
Plantations of Cinnamomum camphora (Linn) Presl with Distinct Soil Bacterial Communities Mitigate Soil Acidity within Polluted Locations in Southwest China

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description and Plot Design
2.2. Soil Samplings and Analysis
2.3. Statistical Analysis
3. Results
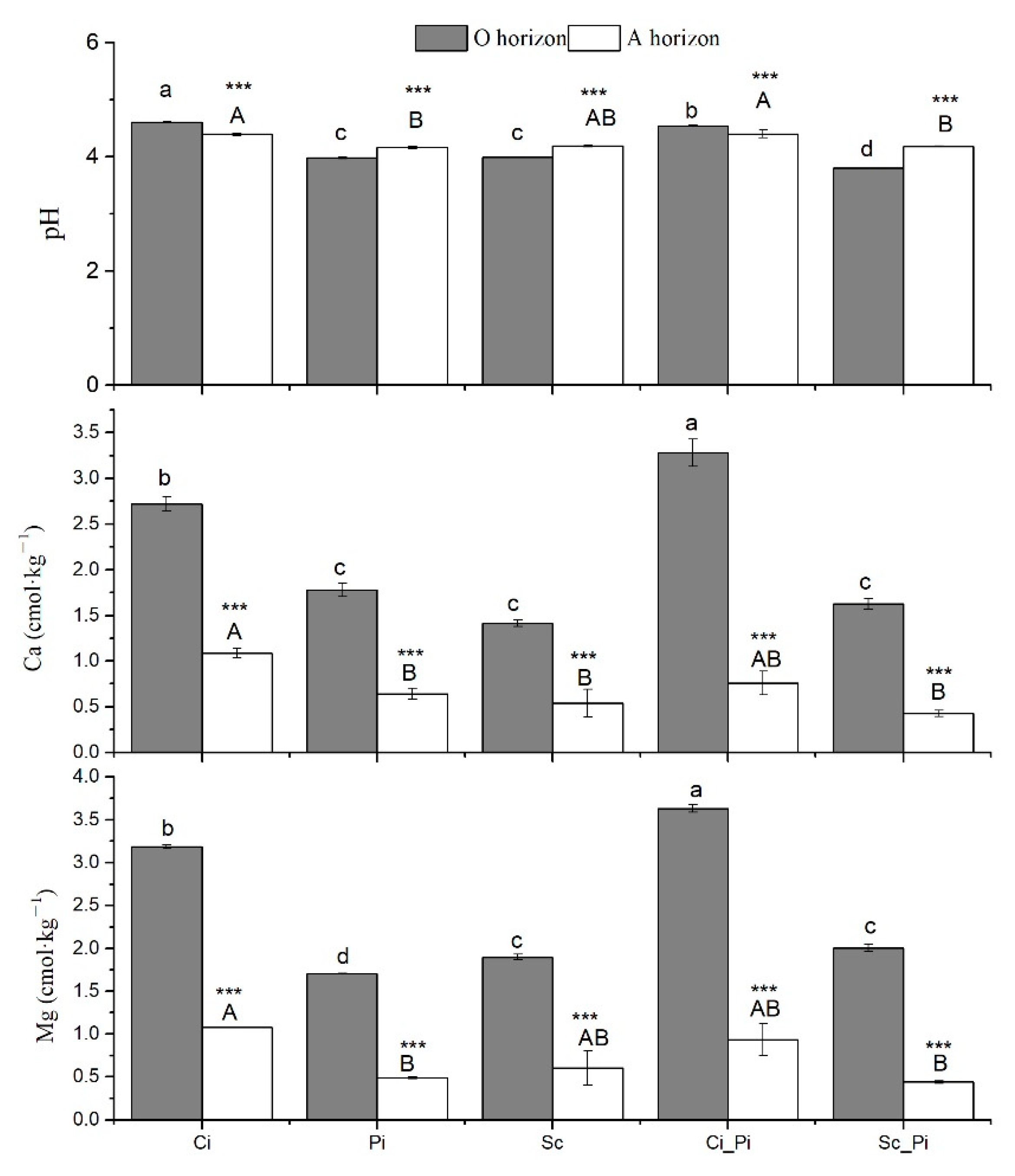
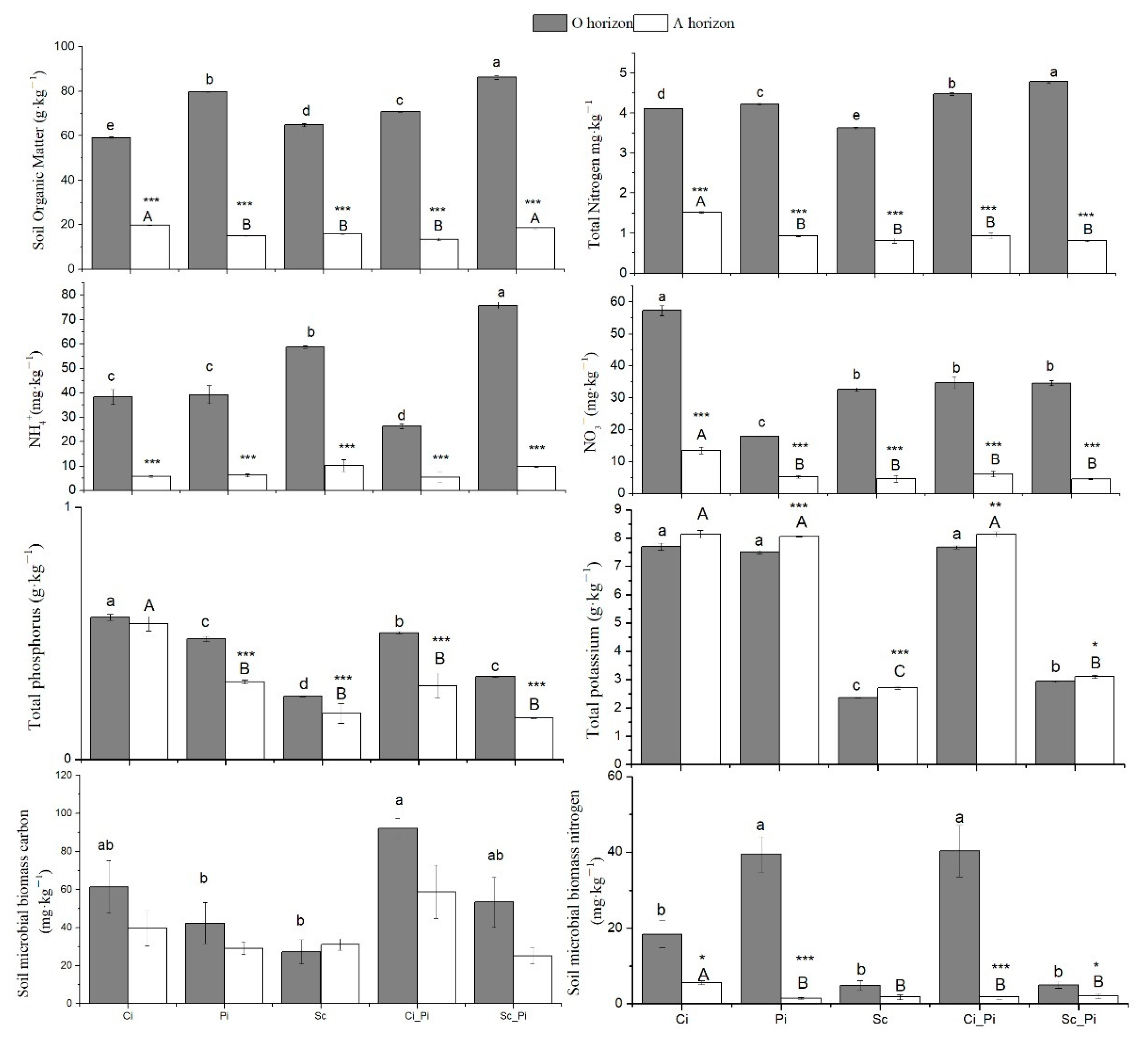
3.1. Soil Physicochemical Properties and Soil Microbial Biomass
3.2. Soil Bacterial Diversity
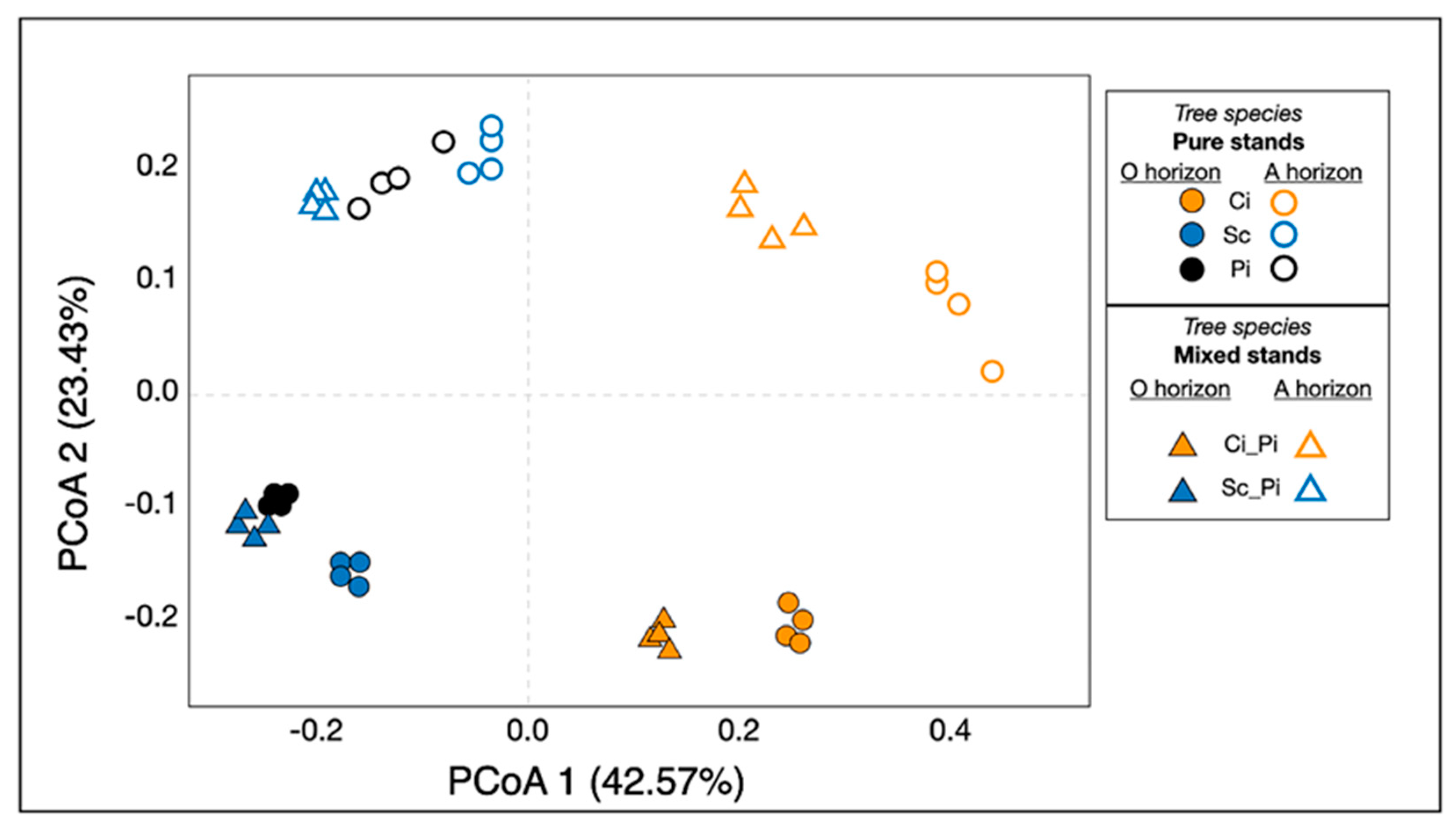
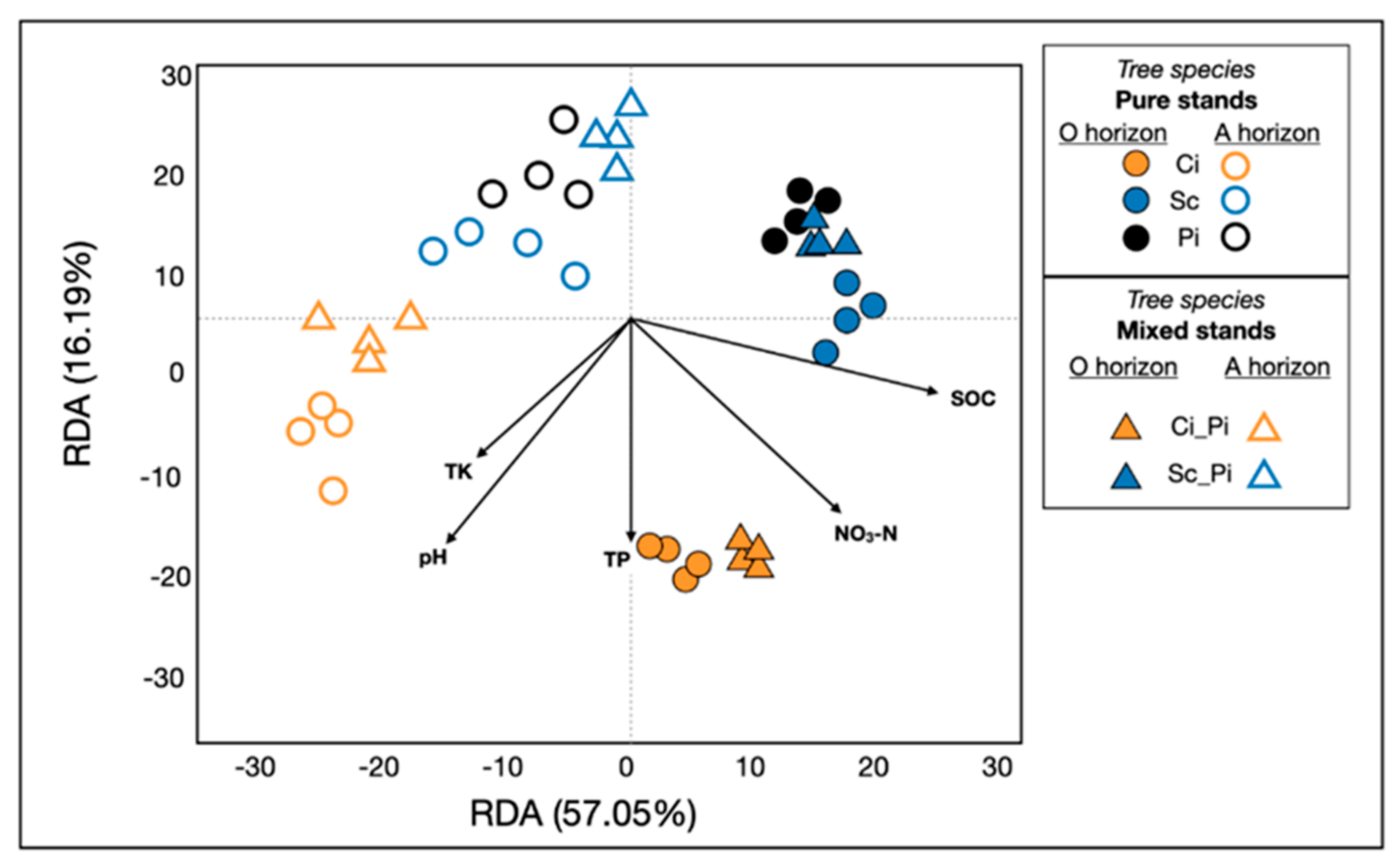
3.3. Soil Bacterial Community Structure and Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duan, L.; Yu, Q.; Zhang, Q.; Wang, Z.; Pan, Y.; Larssen, T.; Tang, J.; Mulder, J. Acid deposition in Asia: Emissions, deposition, and ecosystem effects. Atmos. Environ. 2016, 146, 55–69. [Google Scholar] [CrossRef]
- Yu, H.; He, N.; Wang, Q.; Zhu, J.; Gao, Y.; Zhang, Y.; Jia, Y.; Yu, G. Development of atmospheric acid deposition in China from the 1990s to the 2010s. Environ. Pollut. 2017, 231, 182–190. [Google Scholar] [CrossRef]
- Wang, S.X.; Zhao, B.; Cai, S.Y.; Klimont, Z.; Nielsen, C.P.; Morikawa, T.; Woo, J.H.; Kim, Y.; Fu, X.; Xu, J.Y.; et al. Emission trends and mitigation options for air pollutants in East Asia. Atmos. Chem. Phys. 2014, 14, 6571–6603. [Google Scholar] [CrossRef]
- Kang, Y.; Liu, M.; Song, Y.; Huang, X.; Yao, H.; Cai, X.; Zhang, H.; Kang, L.; Liu, X.; Yan, X.; et al. High-resolution ammonia emissions inventories in China from 1980 to 2012. Atmos. Chem. Phys. 2016, 16, 2043–2058. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [PubMed]
- Simkin, S.M.; Allen, E.B.; Bowman, W.D.; Clark, C.M.; Belnap, J.; Brooksf, M.L.; Cade, B.S.; Collins, S.L.; Geiser, L.H.; Gilliam, F.S.; et al. Conditional vulnerability of plant diversity to Atmospheric nitrogen deposition across the United States. Proc. Natl. Acad. Sci. USA 2016, 113, 4086–4091. [Google Scholar] [CrossRef]
- Huang, Y.; Kang, R.; Mulder, J.; Zhang, T.; Duan, L. Nitrogen saturation, soil acidification, and ecological effects in a subtropical pine forest on acid soil in southwest China. J. Geophys. Res. G Biogeosci. 2015, 120, 2457–2472. [Google Scholar] [CrossRef]
- Berger, T.W.; Türtscher, S.; Berger, P.; Lindebner, L. A slight recovery of soils from Acid Rain over the last three decades is not reflected in the macro nutrition of beech (Fagus sylvatica) at 97 forest stands of the Vienna Woods. Environ. Pollut. 2016, 216, 624–635. [Google Scholar] [CrossRef]
