
Climate Warming Impacts on Distributions of Scots Pine (Pinus sylvestris L.) Seed Zones and Seed Mass across Russia in the 21st Century


Abstract
:1. Introduction
2. Data and Methods
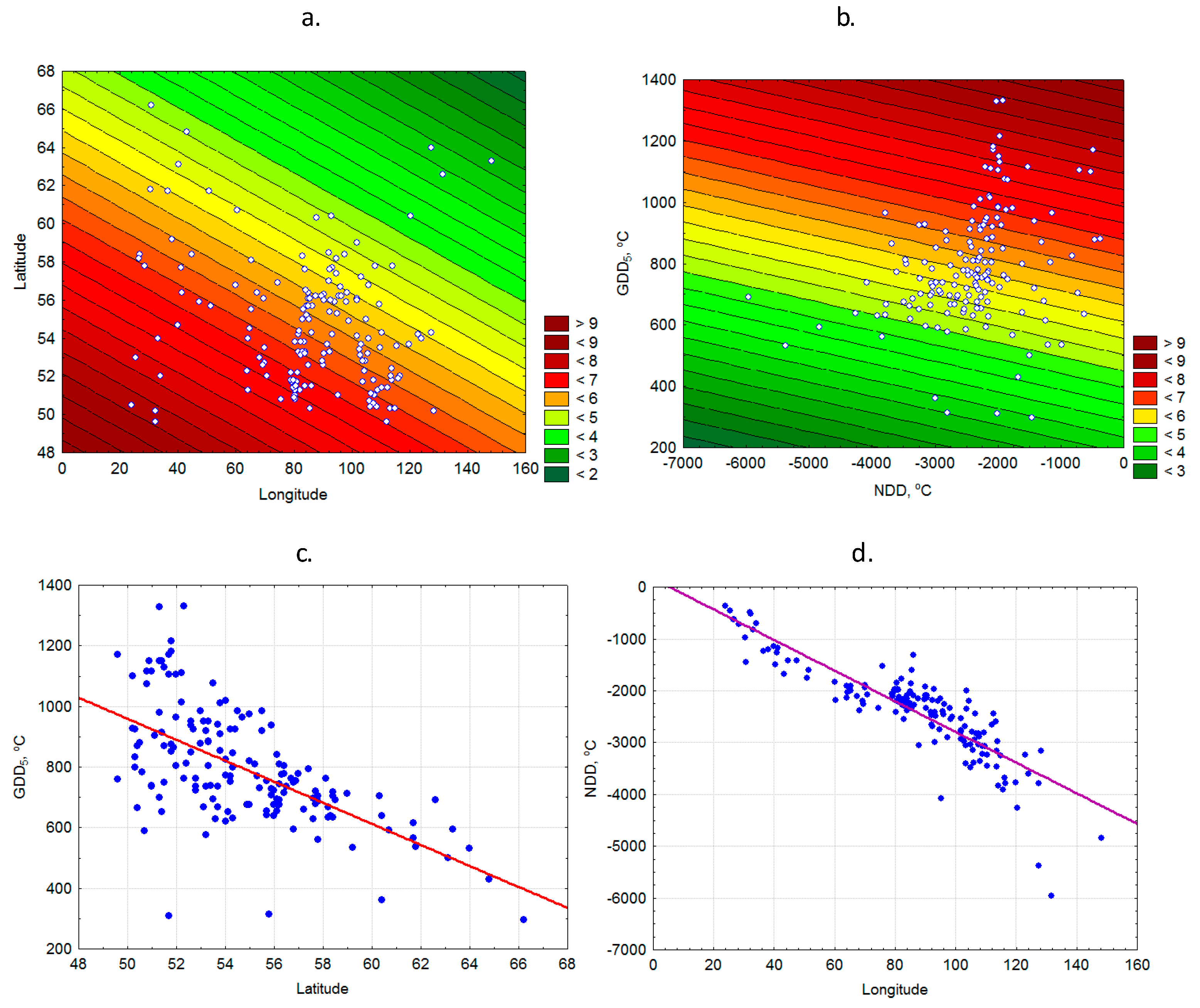
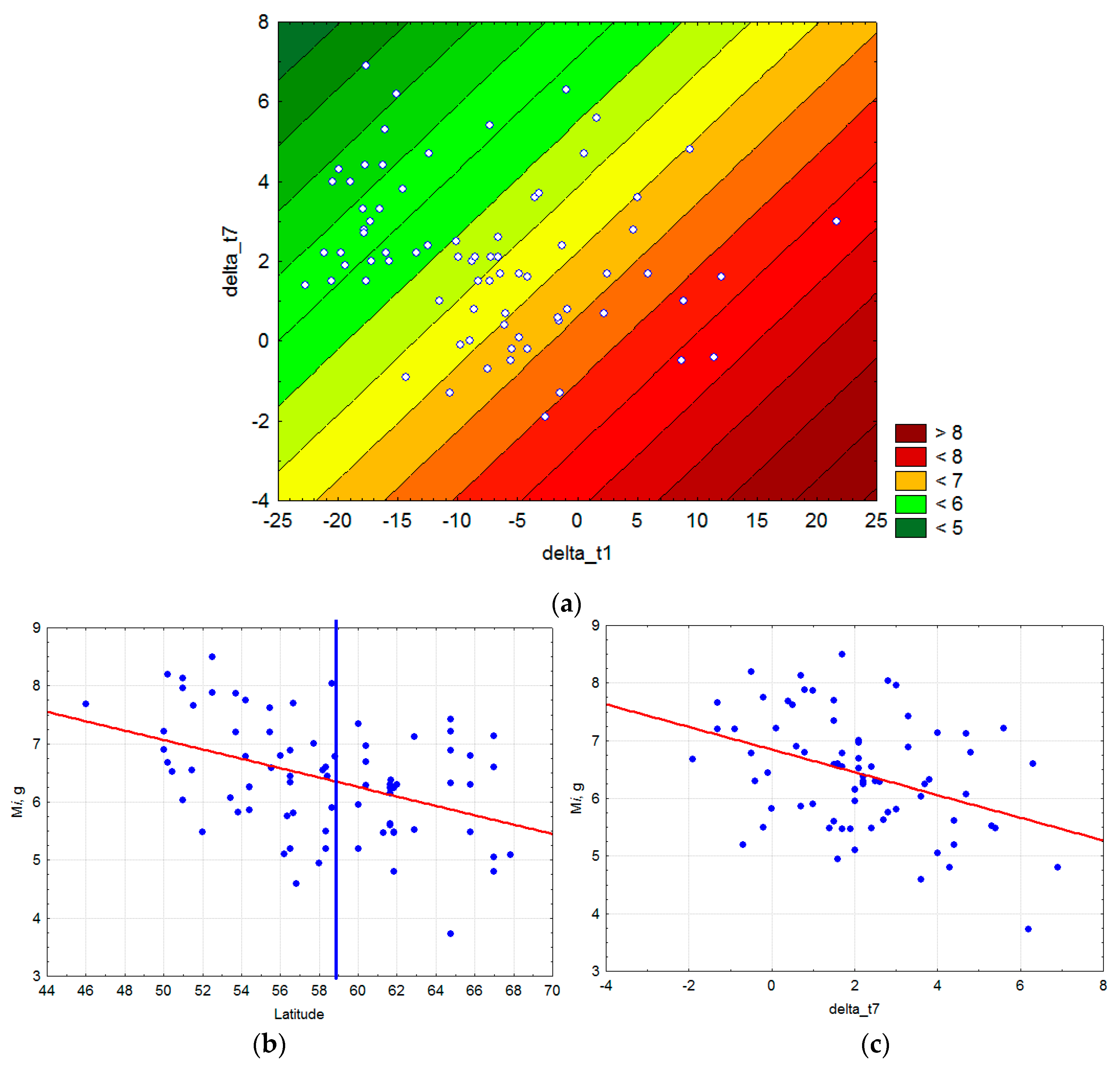
2.1. Seed Mass Data
2.2. Climate Data
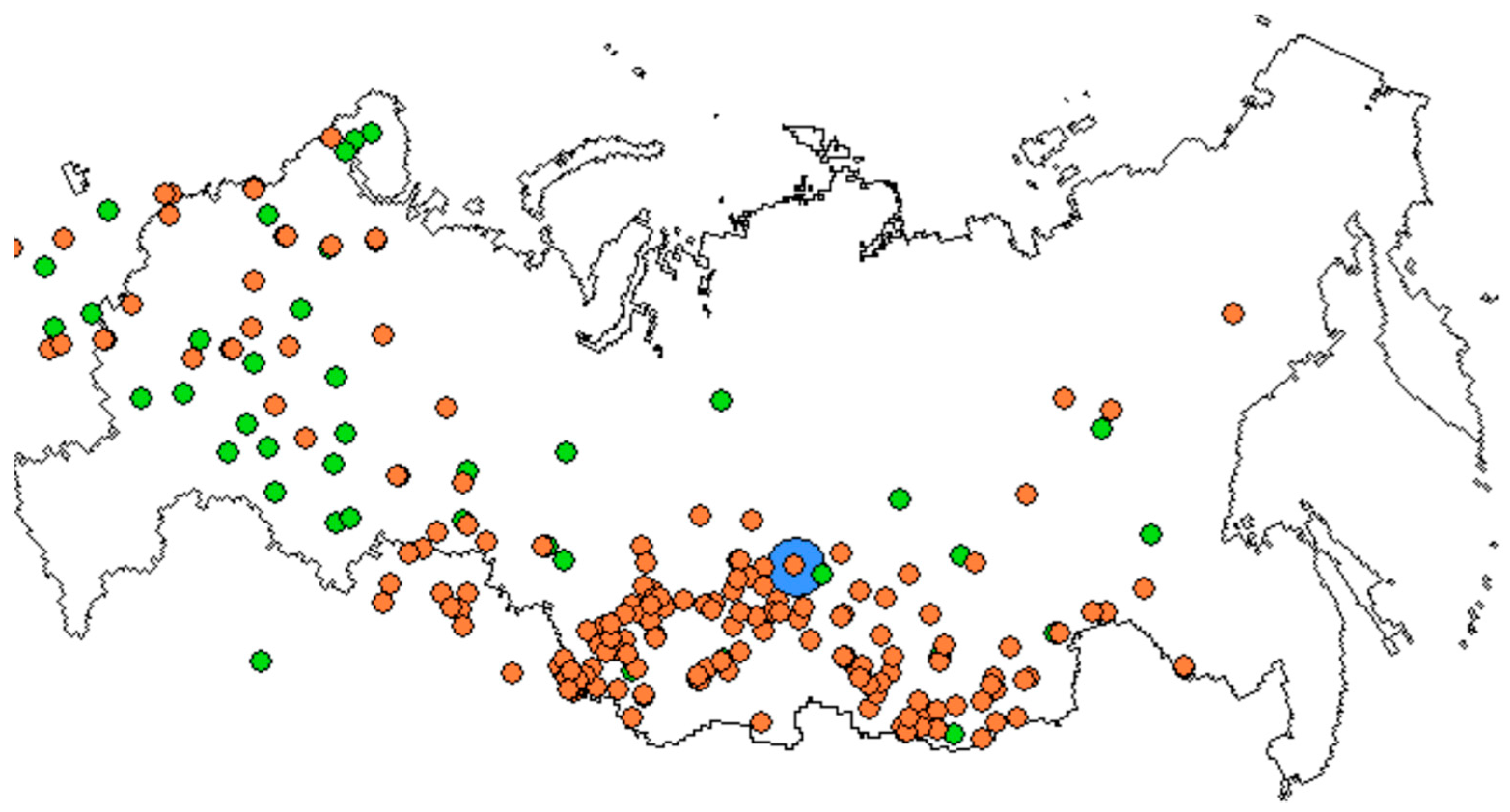
2.3. P. sylvestris Range and Seed Zones across Russia
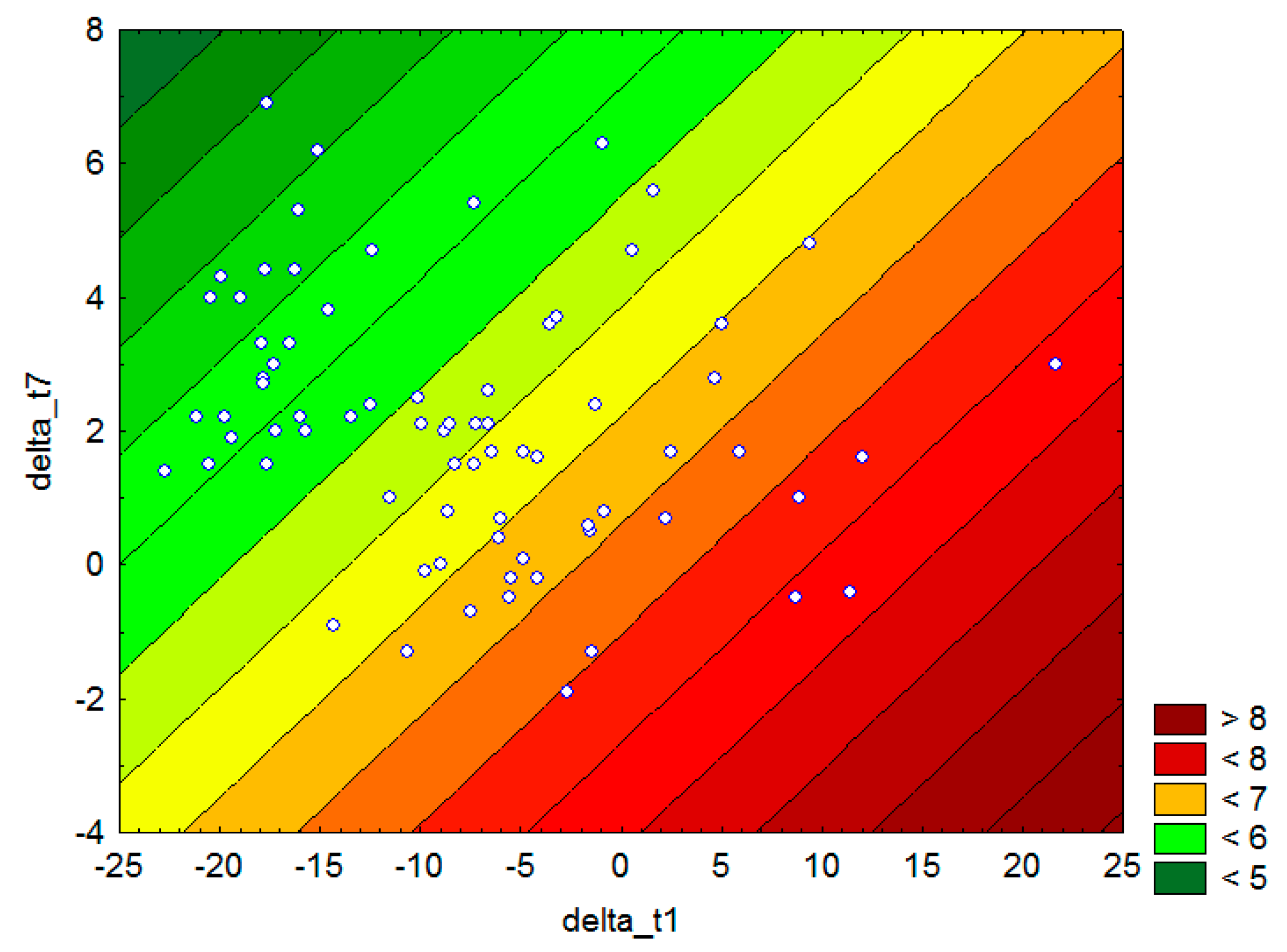
2.4. Experiments with Seed Mass during the Common Garden Trial at the Test Site
2.5. Mapping Seed Zones and Seed Mass across Russia
3. Results
3.1. Bioclimatic Models of P. sylvestris Seed Mass
3.2. Distribution of P. sylvestris Seed Zones in a Warming Climate by 2080
3.3. Coupled Seed Zones and Seed Mass Distributions in Contemporary and Future Climates
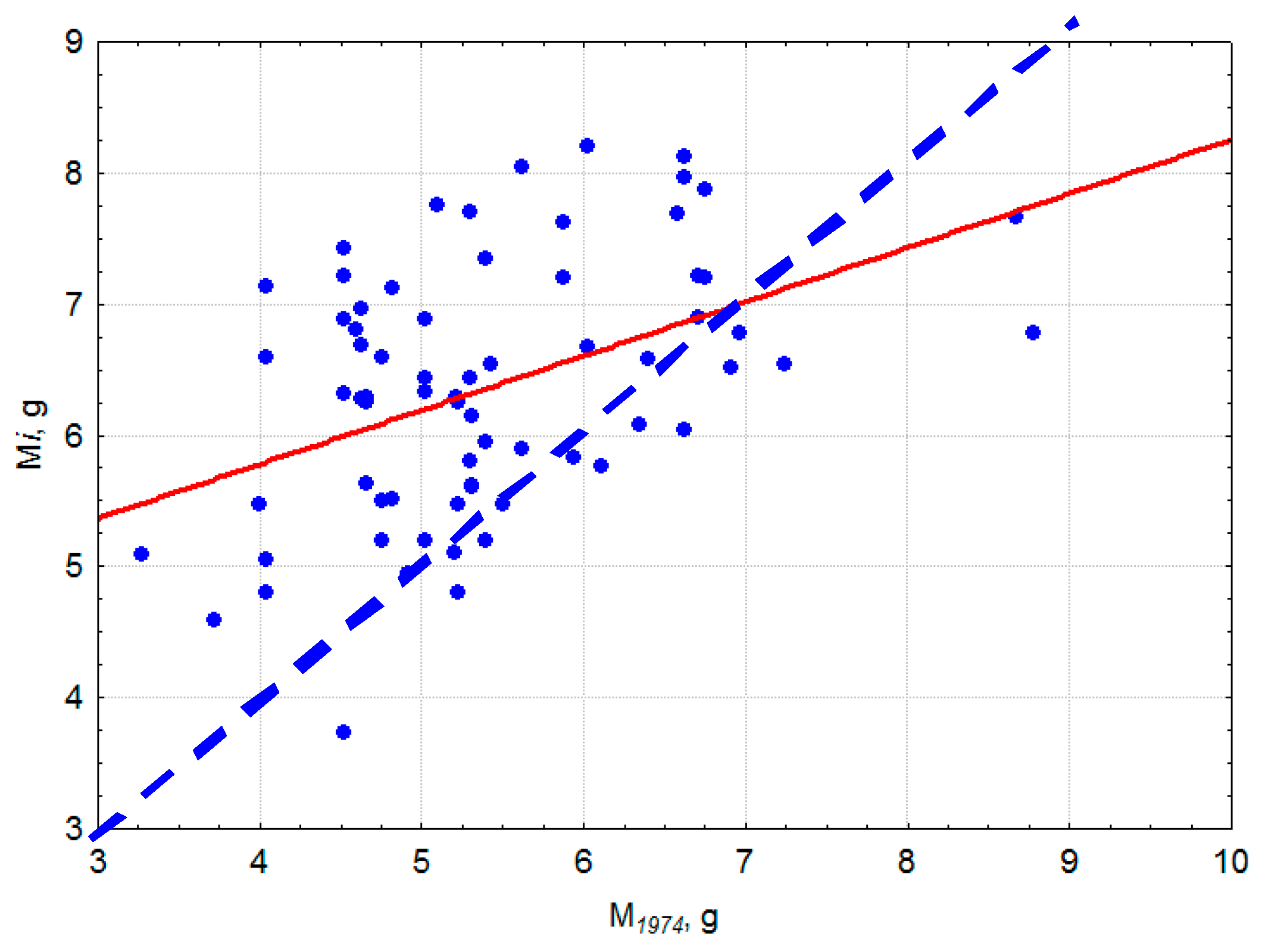
3.4. The 40-Year Common Garden Trial in the Boguchany
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pravdin, L.F. Pinus sylvestris; Nauka: Moscow, Russia, 1964; p. 192. [Google Scholar]
- Mamaev, S.A. Intraspecies Variability of Woody Plants (A Case Study of the Family Pinaceae in the Ural mountains); Nauka: Moscow, Russia, 1973; p. 284. [Google Scholar]
- Cherepnin, V.L. Pinus sylvestris Seeds Variability; Nauka: Novosibirsk, Russia, 1980; p. 182. [Google Scholar]
- Giertych, M.; Matyas, C. Genetics of Scots Pine. Developments of Plant Genetics and Breeding; Elsevier: Amsterdam, The Netherlands, 1991; Volume 3, p. 260. [Google Scholar]
- Mátyás, C.; Ackzell, L.; Samuel, C.J.A. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Scots Pine (Pinus sylvestris); International Plant Genetic Resources Institute: Rome, Italy, 2004; p. 6. [Google Scholar]
- Moles, A.T.; Westoby, M. Latitude, seed predation and seed mass. J. Biogeogr. 2003, 30, 105–128. [Google Scholar] [CrossRef]
