
Contrasting Litter Nutrient and Metal Inputs and Soil Chemistry among Five Common Eastern North American Tree Species


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Plot Construction and Field Methods
2.3. Litter and Soil Analyses
2.4. Data Analysis
3. Results
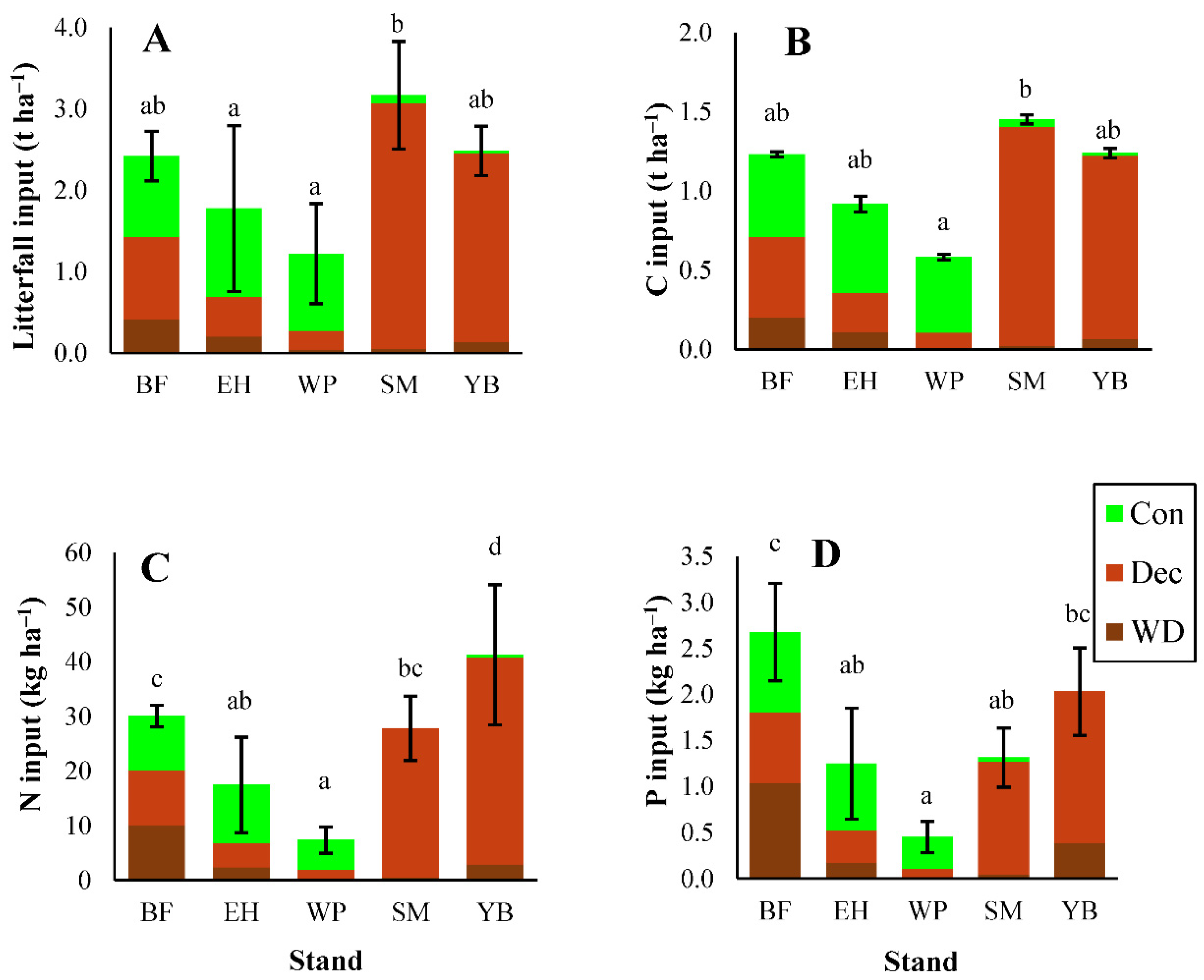
3.1. Litterfall Characteristics
3.1.1. Litterfall Mass
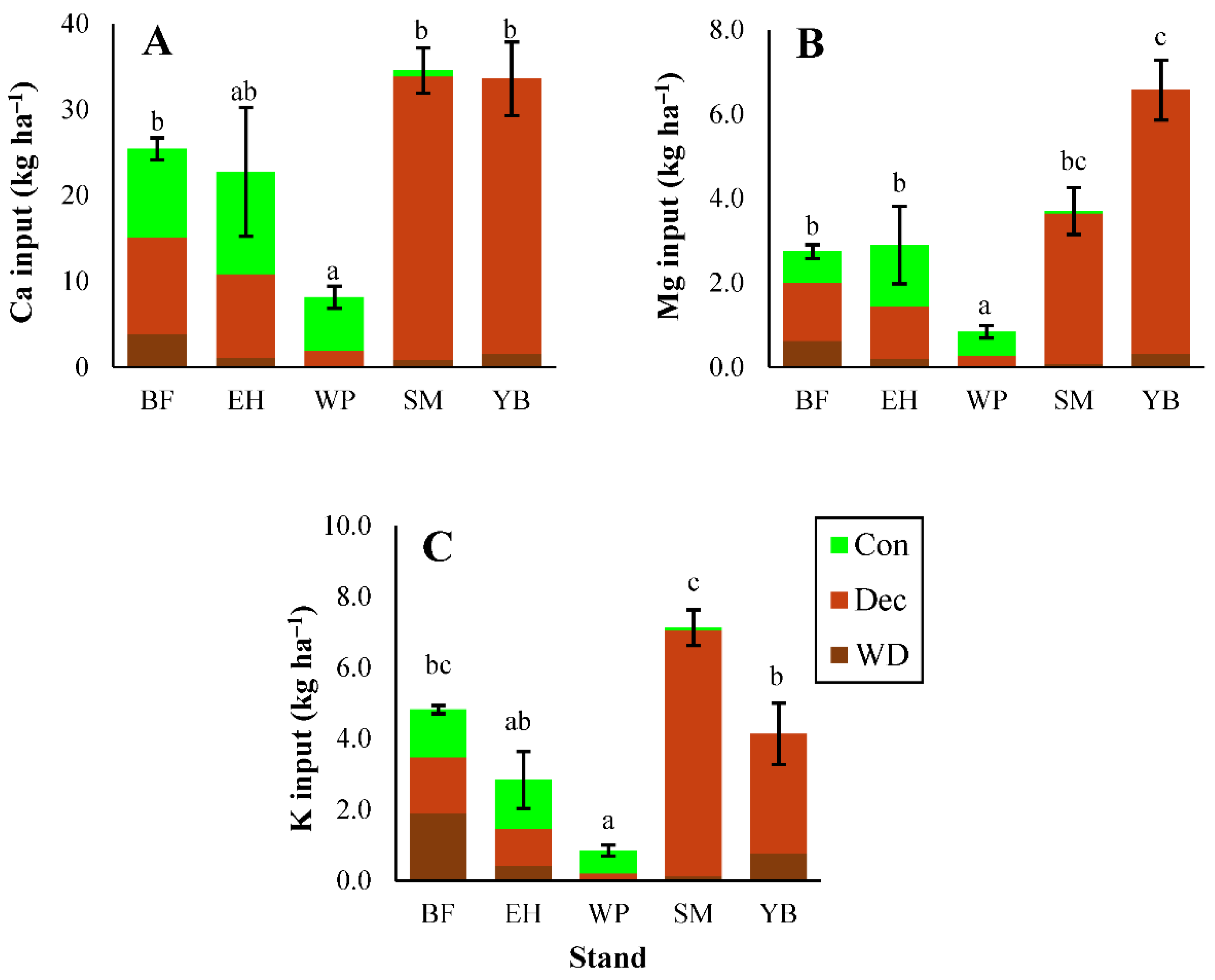
3.1.2. Carbon and Macronutrients
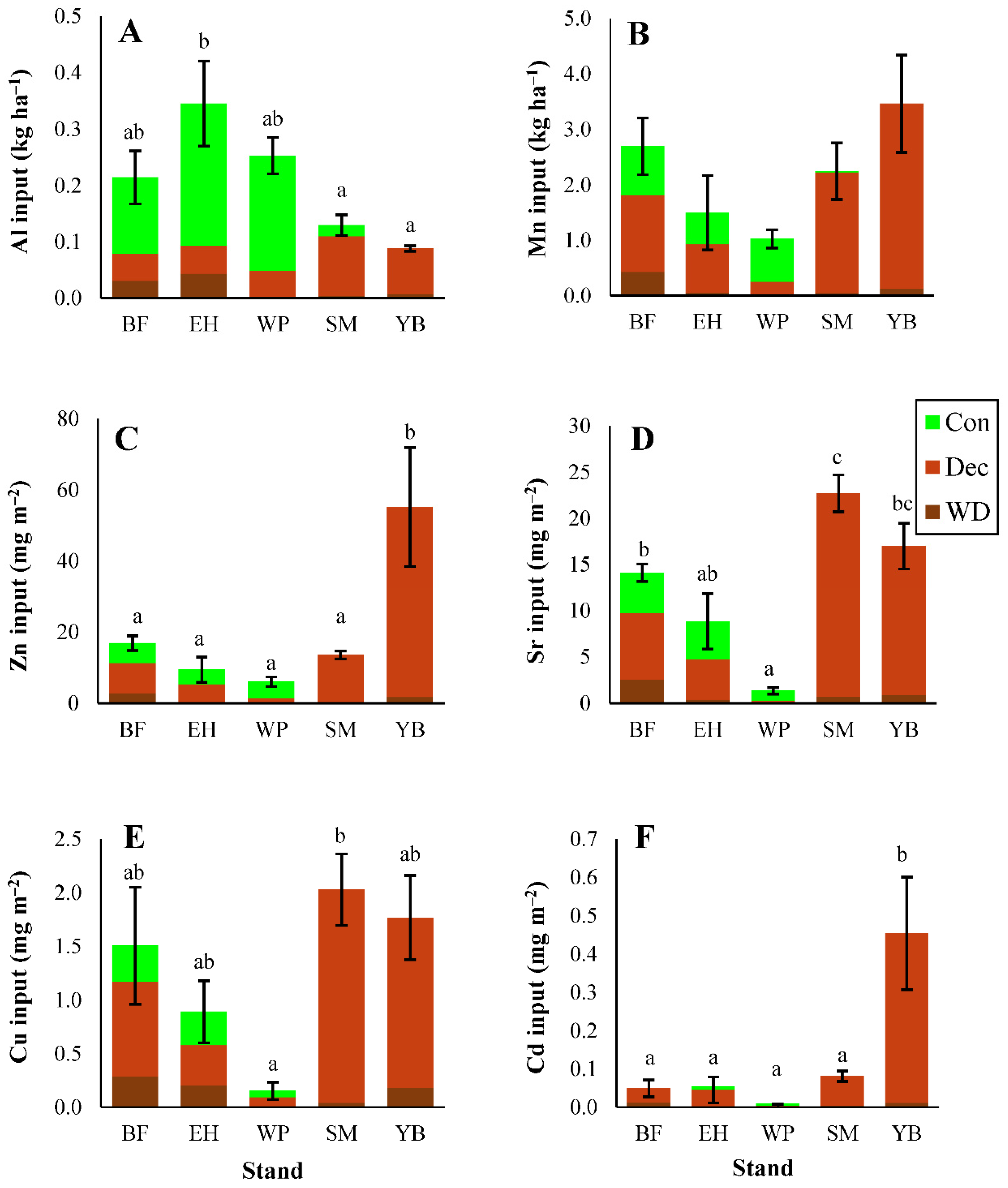
3.1.3. Micronutrients and Trace Metals
