derivatives as naturally occurring compounds in foods and as additives: Uses, exposure, and controversy. Crit. Rev. Food Sci. Nutr. 2017, 57, 3084–3103. [Google Scholar] [CrossRef]
- Surendran, A.; Siddiqui, Y.; Saud, H.M.; Ali, N.S.; Manickam, S. The antagonistic effect of phenolic compounds on ligninolytic and cellulolytic enzymes of Ganoderma boninense, causing basal stem rot in oil palm. Int. J. Agric. Biol. 2017, 19, 1437–1446. [Google Scholar]
- Balamurugan, S. In Vitro Antifungal Activity of Citrus aurantifolia Linn Plant Extracts against Phytopathogenic Fungi Macrophomina phaseolina. Int. Lett. Nat. Sci. 2014, 13, 70–74. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Lee, D.G. An antifungal mechanism of curcumin lies in membrane-targeted action within Candida albicans. IUBMB Life 2014, 66, 1521–6551. [Google Scholar] [CrossRef] [PubMed]
- Schirp, A.; Wolcott, M.P. Influence of fungal decay and moisture absorption on mechanical properties of extruded wood-plastic composites. Wood Fiber Sci. 2005, 37, 643–652. [Google Scholar]
- Naidu, Y.; Siddiqui, Y.; Rafii, M.Y.; Saud, H.M.; Idris, A.S. Investigating the effect of white-rot hymenomycetes biodegradation on basal stem rot infected oil palm wood blocks: Biochemical and anatomical characterization. Ind. Crop. Prod. 2017, 108, 872–882. [Google Scholar] [CrossRef]
- More, S.S.; Renuka, P.S.; Pruthvi, K.; Swetha, M.; Malini, S.; Veena, S.M. Isolation, Purification, and Characterization of Fungal Laccase from Pleurotus sp. Enzym. Res. 2011, 2011, 248735. [Google Scholar] [CrossRef] [Green Version]
- Pizzolitto, R.P.; Barberis, C.L.; Dambolena, J.S.; Herrera, J.M.; Zunino, M.P.; Magnoli, C.E.; Rubinstein, H.R.; Zygadlo, J.A.; Dalcero, A.M. Inhibitory Effect of Natural Phenolic Compounds on Aspergillus parasiticus Growth. J. Chem. 2015, 2015, 547925. [Google Scholar] [CrossRef] [Green Version]
- Dambolena, J.S.; López, A.G.; Meriles, J.M.; Rubinstein, H.R.; Zygadlo, J.A. Inhibitory effect of 10 natural phenolic compounds on Fusarium verticillioides. A structure–property–activity relationship study. Food Control 2012, 28, 163–170. [Google Scholar] [CrossRef]
- Hou, R.; Shi, J.; Ma, X.; Wei, H.; Hu, J.; Tsang, Y.F.; Gao, M.T. Effect of Phenolic Acids Derived from Rice Straw on Botrytis cinerea and Infection on Tomato. Waste Biomass Valorization 2020, 11, 6555–6563. [Google Scholar] [CrossRef]
- Amborabé, B.E.; Fleurat-Lessard, P.; Chollet, J.F.; Roblin, G. Antifungal effects of salicylic acid and other benzoic acid derivatives towards Eutypa lata: Structure-activity relationship. Plant Physiol. Biochem. 2002, 40, 1051–1060. [Google Scholar] [CrossRef]
- Siddiqui, Y.; Surendran, A.; Masya, E.; Agricultural, F.; Sdn, S. Inhibition of Lignin Degrading Enzymes of Ganoderma spp.: An Alternative Control of Basal Stem Rot Disease of Oil Palm gth Article. Int. J. Agric. Biol. 2019, 3, 523–530. [Google Scholar]
- Mikulic-Petkovsek, M.; Schmitzer, V.; Stampar, F.; Veberic, R.; Koron, D. Changes in phenolic content induced by infection with Didymella applanata and Leptosphaeria coniothyrium, the causal agents of raspberry spur and cane blight. Plant Pathol. 2014, 63, 185–192. [Google Scholar] [CrossRef]