- Castro, J.; Reich, P.B.; Sanchez-Miranda, A.; Guerrero, J.D. Evidence that the negative relationship between seed mass and relative growth rate is not physiological but linked to species identity: A within-family analysis of Scots pine. Tree Physiol. 2008, 28, 1077–1082. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Bai, Y.; Lamb, E.G.; Simpson, D.; Liu, G.; Wei, Y.; Wang, D.; McKenney, D.W.; Papadopol, P. Patterns of Cross-Continental Variation in Tree Seed Mass in the Canadian Boreal Forest. PLoS ONE 2013, 8, e61060. [Google Scholar] [CrossRef] [PubMed]
- Himanen, K.; Helenius, P.; Ylioja, T.; Nygren, M. Intracone variation explains most of the variance in Picea abies seed weight: Implications for seed sorting. Can. J. For. Res. 2016, 46, 470–477. [Google Scholar] [CrossRef]
- Freire, J.A.; Rodrigues, G.C.; Tome, M. Climate change impacts on Pinus pinea L. silvicultural system for cone production and ways to contour those impacts: A review complemented with data from permanent plots. Forests 2019, 10, 169. [Google Scholar] [CrossRef] [Green Version]
- Balekoglu, S.; Caliskan, S.; Dirik, H. Effects of geoclimatic factors on the variability in Pinus pinea cone, seed, and seedling traits in Turkey native habitats. Ecol. Process. 2020, 9, 55. [Google Scholar] [CrossRef]
- Suarez-Vidal, E.; Sampedro, L.; Zas, R. Is the benefit of larger seed provisioning on seedling performance greater stress? Environ. Exp. Bot. 2017, 134, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Loewe, M.V.; Balzarini, M.; Alvarez, C.A.; Delard, R.C.; Navarro, C.R. Fruit productivity of Stone pine (Pinus pinea L.) along a climatic gradient in Chile. Agric. For. Meteorol. 2016, 223, 203–216. [Google Scholar] [CrossRef]
- Iroshnikov, A.I. Provenance Trials of Conifers in South Siberia. Provenance Trials and Plantations of Conifers in Siberia; Nauka: Novosibirsk, Russia, 1977; pp. 4–110. [Google Scholar]
- Kuzmina, N.A. Pinus sylvestris variability in Angara region. Lesovedenie 1985, 2, 40–46. [Google Scholar]
- Sivacioğlu, A.; Ayan, S. Evaluation of seed production of scots pine (Pinus sylvestris L.) clonal seed orchard with cone analysis method. African J. Biotechnol. 2008, 7, 4393–4399. [Google Scholar] [CrossRef]
