Classification of Tree Functional Types in a Megadiverse Tropical Mountain Forest from Leaf Optical Metrics and Functional Traits for Two Related Ecosystem Functions

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
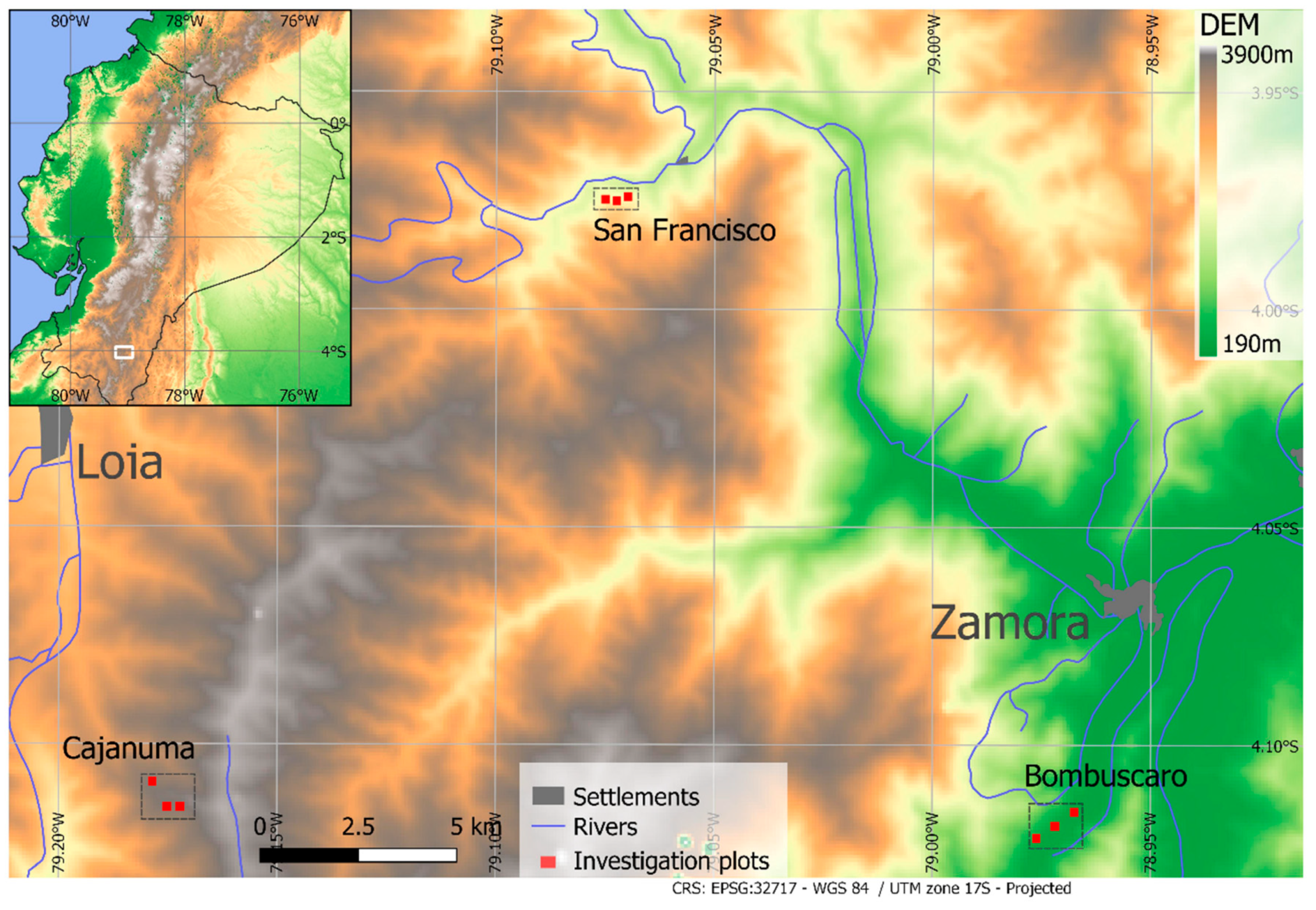
2.1. Study Area
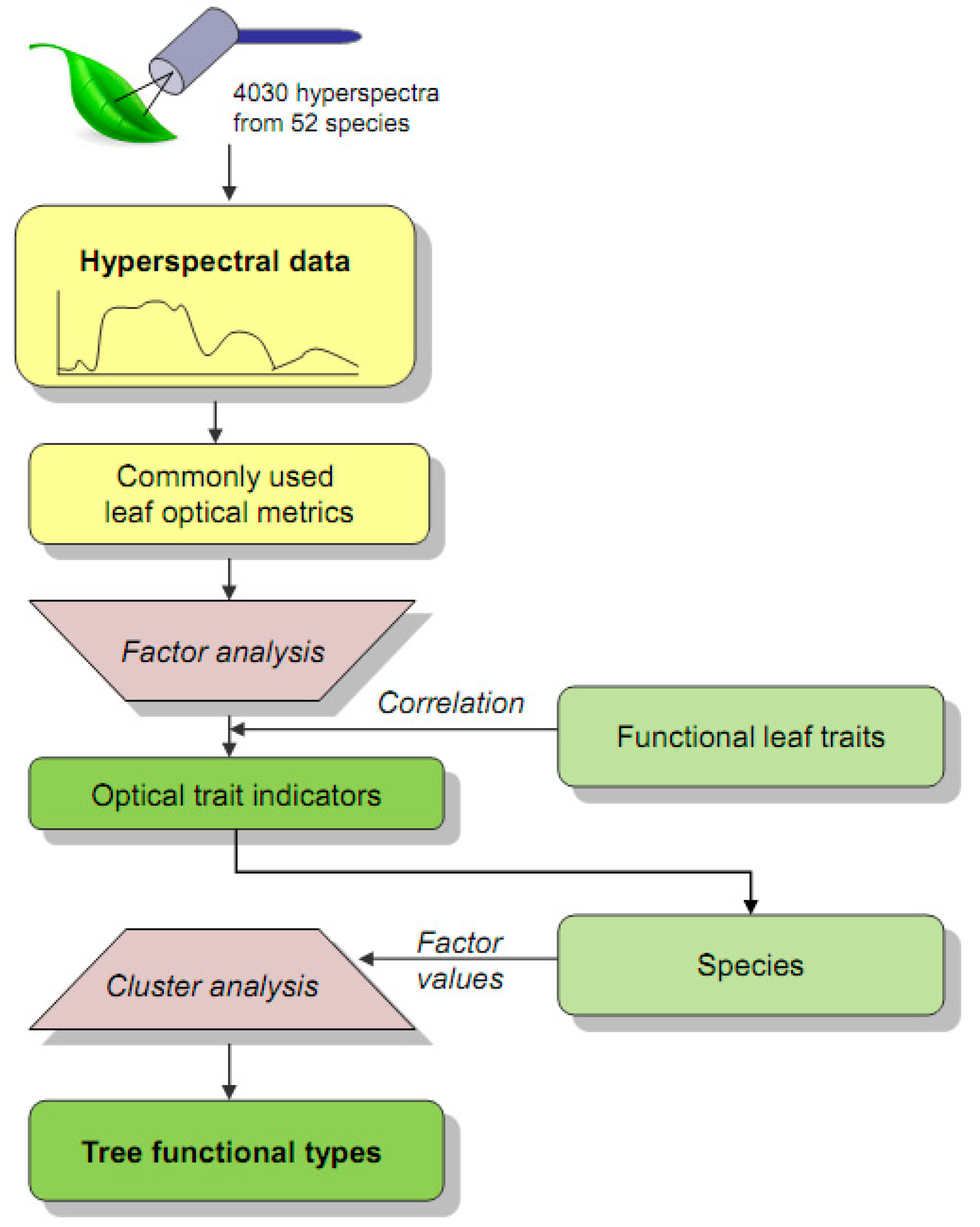
2.2. Workflow and Data
2.2.1. Leaf Spectroscopy
2.2.2. Leaf Trait Measurements
2.3. Data Analysis
2.3.1. Determination of Optical Trait Indicators (OTI) by Factor Analysis
2.3.2. Classification of tree Functional Types (TFT) by Cluster Analysis
3. Results
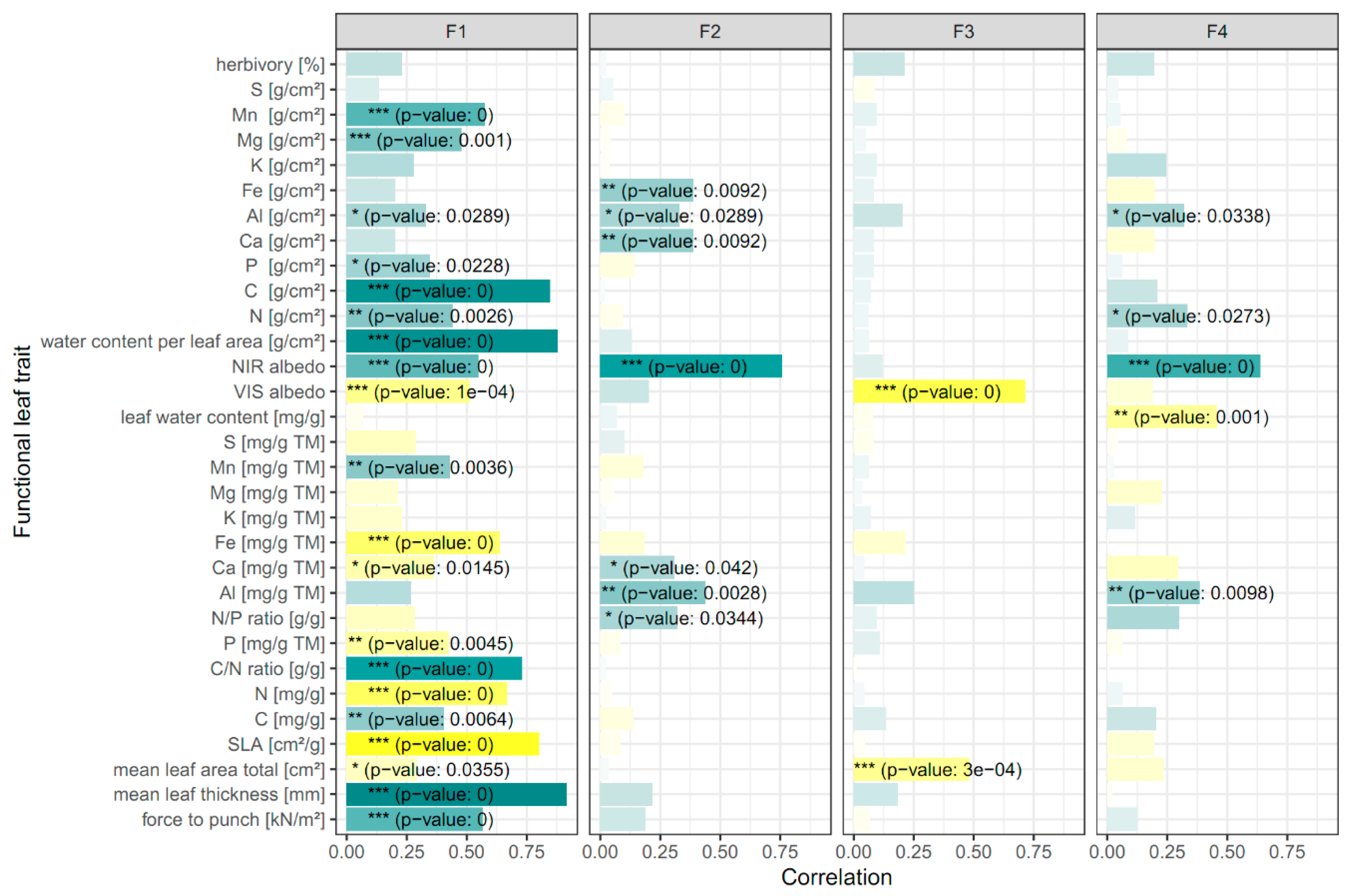
3.1. Relating Optical Metrics to Functional Traits
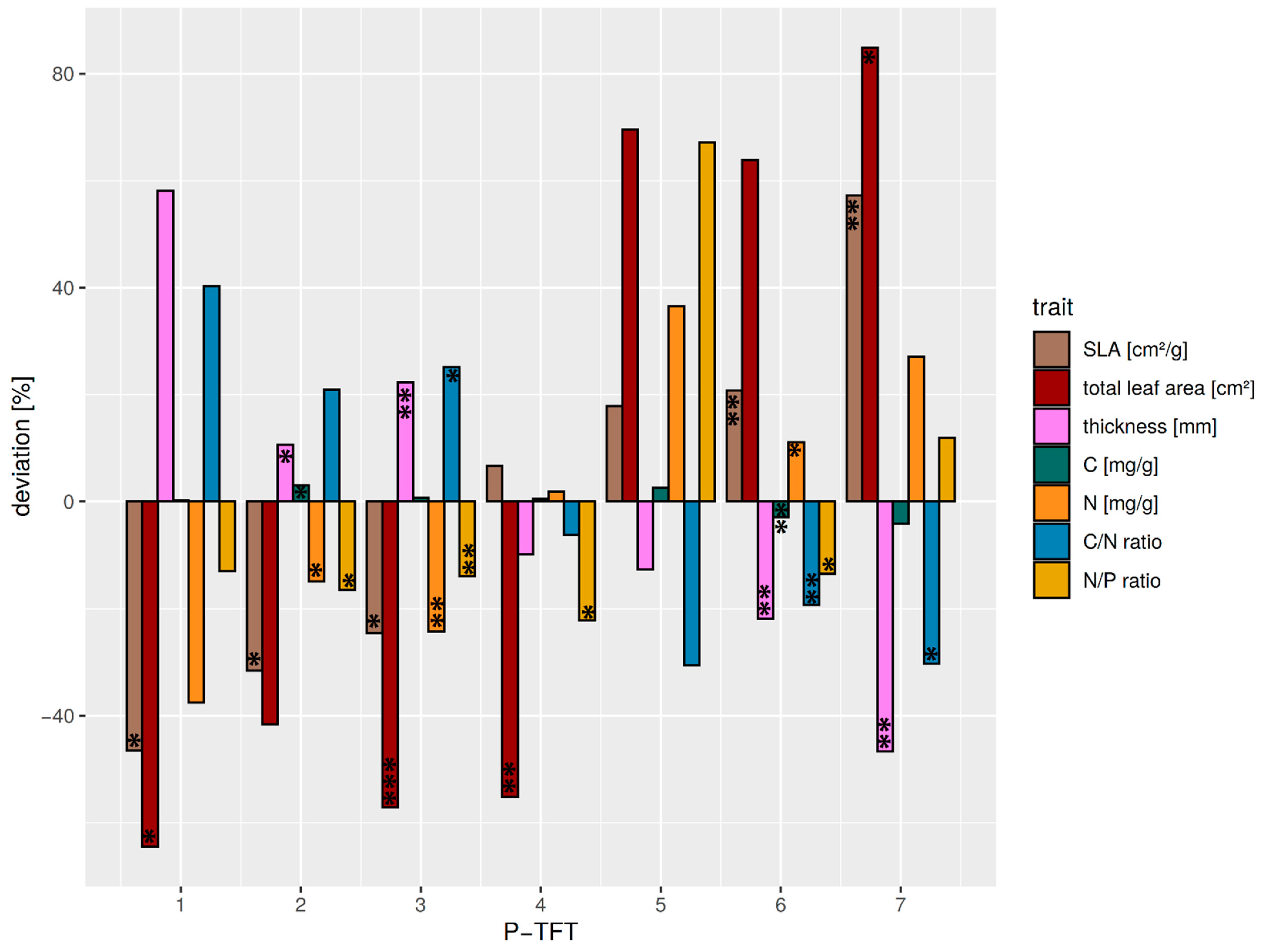
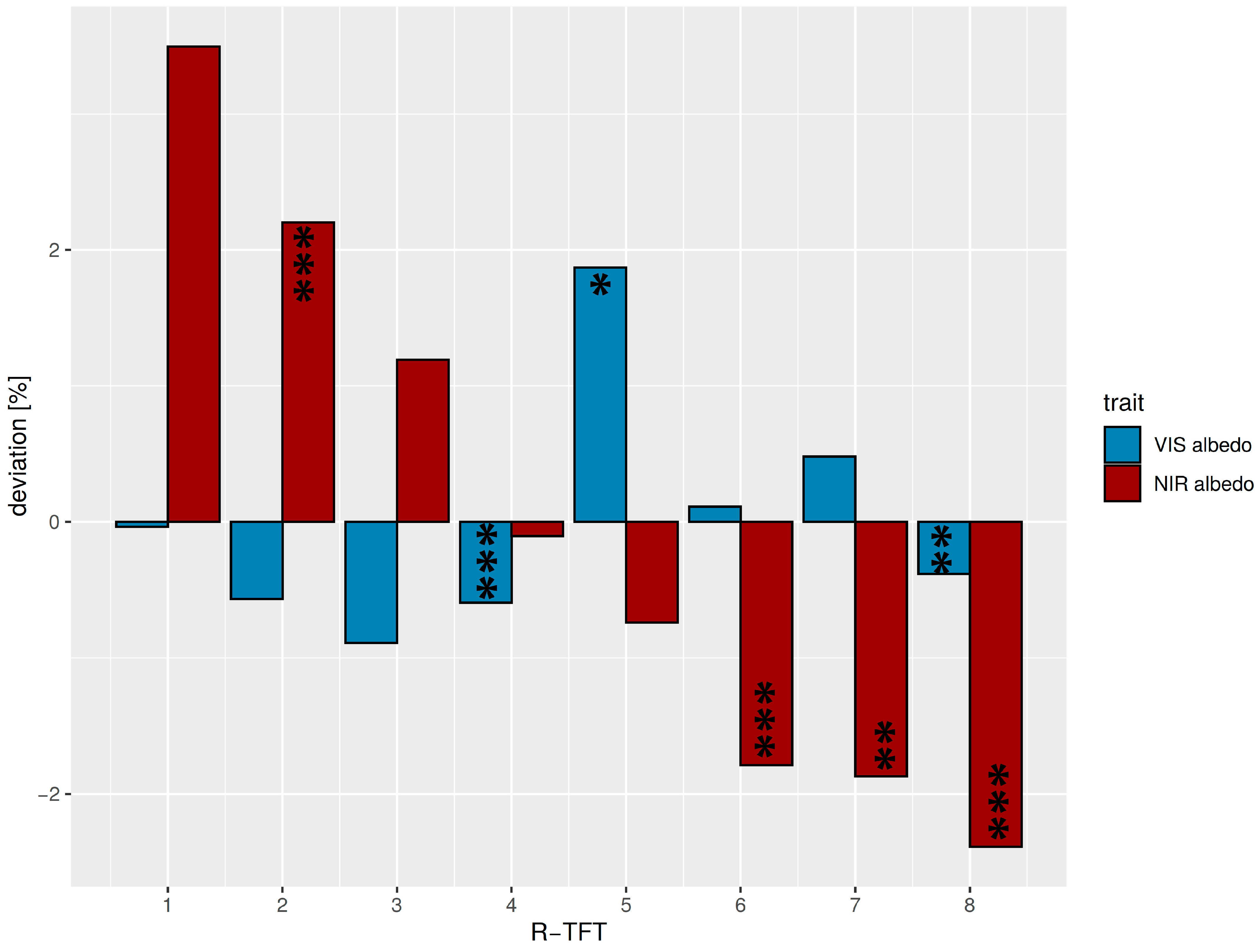
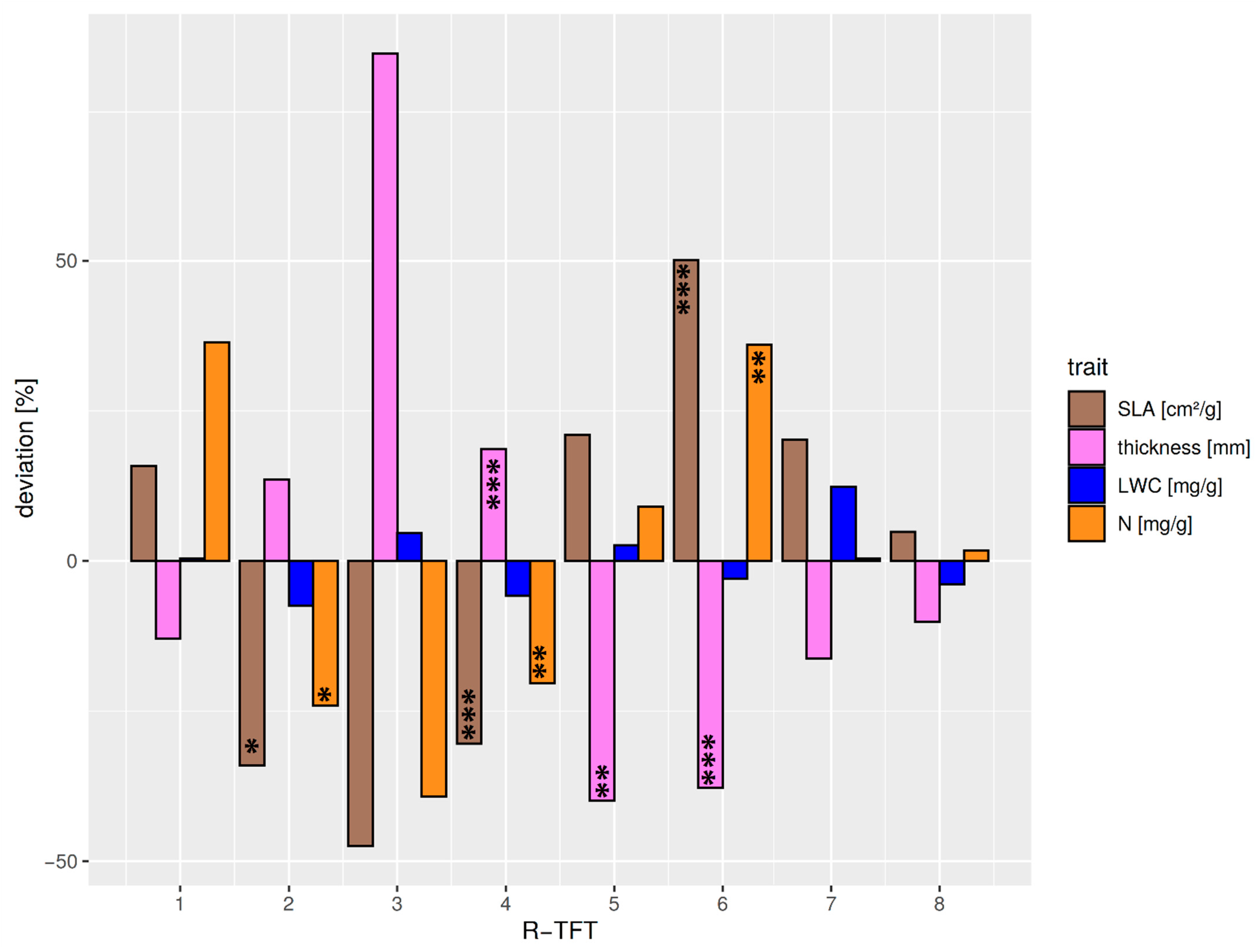
3.2. Classification of Leaf Optical Trait Indicators to Tree Functional Types (TFTs)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lavorel, S.; Díaz, S.; Cornelissen, J.H.C.; Garnier, E.; Harrison, S.P.; McIntyre, S.; Pausas, J.G.; Pérez-Harguindeguy, N.; Roumet, C.; Urcelay, C. Plant Functional Types: Are We Getting Any Closer to the Holy Grail? In Terrestrial Ecosystems in a Changing World; Springer: Berlin/Heidelberg, Germany, 2007; pp. 149–164. [Google Scholar]
- Lewis, S.L.; Lopez-Gonzalez, G.; Sonké, B.; Affum-Baffoe, K.; Baker, T.R.; Ojo, L.O.; Phillips, O.L.; Reitsma, J.M.; White, L.; Wöll, H.; et al. Increasing carbon storage in intact African tropical forests. Nature 2009, 457, 1003–1006. [Google Scholar] [CrossRef] [PubMed]
- Mitchard, E.T. The tropical forest carbon cycle and climate change. Nature 2018, 559, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Levine, N.M.; Zhang, K.; Longo, M.; Baccini, A.; Phillips, O.L.; Lewis, S.L.; Alvarez-Dávila, E.; Brienen, R.J.W.; Erwin, T.L.; Moorcroft, P.R.; et al. Ecosystem heterogeneity determines the ecological resilience of the Amazon to climate change. Proc. Natl. Acad. Sci. USA 2016, 113, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Doughty, C.E.; Santos-Andrade, P.E.; Shenkin, A.; Goldsmith, G.R.; Bentley, L.P.; Blonder, B.; Díaz, S.; Salinas, N.; Enquist, B.J.; Martin, R.E.; et al. Tropical forest leaves may darken in response to climate change. Nat. Ecol. Evol. 2018, 2, 1918–1924. [Google Scholar] [CrossRef]
- Aguilos, M.; Hérault, B.; Burban, B.; Wagner, F.; Bonal, D. What drives long-term variations in carbon flux and balance in a tropical rainforest in French Guiana? Agric. Meteorol. 2018, 253, 114–123. [Google Scholar] [CrossRef]
- Cunningham, S.A.; Summerhayes, B.; Westoby, M. Evolutionary divergences in leaf structure and chemistry, comparing rainfall and soil nutrient gradients. Ecol. Monogr. 1999, 69, 569–588. [Google Scholar] [CrossRef]
- Feng, Y.-L.; Fu, G.-L.; Zheng, Y.-L. Specific leaf area relates to the differences in leaf construction cost, photosynthesis, nitrogen allocation, and use efficiencies between invasive and noninvasive alien congeners. Planta 2008, 228, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Ustin, S. Leaf Otpical Properties; Cambridge University Press: Cambridge, UK, 2019; 555p. [Google Scholar]
- Curran, P.J. Remote sensing of foliar chemistry. Remote Sens. Environ. 1989, 30, 271–278. [Google Scholar] [CrossRef]
- Ollinger, S. Sources of variability in canopy reflectance and the convergent properties of plants. New Phytol. 2011, 189, 375–394. [Google Scholar] [CrossRef]