- Liu, J.; Zong, Y.; Qin, G.; Li, B.; Tian, S. Plasma membrane damage contributes to antifungal activity of silicon against penicillium digitatum. Curr. Microbiol. 2010, 61, 274–279. [Google Scholar] [CrossRef]
- You, Z.; Zhang, C.; Ran, Y. The effects of clioquinol in morphogenesis, cell membrane and ion homeostasis in Candida albicans. BMC Microbiol. 2020, 20, 165. [Google Scholar] [CrossRef]
- Alcazar-Fuoli, L.; Mellado, E. Ergosterol biosynthesis in Aspergillus fumigatus: Its relevance as an antifungal target and role in antifungal drug resistance. Front Microbiol 2013, 3, 439. [Google Scholar] [CrossRef] [Green Version]
- Kazan, K.; Gardiner, D.M. Targeting pathogen sterols: Defence and counterdefence? PLoS Pathog. 2017, 13, e1006297. [Google Scholar] [CrossRef]
- Jordá, T.; Puig, S. Regulation of ergosterol biosynthesis in saccharomyces cerevisiae. Genes 2020, 11, 795. [Google Scholar] [CrossRef]
- Yang, C.; Hamel, C.; Vujanovic, V.; Gan, Y. Fungicide: Modes of Action and Possible Impact on Nontarget Microorganisms. ISRN Ecol. 2011, 2011, 130289. [Google Scholar] [CrossRef] [Green Version]
- Villavicencio, E.V.; Mali, T.; Mattila, H.K.; Lundell, T. Enzyme activity profiles produced on wood and straw by four fungi of different decay strategies. Microorganisms 2020, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Dashtban, M.; Schraft, H.; Syed, T.A.; Qin, W. Fungal biodegradation and enzymatic modification of lignin. Int. J. Biochem. Mol. Biol. 2010, 1, 36–50. [Google Scholar]
- Martínez, Á.T.; Speranza, M.; Ruiz-Dueñas, F.J.; Ferreira, P.; Camarero, S.; Guillén, F.; Martínez, M.J.; Gutiérrez, A.; Del Río, J.C. Biodegradation of lignocellulosics: Microbial, chemical, and enzymatic aspects of the fungal attack of lignin. Int. Microbiol. 2005, 8, 195–204. [Google Scholar] [PubMed]
- Sanghvi, G.V.; Koyani, R.D.; Rajput, K.S. Anatomical characterisation of teak (Tectona grandis) wood decayed by fungus Chrysosporium asperatum. J. Trop. For. Sci. 2013, 25, 547–553. [Google Scholar]
- Widhalm, J.R.; Dudareva, N. A familiar ring to it: Biosynthesis of plant benzoic acids. Mol. Plant 2015, 8, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Surendran, A.; Siddiqui, Y.; Manickam, S.; Ali, A. Role of benzoic and salicylic acids in the immunization of oil palm seedlings-challenged by Ganoderma boninense. Ind. Crop. Prod. 2018, 122, 358–365. [Google Scholar] [CrossRef]
- Ding, P.; Ding, Y. Stories of Salicylic Acid: A Plant Defense Hormone. Trends Plant Sci. 2020, 25, 549–565. [Google Scholar] [CrossRef]
- Bari, E.; Nazarnezhad, N.; Kazemi, S.M.; Tajick Ghanbary, M.A.; Mohebby, B.; Schmidt, O.; Clausen, C.A. Comparison between degradation capabilities of the white rot fungi Pleurotus ostreatus and Trametes versicolor in beech wood. Int. Biodeterior. Biodegrad. 2015, 104, 231–237. [Google Scholar] [CrossRef]
- Andlar, M.; Rezić, T.; Marđetko, N.; Kracher, D.; Ludwig, R.; Šantek, B. Lignocellulose degradation: An overview of fungi and fungal enzymes involved in lignocellulose degradation. Eng. Life Sci. 2018, 18, 768–778. [Google Scholar] [CrossRef]