- Matías, L.; Jump, A.S. Impacts of predicted climate change on recruitment at the geographical limits of Scots pine. J. Exp. Bot. 2014, 65, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batkhuu, N.; Udval, B.; Jigjid, B.; Jamyansuren, S.; Fischer, M. Seed and cone morphological variation and seed germination characteristics of scots pine populations (Pinus sylvestris L.) in Mongolia. Mong. J. Biol. Sci. 2020, 18, 41–54. [Google Scholar] [CrossRef]
- Kuzmina, N.A. Specific features of Scots pine provenance trials in Angara river basin. Lesovedenie 1999, 4, 23–29. (In Russian) [Google Scholar]
- Kuzmina, N.A.; Kuzmin, S.R. Features of generative organs of Scots pine of different origin in the provenance trial. Conifers Boreal Area 2007, 24, 225–234. [Google Scholar]
- Kuzmina, N.A.; Senashova, V.A.; Kuzmin, S.R. Distribution of Lophodermium needle cast agents in Scots pine stands in Central Siberia. Contemp. Probl. Ecol. 2015, 8, 909. [Google Scholar] [CrossRef]
- Kuzmin, S.R.; Kuzmina, N.A. Choosing of Promising Climatypes of Scots Pine in Geographic Cultures in Different Forest Growth Conditions. Lesovedenie 2020, 451–465. [Google Scholar] [CrossRef]
- Kuzmin, S.R.; Kuzmina, N.A. Forest Seed Regions of Scots Pine Based on Growth Assessement in the Provenance Trial in Siberia. Sibirskij Lesnoj Zurnal 2020, 6, 3–15. [Google Scholar] [CrossRef]
- Kuzmin, S.R. Response of Annual Ring Width and Latewood Content of Scots pine to Weather Conditions in Provenance Trials. Lesn. Zhurnal 2020, 5, 64–80. [Google Scholar] [CrossRef]
- Pobedinsky, A.V. Pinus sylvestris; Forestry Publishers: Moscow, Russia, 1979; p. 128. [Google Scholar]
- Tchebakova, N.M.; Parfenova, E.; Soja, A.J. The effects of climate, permafrost and fire on vegetation change in Siberia in a changing climate. Environ. Res. Lett. 2009, 4, 045013. [Google Scholar] [CrossRef]