- Maltz, M.R.; Chen, Z.; Cao, J.; Arogyaswamy, K.; Shulman, H.; Aronson, E.L. Inoculation with Pisolithus tinctorius may ameliorate acid rain impacts on soil microbial communities associated with Pinus massoniana seedlings. Fungal Ecol. 2019, 40, 50–61. [Google Scholar] [CrossRef]
- Graf Pannatier, E.; Thimonier, A.; Schmitt, M.; Walthert, L.; Waldner, P. A decade of monitoring at Swiss Long-Term Forest Ecosystem Research (LWF) sites: Can we observe trends in atmospheric acid deposition and in soil solution acidity? Environ. Monit. Assess. 2011, 174, 3–30. [Google Scholar] [CrossRef]
- Marx, A.; Hintze, S.; Sanda, M.; Jankovec, J.; Oulehle, F.; Dusek, J.; Vitvar, T.; Vogel, T.; van Geldern, R.; Barth, J.A.C. Acid rain footprint three decades after peak deposition: Long-term recovery from pollutant sulphate in the Uhlirska catchment (Czech Republic). Sci. Total Environ. 2017, 598, 1037–1049. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Mulder, J. Atmospheric deposition of nitrogen at five subtropical forested sites in South China. Sci. Total Environ. 2007, 378, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Gundersen, P.; Vogt, R.D.; Koba, K.; Chen, F.; Chen, X.Y.; Yoh, M. Atmospheric deposition and leaching of nitrogen in Chinese forest ecosystems. J. For. Res. 2011, 16, 341–350. [Google Scholar] [CrossRef]
- Tian, D.; Niu, S. A global analysis of soil acidification caused by nitrogen addition. Environ. Res. Lett. 2015, 10, 024019. [Google Scholar] [CrossRef]
- Smith, S.; Read, D. Mycorrhizal Symbiosis; Academic: San Diego, CA, USA, 2008. [Google Scholar] [CrossRef]
- Gruba, P.; Mulder, J. Tree species affect cation exchange capacity (CEC) and cation binding properties of organic matter in acid forest soils. Sci. Total Environ. 2015, 511, 655–662. [Google Scholar] [CrossRef]
- Sayer, E.J. Using experimental manipulation to assess the roles of leaf litter in the functioning of forest ecosystems. Biol. Rev. Camb. Philos. Soc. 2006, 81, 1–31. [Google Scholar] [CrossRef]
- Baar, J.; Ter Braak, C.J.F. Ectomycorrhizal sporocarp occurrence as affected by manipulation of litter and humus layers in scots pine stands of different age. Appl. Soil Ecol. 1996, 4, 61–73. [Google Scholar] [CrossRef]
- Mo, J.; Brown, S.; Peng, S.; Kong, G. Nitrogen availability in disturbed, rehabilitated and mature forests of tropical China. For. Ecol. Manag. 2003, 175, 573–583. [Google Scholar] [CrossRef]
