
Effects of Nitrogen Additions on Soil Respiration in an Asian Tropical Montane Rainforest

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Field Manipulations
2.3. Soil Respiration and Environmental Parameter Measurements
2.4. Soil Characteristics
2.5. Statistical Analysis
3. Results
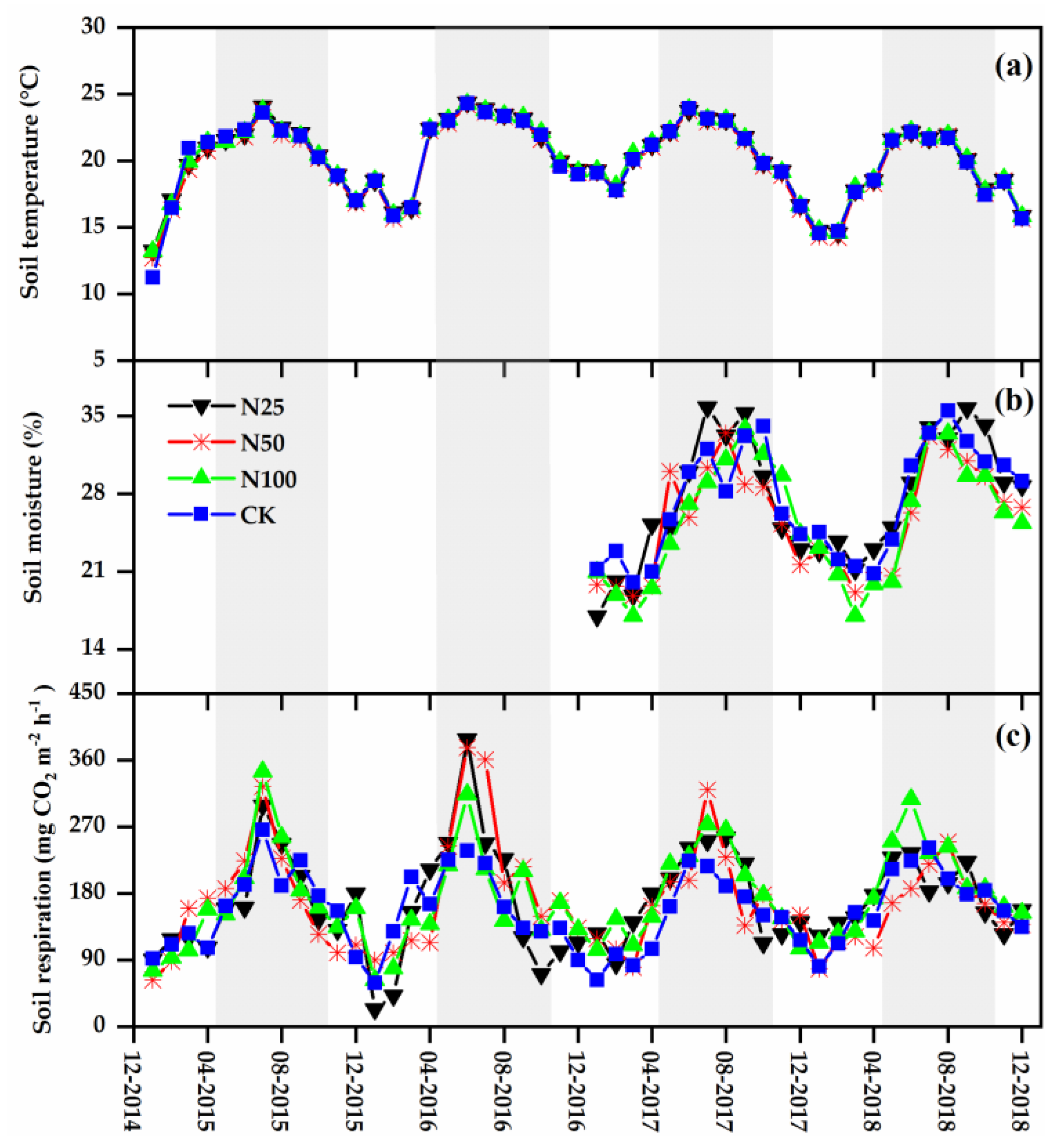
3.1. Soil Environmental Conditions
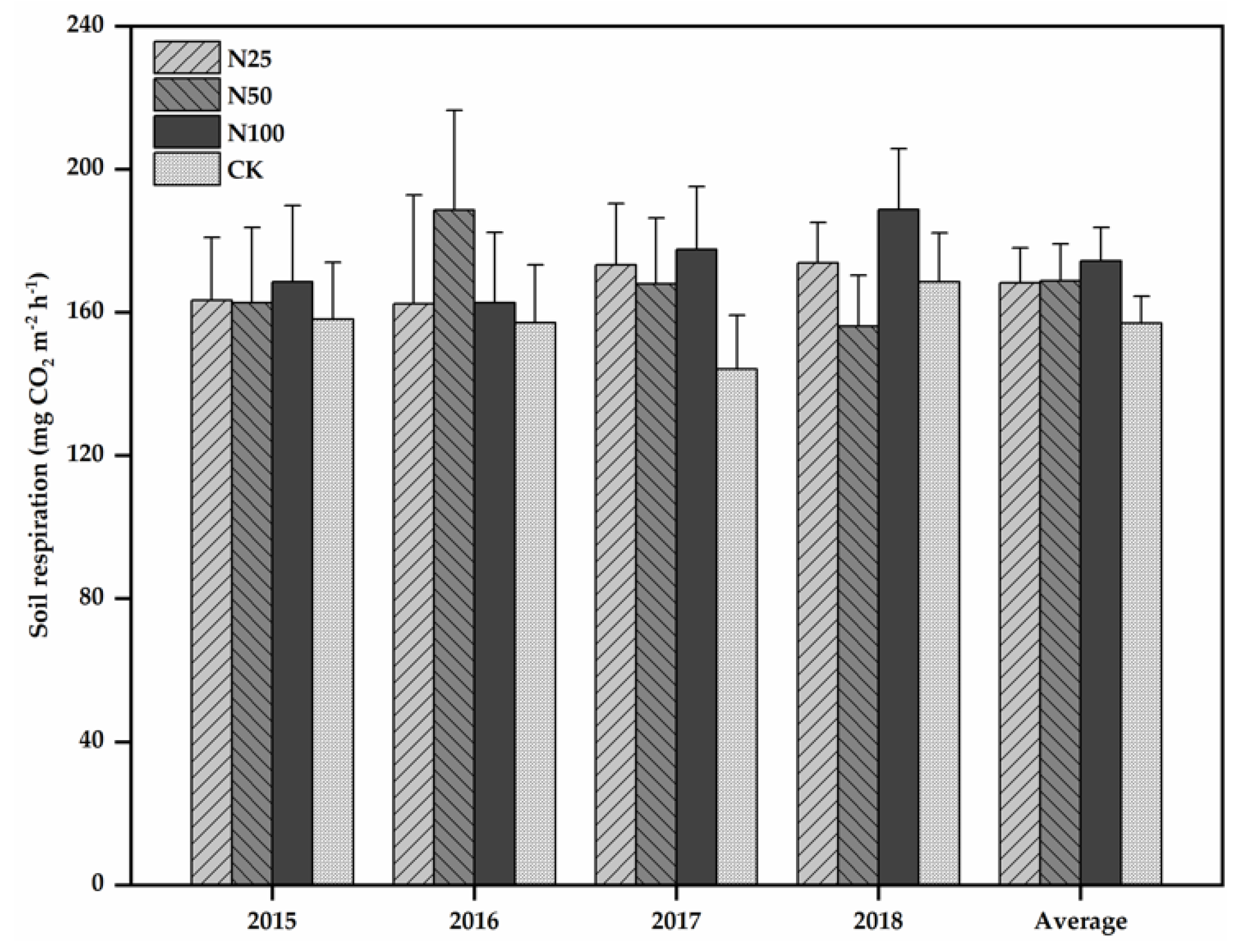
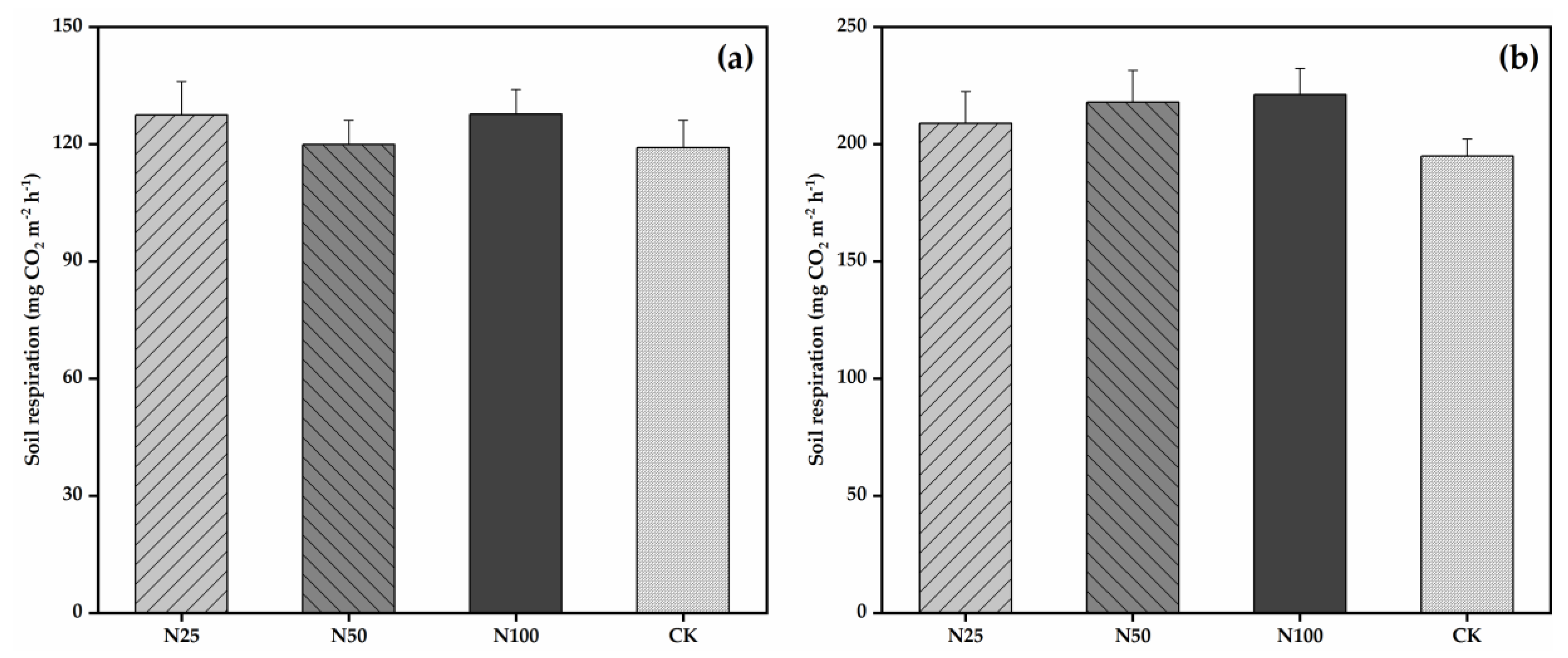
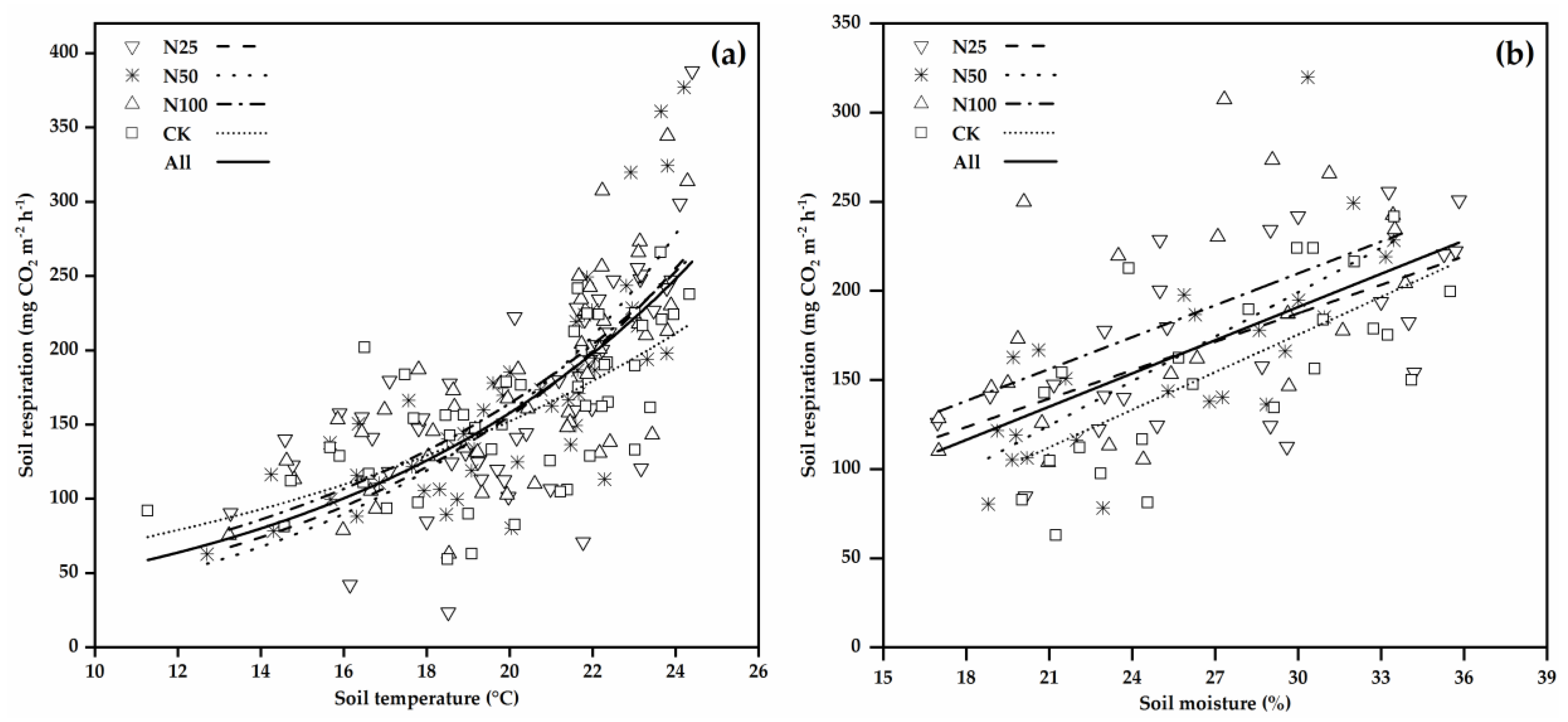
3.2. Soil Respiration and Its Temperature Sensitivity
4. Discussion
4.1. Comparison with Previous Studies
4.2. Environmental Controls on Soil Respiration
4.3. Effects of N Additions on Soil Respiration
4.4. Effects of N Additions on Temperature Sensitivity
4.5. Uncertainties and Limitations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Chaturvedi, A. Will Carbon Market Change the Fate of This Planet? Available online: https://ssrn.com/abstract=3632416; http://dx.doi.org/10.2139/ssrn.3632416 (accessed on 4 March 2021).
- Huntzinger, D.; Michalak, A.; Schwalm, C.; Ciais, P.; King, A.; Fang, Y.; Schaefer, K.; Wei, Y.; Cook, R.; Fisher, J.; et al. Uncertainty in the response of terrestrial carbon sink to environmental drivers undermines carbon-climate feedback predictions. Sci. Rep. 2017, 7, 4765. [Google Scholar] [CrossRef] [PubMed]
- Trumper, K.; Bertzky, M.; Dickson, B.; van der Heijden, G.; Jenkins, M.; Manning, P. The Natural Fix? The Role of Ecosystems in Climate Mitigation; A UNEP Rapid Response Assessment; United Nations Environment Programme, UNEP-WCMC: Cambridge, UK, 2009. [Google Scholar]
- Xu, M.; Shang, H. Contribution of soil respiration to the global carbon equation. J. Plant Physiol. 2016, 203, 16–28. [Google Scholar] [CrossRef]
- Bowden, R.; Davidson, E.; Savage, K.; Arabia, C.; Steudler, P. Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest. Forest Ecol. Manag. 2004, 196, 43–56. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, L.; Ma, Y.; Yin, G.; Zhao, C.; Zhang, Y.; Piao, S. The effect of nitrogen addition on soil respiration from a nitrogen-limited forest soil. Agric. For. Meteorol. 2014, 197, 103–110. [Google Scholar] [CrossRef]
