
When Density Matters: The Spatial Balance between Early and Latewood

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
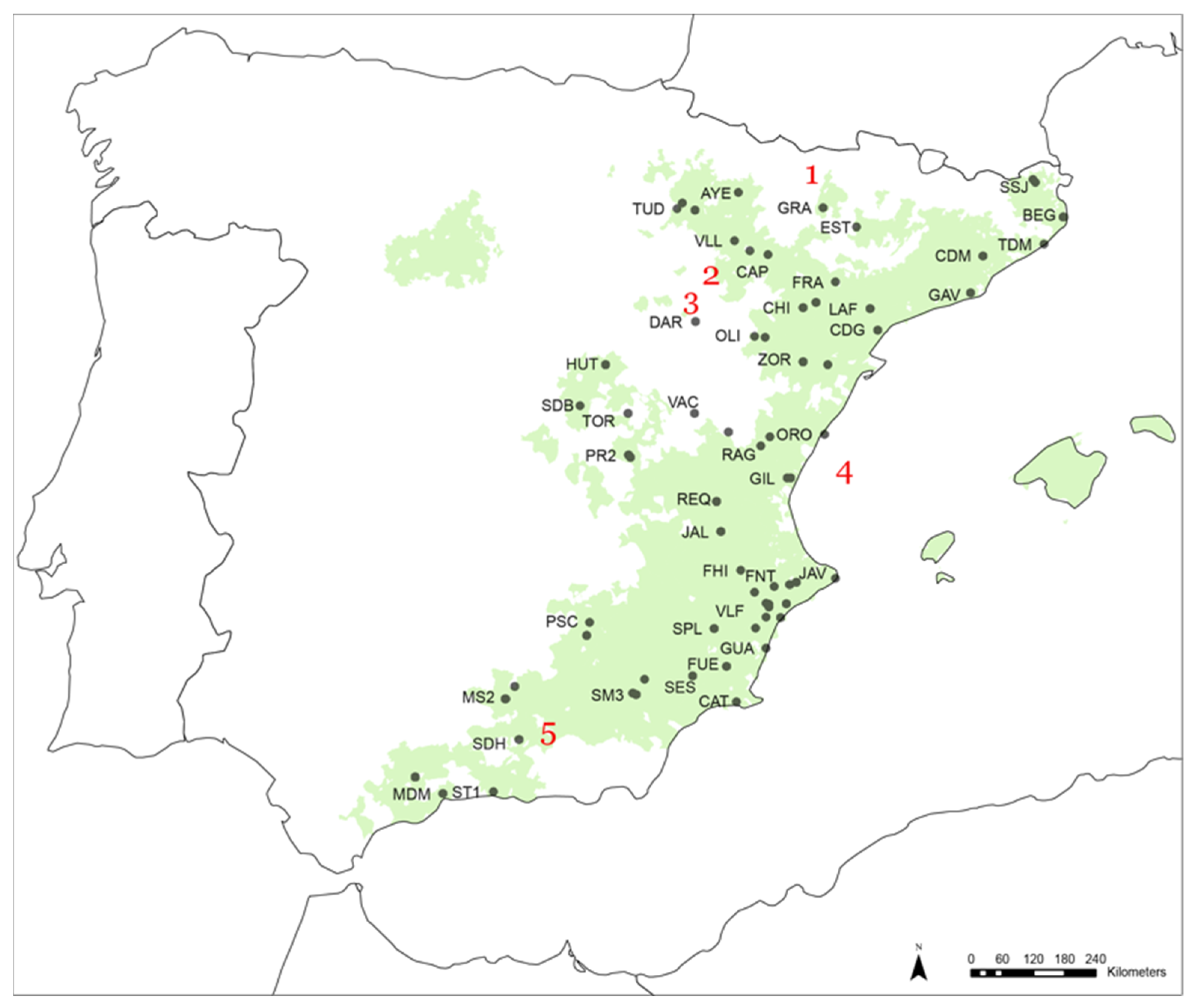
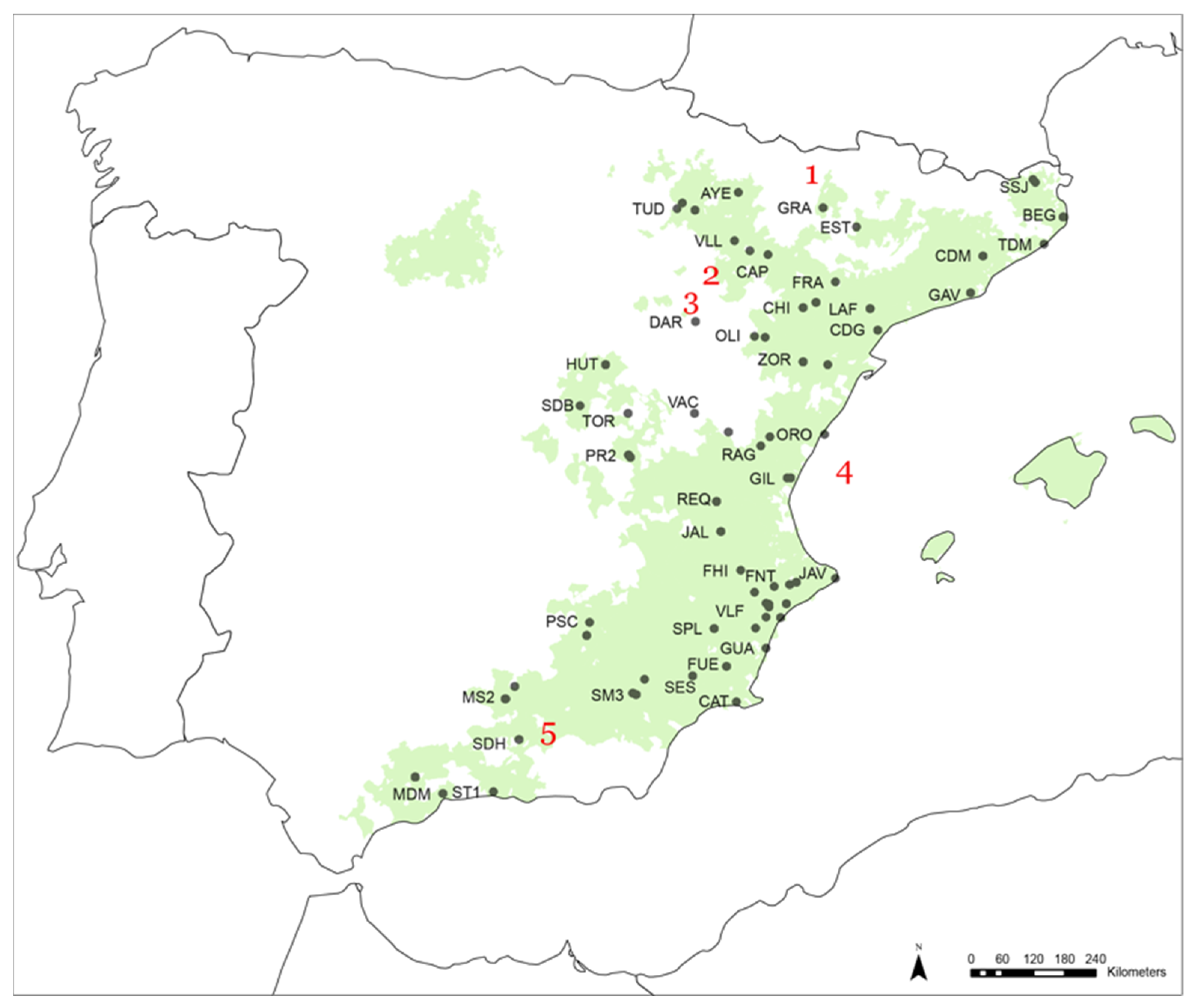
2.1.1. Dendroclimatic Dataset
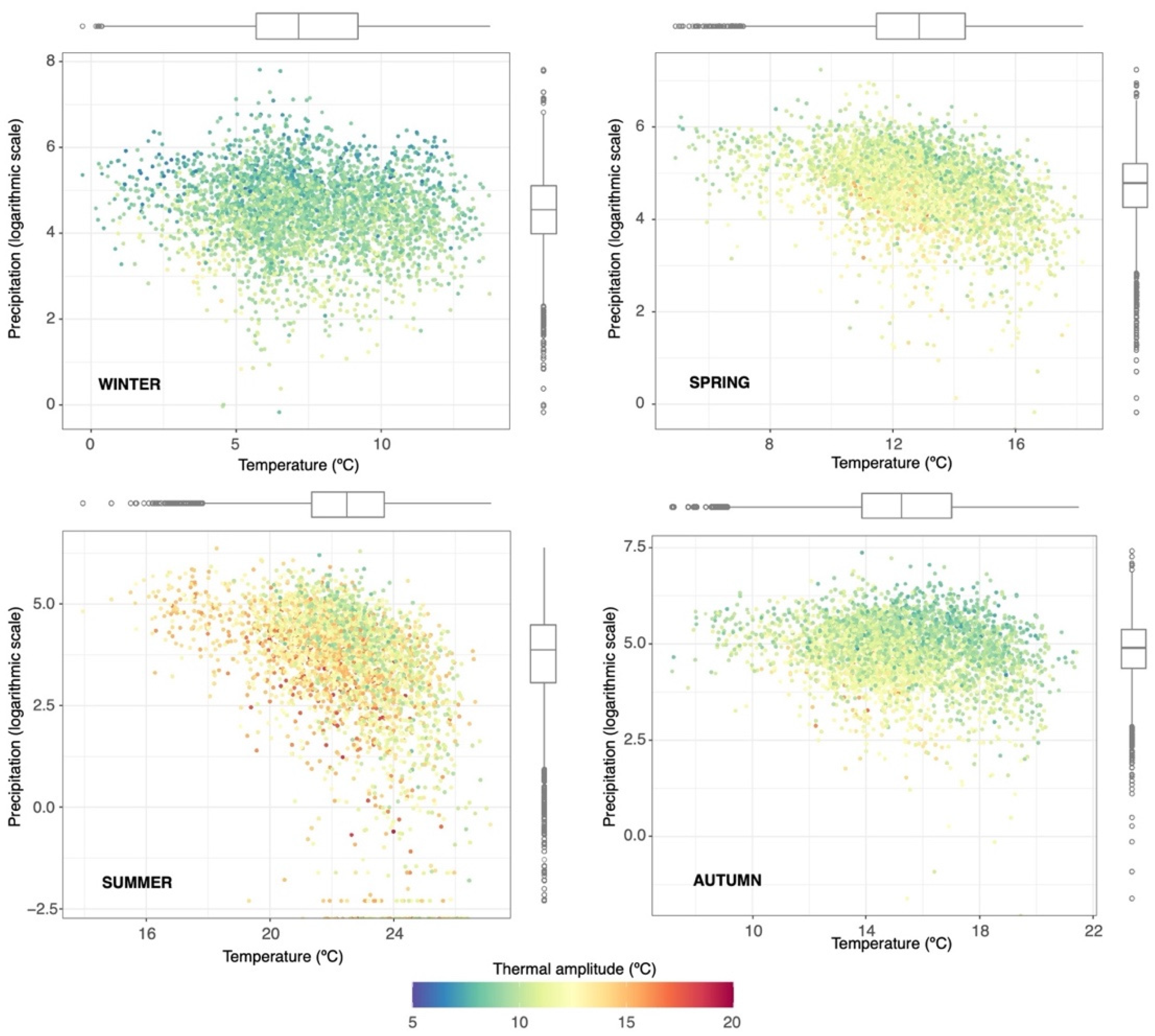