- Dzwonko, Z.; Gawroński, S. Effect of litter removal on species richness and acidification of a mixed oak-pine woodland. Biol. Conserv. 2002, 106, 389–398. [Google Scholar] [CrossRef]
- Mueller, K.E.; Eissenstat, D.M.; Hobbie, S.E.; Oleksyn, J.; Jagodzinski, A.M.; Reich, P.B.; Chadwick, O.A.; Chorover, J. Tree species effects on coupled cycles of carbon, nitrogen, and acidity in mineral soils at a common garden experiment. Biogeochemistry 2012, 111, 601–614. [Google Scholar] [CrossRef]
- De Schrijver, A.; De Frenne, P.; Staelens, J.; Verstraeten, G.; Muys, B.; Vesterdal, L.; Wuyts, K.; van Nevel, L.; Schelfhout, S.; De Neve, S.; et al. Tree species traits cause divergence in soil acidification during four decades of postagricultural forest development. Glob. Chang. Biol. 2012, 18, 1127–1140. [Google Scholar] [CrossRef]
- Van Nevel, L.; Mertens, J.; De Schrijver, A.; Baeten, L.; De Neve, S.; Tack, F.M.G.; Meers, E.; Verheyen, K. Forest floor leachate fluxes under six different tree species on a metal contaminated site. Sci. Total Environ. 2013, 447, 99–107. [Google Scholar] [CrossRef]
- Schelfhout, S.; Mertens, J.; Verheyen, K.; Vesterdal, L.; Baeten, L.; Muys, B.; De Schrijver, A. Tree species identity shapes earthworm communities. Forests 2017, 8, 85. [Google Scholar] [CrossRef]
- Ma, Z.L.; Gao, S.; Yang, W.Q.; Wu, F.Z. Seasonal release characteristics of Ca, Mg and Mn of foliar litter of six tree species in subtropical evergreen broadleaved forest. Chin. J. Appl. Ecol. 2015, 26, 2913–2920. [Google Scholar]
- Finzi, A.C.; Van Breemen, N.; Canham, C.D. Canopy tree-soil interactions within temperate forests: Species effects on soil carbon and nitrogen. Ecol. Appl. 1998, 8, 440–446. [Google Scholar] [CrossRef]
- Currie, W.S.; Aber, J.D.; Mcdowell, W.H.; Boone, R.D.; Magill, A.H. Vertical transport of dissolved organic C and N under long-term N amendments in pine and hardwood forests. Biogeochemistry 1996, 35, 471–505. [Google Scholar] [CrossRef]
- Fröberg, M.; Hansson, K.; Kleja, D.B.; Alavi, G. Dissolved organic carbon and nitrogen leaching from Scots pine, Norway spruce and silver birch stands in southern Sweden. For. Ecol. Manag. 2011, 262, 1742–1747. [Google Scholar] [CrossRef]
- Lindroos, A.J.; Derome, J.; Derome, K.; Smolander, A. The effect of scots pine, norway spruce and silver birch on the chemical composition of stand throughfall and upper soil percolation water in northern Finland. Boreal Environ. Res. 2011, 16, 240–250. [Google Scholar]