- Mielke, M.; Schaffer, B.; Schilling, A. Evaluation of reflectance spectroscopy indices for estimation of chlorophyll content in leaves of a tropical tree species. Photosynthetica 2012, 50, 343–352. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Knapp, D.E.; Tupayachi, R.; Anderson, C.; Carranza, L.; Martinez, P.; Houcheime, M.; Sinca, F.; Weiss, P. Spectroscopy of canopy chemicals in humid tropical forests. Remote Sens. Environ. 2011, 115, 3587–3598. [Google Scholar] [CrossRef]
- Wang, Z.; Chlus, A.; Geygan, R.; Ye, Z.; Zheng, T.; Singh, A.; Couture, J.J.; Cavender-Bares, J.; Kruger, E.L.; Townsend, P.A. Foliar functional traits from imaging spectroscopy across biomes in eastern North America. New Phytol. 2020, 228, 494–511. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro da Luz, B. Attenuated total reflectance spectroscopy of plant leaves: A tool for ecological and botanical studies. New Phytol. 2006, 172, 305–318. [Google Scholar] [CrossRef]
- Arai, E.; Pereira, G.; Coura, S.M.; Cardozo, F.S.; Silva, F.B.; Shimabukuro, Y.E.; Espirito-Santo, F.D. Spectral Signature of Leaves of Amazon Rainforest Tree Species. In Proceedings of the 2010 IEEE International Geoscience and Remote Sensing Symposium, Honolulu, HI, USA, 25–30 July 2010; IEEE: Piscataway, NJ, USA, 2010; pp. 4788–4791. [Google Scholar]
- Harrison, D.; Rivard, B.; Sanchez-Azofeifa, A. Classification of tree species based on longwave hyperspectral data from leaves, a case study for a tropical dry forest. Int. J. Appl. Earth Obs. Geoinf. 2018, 66, 93–105. [Google Scholar] [CrossRef]
- Durgante, F.M.; Higuchi, N.; Almeida, A.; Vicentini, A. Species spectral signature: Discriminating closely related plant species in the Amazon with near-infrared leaf-spectroscopy. For. Ecol. Manag. 2013, 291, 240–248. [Google Scholar] [CrossRef]
- Lang, C.; Costa, F.R.C.; Camargo, J.L.C.; Durgante, F.M.; Vicentini, A. Near infrared spectroscopy facilitates rapid identification of both young and mature Amazonian tree species. PLoS ONE 2015, 10, e0134521. [Google Scholar] [CrossRef]
- Castro-Esau, K.L.; Sánchez-Azofeifa, G.A.; Rivard, B.; Wright, S.J.; Quesada, M. Variability in leaf optical properties of Mesoamerican trees and the potential for species classification. Am. J. Bot. 2006, 93, 517–530. [Google Scholar] [CrossRef]
- Clark, M.L.; Roberts, D.A. Species-level differences in hyperspectral metrics among tropical rainforest trees as determined by a tree-based classifier. Remote Sens. 2012, 4, 1820–1855. [Google Scholar] [CrossRef]
- Ustin, S.L.; Gamon, J.A. Remote sensing of plant functional types. New Phytol. 2010, 186, 795–816. [Google Scholar] [CrossRef]
- Roth, K.L.; Casas, A.; Huesca, M.; Ustin, S.L.; Alsina, M.M.; Mathews, S.A.; Whiting, M.L. Leaf spectral clusters as potential optical leaf functional types within California ecosystems. Remote Sens. Environ. 2016, 184, 229–246. [Google Scholar] [CrossRef]
- Feilhauer, H.; Somers, B.; van der Linden, S. Optical trait indicators for remote sensing of plant species composition: Predictive power and seasonal variability. Ecol. Indic. 2017, 73, 825–833. [Google Scholar] [CrossRef]
- Kattenborn, T.; Fassnacht, F.E.; Schmidtlein, S. Differentiating plant functional types using reflectance: Which traits make the difference? Remote Sens. Ecol. Conserv. 2019, 5, 5–19. [Google Scholar] [CrossRef]
- Bendix, J.; Aguirre, N.; Beck, E.; Bräuning, A.; Brandl, R.; Breuer, L.; Boehning-Gaese, K.; Dantas De Paula, M.; Hickler, T.; Homeier, J.; et al. A research framework for projecting ecosystem change in highly diverse tropical mountain ecosystems. Oecologia 2021, 195, 589–600. [Google Scholar] [CrossRef]