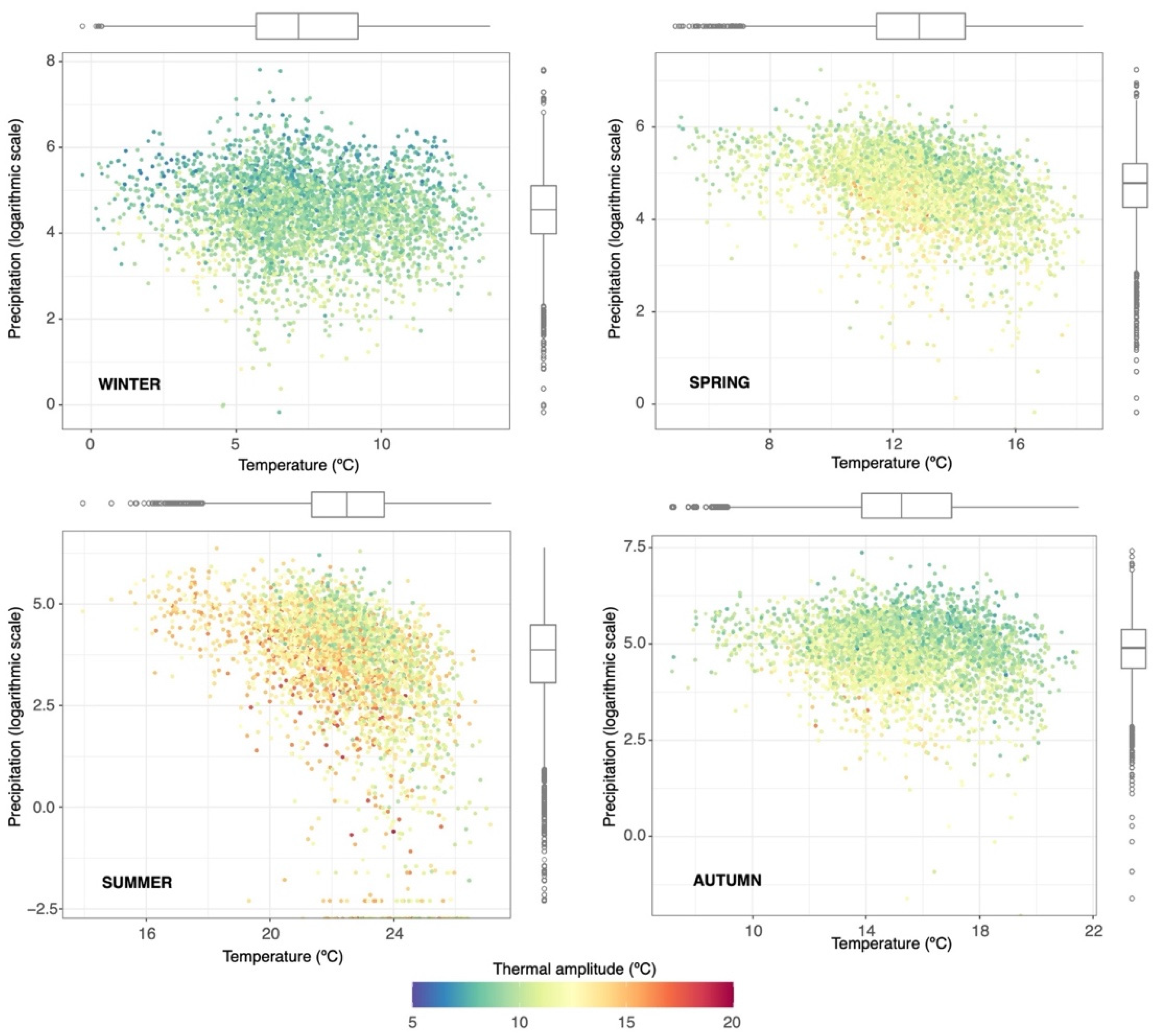
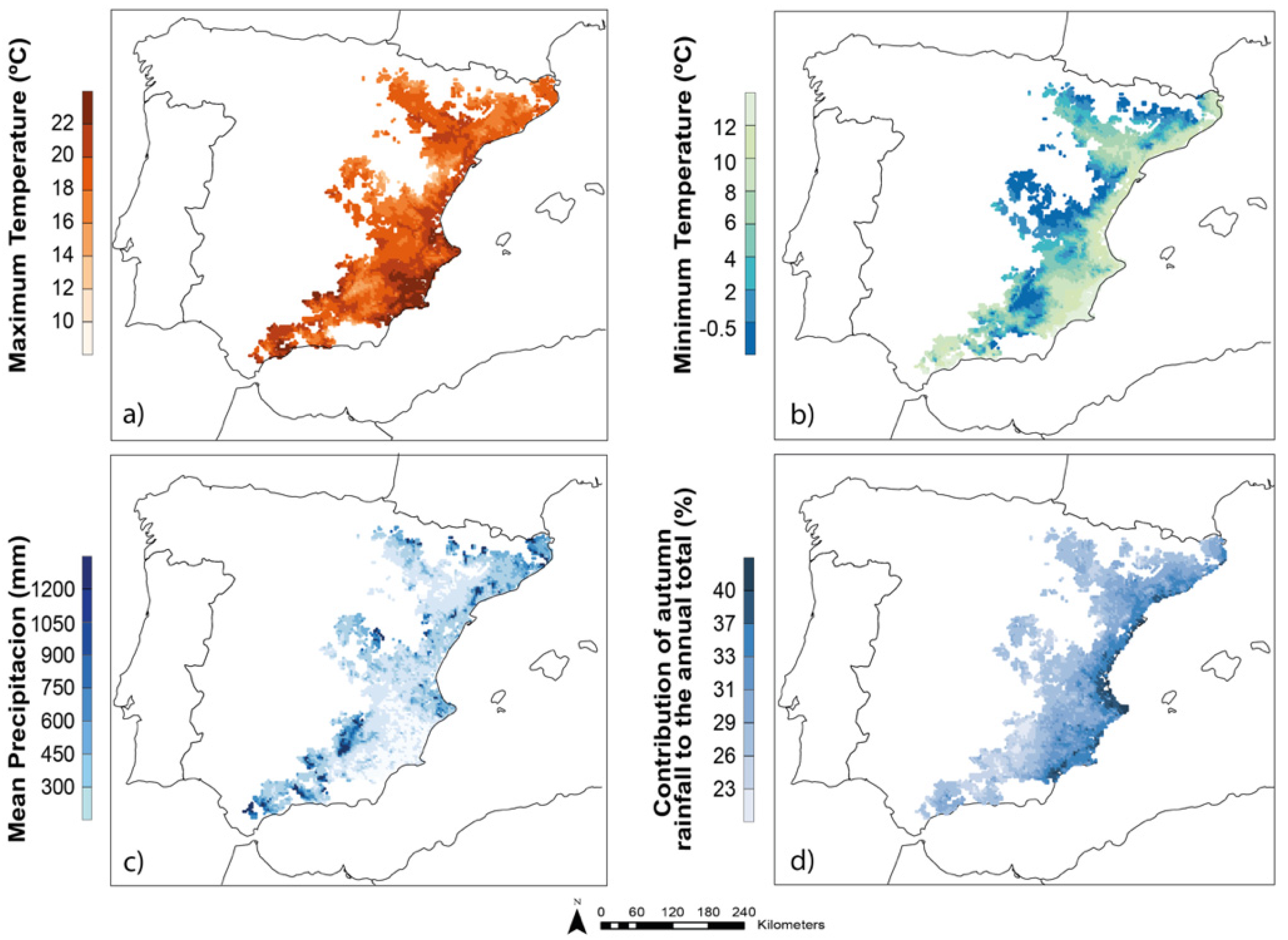
2.1.2. Climatic Domain and Sources
2.1.3. Dendrochronological Methods
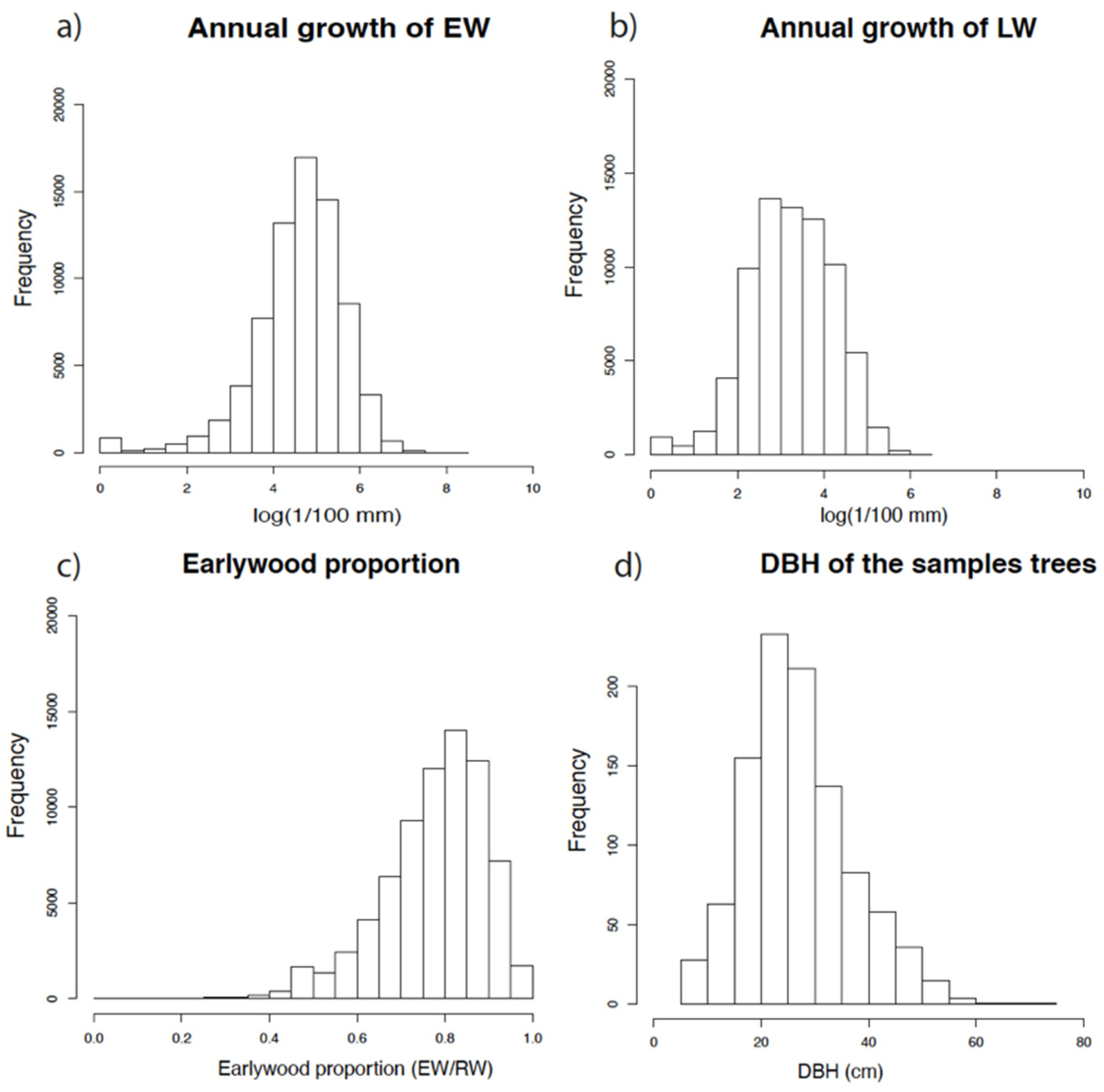
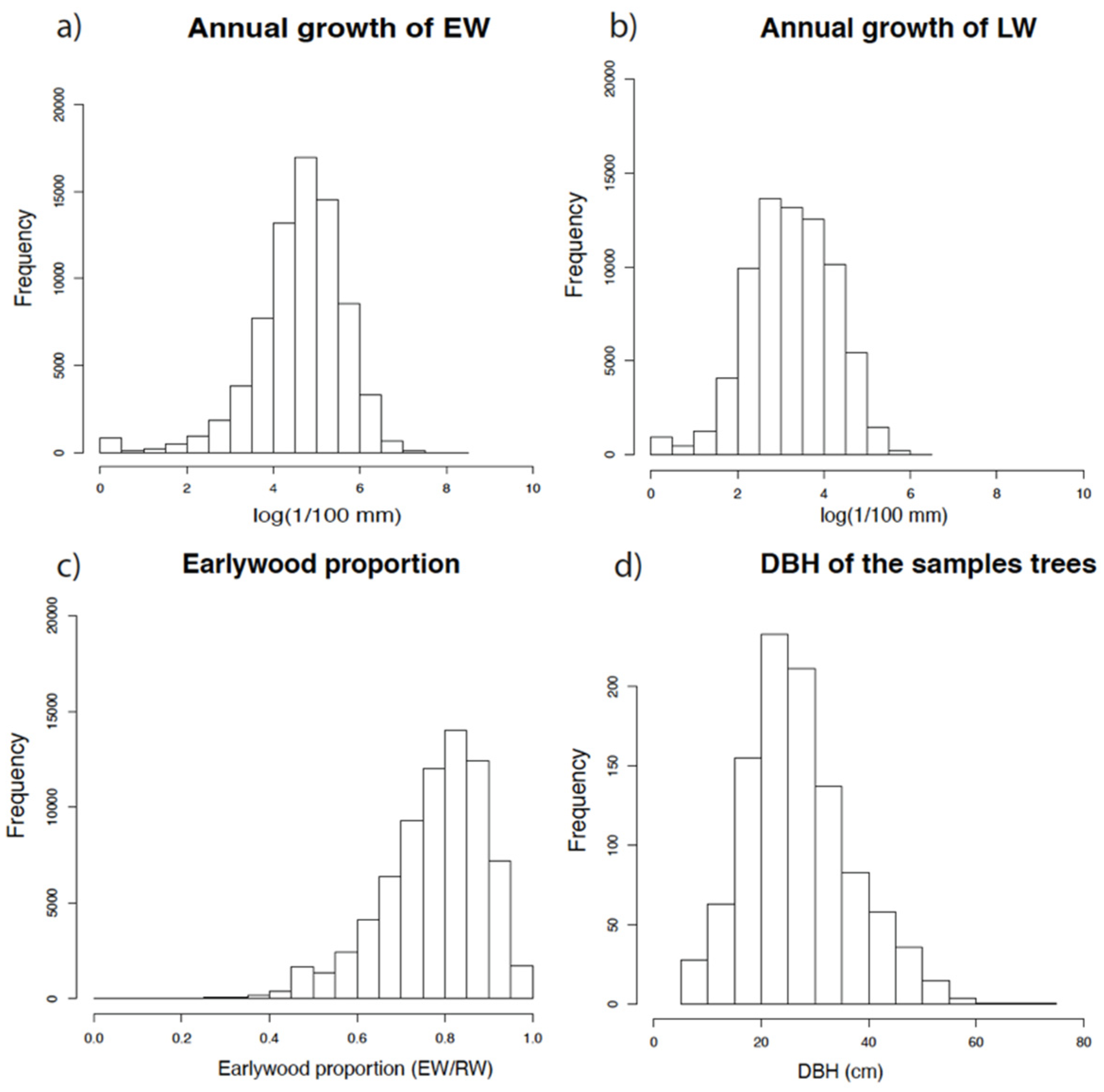
2.1.4. Dendrochronological Dataset