- Yu, X.L. The Buffering Capacity of Acid Rain and Its Chemical Composition Research of Common Species Forest Litter Leaves in Subtropical Area. Master’s Thesis, Zhejiang A&F University, Hangzhou, China, 2011. [Google Scholar]
- Li, Z.; Chen, J.; Wang, Y.; Yu, P.; Du, S.; He, P.; Duan, J. Effects of Schima superba plantations on soil chemical properties in the acid rain region of Chongqing, southwestern China. J. Plant Ecol. 2008, 32, 632–638, (In Chinese with English Abstract). [Google Scholar]
- Wang, L.; Chen, Z.; Shang, H.; Wang, J.; Zhang, P.Y. Impact of simulated acid rain on soil microbial community function in Masson pine seedlings. Electron. J. Biotechnol. 2014, 17, 199–203. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, W.; Zhang, G.; Jiang, L.; Han, X. Mechanisms of soil acidification reducing bacterial diversity. Soil Biol. Biochem. 2015, 81, 275. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Z.; He, J.Z.; Wang, Q.; Shen, C.; Ge, Y. Ectomycorrhizal fungi inoculation alleviates simulated acid rain effects on soil ammonia oxidizers and denitrifiers in Masson pine forest. Environ. Microbiol. 2019, 21, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.Q.; Zhang, J.E.; Ouyang, Y.; Lin, L.; Quan, G.M.; Zhao, B.L.; Yu, J.Y. Effects of simulated acid rain on microbial characteristics in a lateritic red soil. Environ. Sci. Pollut. Res. 2015, 22, 18260–18266. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, B.; Zhao, W.; Wang, L.; Xie, D.; Huo, W.; Wu, Y.; Zhang, J. Comparative effects of sulfuric and nitric acid rain on litter decomposition and soil microbial community in subtropical plantation of Yangtze River Delta region. Sci. Total Environ. 2017, 601–602, 669–678. [Google Scholar] [CrossRef]
- Larssen, T.; Lydersen, E.; Tang, D.; He, Y.; Gao, J.; Liu, H.; Duan, L.; Seip, H.M.; Vogt, R.D.; Mulder, J.; et al. Acid rain in China. Environ. Sci. Technol. 2006, 40, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, Y.; Liu, Y.; Guo, H.; Li, T.; Li, Z.H.; Shi, G. Long-term effects of liming on health and growth of a Masson pine stand damaged by soil acidification in Chongqing, China. PLoS ONE 2014, 9, e94230. [Google Scholar] [CrossRef]
- Wang, Y.; Solberg, S.; Yu, P.; Myking, T.; Vogt, R.D.; Du, S. Assessments of tree crown condition of two Masson pine forests in the acid rain region in south China. For. Ecol. Manag. 2007, 242, 530–540. [Google Scholar] [CrossRef]