- Scott-Denton, L.; Sparks, K.; Monson, R. Spatial and temporal controls over soil respiration rate in a high-elevation, subalpine forest. Soil Biol. Biochem. 2003, 35, 525–534. [Google Scholar] [CrossRef]
- Monson, R.; Lipson, D.; Burns, S.; Turnipseed, A.; Delany, A.; Williams, M.; Schmidt, S. Winter forest soil respiration controlled by climate and microbial community composition. Nature 2006, 439, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Baishya, R. Interactive effects of soil moisture and temperature on soil respiration under native and non-native tree species in semi-arid forest of Delhi, India. Trop. Ecol. 2019, 60, 252–260. [Google Scholar] [CrossRef]
- Čater, M.; Darenova, E.; Simončič, P. Harvesting intensity and tree species affect soil respiration in uneven-aged Dinaric forest stands. Forest Ecol. Manag. 2021, 480, 118638. [Google Scholar] [CrossRef]
- Vitousek, P.; Howarth, R. Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry 1991, 13, 87–115. [Google Scholar] [CrossRef]
- Tu, L.; Hu, T.; Zhang, J.; Li, X.; Hu, H.; Liu, L.; Xiao, Y. Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem. Soil Biol. Biochem. 2013, 58, 255–264. [Google Scholar] [CrossRef]
- Zhou, S.; Xiang, Y.; Tie, L.; Han, B.; Huang, C. Simulated nitrogen deposition significantly reduces soil respiration in an evergreen broadleaf forest in western China. PLoS ONE 2018, 13, e0204661. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, C.; Townsend, A. Nutrient additions to a tropical rain forest drive substantial soil carbon dioxide losses to the atmosphere. Proc. Natl. Acad. Sci. USA 2006, 103, 10316–10321. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Hasselquist, N.; Palmroth, S.; Zheng, Z.; You, W. Short-term response of soil respiration to nitrogen fertilization in a subtropical evergreen forest. Soil Biol. Biochem. 2014, 76, 297–300. [Google Scholar] [CrossRef]
- Ambus, P.; Robertson, G. The effect of increased N deposition of nitrous oxide, methane and carbon dioxide fluxes from unmanaged forest and grassland communities in Michigan. Biogeochemistry 2006, 79, 315–337. [Google Scholar] [CrossRef]