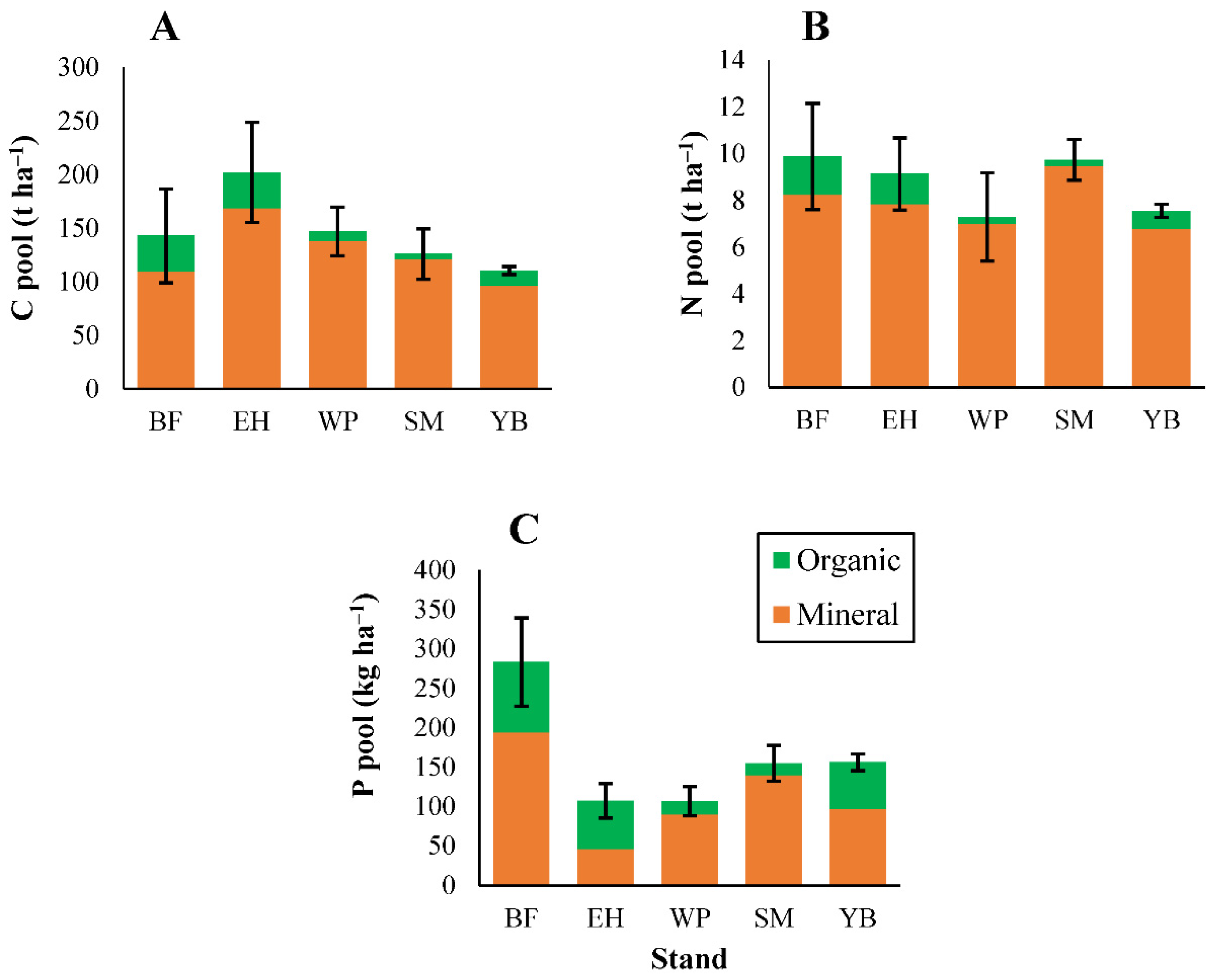
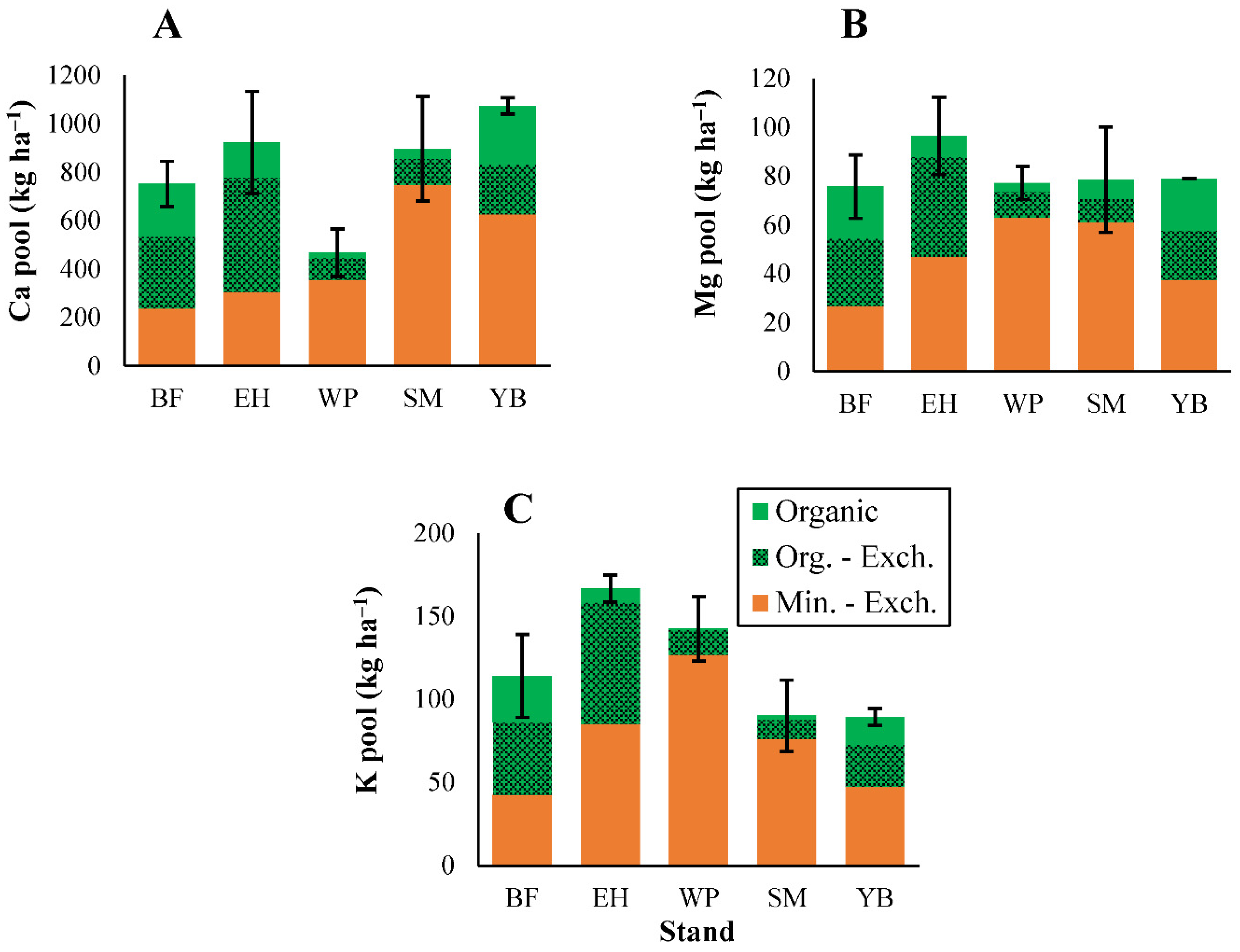
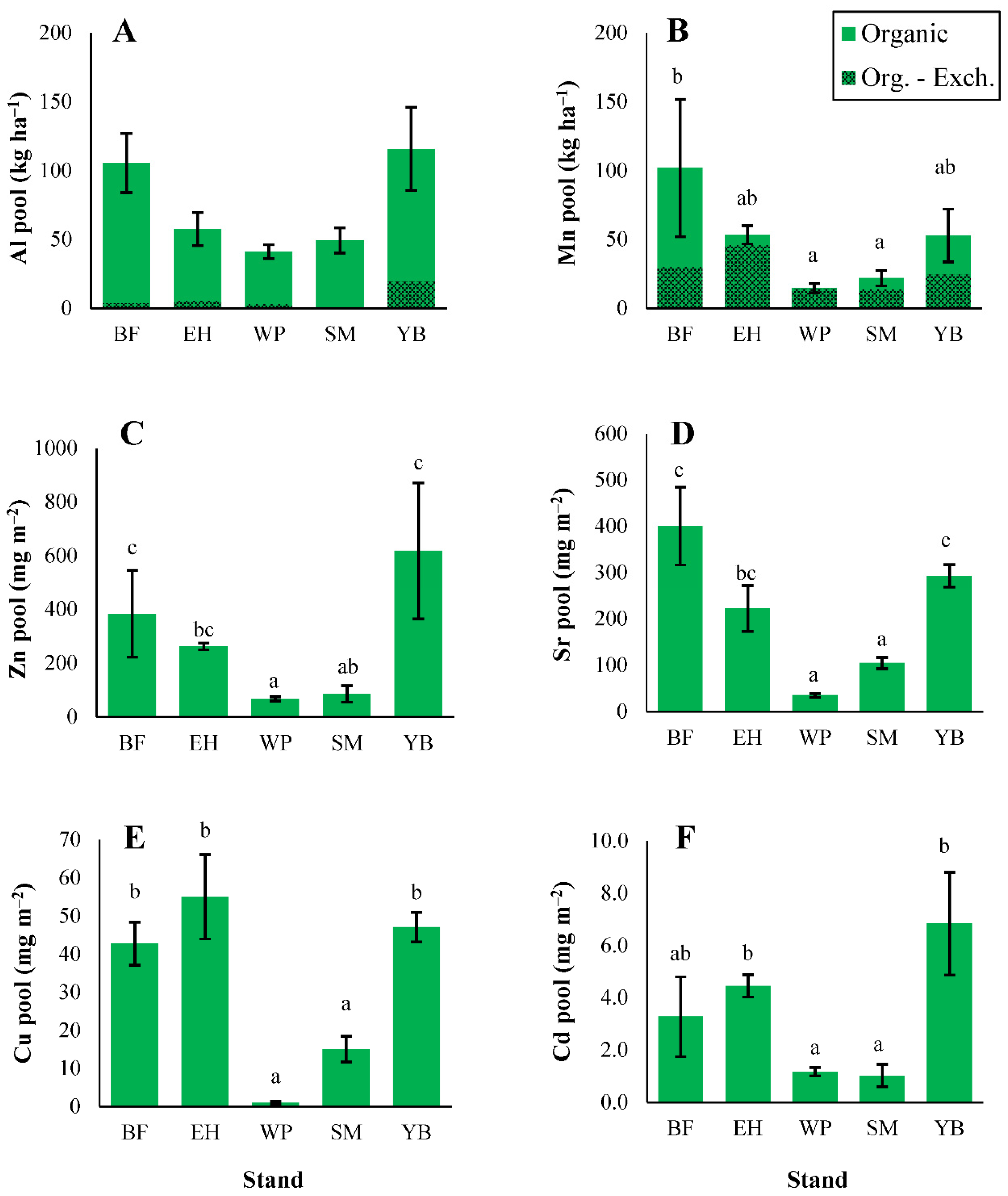
3.2. Soil Properties
3.2.1. Physical and Chemical Properties
3.2.2. Carbon and Macronutrients
3.2.3. Micronutrients and Trace Metals
3.2.4. Organic Horizon Elemental Residence Times
4. Discussion
4.1. Litterfall Chemistry
4.2. Soil Chemistry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boucher, Y.; Arseneault, D.; Sirois, L. Logging–induced change (1930–2002) of a preindustrial landscape at the northern