- Acosta-Martínez, V.; Dowd, S.; Sun, Y.; Allen, V. Tag-encoded pyrosequencing analysis of bacterial diversity in a single soil type as affected by management and land use. Soil Biol. Biochem. 2008, 40, 2762–2770. [Google Scholar] [CrossRef]
- Chen, X.Y.; Mulder, J. Indicators for nitrogen status and leaching in subtropical forest ecosystems, south China. Biogeochemistry 2007, 82, 165–180. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2006; World Soil Resources Reports No. 103; The International Union of Soil Science: Rome, Italy, 2006. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008; pp. 1–214. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Henry, M.; Stevens, H.; et al. Vegan: Community Ecology Package. R Package Version 2.0-3. 2012. Available online: http://CRAN.R-project.org/package=vegan (accessed on 8 January 2013).
- Thomas, G.W. Soil pH and Soil Acidity. In Methods of Soil Analysis: Part 3 Chemical Methods, 5.3; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, T., Sumner, M.E., Soil Science Society of America, Inc., Eds.; American Society of Agronomy, Inc.: Madison, WI, USA, 2018; pp. 475–490. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Allison, S.D.; Martiny, J.B.H. Resistance, resilience, and redundancy in microbial communities. Proc. Natl. Acad. Sci. USA 2008, 105, 11512–11519. [Google Scholar] [CrossRef] [PubMed]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; Van Der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Lejon, D.P.H.; Chaussod, R.; Ranger, J.; Ranjard, L. Microbial community structure and density under different tree species in an acid forest soil (Morvan, France). Microb. Ecol. 2005, 50, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Röling, W.F.M. Hydrocarbon-Degradation by Acidophilic Microorganisms. In Handbook of Hydrocarbon and Lipid Microbiology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1923–1930. [Google Scholar] [CrossRef]
- Singh, O.V. Extremophiles: Sustainable Resources and Biotechnological Implications; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar] [CrossRef]
- Bardavid, R.E.; Oren, A. The amino acid composition of proteins from anaerobic halophilic bacteria of the order Halanaerobiales. Extremophiles 2012, 16, 567–572. [Google Scholar] [CrossRef]
- Falagán, C.; Johnson, D.B. Acidibacter ferrireducens gen. nov., sp. nov.: An acidophilic ferric iron-reducing gammaproteobacterium. Extremophiles 2014, 18, 1067–1073. [Google Scholar] [CrossRef]