- Gao, W.; Yang, H.; Li, S.; Kou, L. Responses of soil CO2, CH4 and N2O fluxes to N, P, and acid additions in mixed forest in subtropical China. J. Resour. Ecol. 2017, 8, 154–164. [Google Scholar]
- Wang, C.; Yang, X.; Xu, K. Effect of chronic nitrogen fertilization on soil CO2 flux in a temperate forest in North China: A 5-year nitrogen addition experiment. J. Soil Sed. 2018, 18, 506–516. [Google Scholar] [CrossRef]
- Mo, J.; Zhang, W.; Zhu, W.; Fang, Y.; Li, D.; Zhao, P. Response of soil respiration to simulated N deposition in a disturbed and a rehabilitated tropical forest in southern China. Plant Soil 2007, 296, 125–135. [Google Scholar] [CrossRef]
- Lü, C.; Tian, H. Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data. J. Geophys. Res. 2007, 112, D22S05. [Google Scholar] [CrossRef]
- Du, Y.; Guo, P.; Liu, J.; Wang, C.; Yang, N.; Jiao, Z. Different types of nitrogen deposition show variable effects on the soil carbon cycle process of temperate forests. Glob. Chang. Biol. 2014, 20, 3222–3228. [Google Scholar] [CrossRef]
- Liu, B.; Mou, C.; Yan, G.; Xu, L.; Jiang, S.; Xing, Y.; Han, S.; Yu, J.; Wang, Q. Annual soil CO2 efflux in a cold temperate forest in northeastern China: Effects of winter snowpack and artificial nitrogen deposition. Sci. Rep. 2016, 6, 18957. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Zhou, G.; Liu, J.; Liu, S.; Duan, H.; Zhang, D. Responses of soil respiration to elevated carbon dioxide and nitrogen addition in young subtropical forest ecosystems in China. Biogeosciences 2010, 7, 315–328. [Google Scholar] [CrossRef]
- Peng, Y.; Song, S.; Li, Z.; Li, S.; Chen, G.; Hu, H.; Xie, J.; Chen, G.; Xiao, Y.; Liu, L.; et al. Influences of nitrogen addition and aboveground litter-input manipulations on soil respiration and biochemical properties in a subtropical forest. Soil Biol. Biochem. 2020, 142, 107694. [Google Scholar] [CrossRef]
- Tang, W.; Chen, D.; Phillips, O.; Liu, X.; Zhou, Z.; Li, Y.; Xi, D.; Zhu, F.; Fang, J.; Zhang, L.; et al. Effects of long-term increased N deposition on tropical montane forest soil N2 and N2O emissions. Soil Biol. Biochem. 2018, 126, 194–203. [Google Scholar] [CrossRef]