- Bendix, J.; Beck, E.; Bräuning, A.; Makeschin, F.; Mosandl, R.; Scheu, S.; Wilcke, W. Ecosystem Services, Biodiversity and Environmental Change in a Tropical Mountain Ecosystem of South Ecuador; Springer Science & Business Media: Berlin, Germany, 2013; Volume 221. [Google Scholar]
- Beck, E.; Bendix, J.; Kottke, I.; Makeschin, F.; Mosandl, R. Gradients in a Tropical Mountain Ecosystem of Ecuador; Springer Science & Business Media: Berlin, Germany, 2008; Volume 198. [Google Scholar]
- Rollenbeck, R.; Bendix, J. Rainfall distribution in the Andes of southern Ecuador derived from blending weather radar data and meteorological field observations. Atmos. Res. 2011, 99, 277–289. [Google Scholar] [CrossRef]
- Fries, A.; Rollenbeck, R.; Bayer, F.; Gonzalez, V.; Onate-Valivieso, F.; Peters, T.; Bendix, J. Catchment precipitation processes in the San Francisco valley in southern Ecuador: Combined approach using high-resolution radar images and in situ observations. Meteorol. Atmos. Phys. 2014, 126, 13–29. [Google Scholar] [CrossRef]
- Giannoni, S.M.; Rollenbeck, R.; Fabian, P.; Bendix, J. Complex topography influences atmospheric nitrate deposition in a neotropical mountain rainforest. Atmos. Environ. 2013, 79, 385–394. [Google Scholar] [CrossRef]
- Makowski Giannoni, S.; Rollenbeck, R.; Trachte, K.; Bendix, J. Natural or anthropogenic? On the origin of atmospheric sulfate deposition in the Andes of southeastern Ecuador. Atmos. Chem. Phys. 2014, 11297–11312. [Google Scholar] [CrossRef]
- Werner, E.; Paladines, F.A.; Homeier, J. Is tropical montane forest heterogeneity promoted by a resource-driven feedback cycle? Evidence from nutrient relations, herbivory and litter decomposition along a topographical gradient. Funct. Ecol. 2015, 29, 430–440. [Google Scholar] [CrossRef]
- Beck, E.; Paladines, P.; Paladines, R.; Matt, F.; Farwig, N.; Bendix, J. Alexander von Humboldt would have loved it: Estación Científica San Francisco: Estación Científica San Franscisco (ECSF). ECOTROPICA 2019, 21, 201909. [Google Scholar]
- Philipson, W.R.; Gordon, D.K.; Philpot, W.D.; Duggin, M.J. Technical note Field reflectance calibration with grey standard reflectors. Int. J. Remote Sens. 1989, 10, 1035–1039. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef]
- Oleson, K.W.; Lawrence, D.M.; Bonan, G.B.; Flanner, M.G.; Kluzek, E.; Lawrence, P.J.; Levis, S.; Swenson, S.C.; Thornton, P.E.; Dai, A. Technical Description of Version 4.5 of the Community Land Model (CLM); NCAR Technical Notes (NCAR/TN-478+ STR); National Center for Atmospheric Research (NCAR): Boulder, CO, USA, 2010. [Google Scholar] [CrossRef]
- Lehmann, J.; Große-Stoltenberg, A.; Römer, M.; Oldeland, J. Field spectroscopy in the VNIR-SWIR region to discriminate between Mediterranean native plants and exotic-invasive shrubs based on leaf tannin content. Remote Sens. 2015, 7, 1225–1241. [Google Scholar] [CrossRef]
- Tucker, C. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Clark, M.L.; Roberts, D.A.; Ewel, J.J.; Clark, D.B. Estimation of tropical rain forest aboveground biomass with small-footprint lidar and hyperspectral sensors. Remote Sens. Environ. 2011, 115, 2931–2942. [Google Scholar] [CrossRef]