2.2. Statistical Analysis
Models
3. Results
3.1. Main Factors Controlling EW and LW Growth in Pinus Halepensis
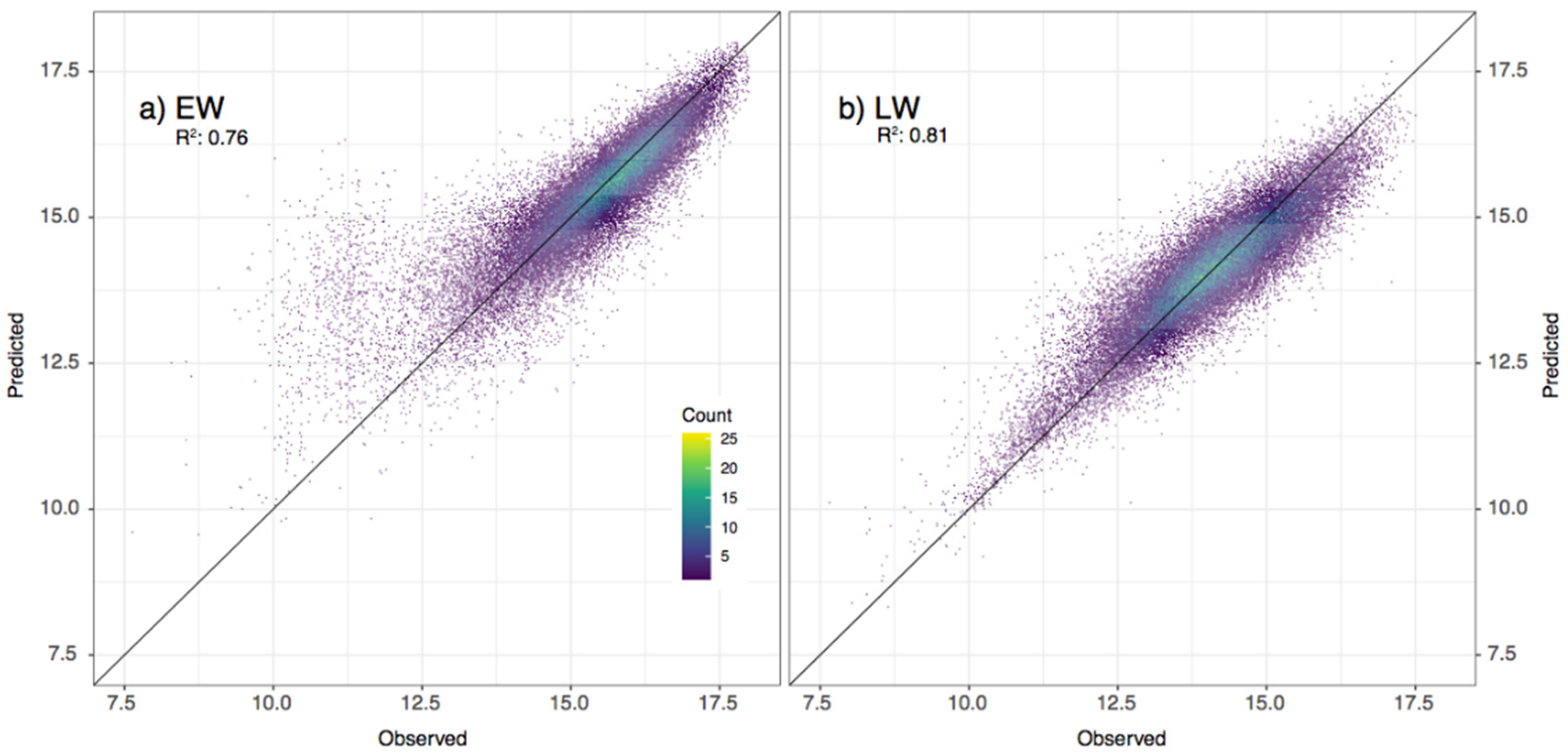
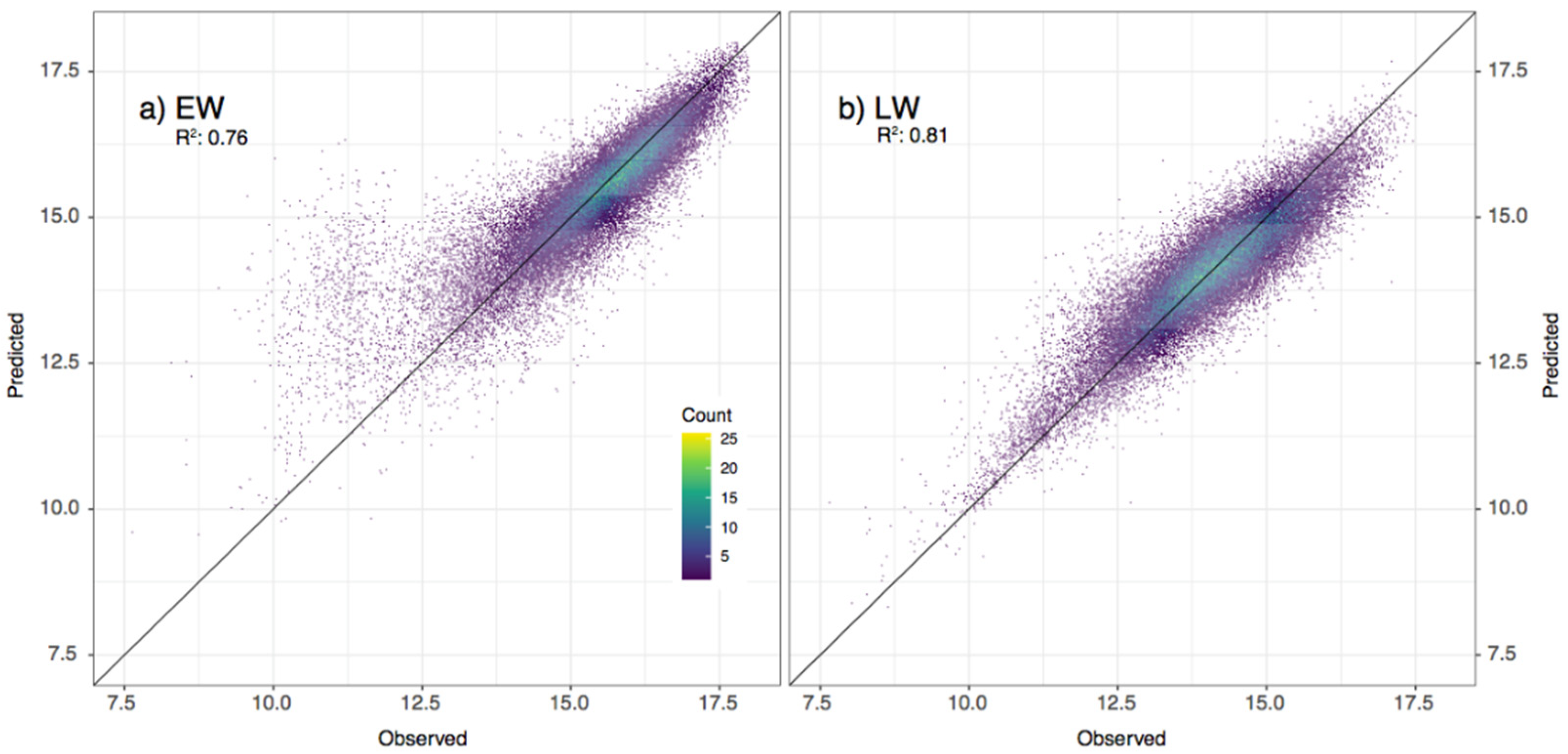
3.2. Models’ Accuracy Assessment and Validation