- Kracher, D.; Ludwig, R. Cellobiose dehydrogenase: An essential enzyme for lignocellulose degradation in nature—A review/Cellobiosedehydrogenase: Ein essentielles Enzym für den Lignozelluloseabbau in der Natur—Eine Übersicht. Die Bodenkult. J. Land Manag. Food Environ. 2016, 67, 145–163. [Google Scholar] [CrossRef] [Green Version]
- Govender, N.; Wong, M.Y. Detection of oil palm root penetration by agrobacterium-mediated transformed Ganoderma boninense, expressing green fluorescent protein. Phytopathology 2017, 107, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, O. Indoor wood-decay basidiomycetes: Damage, causal fungi, physiology, identification and characterization, prevention and control. Mycol. Prog. 2007, 6, 261. [Google Scholar] [CrossRef]
- Naidu, Y.; Idris, A.S.; Nusaibah, S.A.; Norman, K.; Siddiqui, Y. In vitro screening of biocontrol and biodegradation potential of selected hymenomycetes against Ganoderma boninense and infected oil palm waste. For. Pathol. 2015, 45, 474–483. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Benzoic Acid Concentrations (mM) | Growth Rate (cm Day−1) | PIDG (%) |
|---|---|---|
| 0.00 | 0.83 | 0 |
| 4.00 | 0.40 | 62.37 ± 1.4 e |
| 4.25 | 0.35 | 68.21 ± 1.7 d |
| 4.50 | 0.27 | 69.43 ± 1.5 d |
| 4.75 | 0.13 | 75.31 + 1.6 c |
| 5.00 | 0.10 | 80.00 + 1.0 b |
| 5.25 | 0 | 100 a |
| 5.50 | 0 | 100 a |
| Benzoic Acid Concentration (mM) | Enzyme Activity | ||
|---|---|---|---|
| Laccase (U/L) | Lignin Peroxidase (U/mL) | Manganese Peroxidase (U/L) | |
| 0.00 | 0.67 b | 0.001 b | 5.36 a |
| 2.50 | 0.33 c | 0.066 a | 0.49 e |
| 3.50 | 0.25 d | 0.109 ba | 1.57 b |
| 4.50 | 1.18 a | 0.107 a | 1.25 d |
| 5.00 | 0.26d | 0.079 ab | 1.49 c |
| Benzoic Acid Concentration (mM) | Enzymes Activity (U/mL) | ||
|---|---|---|---|
| Cellulase | Amylase | Xylanase | |
| 0.00 | 0.64 a | 2.04 a | 1.45 a |
| 2.50 | 0.19 c | 2.24 a | 1.16 b |
| 3.50 | 0.09 e | 2.02 a | 1.51 a |
| 4.50 | 0.10 d | 1.92 a | 1.03 b |
| 5.00 | 0.23 b | 2.10 a | 0.50 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernanda, R.; Siddiqui, Y.; Ganapathy, D.; Ahmad, K.; Surendran, A. Suppression of Ganoderma boninense Using Benzoic Acid: Impact on Cellular Ultrastructure and Anatomical Changes in Oil Palm Wood. Forests 2021, 12, 1231. https://doi.org/10.3390/f12091231
Fernanda R, Siddiqui Y, Ganapathy D, Ahmad K, Surendran A. Suppression of Ganoderma boninense Using Benzoic Acid: Impact on Cellular Ultrastructure and Anatomical Changes in Oil Palm Wood. Forests. 2021; 12(9):1231. https://doi.org/10.3390/f12091231
Chicago/Turabian StyleFernanda, Rozi, Yasmeen Siddiqui, Daarshini Ganapathy, Khairulmazmi Ahmad, and Arthy Surendran. 2021. "Suppression of Ganoderma boninense Using Benzoic Acid: Impact on Cellular Ultrastructure and Anatomical Changes in Oil Palm Wood" Forests 12, no. 9: 1231. https://doi.org/10.3390/f12091231
APA StyleFernanda, R., Siddiqui, Y., Ganapathy, D., Ahmad, K., & Surendran, A. (2021). Suppression of Ganoderma boninense Using Benzoic Acid: Impact on Cellular Ultrastructure and Anatomical Changes in Oil Palm Wood. Forests, 12(9), 1231. https://doi.org/10.3390/f12091231