- Parfenova, E.I.; Tchebakova, N.M.; Soja, A.J. Assessing landscape potential for human sustainability and ‘attractiveness’ across Asian Russia in a warmer 21st century. Environ. Res. Lett. 2019, 14, 065004. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Tchebakova, N.M.; Barnhardt, L.K. Efficacy of climate transfer functions: Introduction of Eurasian populations of Larix into Alberta. Can. J. For. Res. 1999, 29, 1660–1668. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Tchebakova, N.M.; Parfenova, E.I.; Wykoff, W.R.; Kuzmina, N.A.; Milyutin, L.I. Intraspecific responses to climate in Pinus sylvestris. Glob. Chang. Biol. 2002, 8, 912–929. [Google Scholar] [CrossRef] [Green Version]
- Rehfeldt, G.E.; Tchebakova, N.M.; Milyutin, L.I.; Parfenova, E.I.; Wykoff, W.R.; Kouzmina, N.A. Assessing population responses to climate in Pinus sylvestris and Larix spp. of Eurasia with Climate-Transfer Models. Eurasian J. Res. 2003, 6, 83–98. [Google Scholar]
- Tchebakova, N.M.; Rehfeldt, G.E.; Parfenova, E.I. From vegetation zones to climatypes: Effects of climate warming on Siberian ecosystems. In Permafrost Ecosystems. Siberian Larch Forests; Osawa, A., Zyryanova, O.A., Matsuura, Y., Kajimoto, T., Wein, R.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 427–447. [Google Scholar]
- Pozdnyakov, L.K. Permafrost Forestry; Nauka: Novosibirsk, Russia, 1986; p. 192. [Google Scholar]
- Khotinsky, N.A. The Holocene of the Northern Eurasia; Nauka: Moscow, Russia, 1977; p. 200. [Google Scholar]
- Tchebakova, N.M.; Blyakharchuk, T.A.; Parfenova, E.I. Reconstruction and prediction of climate and vegetation change in the Holocene in the Altai-Sayan mountains, Central Asia. Environ. Res. Lett. 2009, 4, 045025. [Google Scholar] [CrossRef] [Green Version]
- Boratynski, A. Range of natural distribution. In Developments in Plant Genetics and Breeding; Giertych, M., Mâtyâs, C., Eds.; Elsevier: Amsterdam, The Netherlands, 1991; pp. 19–30. [Google Scholar]