- Mo, J.; Zhang, W.; Zhu, W.; Gundersen, P.; Fang, Y.; Li, D.; Wang, H. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Glob. Chang. Biol. 2008, 14, 403–412. [Google Scholar] [CrossRef]
- Du, E.; Terrer, C.; Pellegrini, A.; Ahlström, A.; van Lissa, C.; Zhao, X.; Xia, N.; Wu, X.; Jackson, R. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 2020, 13, 221–226. [Google Scholar] [CrossRef]
- Du, E.; Zhou, Z.; Li, P.; Hu, X.; Ma, Y.; Wang, W.; Zheng, C.; Zhu, J.; He, J.; Fang, J. NEECF: A project of nutrient enrichment experiments in China’s forests. J. Plant Ecol. 2013, 6, 428–435. [Google Scholar] [CrossRef]
- Wang, A.; Zhu, W.; Gundersen, P.; Phillips, O.; Chen, D.; Fang, Y. Fates of atmospheric deposited nitrogen in an Asian tropical primary forest. For. Ecol. Manag. 2018, 411, 213–222. [Google Scholar] [CrossRef]
- Chen, D.; Li, Y.; Liu, H.; Xu, H.; Xiao, W.; Luo, T.; Zhou, Z.; Lin, M. Biomass and carbon dynamics of a tropical mountain rain forest in China. Sci. China Life Sci. 2010, 53, 798–810. [Google Scholar] [CrossRef]
- Fang, J.; Li, Y.; Zhu, B.; Liu, G.; Zhou, G. Community structures and species richness in the montane rain forest of Jianfengling, Hainan Island, China. Biodivers. Sci. 2004, 12, 29–43. [Google Scholar]
- Shi, J.; Xu, H.; Lin, M.; Li, Y. Dynamics of litterfall production in the tropical mountain rainforest of Jianfengling, Hainan Island, China. Plant Sci. J. 2019, 37, 593–601. [Google Scholar]
- Zhou, Z. Effects of Nitrogen and Phosphorus Additions on Carbon Cycling of Tropical Mountain Rainforests in Hainan Island, China. Ph.D. Thesis, Peking University, Beijing, China, June 2013. [Google Scholar]
- Bai, Z.; Yang, G.; Chen, H.; Zhu, Q.; Chen, D.; Li, Y.; Wang, X.; Wu, Z.; Zhou, G.; Peng, C. Nitrous oxide fluxes from three forest types of the tropical mountain rainforests on Hainan Island, China. Atmos. Environ. 2014, 92, 469–477. [Google Scholar] [CrossRef]
- Hutchinson, G.; Mosier, A. Improved soil cover method for field measurement of nitrous oxide fluxes. Soil Sci. Soc. Am. J. 1981, 45, 311–316. [Google Scholar] [CrossRef]