- Jordan, C.F. Derivation of leaf-area index from quality of light on the forest floor. Ecology 1969, 50, 663–666. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Wu, C.; Niu, Z.; Tang, Q.; Huang, W. Estimating chlorophyll content from hyperspectral vegetation indices: Modeling and validation. Agric. For. Meteorol. 2008, 148, 1230–1241. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Keydan, G.P.; Merzlyak, M.N. Three-band model for noninvasive estimation of chlorophyll, carotenoids, and anthocyanin contents in higher plant leaves. Geophys. Res. Lett. 2006, 33, L11402. [Google Scholar] [CrossRef]
- Blackburn, G.A. Quantifying chlorophylls and caroteniods at leaf and canopy scales: An evaluation of some hyperspectral approaches. Remote Sens. Environ. 1998, 66, 273–285. [Google Scholar] [CrossRef]
- Serrano, L.; Penuelas, J.; Ustin, S.L. Remote sensing of nitrogen and lignin in Mediterranean vegetation from AVIRIS data: Decomposing biochemical from structural signals. Remote Sens. Environ. 2002, 81, 355–364. [Google Scholar] [CrossRef]
- Galvao, L.S.; Formaggio, A.R.; Tisot, D.A. Discrimination of sugarcane varieties in Southeastern Brazil with EO-1 Hyperion data. Remote Sens. Environ. 2005, 94, 523–534. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Biel, C.; Serrano, L.; Save, R. The reflectance at the 950–970 nm region as an indicator of plant water status. Int. J. Remote Sens. 1993, 14, 1887–1905. [Google Scholar] [CrossRef]
- Rosso, P.H.; Pushnik, J.C.; Lay, M.; Ustin, S.L. Reflectance properties and physiological responses of Salicornia virginica to heavy metal and petroleum contamination. Environ. Pollut. 2005, 137, 241–252. [Google Scholar] [CrossRef]
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research; Northwestern University: Evanston, IL, USA, 2019. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Xu, D.; Tian, Y. A comprehensive survey of clustering algorithms. Ann. Data Sci. 2015, 2, 165–193. [Google Scholar] [CrossRef]
- Frey, B.J.; Dueck, D. Clustering by passing messages between data points. Science 2007, 315, 972–976. [Google Scholar] [CrossRef]
- Bodenhofer, U.; Kothmeier, A.; Hochreiter, S. APCluster: An R package for affinity propagation clustering. Bioinformatics 2011, 27, 2463–2464. [Google Scholar] [CrossRef]
- Ester, M.; Sander, J. Knowledge Discovery in Databases; Springer: Berlin, Germany, 2000; p. 275. [Google Scholar]
- Rousseeuw, P.J. Silhouettes: A graphical aid to the interpretation and validation of cluster analysis. J. Comput. Appl. Math. 1987, 20, 53–65. [Google Scholar] [CrossRef]
- Paul, M.J.; Foyer, C.H. Sink regulation of photosynthesis. J. Exp. Bot. 2001, 52, 1383–1400. [Google Scholar] [CrossRef]
- Güsewell, S. N:P ratios in terrestrial plants: Variation and functional significance: Tansley review. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Winter, K.; Turner, B.L. Leaf nitrogen to phosphorus ratios of tropical trees: Experimental assessment of physiological and environmental controls. New Phytol. 2010, 185, 770–779. [Google Scholar] [CrossRef]
- Abadía, J.; Morales, F.; Abadía, A. Photosystem II efficiency in low chlorophyll, iron-deficient leaves. Plant Soil 1999, 183–192. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, S.; Wan, S.; Li, X. The significance of calcium in photosynthesis. Int. J. Mol. Sci. 2019, 20, 1353. [Google Scholar] [CrossRef]
- Jiang, H.X.; Chen, L.S.; Zheng, J.G.; Han, S.; Tang, N.; Smith, B.R. Aluminum-induced effects on Photosystem II photochemistry in Citrus leaves assessed by the chlorophyll a fluorescence transient. Tree Physiol. 2008, 215, 1863–1871. [Google Scholar] [CrossRef]