range limit of northern hardwoods, eastern Canada. Can. J. For. Res. 2006, 36, 505–517. [Google Scholar] [CrossRef]
- Dieler, J.; Uhl, E.; Biber, P.; Müller, J.; Rötzer, T.; Pretzsch, H. Effect of forest stand management on species composition, structural diversity, and productivity in the temperate zone of Europe. Eur. J. For. Res. 2017, 136, 739–766. [Google Scholar] [CrossRef]
- Coates, T.A.; Johnson, A.; Aust, W.M.; Hagan, D.L.; Chow, A.T.; Trettin, C. Forest composition, fuel loading, and soil chemistry resulting from 50 years of forest management and natural disturbance in two southeastern coastal plain watersheds, USA. Forest Ecol. Manag. 2020, 473, 118337. [Google Scholar] [CrossRef]
- Leadbitter, P.; Euler, D.; Naylor, B. A comparison of historical and current forest cover in selected areas of the Great Lakes–St. Lawrence forest of central Ontario. For. Chron. 2002, 78, 522–529. [Google Scholar] [CrossRef]
- Pinto, F.; Romaniuk, S.; Ferguson, M. Changes to preindustrial forest tree composition in central and northeastern Ontario, Canada. Can. J. For. Res. 2008, 38, 1842–1854. [Google Scholar] [CrossRef]
- Thompson, I.D.; Simard, J.H.; Titman, R.D. Historical changes in white pine (Pinus strobus l.) density in Algonquin Park, Ontario, during the 19th century. Nat. Areas J. 2006, 26, 61–71. [Google Scholar] [CrossRef]
- Ontario Ministry of Natural Resources and Forestry (OMNRF). Forest Management Guide to Silviculture in the Great Lakes–St. Lawrence and Boreal Forests of Ontario; Queens Printer for Ontario: Toronto, ON, Canada, 2015; pp. 1–394.