- Thrash, J.C.; Coates, J.D. Acidobacterium Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc.: Hoboken, 2015. [Google Scholar] [CrossRef]
- Kulichevskaya, I.S.; Danilova, O.V.; Tereshina, V.M.; Kevbrin, V.V.; Dedysh, S.N. Descriptions of Roseiarcus fermentans gen. nov., sp. nov., a bacteriochlorophyll a-containing fermentative bacterium related phylogenetically to alphaproteobacterial methanotrophs, and of the family Roseiarcaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 2558–2565. [Google Scholar] [CrossRef]
- Kim, K.K.; Lee, K.C.; Eom, M.K.; Kim, J.S.; Kim, D.S.; Ko, S.H.; Kim, B.H.; Lee, J.S. Variibacter gotjawalensis gen. nov., sp. nov., isolated from soil of a lava forest. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2014, 105, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.B.; Stallwood, B.; Kimura, S.; Hallberg, K.B. Isolation and characterization of Acidicaldus organivorus, gen. nov., sp. nov.: A novel sulfur-oxidizing, ferric iron-reducing thermo-acidophilic heterotrophic Proteobacterium. Arch. Microbiol. 2006, 185, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Ren, H.; Li, S.; Leng, X.; Yao, X. Soil bacterial community structure and co-occurrence pattern during vegetation restoration in karst rocky desertification area. Front. Microbiol. 2017, 8, 2377. [Google Scholar] [CrossRef]
- Stepkowski, T.; Zak, M.; Moulin, L.; Króliczak, J.; Golińska, B.; Narozna, D.; Safronova, V.I.; Madrzak, C.J. Bradyrhizobium canariense and Bradyrhizobium japonicum are the two dominant rhizobium species in root nodules of lupin and serradella plants growing in Europe. Syst. Appl. Microbiol. 2011, 34, 368–375. [Google Scholar] [CrossRef] [PubMed]
- García-Fraile, P.; Benada, O.; Cajthaml, T.; Baldrian, P.; Lladó, S. Terracidiphilus gabretensis gen. nov., sp. nov., an abundant and active forest soil acidobacterium important in organic matter transformation. Appl. Environ. Microbiol. 2016, 82, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Costa, O.Y.A.; Raaijmakers, J.M.; Kuramae, E.E. Microbial extracellular polymeric substances: Ecological function and impact on soil aggregation. Front. Microbiol. 2018, 9, 1636. [Google Scholar] [CrossRef]
- Albuquerque, L.; França, L.; Rainey, F.A.; Schumann, P.; Nobre, M.F.; Da Costa, M.S. Gaiella occulta gen. nov., sp. nov., a novel representative of a deep branching phylogenetic lineage within the class Actinobacteria and proposal of Gaiellaceae fam. nov. and Gaiellales ord. nov. Syst. Appl. Microbiol. 2011, 34, 595–599. [Google Scholar] [CrossRef]