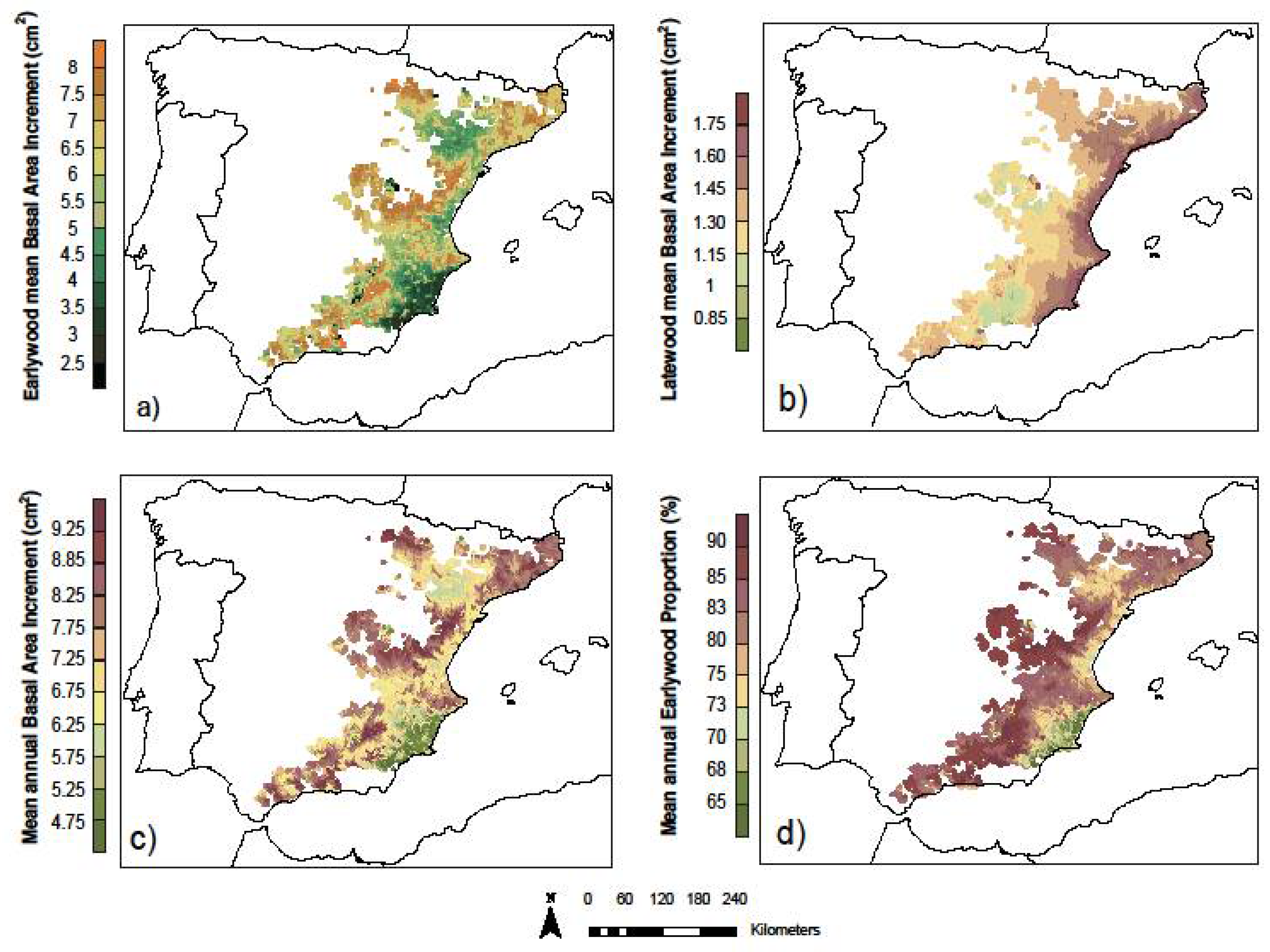
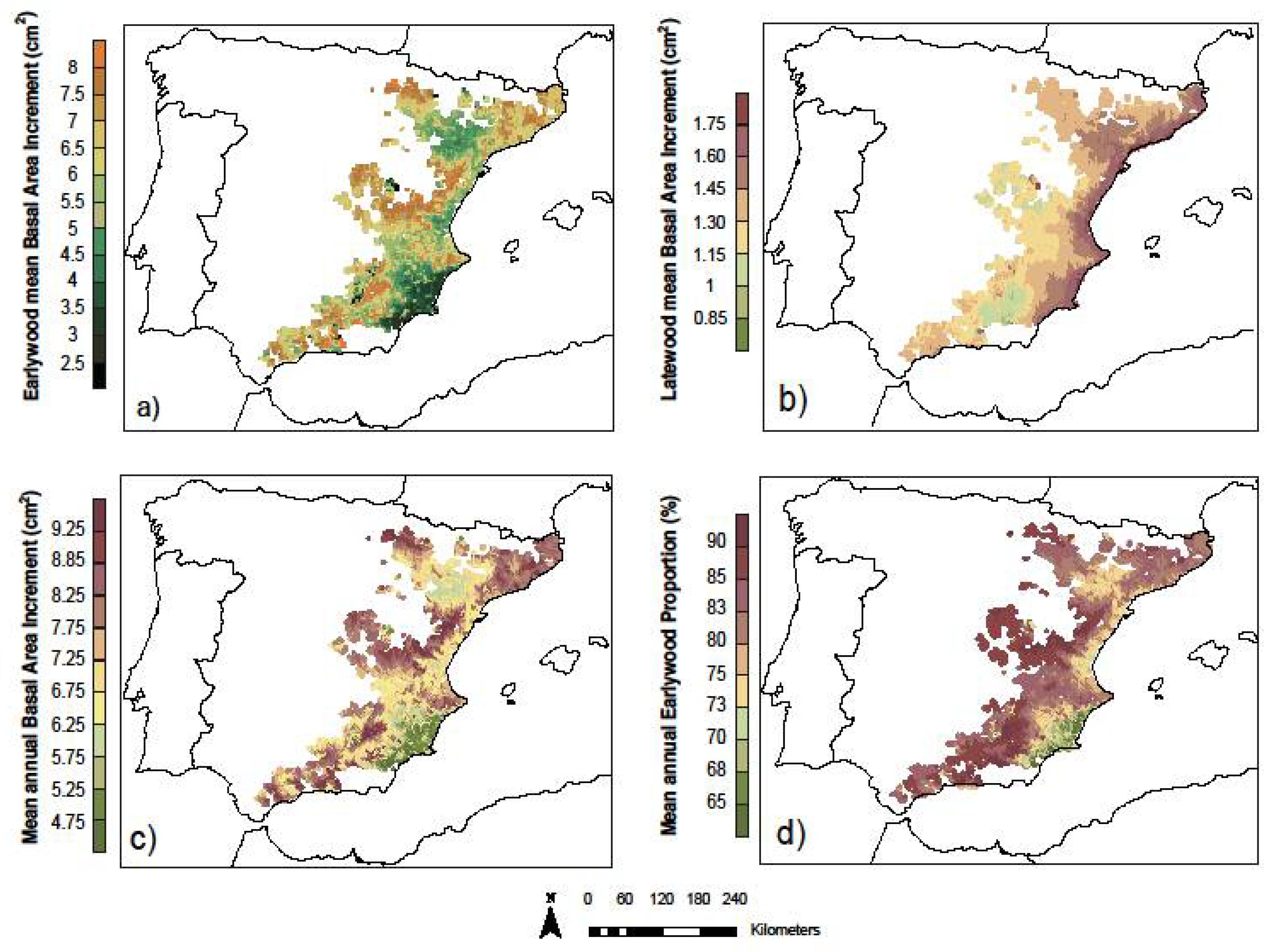
3.3. Models’ Predictions across Species Distribution
4. Discussion
4.1. Tree Acclimation and Spatial Plasticity in Secondary Growth
4.2. Differing Anatomical Strategies
4.3. Effects on the Carbon Balance
5. Conclusions
- -
- in areas with wetter conditions, the trees contained a higher proportion of EW (i.e., water transport efficiency) and a lower response to climatic variations. This fact allowed the trees to maintain a stable growth rate (i.e., an efficiency strategy).