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of Macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: London, UK, 2012; pp. 135–189. [Google Scholar] [CrossRef]
- El–Jaoual, T.; Cox, D.A. Manganese toxicity in plants. J. Plant Nutr. 1998, 21, 353–386. [Google Scholar] [CrossRef]
- Cronan, C.S.; Grigal, D.F. Use of calcium/aluminum ratios as indicators of stress in forest ecosystems. J. Environ. Qual. 1995, 24, 209–226. [Google Scholar] [CrossRef]
- Smith, T.M.; Smith, R.L. Elements of Ecology, 8th ed.; Pearson: San Francisco, CA, USA, 2012; pp. 92–118. [Google Scholar] [CrossRef]
- Morrison, I.K. Addition of organic matter and elements to the forest floor of an old–growth Acer saccharum forest in the annual litter fall. Can. J. For. Res. 1991, 21, 462–468. [Google Scholar] [CrossRef]
- Landre, A.L.; Watmough, S.A.; Dillon, P.J. Metal pools, fluxes, and budgets in an acidified forested catchment on the Precambrian Shield, Central Ontario, Canada. Water Air Soil Pollut. 2010, 209, 209–228. [Google Scholar] [CrossRef]
- Attiwill, P.M. The loss of elements from decomposing litter. Ecology 1968, 49, 142–145. [Google Scholar] [CrossRef]
- Gosz, J.R.; Likens, G.E.; Bormann, F.H. Nutrient content of litter fall on the Hubbard Brook Experimental Forest, New Hampshire. Ecology 1972, 53, 769–784. [Google Scholar] [CrossRef]
- Van Hook, R.I.; Harris, W.F.; Henderson, G.S. Cadmium, lead, and zinc distributions and cycling in a mixed deciduous forest. Ambio 1977, 6, 281–286. [Google Scholar]
- Bigelow, S.W.; Canham, C.D. Litterfall as a niche construction process in a northern hardwood forest. Ecosphere 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Finzi, A.C.; Canham, C.D.; Van Breemen, N. Canopy tree–soil interactions within temperate forests: Species effects on pH and cations. Ecol. App. 1998, 8, 447–454. [Google Scholar] [CrossRef]
- Andersen, M.K.; Raulund–Rasmussen, K.; Strobel, B.W.; Hansen, H.C.B. The effects of tree species and site on the solubility of Cd, Cu, Ni, Pb and Zn in soils. Water Air Soil Pollut. 2004, 154, 357–370. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Geibe, C.; Holmström, S.; Lundström, U.S.; Van Breemen, N. The effect of organic acids on base cation leaching from the forest floor under six North American tree species. Eur. J. Soil Sci. 2001, 52, 205–214. [Google Scholar] [CrossRef]
- Van Breemen, N.; Finzi, A.C.; Canham, C.D. Canopy tree–soil interactions within temperate forests: Effects of soil elemental composition and texture on species distributions. Can. J. For. Res. 1997, 27, 1110–1116. [Google Scholar] [CrossRef]
- Richardson, J.B.; Friedland, A.J. Influence of coniferous and deciduous vegetation on major and trace metals in forests of northern New England, USA. Plant Soil 2016, 402, 363–378. [Google Scholar] [CrossRef]
- Horton, K.W.; Brown, W.G.E. Ecology of White and Red Pine in the Great Lakes–St. Lawrence Forest Region; Department of Northern Affairs and National Resources: Ottawa, ON, Canada, 1960; pp. 1–22. [Google Scholar]
- Remmel, T. An Introduction to the Algonquin Park Ecosystem. In Algonquin Park: The Human Impact; Euler, D., Wilton, M., Eds.; Algonquin Eco Watch: Espanola, ON, Canada, 2009; pp. 14–35. [Google Scholar]
- Melvin, A.M.; Mack, M.C.; Johnstone, J.F.; McGuire, A.D.; Genet, H.; Schuur, E.A.G. Differences in ecosystem carbon distribution and nutrient cycling linked to forest tree species composition in a mid-successional boreal forest. Ecosystems 2015, 18, 1472–1488. [Google Scholar] [CrossRef]
- Lovett, G.M.; Mitchell, M.J. Sugar maple and nitrogen cycling in the forests of eastern North America. Front. Ecol. Environ. 2004, 2, 81–88. [Google Scholar] [CrossRef]
- Hallett, R.A.; Bailey, S.W.; Horsley, S.B.; Long, R.P. Influence of nutrition and stress on sugar maple at a regional scale. Can. J. For. Res. 2006, 36, 2235–2246. [Google Scholar] [CrossRef]
- Environment and Climate Change Canada. Canadian Climate Normals 1981–2010 Station Data. Available online: https://climate.weather.gc.ca/climate_normals/results_1981_2010_e.html?searchType=stnName&txtStationName=haliburton&searchMethod=contains&txtCentralLatMin=0&txtCentralLatSec=0&txtCentralLongMin=0&txtCentralLongSec=0&stnID=5170&dispBack=1 (accessed on 29 March 2021).