- Hauke-Kowalska, M.; Borowiak, E.; Barzdajn, W.; Kowalkwski, W.; Korzeniewicz, R.; Wawro, T. Cone and seeds variability in seed orchards and seed stands of Pinus sylvestris L. Balt. For. 2019, 25, 187–192. [Google Scholar] [CrossRef]
- Przybylski, P.; Masternak, K.; Jastrzębowski, S. Isozyme polymorphism and seed and cone variability of Scots pine (Pinus sylvestris L.) in relation to local environments in Poland. Folia For. Pol. Ser. A 2020, 62, 88–99. [Google Scholar] [CrossRef]
- Mikola, J. The effect of seed size and durarion of growth on the height of Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies (L.) Karst.) provenances and progenies at the nursery stage. Silva Fenn. 1980, 14, 84–94. [Google Scholar] [CrossRef]
- Prescher, F.; Lindgren, D.; Wennstrom, U.; Almqvist, C.; Ruotsalainen, S.; Kroon, J. Seed production in Scots pine seed orchards. In Status, Monitoring and Targets for Breeding Programs, Proceedings of the Meeting of Nordic Forest Tree Breeders and Forest Genetics, Syktyvkar, Russia, 13–15 September 2015; Fedorkov, A., Ed.; Russian Academy of Sciences: Moscow, Russia, 2005; pp. 65–71. [Google Scholar]
- Debain, S.; Curt, T.; Lepart, J.; Prevosto, B. Reproductive variability in Pinus sylvestris in southern France: Implications for invasion. J. Veg. Sci. 2003, 14, 509–516. [Google Scholar] [CrossRef]
- Kaliniewicz, Z.; Rawa, T.; Tylek, P.; Markowski, P.; Anders, A.; Fura, S. The effect of the age of Scots pine (Pinus sylvestris L.) stands on the physical properties of seeds and the operating parameters of cleaning machines. Tech. Sci. Univ. Warm. Mazury Olszt. 2013, 16, 63–72. [Google Scholar]
- Orlov, A.Y.; Koshelkov, S.P. Soil ecology of Scots Pine; ‘Nauka’ Publ.: Moscow, Russia, 1971; p. 324. (In Russian) [Google Scholar]
- Dostovalov, B.N.; Kudryavtsev, V.A. Basics of Geocryology; Obshcheye Merzlotovedeniya (General Permafrost Science): Moscow, Russia, 1967; p. 404. (In Russian) [Google Scholar]