- Horler, D.N.H.; Dockray, M.; Barber, J. The red edge of plant leaf reflectance. Int. J. Remote Sens. 1983, 4, 273–288. [Google Scholar] [CrossRef]
- Wagemann, J.; Thies, B.; Rollenbeck, R.; Peters, T.; Bendix, J. Regionalization of wind-speed data to analyse tree-line wind conditions in the eastern Andes of southern Ecuador. Erdkunde 2015, 64, 3–19. [Google Scholar] [CrossRef]
- Gilliham, M.; Dayod, M.; Hocking, B.J.; Xu, B.; Conn, S.J.; Kaiser, B.N.; Leigh, R.A.; Tyerman, S.D. Calcium delivery and storage in plant leaves: Exploring the link with water flow. J. Exp. Bot. 2011, 62, 2233–2250. [Google Scholar] [CrossRef]
- Basayigit, L.; Albayrak, S.; Senol, H. Analysis of VNIR reflectance for prediction of macro and micro nutrient and chlorophyll contents in apple trees (Malus communis). Asian J. Chem. 2009, 21, 1302–1308. [Google Scholar]
- Kattenborn, T.; Schmidtlein, S. Radiative transfer modelling reveals why canopy reflectance follows function. Sci. Rep. 2019, 9, 6541. [Google Scholar] [CrossRef] [PubMed]
- Croft, H.; Chen, J.M.; Luo, X.; Bartlett, P.; Chen, B.; Staebler, R.M. Leaf chlorophyll content as a proxy for leaf photosynthetic capacity. Glob. Chang. Biol. 2017, 23, 3513–3524. [Google Scholar] [CrossRef]
- Houborg, R.; Cescatti, A.; Migliavacca, M.; Kustas, W. Satellite retrievals of leaf chlorophyll and photosynthetic capacity for improved modeling of GPP. Agric. For. Meteorol. 2013, 177, 10–23. [Google Scholar] [CrossRef]
- Schull, M.; Anderson, M.; Houborg, R.; Gitelson, A.A.; Kustas, W. Thermal-based modeling of coupled carbon, water, and energy fluxes using nominal light use efficiencies constrained by leaf chlorophyll observations. Biogeosciences 2015, 12, 1511–1523. [Google Scholar] [CrossRef]
- Li, Y.; Liu, C.; Zhang, J.; Yang, H.; Xu, L.; Wang, Q.; Sack, L.; Wu, X.; Hou, J.; He, N. Variation in leaf chlorophyll concentration from tropical to cold-temperate forests: Association with gross primary productivity. Ecol. Indic. 2018, 85, 383–389. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Peng, Y.; Arkebauer, T.J.; Schepers, J. Relationships between gross primary production, green LAI, and canopy chlorophyll content in maize: Implications for remote sensing of primary production. Remote Sens. Environ. 2014, 144, 65–72. [Google Scholar] [CrossRef]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple functional roles of anthocyanins in plant-environment interactions. Environ. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Steyn, W.; Wand, S.; Holcroft, D.; Jacobs, G. Anthocyanins in vegetative tissues: A proposed unified function in photoprotection. New Phytol. 2002, 155, 349–361. [Google Scholar] [CrossRef]
- Zygielbaum, A.I.; Gitelson, A.A.; Arkebauer, T.J.; Rundquist, D.C. Non-destructive detection of water stress and estimation of relative water content in maize. Geophys. Res. Lett. 2009, 36, 1–4. [Google Scholar] [CrossRef]
- Pillar, V.D.; Sosinski, E.E., Jr. An improved method for searching plant functional types by numerical analysis. J. Veg. Sci. 2003, 14, 323–332. [Google Scholar] [CrossRef]
- Gitay, H.; Noble, I.; Connell, J. Deriving functional types for rain-forest trees. J. Veg. Sci. 1999, 10, 641–650. [Google Scholar] [CrossRef]
- Girardin, C.A.J.; Malhi, Y.; Aragao, L.E.O.C.; Mamani, M.; Huaraca Huasco, W.; Durand, L.; Feeley, K.J.; Rapp, J.; Silva-Espejo, J.E.; Whittaker, R.J.; et al. Net primary productivity allocation and cycling of carbon along a tropical forest elevational transect in the Peruvian Andes. Glob. Chang. Biol. 2010, 16, 3176–3192. [Google Scholar] [CrossRef]