- Chen, Z.; Ding, W.; Luo, Y.; Yu, H.; Xu, Y.; Müller, C.; Xu, X.; Zhu, T. Nitrous oxide emissions from cultivated blacksoil: A case study in Northeast China and global estimates using empirical model. Glob. Biogeochem. Cycles 2014, 27, 1311–1326. [Google Scholar] [CrossRef]
- Badalucco, L.; Gelsomino, A.; Dellorco, S.; Grego, S.; Nannipieri, P. Biochemical characterization of soil organic compounds extracted by 0.5 M K2SO4 before and after chloroform fumigation. Soil Biol. Beachem. 1992, 24, 569–578. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J. On the temperature dependence of soil respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Wei, H. Soil Greenhouse Gas Emission and Its Influencing Factors from Temperate Deciduous Broad-leaved Forest and Tropical Mountain Rain Forest. Ph.D. Thesis, Northwest A&F University, Yangling, China, September 2018. [Google Scholar]
- Ishizuka, S.; Iswandi, A.; Nakajima, Y.; Yonemura, S.; Sudo, S.; Tsuruta, H.; Muriyarso, D. Spatial patterns of greenhouse gas emission in a tropical rainforest in Indonesia. Nutr. Cycl. Agroecosyst. 2005, 71, 55–62. [Google Scholar] [CrossRef]
- Bekku, Y.; Nakatsubo, T.; Kume, A.; Adachi, M.; Koizumi, H. Effect of warming on the temperature dependence of soil respiration rate in arctic, temperate and tropical soils. Appl. Soil Ecol. 2003, 22, 205–210. [Google Scholar] [CrossRef]
- Wang, C.; Yang, J.; Zhang, Q. Soil respiration in six temperate forests in China. Glob. Chang. Biol. 2006, 12, 2103–2114. [Google Scholar] [CrossRef]
- Yuan, C.; Zhu, G.; Yang, S.; Xu, G.; Li, Y.; Gong, H.; Wu, C. Soil warming increases soil temperature sensitivity in subtropical Forests of SW China. PeerJ 2019, 7, e7721. [Google Scholar] [CrossRef]
- Xu, Z.; Tang, S.; Xiong, L.; Yang, W.; Yin, H.; Tu, L.; Wu, F.; Chen, L.; Tan, B. Temperature sensitivity of soil respiration in China’s forest ecosystems: Patterns and controls. Appl. Soil Ecol. 2015, 93, 105–110. [Google Scholar] [CrossRef]
- Jiang, L.; Ma, S.; Zhou, Z.; Zheng, T.; Jiang, X.; Cai, Q.; Li, P.; Zhu, J.; Li, Y.; Fang, J. Soil respiration and its partitioning in different components in tropical primary and secondary mountain rain forests in Hainan Island, China. J. Plant Ecol. 2017, 10, 791–799. [Google Scholar] [CrossRef]