- Sannikov, S.N.; Petrova, I.V.; Sannikova, N.S.; Afonin, A.N.; Chernodubov, A.I.; Egorov, E.V. Genetic-climatologic-geographical principles of seed zoning of pine forests in Russia. Sib. Lesn. Zhurnal (Sib. J. For. Sci.) 2017, 2, 19–30, (In Russian with English abstracts). [Google Scholar]
- Shmueli, G. To Explain or to Predict? Stat. Sci. 2010, 25, 289–310. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GDD5, °C | AMI | NDD, °C | |||||
|---|---|---|---|---|---|---|---|
| −6000–−4850 | −4850–−3700 | −3700–−2550 | −2550–−1400 | −1400–−250 | >−250 | ||
| 600–1080 | 0.6–1.8 | 1 3.4–5.9 | 2 3.9–6.4 | 3 4.3–6.8 | 4 4.8–7.3 | 5 5.3–7.8 | 6 |
| 1.8–3.0 | 7 4.1–6.6 | 8 4.6–7.1 | 9 5.0–7.5 | 10 | 11 | 12 | |
| 3.0–4.2 | 13 | 14 | 15 | 16 | 17 | 18 | |
| 4.2–5.4 | 19 | 20 | 21 | 22 | 23 | 24 | |
| 5.4–6.6 | 25 | 26 | 27 | 28 | 29 | 30 | |
| 6.6–7.0 | 31 | 32 | 33 | 34 | 35 | 36 | |
| 1080–1560 | 0.6–1.8 | 37 | 38 | 39 5.7–8.2 | 40 6.1–8.6 | 41 6.6–9.1 | 42 |
| 1.8–3.0 | 43 | 44 5.9–8.4 | 45 6.4–8.9 | 46 6.8–9.3 | 47 7.3–9.8 | 48 | |
| 3.0–4.2 | 49 6.1–8.6 | 50 6.6–9.1 | 51 7.1–9.6 | 52 7.5–10.0 | 53 | 54 | |
| 4.2–5.4 | 55 | 56 | 57 | 58 | 59 | 60 | |
| 5.4–6.6 | 61 | 62 | 63 | 64 | 65 | 66 | |
| 6.6–7.0 | 67 | 68 | 69 | 70 | 71 | 72 | |
| 1560–2040 | 0.6–1.8 | 73 | 74 | 75 | 76 | 77 | 78 |
| 1.8–3.0 | 79 | 80 | 81 7.7–10.2 | 82 8.2–10.7 | 83 8.6–11.1 | 84 9.1–11.2 | |
| 3.0–4.2 | 85 | 86 7.9–10.4 | 87 8.4–10.9 | 88 8.9–11.4 | 89 9.3–11.8 | 90 | |
| 4.2–5.4 | 91 | 92 | 93 9.1–11.6 | 94 | 95 | 96 | |
| 5.4–6.6 | 97 | 98 | 99 | 100 | 101 | 102 | |
| 6.6–7.0 | 103 | 104 | 105 | 106 | 107 | 108 | |
| 2040–2520 | 0.6–1.8 | 109 | 110 | 111 | 112 | 113 | 114 |
| 1.8–3.0 | 115 | 116 | 117 | 118 9.5–12.0 | 119 9.9–12.4 | 120 10.4–12.5 | |
| 3.0–4.2 | 121 | 122 | 123 | 124 10.2–12.7 | 125 10.7–13.2 | 126 | |
| 4.2–5.4 | 127 | 128 | 129 | 130 | 131 11.4–13.9 | 132 | |
| 5.4–6.6 | 133 | 134 | 135 | 136 | 137 12.1–14.6 | 138 | |
| 6.6–7.0 | 139 | 140 | 141 | 142 | 143 | 144 | |
| 2520–3000 | 0.6–1.8 | 145 | 146 | 147 | 148 | 149 | 150 |