- Soil Classification Working Group. The Canadian System of Soil Classification, 3rd ed.; Agriculture and Agri–Food Can. Publication 1646 (Revised); NRC Research Press: Ottawa, ON, Canada, 1998; pp. 1–187. [Google Scholar]
- Dillon, P.J.; Watmough, S.A.; Eimers, M.C.; Aherne, J. Long–term changes in boreal lake and stream chemistry: Recovery from acid deposition and the role of climate. In Acid in the Environment; Visgilio, G.R., Whitelaw, D.M., Eds.; Springer: New York, NY, USA, 2007; pp. 59–76. [Google Scholar] [CrossRef]
- Ferrari, J.B. Fine-scale patterns of leaf litterfall and nitrogen cycling in an old-growth forest. Can. J. For. Res. 1999, 29, 291–302. [Google Scholar] [CrossRef]
- Gosz, J.R.; Likens, G.E.; Bormann, F.H. Organic matter and nutrient dynamics of the forest and forest floor in the Hubbard Brook Forest. Oecologia 1976, 22, 305–320. [Google Scholar] [CrossRef] [PubMed]
- Real Statistics Using Excel. Available online: www.real–statistics.com (accessed on 20 October 2020).
- Peichl, M.; Arain, M.A. Above–and belowground ecosystem biomass and carbon pools in an age–sequence of temperate pine plantation forests. Agric. For. Meteor. 2006, 140, 51–63. [Google Scholar] [CrossRef]
- Yanai, R.D.; Arthur, M.A.; Acker, M.; Levine, C.R.; Park, B.B. Variation in Mass and Nutrient Concentration of Leaf Litter across Years and Sites in a Northern Hardwood Forest. Can. J. For. Res. 2012, 42, 1597–1610. [Google Scholar] [CrossRef]
- Hendrickson, O.Q.; Burgess, D.M.; Chatarpaul, L. Biomass and nutrients in Great Lakes–St. Lawrence forest species: Implications for whole–tree and conventional harvest. Can. J. For. Res. 1987, 17, 210–218. [Google Scholar] [CrossRef]
- Neumann, M.; Ukonmaanaho, L.; Johnson, J.; Benham, S.; Vesterdal, L.; Novotný, R.; Verstraeten, A.; Lundin, L.; Thimonier, A.; Michopoulos, P.; et al. Quantifying carbon and nutrient input from litterfall in European forests using field observations and modeling. Glob. Biogeochem. Cycles 2018, 32, 784–798. [Google Scholar] [CrossRef]
- Likens, G.E.; Driscoll, C.T.; Buso, D.C.; Siccama, T.G.; Johnson, C.E.; Fahey, T.J.; Reiners, W.A.; Ryan, D.F.; Martin, C.W.; Bailey, S.W. The biogeochemistry of calcium at Hubbard Brook. Biogeochemistry 1998, 41, 89–173. [Google Scholar] [CrossRef]
- Watmough, S.A.; Dillon, P.J. Base cation losses from a coniferous catchment in central Ontario, Canada. Water Air Soil Pollut. Focus. 2001, 1, 507–524. [Google Scholar] [CrossRef]
- Côté, B.; Fyles, J.W. Nutrient concentration and acid–base status of leaf litter of tree species characteristic of the hardwood forest of southern Quebec. Can. J. For. Res. 1994, 24, 192–196. [Google Scholar] [CrossRef]
- Morrison, I.K. Effect of crown position on foliar concentrations of 11 elements in Acer saccharum and Betula alleghaniensis trees on a till soil. Can. J. For. Res. 1985, 15, 179–183. [Google Scholar] [CrossRef]
- Munro, L.; Courchesne, F. Bioconcentration of Cd and Zn in the soils of an uncontaminated forest in the Quebec Laurentians. Biogeochemistry 2019, 143, 293–312. [Google Scholar] [CrossRef]
- Gosz, J.R.; Likens, G.E.; Bormann, F.H. Nutrient release from decomposing leaf and branch litter in the Hubbard Brook Forest, New Hampshire. Ecol. Monogr. 1973, 43, 173–191. [Google Scholar] [CrossRef]
- Phillips, T.; Watmough, S.A. A nutrient budget for a selection harvest: Implications for long–term sustainability. Can. J. For. Res. 2012, 42, 2064–2077. [Google Scholar] [CrossRef]
- Likens, G.E.; Bormann, F.H. Chemical Analyses of Plant Tissues from the Hubbard Brook Ecosystem in New Hampshire (Bulletin 79); Yale School of Forestry & Environmental Studies Bulletin Series; Yale University: New Haven, CT, USA, 1970; pp. 1–25. [Google Scholar]