- Severino, R.; Froufe, H.J.C.; Barroso, C.; Albuquerque, L.; Lobo-da-Cunha, A.; da Costa, M.S.; Egas, C. High-quality draft genome sequence of Gaiella occulta isolated from a 150 meter deep mineral water borehole and comparison with the genome sequences of other deep-branching lineages of the phylum Actinobacteria. Microbiologyopen 2019, 8, e00840. [Google Scholar] [CrossRef] [PubMed]
- Prosser, J.I.; Head, I.M.; Stein, L.Y. The family Nitrosomonadaceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Springer: Berlin/Heidelberg, Germany, 2014; Volume 9783642301, pp. 901–918. ISBN 9783642301971. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shannon-Wiener Index | Chao | |||
|---|---|---|---|---|
| O horizon | A horizon | O horizon | A horizon | |
| Pi | 5.52 ± 0.013d | 5.63 ± 0.030d | 1238 ± 30d | 1462 ± 64c |
| Ci | 6.26 ± 0.018a | 6.15 ± 0.012a | 2339 ± 21a | 2200 ± 33a |
| Sc | 5.79 ± 0.006c | 5.76 ± 0.019c | 1553 ± 26c | 1415 ± 7c |
| Ci_Pi | 6.17 ± 0.023b | 5.98 ± 0.044b | 2164 ± 39b | 1993 ± 35b |
| Sc_Pi | 5.47 ± 0.011d | 5.62 ± 0.023d | 1172 ± 15d | 1392 ± 32c |
| Rhodospirillales | Halanaerobiales | Acidimicrobiales | Solirubrobacterales | Acidibacter | Variibacter | Acidothermus | Rhizomicrobium | Acidobacterium | Roseiarcus | Acidicaldus | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| O horizon | |||||||||||
| Ci | 3.12c | 0.98c | 1.65c | 0.61d | 3.43b | 4.14b | 2.42c | 2.43b | 0.43e | 1.24c | 0.43c |
| Pi | 5.88a | 2.92a | 3.29a | 1.45ab | 7.46a | 7.10a | 8.15a | 3.22a | 3.17b | 1.70bc | 1.10a |
| Sc | 4.82b | 3.06a | 2.56b | 1.81a | 6.77a | 5.08b | 7.99a | 3.78a | 2.82c | 2.17ab | 0.78b |
| Ci_Pi | 2.97c | 1.73b | 2.04bc | 0.96cd | 3.50b | 5.09b | 3.23c | 3.09ab | 1.05d | 1.39c | 0.67b |
| Sc_Pi | 5.05a | 3.11a | 2.45b | 1.19bc | 8.53a | 6.30a | 6.50b | 3.77a | 3.51a | 2.40a | 0.74b |
| A horizon | |||||||||||
| Ci | 3.22c | 0.49d | 0.84c | 0.17b | 1.85c | 2.51b | 0.87c | 1.67b | 0.11c | 0.40b | 0.11c |
| Pi | 8.48a | 3.25a | 1.47ab | 0.30b | 5.91a | 3.96ab | 3.33b | 2.40a | 2.45a | 0.75ab | 0.48a |
| Sc | 5.46b | 2.16b | 1.65a | 0.28b | 3.78b | 4.94a | 4.47a | 1.60b | 1.05b | 0.82ab | 0.40ab |
| Ci_Pi | 5.29b | 1.19c | 0.96bc | 0.26b | 2.90bc | 2.87b | 1.95c | 2.00ab | 0.41c | 0.52b | 0.35b |
| Sc_Pi | 10.05a | 3.19a | 1.88a | 0.52a | 6.90a | 4.22ab | 4.03ab | 2.09ab | 2.55a | 1.08a | 0.48a |