- Zimmermann, M.; Davies, K.; Peña de Zimmermann, V.; Bird, M. Impact of temperature and moisture on heterotrophic soil respiration along a moist tropical forest gradient in Australia. Soil Res. 2015, 53, 286–297. [Google Scholar] [CrossRef]
- Campbell, J.; Law, B. Forest soil respiration across three climatically distinct chronosequences in Oregon. Biogeochemistry 2005, 73, 109–125. [Google Scholar] [CrossRef]
- Ryan, M.; Law, B. Interpreting, measuring, and modeling soil respiration. Biogeochemistry 2005, 73, 3–27. [Google Scholar] [CrossRef]
- Kittredge, H.; Cannone, T.; Funk, J.; Chapman, S. Soil respiration and extracellular enzyme production respond differently across seasons to elevated temperatures. Plant Soil 2018, 425, 351–361. [Google Scholar] [CrossRef]
- Ilsteld, U.; Nordgren, A.; Malmer, M. Optimum soil water for soil respiration before and after amendment with glucose in humid tropical acrisols and a boreal mor layer. Soil Biol. Biochem. 2000, 32, 1591–1599. [Google Scholar]
- Phillips, C.; Nickerson, N.; Risk, D.; Kayler, Z.; Andersen, C.; Mix, A.; Bond, B. Soil moisture effects on the carbon isotope composition of soil respiration. Rapid Commun. Mass Spectrom. 2010, 24, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Song, X.; Chang, S.; Peng, C.; Xiao, W.; Zhang, J.; Xiang, W.; Li, Y.; Wang, W. Nitrogen depositions increase soil respiration and decrease temperature sensitivity in a Moso bamboo forest. Agric. For. Meteorol. 2019, 268, 48–54. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, W.; Sun, T.; Chen, L.; Pang, X.; Wang, Y.; Xiao, F. N and P fertilization reduced soil autotrophic and heterotrophic respiration in a young Cunninghamia lanceolata forest. Agric. For. Meteorol. 2017, 232, 66–73. [Google Scholar] [CrossRef]
- Yan, W.; Chen, X.; Peng, Y.; Zhu, F.; Zhen, W.; Zhang, X. Response of soil respiration to nitrogen addition in two subtropical forest types. Pedosphere 2020, 30, 478–486. [Google Scholar] [CrossRef]
- Sotta, E.; Meir, P.; Malhi, Y.; Nobre, A.; Hodnett, M.; Grace, J. Soil CO2 efflux in a tropical forest in the central Amazon. Glob. Chang. Biol. 2004, 10, 601–617. [Google Scholar] [CrossRef]