- Wallis, C.I.; Homeier, J.; Peña, J.; Brandl, R.; Farwig, N.; Bendix, J. Modeling tropical montane forest biomass, productivity and canopy traits with multispectral remote sensing data. Remote Sens. Environ. 2019, 225, 77–92. [Google Scholar] [CrossRef]
- Moser, G.; Leuschner, C.; Hertel, D.; Graefe, S.; Soethe, N.; Iost, S. Elevation effects on the carbon budget of tropical mountain forests (S Ecuador): The role of the belowground compartment. Glob. Chang. Biol. 2011, 17, 2211–2226. [Google Scholar] [CrossRef]
- Shen, Y.; Gilbert, G.S.; Li, W.; Fang, M.; Lu, H.; Yu, S. Linking aboveground traits to root traits and local environment: Implications of the plant economics spectrum. Front. Plant Sci. 2019, 10, 1412. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Richardson, S.J.; Wright, E.F.; Bellingham, P.J. Environmental filtering and positive plant litter feedback simultaneously explain correlations between leaf traits and soil fertility. Ecosystems 2015, 18, 1269–1280. [Google Scholar] [CrossRef]
- Reich, P.B. The world-wide ‘fast–slow’plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Pérez-Ramos, I.M.; Roumet, C.; Cruz, P.; Blanchard, A.; Autran, P.; Garnier, E. Evidence for a ‘plant community economics spectrum’driven by nutrient and water limitations in a Mediterranean rangeland of southern France. J. Ecol. 2012, 100, 1315–1327. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Bentley, L.P.; Shenkin, A.; Asner, G.P.; Atkin, O.K.; Díaz, S.; Enquist, B.J.; Farfan-Rios, W.; Gloor, E.; Malhi, Y.; et al. Solar radiation and functional traits explain the decline of forest primary productivity along a tropical elevation gradient. Ecol. Lett. 2017, 20, 730–740. [Google Scholar] [CrossRef]
- Malhi, Y.; Girardin, C.A.; Goldsmith, G.R.; Doughty, C.E.; Salinas, N.; Metcalfe, D.B.; Huaraca Huasco, W.; Silva-Espejo, J.E.; Amézquita, F.F.; Silman, M. The variation of productivity and its allocation along a tropical elevation gradient: A whole carbon budget perspective. New Phytol. 2017, 214, 1019–1032. [Google Scholar] [CrossRef]
- Homeier, J.; Leuschner, C. Factors controlling the productivity of tropical Andean forests: Climate and soil are more important than tree diversity. Biogeosciences 2021, 18, 1525–1541. [Google Scholar] [CrossRef]
- Poorter, L.; Oberbauer, S.F.; Clark, D.B. Leaf optical properties along a vertical gradient in a tropical rain forest canopy in Costa Rica. Am. J. Bot. 1995, 82, 1257–1263. [Google Scholar] [CrossRef]
- Bendix, J.; Rollenbeck, R.; Göttlicher, D.; Cermak, J. Cloud occurence and cloud properties in Ecuador. Clim. Res. 2006, 30, 133–147. [Google Scholar] [CrossRef]
- Göttlicher, D.; Albert, J.; Nauss, T.; Bendix, J. Optical properties of selected plants from a tropical mountain ecosystem—Traits for plant functional types to parametrize a land surface model. Ecol. Model. 2011, 222, 493–502. [Google Scholar] [CrossRef]
- Wright, S.J.; Kitajima, K.; Kraft, N.J.; Reich, P.B.; Wright, I.J.; Bunker, D.E.; Condit, R.; Dalling, J.W.; Davies, S.J.; Zanne, A.E.; et al. Functional traits and the growth–mortality trade-off in tropical trees. Ecology 2010, 91, 3664–3674. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Wright, I.J.; Cavender-Bares, J.; Craine, J.M.; Oleksyn, J.; Westoby, M.; Walters, M.B. The evolution of plant functional variation: Traits, spectra, and strategies. Int. J. Plant. Sci. 2003, 164, S143–S164. [Google Scholar] [CrossRef]