- -
- On the contrary, in areas where climatic conditions were more xeric, the proportion of LW was higher and had a faster response to the climatic variations (i.e., a safety strategy).
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution os species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 361–371. [Google Scholar] [CrossRef] [Green Version]
- De Luis, M.; Cufar, K.; Di Filippo, A.; Novak, K.; Papadopoulos, A.; Piovesan, G.; Rathgeber, C.; Raventós, J.; Saz, M.; Smith, K. Plasticity in dendroclimatic response across the distribution range of Aleppo pine (Pinus halepensis). PLoS ONE 2013, 8, e83550. [Google Scholar] [CrossRef] [Green Version]
- Benito Garzón, M.; Alía, R.; Robson, T.M.; Zavala, M.A. Intra-specific variability and plasticity influence potential tree species distributions under climate change. Glob. Ecol. Biogeogr. 2011, 20, 766–778. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Sanchez-Gomez, D.; Zavala, M.A. Quantitative estimation of phenotypic plasticity: Bridging the gap between the evolutionary concept and its ecological applications. J. Ecol. 2006, 94, 1103–1116. [Google Scholar] [CrossRef]
- Matesanz, S.; Gianoli, E.; Valladares, F. Global change and the evolution of phenotypic plasticity in plants. Ann. N. Y. Acad. Sci. 2010, 1206, 35–55. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Cochard, H.; Mencuccini, M.; Sterck, F.; Herrero, A.; Korhonen, J.F.J.; Llorens, P.; Nikinmaa, E.; Nolè, A.; Poyatos, R.; et al. Hydraulic adjustment of Scots pine across Europe. New Phytol. 2009, 184, 353–364. [Google Scholar] [CrossRef]
- Camarero, J.J.; Collado, E.; Martínez-de-Aragón, J.; de-Miguel, S.; Büntgen, U.; Martinez-Peña, F.; Martín-Pinto, P.; Ohenoja, E.; Romppanen, T.; Salo, K.; et al. Associations between climate and earlywood and latewood width in boreal and Mediterranean Scots pine forests. Trees Struct. Funct. 2021, 35, 155–169. [Google Scholar] [CrossRef]
- Voltas, J.; Lucabaugh, D.; Chambel, M.R.; Ferrio, J.P. Intraspecific variation in the use of water sources by the circum-Mediterranean conifer Pinus halepensis. New Phytol. 2015, 208, 1031–1041. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.M.; Rundel, P.W. Ecology and Biogeography of Pinus: An Introduction; Cambridge University Press: Cambridge, UK, 1998; pp. 3–46. [Google Scholar]
- Taïbi, K.; del Campo, A.D.; Aguado, A.; Mulet, J.M. The effect of genotype by environment interaction, phenotypic plasticity and adaptation on Pinus halepensis reforestation establishment under expected climate drifts. Ecol. Eng. 2015, 84, 218–228. [Google Scholar] [CrossRef]
- Martínez del Castillo, E.; Tejedor, E.; Serrano-Notivoli, R.; Novak, K.; Saz, M.Á.; Longares, L.A.; de Luis, M. Contrasting patterns of tree growth of Mediterranean pine species in the Iberian Peninsula. Forests 2018, 9, 416. [Google Scholar] [CrossRef] [Green Version]
- Zalloni, E.; de Luis, M.; Campelo, F.; Novak, K.; De Micco, V.; Di Filippo, A.; Vieira, J.; Nabais, C.; Rozas, V.; Battipaglia, G. Climatic signals from intra-annual density fluctuation frequency in mediterranean pines at a regional scale. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Voltas, J.; Chambel, M.R.; Prada, M.A.; Ferrio, J.P. Climate-related variability in carbon and oxygen stable isotopes among populations of Aleppo pine grown in common-garden tests. Trees Struct. Funct. 2008, 22, 759–769. [Google Scholar] [CrossRef]
- Liphschitz, N.; Lev-Yadun, S.; Rosen, E.; Waisel, Y. The annual rhythm of activity of the lateral meristems (cambium and phellogen) in Pinus halepensis Mill. and Pinus pinea L. IAWA J. 1984, 5, 263–274. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; de Luis, M.; Novak, K.; Martinez Del Castillo, E.; Schmitt, U.; Mrak, P.; Koch, G.; Štrus, J.; Žnidarič, M.T.; et al. Annual cambial rhythm in Pinus halepensis and Pinus sylvestris as indicator for climate adaptation. Front. Plant Sci. 2016, 7, 1923. [Google Scholar] [CrossRef] [Green Version]
- Cherubini, P.; Gartner, B.L.; Tognetti, R.; Bräker, O.U.; Schoch, W.; Innes, J.L. Identification, measurement and interpretation of tree rings in woody species from mediterranean climates. Biol. Rev. Camb. Philos. Soc. 2003, 78, 119–148. [Google Scholar] [CrossRef] [Green Version]
- De Luis, M.; Gričar, J.; Čufar, K.; Raventós, J. Seasonal dynamics of wood formation in Pinus halepensis from dryand semi-arid ecosystems in Spain. IAWA J. 2007, 28, 389–404. [Google Scholar] [CrossRef] [Green Version]
- Novak, K.; de Luis, M.; Raventós, J.; Čufar, K. Climatic signals in tree-ring widths and wood structure of Pinus halepensis in contrasted environmental conditions. Trees Struct. Funct. 2013, 27, 927–936. [Google Scholar] [CrossRef]
- Novak, K.; De Luis, M.; Gričar, J.; Prislan, P.; Merela, M.; Smith, K.T.; Čufar, K. Missing and dark ringd associated with drought in Pinus halepensis. IAWA J. 2016, 37, 260–274. [Google Scholar] [CrossRef]
- Novak, K.; Čufar, K.; De Luis, M.; Sánchez, M.A.S.; Raventós, J. Age, climate and intra-annual density fluctuations in Pinus halepensis in Spain. IAWA J. 2013, 34, 459–474. [Google Scholar] [CrossRef] [Green Version]
- Antony, F.; Schimleck, L.R.; Daniels, R.F. A comparison of earlywood-latewood demarcation methods—A case study in loblolly pine. IAWA J. 2012, 33, 187–195. [Google Scholar] [CrossRef]
- Babushkina, E.A.; Belokopytova, L.V.; Kostyakova, T.V.; Kokova, V.I. Earlywood and Latewood Features of Pinus sylvestris in Semiarid Natural Zones of South Siberia. Russ. J. Ecol. 2018, 49, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Domec, J.-C.; Gartner, B.L. How do water transport and water storage differ in coniferous earlywood and latewood? J. Exp. Bot. 2002, 53, 2369–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, D.M. Ecology and biogeography of Pinus. Ecol. Biogeogr. Pinus 1998. [Google Scholar] [CrossRef]
- Keeley, J.E.; Ne’eman, G.; Trabaud, L. Ecology, Biogeography and Management of Pinus halepensis and P. brutia Forest Ecosystems in the Mediterranean Basin. J. Veg. Sci. 2001, 12, 151. [Google Scholar] [CrossRef]
- Olivar, J.; Bogino, S.; Spiecker, H.; Bravo, F. Climate impact on growth dynamic and intra-annual density fluctuations in Aleppo pine (Pinus halepensis) trees of different crown classes. Dendrochronologia 2012, 30, 35–47. [Google Scholar] [CrossRef]