- Song, L.; Tian, P.; Zhang, J.; Jin, G. Effects of three years of simulated nitrogen deposition on soil nitrogen dynamics and greenhouse gas emissions in a Korean pine plantation of northeast China. Sci. Total Environ. 2017, 609, 1303–1311. [Google Scholar] [CrossRef]
- Xiao, H.; Shi, Z.; Li, Z.; Wang, L.; Chen, J.; Wang, J. Responses of soil respiration and its temperature sensitivity to nitrogen addition: A meta-analysis in China. Appl. Soil Ecol. 2020, 150, 103484. [Google Scholar] [CrossRef]
- Zhou, T.; Shi, P.; Hui, D.; Luo, Y. Global pattern of temperature sensitivity of soil heterotrophic respiration (Q10) and its implications for carbon-climate feedback. J. Geophys. Res. 2009, 114, G02016. [Google Scholar] [CrossRef]
- Cusack, D.; Torn, M.; McDowell, W.; Silver, W. The response of heterotrophic activity and carbon cycling to nitrogen additions and warming in two tropical soils. Glob. Chang. Biol. 2010, 16, 2555–2572. [Google Scholar] [CrossRef]
- Sinsabaugh, R.; Gallo, M.; Lauber, C.; Waldrop, M.; Zak, D. Extracellular enzyme activities and soil organic matter dynamics for northern hardwood forests receiving simulated nitrogen deposition. Biogeochemistry 2005, 75, 201–215. [Google Scholar] [CrossRef]
- Fang, C.; Moncrieff, J. An open-top chamber for measuring soil respiration and the influence of pressure difference on CO2 efflux measurement. Funct. Ecol. 1998, 12, 319–325. [Google Scholar] [CrossRef]
- Davidson, E.; Savage, K.; Verchot, L.; Navarro, R. Minimizing artifacts and biases in chamber-based measurements of soil respiration. Agric. For. Meteorol. 2002, 113, 21–37. [Google Scholar] [CrossRef]
- Liang, N.; Nakadai, T.; Hirano, T.; Qu, L.; Koike, T.; Fujinuma, Y.; Inoue, G. In situ comparison of four approaches to estimating soil CO2 efflux in a northern larch (Larix kaempferi Sarg.) forest. Agric. For. Meteorol. 2004, 123, 97–117. [Google Scholar] [CrossRef]
- Janssens, I.; Kowalski, A.; Longdoz, B.; Ceulemans, R. Assessing forest soil CO2 efflux: An in-situ comparison of four techniques. Tree Physiol. 2000, 20, 23–32. [Google Scholar] [CrossRef]