| Bradyrhizobium canariense | Gaiellales | Chitinophagaceae | Nitrosomonadaceae | Terracidiphilus | Burkholderia | |
|---|---|---|---|---|---|---|
| O horizon | ||||||
| Ci | 4.07 ± 0.280a | 2.93 ± 0.198a | 1.26 ± 0.108a | 1.42 ± 0.036a | 1.01 ± 0.029b | 1.93 ± 0.334a |
| Pi | 1.45 ± 0.065c | 0.61 ± 0.082c | 0.58 ± 0.067b | 0.018 ± 0.007c | 0.71 ± 0.014c | 0.50 ± 0.060b |
| Sc | 2.87 ± 0.246b | 0.51 ± 0.038c | 0.53 ± 0.035b | 0.032 ± 0.011c | 0.73 ± 0.062c | 2.05 ± 0.235a |
| Ci_Pi | 4.75 ± 0.249a | 2.24 ± 0.146b | 1.49 ± 0.092a | 0.84 ± 0.067b | 1.39 ± 0.060a | 2.87 ± 0.262a |
| Sc_Pi | 0.94 ± 0.081c | 0.27 ± 0.024c | 0.56 ± 0.029b | 0.00 ± 0.00c | 0.75 ± 0.086c | 0.45 ± 0.056b |
| A horizon | ||||||
| Ci | 2.07 ± 0.063a | 0.92 ± 0.121a | 0.65 ± 0.10a | 1.81 ± 0.501a | 0.69 ± 0.048b | 0.61 ± 0.069 |
| Pi | 1.04 ± 0.107b | 0.48 ± 0.090b | 0.50 ± 0.065ab | 0.00 ± 0.00b | 1.38 ± 0.164a | 0.81 ± 0.381 |
| Sc | 1.11 ± 0.111b | 0.22 ± 0.028b | 0.25 ± 0.032b | 0.00 ± 0.00b | 1.36 ± 0.135a | 0.16 ± 0.023 |
| Ci_Pi | 2.26 ± 0.146a | 0.99 ± 0.072a | 0.63 ± 0.067a | 0.37 ± 0.099b | 1.48 ± 0.131a | 0.87 ± 0.038 |
| Sc_Pi | 0.94 ± 0.073b | 0.35 ± 0.032b | 0.28 ± 0.030b | 0.00 ± 0.00b | 1.20 ± 0.011a | 0.40 ± 0.070 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Maltz, M.R.; Zhang, Y.; O’Brien, B.J.; Neff, M.; Wang, Y.; Cao, J. Plantations of Cinnamomum camphora (Linn) Presl with Distinct Soil Bacterial Communities Mitigate Soil Acidity within Polluted Locations in Southwest China. Forests 2021, 12, 657. https://doi.org/10.3390/f12060657
Chen Z, Maltz MR, Zhang Y, O’Brien BJ, Neff M, Wang Y, Cao J. Plantations of Cinnamomum camphora (Linn) Presl with Distinct Soil Bacterial Communities Mitigate Soil Acidity within Polluted Locations in Southwest China. Forests. 2021; 12(6):657. https://doi.org/10.3390/f12060657
Chicago/Turabian StyleChen, Zhan, Mia R. Maltz, Yuguang Zhang, Brendan J. O’Brien, Monica Neff, Yihao Wang, and Jixin Cao. 2021. "Plantations of Cinnamomum camphora (Linn) Presl with Distinct Soil Bacterial Communities Mitigate Soil Acidity within Polluted Locations in Southwest China" Forests 12, no. 6: 657. https://doi.org/10.3390/f12060657
APA StyleChen, Z., Maltz, M. R., Zhang, Y., O’Brien, B. J., Neff, M., Wang, Y., & Cao, J. (2021). Plantations of Cinnamomum camphora (Linn) Presl with Distinct Soil Bacterial Communities Mitigate Soil Acidity within Polluted Locations in Southwest China. Forests, 12(6), 657. https://doi.org/10.3390/f12060657