- Alton, P.B. How useful are plant functional types in global simulations of the carbon, water, and energy cycles? J. Geophys. Res. Biogeosc. 2011, 116, G01030. [Google Scholar] [CrossRef]
- Lawrence, D.; Fisher, R.; Koven, C.; Oleson, K.; Swenson, S.; Vertenstein, M.; Andre, B.; Bonan, G.; Ghimire, B.; van Kampenhout, L.; et al. Technical Description of Version 5.0 of the Community Land Model (CLM); NCAR Technical Note NCAR/TN-478+ STR.; National Center for Atmospheric Research (NCAR): Boulder, CO, USA, 2018; p. 257. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Optical Metric | Formula | Absorption Bands | Functional Trait Relation | Source |
|---|---|---|---|---|
| NDVI | (R800 − R680)/(R800 + R680) | Chlorophyll | AGB (+) | Tucker (1979) [41] Clark et al. (2011) [42] |
| SR680 | R800/R680 | Chlorophyll | Chlorophyll (+) | Jordan (1969) [43] Mielke et al. (2012) [14] |
| SR705 | R750/R705 | Chlorophyll | Chlorophyll (+) | Sims and Gamon (2002) [44] Mielke et al. (2012) [14] |
| mCARI | ((R750 − R705) − 0.2 × (R750 − 550)) × (R750/705) | Chlorophyll | Chlorophyll (+) | Wu et al. (2008) [45] Mielke et al. (2012) [14] |
| SR798 | R798/R679 | Chlorophyll | AGB (+) | Clark et al. (2011) [42] |
| ARI | (1/R550) − (1/R700) | Anthocyanins | Anthocyanin (+) | Gitelson et al. (2006) [46] Mielke et al. (2012) [14] |
| BlackburnCar2 | (R804 − R484)/(R804 + R484) | Carotenoids | Carotenoids (+) | Blackburn (1998) [47] |
| GitelsonCar1 | (R484−1 − R571−1) × R746 | Carotenoids | Carotenoids (+) | Gitelson et al. (2006) [46] |
| GitelsonCar2 | (R484−1 − R689−1) × R746 | Carotenoids Chlorophyll | Carotenoids (+) | Gitelson et al. (2006) [46] |
| D1040 | D1040 | Lignin Proteins | Structural carbohydrates (+) | Curran (1987) [12] |
| D1690 | D1690 | Lignin, sugars, starch, proteins, N | Structural carbohydrates (+) | Curran (1987) [12] |
| D1420 | D1420 | Lignin | Structural carbohydrates (+) | Curran (1987) [12] |
| D1490 | D1490 | Cellulose | Structural carbohydrates (+) | Curran (1987) [12] |
| D1460 | D1460 | Sugar, starch, tannins lignin | Phenolic compounds (+) | Lehmann et al. (2015) [40] |
| NDNI | (log(1/R1510) − log(1/R1680))/(log(1/R1510) + log(1/R1680)) | N | Nitrogen (−) | Serrano et al. (2002) [48] |
| D1510 | D1510 | N | Nitrogen (−) | Curran (1987) [29] |
| D1020 | D1020 | Proteins | Nitrogen (−) | Curran (1987) [29] |
| LWVI_1 | (R1094-R983)/(R1094 + R983) | Sugar, starch, protein, water | Water content per leaf area (+) | Galvao et al. (2005) [49] |
| LWVI_2 | (R1094-R1205)/(R1094 + R1205) | Cellulose, lignin, starch, sugar | Water content per leaf area (+) | Galvao et al. (2005) [49] |
| WBI | R902/R973 | Sugar, starch | Water content per leaf area (+) | Peñuelas et al. (1993) [50] |
| D970 | D970 | water | Water content per leaf area (−) | Curran (1987) [12] |
| D1200 | D1200 | Cellulose | Water content per leaf area (−) | Curran (1987) [12] |
| D1400 | D1400 | water | Leaf water content (+) | Curran (1987) [12] |
| D1240 | D1240 | -- | Heavy metals (+) | Rosso et al. (2005) [51] |
| F1 | F2 | F3 | F4 | |
|---|---|---|---|---|
| NDVI | 0.10 | 0.86 | 0.19 | 0.12 |
| SR680 | 0.09 | 0.83 | 0.21 | 0.14 |
| SR705 | 0.23 | 0.64 | 0.17 | 0.14 |
| mCARI | −0.27 | 0.94 | −0.32 | 0.06 |
| SR798 | 0.09 | 0.83 | 0.22 | 0.14 |
| ARI | 0.22 | −0.42 | 0.78 | 0.07 |
| BlackburnCar2 | 0.23 | 0.14 | 0.85 | 0.05 |
| GitelsonCar1 | −0.22 | 0.13 | 0.93 | 0.01 |