- Zheng, S.; Bian, H.; Quan, Q.; Xu, L.; Chen, Z.; He, N. Effect of nitrogen and acid deposition on soil respiration in a temperate forest in China. Geoderma 2018, 329, 82–90. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N Treatment | ||||
|---|---|---|---|---|
| N25 | N50 | N100 | CK | |
| Soil temperature (°C) | 20.1 ± 0.4 | 19.9 ± 0.4 | 20.1 ± 0.4 | 20.0 ± 0.4 |
| Soil moisture (%) | 27.4 ± 1.2 | 25.5 ± 1.0 | 25.5 ± 1.1 | 27.3 ± 1.0 |
| NH4+-N (mg kg−1) | 18.9 ± 2.0 | 16.1 ± 1.2 | 19.0 ± 2.9 | 15.5 ± 1.3 |
| NO3−-N (mg kg−1) | 14.4 ± 1.8 | 15.4 ± 2.4 | 13.8 ± 2.8 | 11.5 ± 1.0 |
| MBC (mg kg−1) | 108.5 ± 17.0 | 81.5 ± 19.6 | 87.8 ± 15.9 | 98.6 ± 24.5 |
| MBN (mg kg−1) | 105.4 ± 18.2 | 101.4 ± 19.3 | 88.4 ± 9.4 | 106.5 ± 18.8 |
| Source of Variation | RS | |
|---|---|---|
| F | p | |
| Month | 35.44 | <0.001 |
| Treatment | 1.84 | 0.19 |
| Month × Treatment | 0.72 | 0.70 |
| m | n | R2 | p | Q10 | a | b | R2 | p | |
|---|---|---|---|---|---|---|---|---|---|
| N25 | 13.11 | 0.1237 | 0.50 | <0.001 | 3.45 | 5.32 | 27.83 | 0.35 | <0.005 |
| N50 | 9.36 | 0.1413 | 0.64 | <0.001 | 4.11 | 8.31 | −50.06 | 0.51 | <0.001 |
| N100 | 19.02 | 0.1078 | 0.52 | <0.001 | 2.94 | 5.96 | 31.01 | 0.27 | <0.005 |
| CK | 29.47 | 0.0821 | 0.41 | <0.001 | 2.27 | 7.03 | −35.37 | 0.46 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, F.; Peng, C.; Liu, W.; Liu, Z.; Wang, H.; Chen, D.; Li, Y. Effects of Nitrogen Additions on Soil Respiration in an Asian Tropical Montane Rainforest. Forests 2021, 12, 802. https://doi.org/10.3390/f12060802
Wu F, Peng C, Liu W, Liu Z, Wang H, Chen D, Li Y. Effects of Nitrogen Additions on Soil Respiration in an Asian Tropical Montane Rainforest. Forests. 2021; 12(6):802. https://doi.org/10.3390/f12060802
Chicago/Turabian StyleWu, Fangtao, Changhui Peng, Weiguo Liu, Zhihao Liu, Hui Wang, Dexiang Chen, and Yide Li. 2021. "Effects of Nitrogen Additions on Soil Respiration in an Asian Tropical Montane Rainforest" Forests 12, no. 6: 802. https://doi.org/10.3390/f12060802
APA StyleWu, F., Peng, C., Liu, W., Liu, Z., Wang, H., Chen, D., & Li, Y. (2021). Effects of Nitrogen Additions on Soil Respiration in an Asian Tropical Montane Rainforest. Forests, 12(6), 802. https://doi.org/10.3390/f12060802