| 1.8–3.0 | 151 | 152 | 153 | 154 | 155 | 156 | |
| 3.0–4.2 | 157 | 158 | 159 | 160 | 161 | 162 | |
| 4.2–5.4 | 163 | 164 | 165 | 166 | 167 12.7–15.2 | 168 | |
| 5.4–6.6 | 169 | 170 | 171 | 172 | 173 13.4–15.9 | 174 13.9–16.0 | |
| 6.6–7.0 | 175 | 176 | 177 | 178 | 179 | 180 | |
| Current Climate Seed Zone, % of Each Mass Grade | Seed Zone, % of Total Area | Scenario RCP 2.6 Seed Zone, % of Each Mass Grade | Seed zone, | Scenario RCP 8.5 Seed Zone, % of Each Mass Grade | Seed Zone, % of Total Area | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Seed Zone | Seed Mass Grades | Seed Mass Grades | % of Total Area | Seed Mass Grades | ||||||||||||||
| <2 | 2–4 | 4–6 | 6–8 | 8–10 | 10–12 | 2–4 | 4–6 | 6–8 | 8–10 | 10–12 | 6–8 | 8–10 | 10–12 | >12 | ||||
| 1 | 52.9 | 47.1 | 3.36 | |||||||||||||||
| 2 | 16.9 | 82.3 | 0.8 | 6.75 | 96.6 | 3.4 | 5.43 | |||||||||||
| 3 | 2.1 | 87.9 | 10.0 | 7.78 | 51.8 | 48.2 | 4.59 | |||||||||||
| 4 | 39.6 | 60.4 | 6.53 | 3.4 | 96.6 | 1.24 | ||||||||||||
| 5 | 9.8 | 90.2 | 1.99 | |||||||||||||||
| 7 | 12.8 | 87.2 | 5.22 | |||||||||||||||
| 8 | 1.8 | 87.6 | 10.6 | 2.36 | 62.4 | 37.6 | 4.61 | |||||||||||
| 9 | 36.0 | 64.0 | 1.89 | |||||||||||||||
| 39 | 28.1 | 71.9 | 1.88 | 98.7 | 1.3 | 3.29 | 98.9 | 1.2 | 1.22 | |||||||||
| 40 | 0.6 | 99.4 | 2.29 | 88.2 | 11.8 | 2.60 | ||||||||||||
| 41 | 100 | 1.74 | ||||||||||||||||
| 44 | 28.8 | 71.2 | 5.97 | 100 | 3.01 | 97.4 | 2.6 | 2.17 | ||||||||||
| 45 | 0.4 | 98.8 | 0.7 | 13.06 | 89.2 | 10.8 | 11.10 | 68.7 | 31.3 | 5.43 | ||||||||
| 46 | 76.3 | 23.7 | 11.29 | 35.3 | 64.7 | 15.66 | ||||||||||||
| 47 | 24.5 | 75.5 | 3.74 | 16.7 | 83.3 | 8.14 | ||||||||||||
| 49 | 50.9 | 49.1 | 3.19 | |||||||||||||||
| 50 | 3.9 | 96.1 | 2.47 | 82.5 | 17.5 | 3.36 | ||||||||||||
| 51 | 80.2 | 19.8 | 2.25 | 19.6 | 80.4 | 2.07 | ||||||||||||
| 52 | 4.8 | 95.2 | 1.74 | |||||||||||||||
| 81 | 30.8 | 69.2 | 1.08 | 100 | 1.17 | 100 | 4.97 | |||||||||||
| 82 | 100 | 1.15 | 97.4 | 2.6 | 8.21 | 78.9 | 21.1 | 12.08 | ||||||||||