- Papadopoulos, A.; Serre-Bachet, F.; Tessier, L. Tree ring to climate relationships of Aleppo pine (Pinus halepensis Mill.) in Greece. Ecol. Mediterr. 2001, 27, 89–98. [Google Scholar] [CrossRef]
- Pasho, E.; Camarero, J.J.; de Luis, M.; Vicente-Serrano, S.M. Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain. Agric. For. Meteorol. 2011, 151, 1800–1811. [Google Scholar] [CrossRef]
- González-González, B.D.; Vázquez-Ruiz, R.A.; García-González, I. Effects of climate on earlywood vessel formation of quercus robur and q. Pyrenaica at a site in the northwestern iberian peninsula. Can. J. For. Res. 2015, 45, 698–709. [Google Scholar] [CrossRef]
- Souto-Herrero, M.; Rozas, V.; García-González, I. Earlywood vessels and latewood width explain the role of climate on wood formation of Quercus pyrenaica Willd. across the Atlantic-Mediterranean boundary in NW Iberia. For. Ecol. Manage. 2018, 425, 126–137. [Google Scholar] [CrossRef]
- Serrano-Notivoli, R.; Beguería, S.; De Luis, M. STEAD: A high-resolution daily gridded temperature dataset for Spain. Earth Syst. Sci. Data Discuss 2019, 1171–1188. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Notivoli, R.; Beguería, S.; Saz, M.Á.; Longares, L.A.; De Luis, M. SPREAD: A high-resolution daily gridded precipitation dataset for Spain—An extreme events frequency and intensity overview. Earth Syst. Sci. Data 2017, 9, 721–738. [Google Scholar] [CrossRef] [Green Version]
- Larsson, L.A. CoRecorder&CDendro Program; Cybis Elektron; Data AB; Version 7.6; 2012. [Google Scholar]
- Holmes, R. Computer-Assisted Quality Control in Tree-Ring Dating and Measurement. Tree Ring Bull. 1983, 43, 51–67. [Google Scholar]
- Pallardy, S.G. Physiology of Woody Plants, 3rd ed.; Elsevier: Amsterdam, The Netherlands; Boston, MA, USA, 2008; ISBN 9780120887651. [Google Scholar]
- Biondi, F.; Qeadan, F. Removing the Tree-Ring Width Biological Trend Using Expected Basal Area Increment. In Fort Valley Experimental Forest—A Century of Research 1908–2008; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2008; pp. 124–131. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Martínez del Castillo, E.; Longares, L.A.; Serrano-Notivoli, R.; de Luis, M. Modeling tree-growth: Assessing climate suitability of temperate forests growing in Moncayo Natural Park (Spain). For. Ecol. Manag. 2019, 435, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Tejedor, E.; Serrano-Notivoli, R.; de Luis, M.; Saz, M.A.; Hartl, C.; St George, S.; Büntgen, U.; Liebhold, A.M.; Vuille, M.; Esper, J. A global perspective on the climate-driven growth synchrony of neighbouring trees. Glob. Ecol. Biogeogr. 2020, 29, 1114–1125. [Google Scholar] [CrossRef]
- Barbéro, M.; Loisel, R.; Quézel, P.; Richardson, D.; Romane, F. Pines of the Mediterranean Basin. In Ecology and Biogeography of Pinus; Cambridge University Press: Cambridge, UK, 1998; pp. 153–170. [Google Scholar]
- Quézel, P. Taxonomy and biogeography of Mediterranean pines (Pinus halepensis and P. brutia). In Ecology, Biogeography and Management of Pinus Halepensis and P. Brutia Forest Ecosystems in the Mediterranean Basin; Backhuys Publishers: Leiden, The Netherlands, 2000; pp. 1–12. [Google Scholar]
- De Luis, M.; Novak, K.; Čufar, K.; Raventós, J. Size mediated climate-growth relationships in Pinus halepensis and pinus pinea. Trees Struct. Funct. 2009, 23, 1065–1073. [Google Scholar] [CrossRef]
- Zobel, B.J.; van Buijtenen, J.P. Wood Variation and Wood Properties. In Wood Variation; Springer: Berlin/Heidelberg, Germany, 1989; pp. 1–32. [Google Scholar] [CrossRef]
- Bannan, M.W. Sequential Changes in Rate of Anticlinal Division, Cambial Cell Length, and Ring Width in the Growth of Coniferous Trees. Can. J. Bot. 1967, 45, 1359–1369. [Google Scholar] [CrossRef]
- Lebourgeois, F. Climatic signals in earlywood, latewood and total ring width of Corsican pine from western France. Ann. For. Sci. 2000, 57, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Kagawa, A.; Sugimoto, A.; Maximov, T.C. 13CO2 pulse-labelling of photoassimilates reveals carbon allocation within and between tree rings. Plant Cell Environ. 2006, 29, 1571–1584. [Google Scholar] [CrossRef]
- Kress, A.; Young, G.H.F.; Saurer, M.; Loader, N.J.; Siegwolf, R.T.W.; McCarroll, D. Stable isotope coherence in the earlywood and latewood of tree-line conifers. Chem. Geol. 2009, 268, 52–57. [Google Scholar] [CrossRef]
- Oberhuber, W.; Swidrak, I.; Pirkebner, D.; Gruber, A. Temporal dynamics of nonstructural carbohydrates and xylem growth in pinus sylvestris exposed to drought. Can. J. For. Res. 2011, 41, 1590–1597. [Google Scholar] [CrossRef] [Green Version]
- Manrique-Alba, À.; Beguería, S.; Molina, A.J.; González-Sanchis, M.; Tomàs-Burguera, M.; del Campo, A.D.; Colangelo, M.; Camarero, J.J. Long-term thinning effects on tree growth, drought response and water use efficiency at two Aleppo pine plantations in Spain. Sci. Total Environ. 2020, 728. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Fernández, J.; Almendra-Martín, L.; de Luis, M.; González-Zamora, A.; Herrero-Jiménez, C. Tracking tree growth through satellite soil moisture monitoring: A case study of Pinus halepensis in Spain. Remote Sens. Environ. 2019, 235, 111422. [Google Scholar] [CrossRef]
- De Luis, M.; Novak, K.; Raventós, J.; Gričar, J.; Prislan, P.; Čufar, K. Cambial activity, wood formation and sapling survival of Pinus halepensis exposed to different irrigation regimes. For. Ecol. Manag. 2011, 262, 1630–1638. [Google Scholar] [CrossRef]
- de Luis, M.; Brunetti, M.; Gonzalez-Hidalgo, J.C.; Longares, L.A.; Martin-Vide, J. Changes in seasonal precipitation in the Iberian Peninsula during 1946–2005. Glob. Planet. Chang. 2010, 74, 27–33. [Google Scholar] [CrossRef]
- Zweifel, R.; Sterck, F. A Conceptual Tree Model Explaining Legacy Effects on Stem Growth. Front. For. Glob. Chang. 2018, 1, 1–9. [Google Scholar] [CrossRef]
- Sperry, J.; Saliendra, N.Z. Intra- and inter-plant variation in xylem cavitation in Betula occidentalis. Plant Cell Environ. 1994, 17, 1233–1241. [Google Scholar] [CrossRef]
- Hargrave, K.R.; Kolb, K.J.; Ewers, F.W.; Davis, S.D. Conduit diameter and drought-induced embolism in Salvia mellifera Greene (Labiatae). New Phytol. 1994, 126, 695–705. [Google Scholar] [CrossRef]