| GitelsonCar2 | −0.19 | 0.08 | 0.98 | 0.00 |
| D1040 | 0.92 | 0.02 | −0.02 | 0.21 |
| D1690 | 0.82 | −0.07 | −0.03 | 0.37 |
| D1420 | 0.70 | 0.01 | −0.05 | 0.08 |
| D1490 | 0.57 | 0.29 | 0.08 | 0.38 |
| D1460 | 0.23 | 0.34 | 0.12 | 0.65 |
| NDNI | 1.00 | 0.03 | −0.02 | −0.15 |
| D1510 | 0.88 | 0.24 | 0.06 | −0.15 |
| D1020 | 0.89 | 0.15 | 0.00 | 0.17 |
| LWVI_1 | 0.96 | −0.07 | −0.05 | 0.00 |
| LWVI_2 | 0.99 | 0.15 | −0.01 | −0.14 |
| WBI | 0.97 | −0.29 | 0.02 | −0.11 |
| D970 | −0.30 | −0.83 | −0.06 | 0.24 |
| D1200 | −0.72 | −0.02 | −0.06 | −0.26 |
| D1400 | 0.04 | 0.04 | −0.03 | −0.97 |
| D1240 | 0.06 | −0.12 | −0.05 | −0.67 |
| TFT No. | No. Species | F1 | F2 | Avg. within Cluster Distance | Avg. between Cluster Distance | Avg. Silhouette Width | Productivity |
|---|---|---|---|---|---|---|---|
| P1 | 3 | 1.95 | 1.51 | 0.78 | 2.45 | 0.49 | very low |
| P2 | 8 | 0.56 | −0.48 | 0.33 | 1.75 | 0.65 | low |
| P3 | 11 | 1.04 | 0.22 | 0.60 | 1.81 | 0.35 | low, P-limited |
| P4 | 5 | 0.28 | −1.70 | 0.53 | 2.44 | 0.62 | intermediate |
| P5 | 1 | −0.06 | 3.38 | NA | 3.17 | 0.00 | high, P-limited |
| P6 | 15 | −0.78 | 0.32 | 0.71 | 1.88 | 0.41 | high |
| P7 | 9 | −1.22 | −0.64 | 0.75 | 2.24 | 0.36 | very high |
| R-TFT No. | No. of Species | F1 | F2 | F3 | F4 | Avg. within Cluster Distance | Avg. between Cluster Distance | Avg. Silhouette Width | Albedo Difference |
|---|---|---|---|---|---|---|---|---|---|
| R1 | 1 | −0.06 | 3.38 | 1.33 | 2.40 | NA | 4.17 | 0.00 | very high NIR intermediate VIS |
| R2 | 5 | 0.80 | 0.82 | 0.68 | 1.61 | 1.05 | 2.87 | 0.49 | high NIR low VIS |
| R3 | 1 | 2.17 | 2.04 | 0.47 | −0.80 | NA | 3.40 | 0.00 | high NIR very low VIS |
| R4 | 15 | 0.80 | −0.33 | 0.30 | 0.10 | 1.28 | 2.56 | 0.27 | intermediate NIR low VIS |
| R5 | 4 | −1.00 | 0.33 | −2.30 | −0.66 | 2.22 | 3.67 | 0.21 | low NIR very high VIS |
| R6 | 12 | −0.93 | 0.20 | 0.77 | 0.02 | 1.50 | 2.82 | 0.33 | very low NIR intermediate VIS |
| R7 | 9 | −0.68 | 0.10 | −0.47 | −1.36 | 1.67 | 2.99 | 0.26 | very low NIR high VIS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Limberger, O.; Homeier, J.; Farwig, N.; Pucha-Cofrep, F.; Fries, A.; Leuschner, C.; Trachte, K.; Bendix, J. Classification of Tree Functional Types in a Megadiverse Tropical Mountain Forest from Leaf Optical Metrics and Functional Traits for Two Related Ecosystem Functions. Forests 2021, 12, 649. https://doi.org/10.3390/f12050649
Limberger O, Homeier J, Farwig N, Pucha-Cofrep F, Fries A, Leuschner C, Trachte K, Bendix J. Classification of Tree Functional Types in a Megadiverse Tropical Mountain Forest from Leaf Optical Metrics and Functional Traits for Two Related Ecosystem Functions. Forests. 2021; 12(5):649. https://doi.org/10.3390/f12050649
Chicago/Turabian StyleLimberger, Oliver, Jürgen Homeier, Nina Farwig, Franz Pucha-Cofrep, Andreas Fries, Christoph Leuschner, Katja Trachte, and Jörg Bendix. 2021. "Classification of Tree Functional Types in a Megadiverse Tropical Mountain Forest from Leaf Optical Metrics and Functional Traits for Two Related Ecosystem Functions" Forests 12, no. 5: 649. https://doi.org/10.3390/f12050649
APA StyleLimberger, O., Homeier, J., Farwig, N., Pucha-Cofrep, F., Fries, A., Leuschner, C., Trachte, K., & Bendix, J. (2021). Classification of Tree Functional Types in a Megadiverse Tropical Mountain Forest from Leaf Optical Metrics and Functional Traits for Two Related Ecosystem Functions. Forests, 12(5), 649. https://doi.org/10.3390/f12050649