| 83 | 98.8 | 1.2 | 1.18 | 76.5 | 23.5 | 9.22 | 26.5 | 73.5 | 7.79 | |||||||||
| 84 | 10.3 | 89.7 | 4.62 | |||||||||||||||
| 86 | 100 | 1.23 | ||||||||||||||||
| 87 | 80.1 | 19.9 | 7.27 | |||||||||||||||
| 88 | 92.6 | 7.4 | 2.25 | 55.6 | 44.4 | 3.97 | 35.9 | 64.1 | 3.52 | |||||||||
| 89 | 0.6 | 99.4 | 2.35 | |||||||||||||||
| 93 | 19.8 | 80.2 | 1.21 | |||||||||||||||
| 118 | 4.5 | 95.5 | 4.38 | |||||||||||||||
| 119 | 100 | 7.34 | ||||||||||||||||
| 120 | 93.8 | 6.2 | 4.06 | |||||||||||||||
| 124 | 100 | 3.13 | ||||||||||||||||
| 125 | 92.4 | 7.6 | 1.29 | 70.7 | 29.3 | 12.88 | ||||||||||||
| 126 | 9.0 | 91.0 | 1.71 | |||||||||||||||
| 131 | 18.3 | 81.7 | 1.30 | 100 | 2.15 | |||||||||||||
| 137 | 55.1 | 44.9 | 1.08 | |||||||||||||||
| 167 | 100 | 1.16 | ||||||||||||||||
| 173 | 100 | 1.05 | ||||||||||||||||
| 174 | 100 | 0.98 | ||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parfenova, E.I.; Kuzmina, N.A.; Kuzmin, S.R.; Tchebakova, N.M. Climate Warming Impacts on Distributions of Scots Pine (Pinus sylvestris L.) Seed Zones and Seed Mass across Russia in the 21st Century. Forests 2021, 12, 1097. https://doi.org/10.3390/f12081097
Parfenova EI, Kuzmina NA, Kuzmin SR, Tchebakova NM. Climate Warming Impacts on Distributions of Scots Pine (Pinus sylvestris L.) Seed Zones and Seed Mass across Russia in the 21st Century. Forests. 2021; 12(8):1097. https://doi.org/10.3390/f12081097
Chicago/Turabian StyleParfenova, Elena I., Nina A. Kuzmina, Sergey R. Kuzmin, and Nadezhda M. Tchebakova. 2021. "Climate Warming Impacts on Distributions of Scots Pine (Pinus sylvestris L.) Seed Zones and Seed Mass across Russia in the 21st Century" Forests 12, no. 8: 1097. https://doi.org/10.3390/f12081097
APA StyleParfenova, E. I., Kuzmina, N. A., Kuzmin, S. R., & Tchebakova, N. M. (2021). Climate Warming Impacts on Distributions of Scots Pine (Pinus sylvestris L.) Seed Zones and Seed Mass across Russia in the 21st Century. Forests, 12(8), 1097. https://doi.org/10.3390/f12081097