- Schulte, P.J.; Gibson, A.C. Hydraulic conductance and tracheid anatomy in six species of extant seed plants. Can. J. Bot. 1988, 1073–1079. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Pockman, W.T.; Davis, S.D.; McCulloh, K.A. Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 2001, 126, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Cuny, H.E.; Rathgeber, C.B.K.; Frank, D.; Fonti, P.; Makinen, H.; Prislan, P.; Rossi, S.; Del Castillo, E.M.; Campelo, F.; Vavrčík, H.; et al. Woody biomass production lags stem-girth increase by over one month in coniferous forests. Nat. Plants 2015, 1, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lutz, J. How growth rate affects properties of softwood veneer. For. Prod. J. 1964, 24, 94–102. [Google Scholar]
- Takjima, T. Tree Growth and Wood Properties; Research Report; Faculty of Agriculture, Tokyo University: Tokyo, Japan, 1967. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Earlywood BAI | Latewood BAI | |||||
|---|---|---|---|---|---|---|
| Estimate | Std. Error | sig | Estimate | Std. Error | sig | |
| (Intercept) | 2.75 × 100 | 6.46 × 10−4 | *** | 2.65 × 100 | 6.65 × 10−4 | *** |
| Aridity gradient (AI) | −1.19 × 10−2 | 7.06 × 10−4 | *** | −3.96 × 10−3 | 7.04 × 10−4 | *** |
| Earlywood proportion (Ep) | 2.96 × 10−3 | 4.70 × 10−4 | *** | −6.66 × 10−3 | 4.37 × 10−4 | *** |
| AI: Ep | 3.70 × 10−3 | 5.89 × 10−4 | *** | 1.21 × 10−3 | 5.26 × 10−4 | * |
| PP Autumn-1 | 1.12 × 10−2 | 2.52 × 10−4 | *** | |||
| PP Win | 1.25 × 10−2 | 2.68 × 10−4 | *** | 7.89 × 10−4 | 2.43 × 10−4 | ** |
| PP Spr | 1.19 × 10−2 | 2.72 × 10−4 | *** | 1.50 × 10−3 | 2.52 × 10−4 | *** |
| PP Summer | 3.86 × 10−3 | 2.59 × 10−4 | *** | 5.70 × 10−3 | 2.36 × 10−4 | *** |
| PP Autumn | 6.56 × 10−3 | 2.29 × 10−4 | *** | |||
| Tmax Autumn-1 | −9.37 × 10−4 | 6.39 × 10−4 | ||||
| Tmax Win | 1.84 × 10−4 | 7.08 × 10−4 | −3.77 × 10−3 | 6.47 × 10−4 | *** | |
| Tmax Spr | −8.18 × 10−3 | 6.50 × 10−4 | *** | 4.74 × 10−3 | 5.55 × 10−4 | *** |
| Tmax Summer | 3.39 × 10−4 | 5.37 × 10−4 | −3.89 × 10−3 | 5.04 × 10−4 | *** | |
| Tmax Autumn | −1.27 × 10−3 | 5.98 × 10−4 | * | |||
| Tmin Autumn-1 | −7.21 × 10−3 | 9.58 × 10−4 | *** | |||
| Tmin Win | 7.27 × 10−3 | 6.69 × 10−4 | *** | 2.44 × 10−3 | 5.99 × 10−4 | *** |
| Tmin Spr | 2.58 × 10−3 | 9.45 × 10−4 | *** | −5.68 × 10−3 | 8.51 × 10−4 | *** |
| Tmin Summer | −5.92 × 10−3 | 8.85 × 10−4 | *** | 2.74 × 10−3 | 8.43 × 10−4 | ** |
| Tmin Autumn | 7.21 × 10−3 | 9.46 × 10−4 | *** | |||
| AI: PP Autumn-1 | −3.46 × 10−3 | 2.65 × 10−4 | *** | |||
| AI: PP Win | −4.44 × 10−3 | 2.84 × 10−4 | *** | 4.31 × 10−4 | 2.62 × 10−4 | |
| AI: PP Spr | −2.20 × 10−3 | 2.65 × 10−4 | *** | −1.70 × 10−4 | 2.46 × 104 | |
| AI: PP Summer | −1.50 × 10−3 | 2.56 × 10−4 | *** | 1.42 × 10−4 | 2.33 × 10−4 | |
| AI: PP Autumn | −2.68 × 10−3 | 2.42 × 10−4 | *** | |||
| AI: Tmax Autumn-1 | −1.15 × 10−3 | 6.84 × 10−4 | ||||
| AI: Tmax Win | 5.53 × 10−3 | 7.53 × 10−4 | *** | 1.38 × 10−3 | 6.86 × 10−4 | * |
| AI: Tmax Spr | 3.77 × 10−3 | 7.00 × 10−4 | *** | −1.62 × 10−3 | 6.00 × 10−4 | ** |
| AI: Tmax Summer | −4.86 × 10−3 | 5.93 × 10−4 | *** | 2.01 × 10−3 | 5.60 × 10−4 | *** |
| AI: Tmax Autumn | −1.61 × 10−3 | 6.47 × 10−4 | ||||
| AI: Tmin Autumn-1 | 1.48 × 10−4 | 1.05 × 10−3 | * | |||
| AI: Tmin Win | −1.07 × 10−3 | 7.25 × 10−4 | −1.78 × 10−3 | 6.47 × 10−4 | ** | |
| AI: Tmin Spr | −5.99 × 10−3 | 1.01 × 10−3 | *** | 2.10 × 10−3 | 9.02 × 10−4 | * |
| AI: Tmin Summer | 6.19 × 10−3 | 9.50 × 10−4 | *** | −1.23 × 10−3 | 9.07 × 10−4 | |
| AI: Tmin Autumn | 7.42 × 10−4 | 1.03 × 10−3 | ||||
| Ep: PP Autumn-1 | −3.01 × 10−4 | 2.63 × 10−4 | ||||
| Ep: PP Win | 2.16 × 10−4 | 2.83 × 10−4 | 5.30 × 10−4 | 2.55 × 10−4 | * | |
| Ep: PP Spr | −3.20 × 10−4 | 2.71 × 10−4 | 5.64 × 10−4 | 2.51 × 10−4 | * | |
| Ep: PP Summer | −3.90 × 10−4 | 2.56 × 10−4 | −1.13 × 10−3 | 2.32 × 10−4 | *** | |
| Ep: PP Autumn | 2.63 × 10−4 | 2.37 × 10−4 | ||||
| Ep: Tmax Autumn-1 | 1.50 × 10−4 | 6.50 × 10−4 | ||||
| Ep: Tmax Win | −8.95 × 10−4 | 7.16 × 10−4 | 1.93 × 10−3 | 6.53 × 10−4 | ** | |
| Ep: Tmax Spr | 1.35 × 10−3 | 6.70 × 10−4 | * | −1.20 × 10−3 | 5.73 × 10−4 | * |
| Ep: Tmax Summer | 1.83 × 10−4 | 5.42 × 10−4 | −9.88 × 10−5 | 5.03 × 10−4 | ||
| Ep: Tmax Autumn | 8.41 × 10−4 | 6.17 × 10−4 | ||||
| Ep: Tmin Autumn-1 | 1.46 × 10−3 | 1.00 × 10−3 | ||||
| Ep: Tmin Win | 4.33 × 10−3 | 6.98 × 10−4 | *** | 7.77 × 10−4 | 6.22 × 10−4 | |
| Ep: Tmin Spr | −4.32 × 10−3 | 9.93 × 10−4 | *** | −3.43 × 10−4 | 8.90 × 10−4 | |
| Ep: Tmin Summer | −1.57 × 10−3 | 8.89 × 10−4 | 1.56 × 10−3 | 8.34 × 10−4 | ||
| Ep: Tmin Autumn | −1.31 × 10−3 | 9.79 × 10−4 | ||||
| AIC | BIC | logLik | Deviance | df.resid | ||
|---|---|---|---|---|---|---|
| Earlywood | Full | 102,951.6 | 103,402.9 | −51,424.8 | 102,849.6 | 51,435 |
| Null | 115,841.8 | 115,948.0 | −57,908.9 | 115,817.8 | 51,474 | |
| no AI | 104,370.0 | 104,697.4 | −52,148.0 | 104,296.0 | 51,449 | |
| no Ewp | 103,000.0 | 103,327.4 | −51,463.0 | 102,926.0 | 51,449 | |
| no Clima | 115,767.0 | 115,899.8 | −57,868.5 | 115,737.0 | 51,471 | |
| Latewood | Full | 79,472.3 | 79,799.7 | −39,699.1 | 79,398.3 | 51,449 |
| Null | 82,315.1 | 82,421.3 | −41,145.6 | 82,291.1 | 51,474 | |
| no AI | 79,919.5 | 80,247.0 | −39,922.8 | 79,845.5 | 51,449 | |
| no Ewp | 79,710.7 | 80,162.1 | −39,804.4 | 79,608.7 | 51,435 | |
| no Clima | 82,665.0 | 82,797.7 | −41,317.5 | 82,635.0 | 51,471 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Royo-Navascues, M.; Martinez del Castillo, E.; Serrano-Notivoli, R.; Tejedor, E.; Novak, K.; Longares, L.A.; Saz, M.A.; de Luis, M. When Density Matters: The Spatial Balance between Early and Latewood. Forests 2021, 12, 818. https://doi.org/10.3390/f12070818
Royo-Navascues M, Martinez del Castillo E, Serrano-Notivoli R, Tejedor E, Novak K, Longares LA, Saz MA, de Luis M. When Density Matters: The Spatial Balance between Early and Latewood. Forests. 2021; 12(7):818. https://doi.org/10.3390/f12070818
Chicago/Turabian StyleRoyo-Navascues, Maria, Edurne Martinez del Castillo, Roberto Serrano-Notivoli, Ernesto Tejedor, Klemen Novak, Luis Alberto Longares, Miguel Angel Saz, and Martin de Luis. 2021. "When Density Matters: The Spatial Balance between Early and Latewood" Forests 12, no. 7: 818. https://doi.org/10.3390/f12070818
APA StyleRoyo-Navascues, M., Martinez del Castillo, E., Serrano-Notivoli, R., Tejedor, E., Novak, K., Longares, L. A., Saz, M. A., & de Luis, M. (2021). When Density Matters: The Spatial Balance between Early and Latewood. Forests, 12(7), 818. https://doi.org/10.3390/f12070818