- McGee, C.J.; Fernandez, I.J.; Norton, S.A.; Stubbs, C.S. Cd, Ni, Pb, and Zn Concentrations in forest vegetation and soils in Maine. Water Air Soil Pollut. 2007, 180, 141–153. [Google Scholar] [CrossRef]
- Crim, P.M.; McDonald, L.M.; Cumming, J.R. Soil and tree nutrient status of high elevation mixed red spruce (Picea rubens Sarg.) and broadleaf deciduous forests. Soil Syst. 2019, 3, 80. [Google Scholar] [CrossRef]
- Ukonmaanaho, L.; Merilä, P.; Nöjd, P.; Nieminen, T.M. Litterfall production and nutrient return to the forest floor in Scots pine and Norway spruce stands in Finland. Boreal Environ. Res. 2008, 13, 67–91. [Google Scholar]
- Schroth, A.W.; Friedland, A.J.; Bostick, B.C. Macronutrient Depletion and Redistribution in Soils under Conifer and Northern Hardwood Forests. Soil Sci. Soc. Am. J. 2007, 71, 457–468. [Google Scholar] [CrossRef]
- Freedman, B. Environmental Ecology: The Impacts of Pollution and Other Stresses on Ecosystem Structure and Function; Academic Press: San Diego, CA, USA, 1989; pp. 1–424. [Google Scholar] [CrossRef]
- Li, W.; Johnson, C.E. Relationships among pH, aluminum solubility and aluminum complexation with organic matter in acid forest soils of the Northeastern United States. Geoderma 2016, 271, 234–242. [Google Scholar] [CrossRef]
- Lawrence, G.B.; David, M.B.; Shortle, W.C. A new mechanism for calcium loss in forest–floor soils. Nature 1995, 378, 162–165. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Smits, M.M. Tree species effects on calcium cycling: The role of calcium uptake in deep soils. Ecosystems 2002, 5, 385–398. [Google Scholar] [CrossRef]
- Burgess–Conforti, J.R.; Moore, P.A.; Owens, P.R.; Miller, D.M.; Ashworth, A.J.; Hays, P.D.; Evans–White, M.A.; Anderson, K.R. Are soils beneath coniferous tree stands more acidic than soils beneath deciduous tree stands? Environ. Sci. Pollut. Res. 2019, 26, 14920–14929. [Google Scholar] [CrossRef]
- Finzi, A.C.; Van Breemen, N.; Canham, C.D. Canopy tree–soil interactions within temperate forests: Species effects on soil carbon and nitrogen. Ecol. App. 1998, 8, 440–446. [Google Scholar] [CrossRef]
- Lovett, G.M.; Weathers, K.C.; Arthur, M.A.; Schultz, J.C. Nitrogen cycling in a northern hardwood forest: Do species matter? Biogeochemistry 2004, 67, 289–308. [Google Scholar] [CrossRef]
- Pastor, J.; Aber, J.D.; Mcclaugherty, C.A.; Melillo, J.M. Aboveground production and N and p cycling along a nitrogen mineralization gradient on Blackhawk Island, Wisconsin. Ecology 1984, 65, 256–268. [Google Scholar] [CrossRef]
- Fassnacht, K.S.; Gower, S.T. Comparison of the litterfall and forest floor organic matter and nitrogen dynamics of upland forest ecosystems in north central Wisconsin. Biogeochemistry 1999, 45, 265–284. [Google Scholar] [CrossRef]
- Gradowski, T.; Thomas, S.C. phosphorus limitation of sugar maple growth in central Ontario. For. Ecol. Manag. 2006, 226, 104–109. [Google Scholar] [CrossRef]
- Fujinuma, R.; Bockheim, J.G.; Balster, N. Base–cation cycling by individual tree species in old–growth forests of Upper Michigan, USA. Biogeochemistry 2005, 74, 357–376. [Google Scholar] [CrossRef]
- Maclean, D.A.; Wein, R.W. Litter production and forest floor nutrient dynamics in pine and hardwood stands of New Brunswick, Canada. Holarct. Ecol. 1978, 1, 1–15. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Number | Balsam Fir | Eastern Hemlock | White Pine | Sugar Maple | Yellow Birch |
|---|---|---|---|---|---|
| 1 | BF 79%, SM 21% | EH 76% WC 23% BF 1% | WP 95% AB 4% EH 2% | SM 65% BF 35% | YB 95% BF 5% |
| 2 | BF 58% SM 24% YB 19% | EH 100% | WP 74% WB 12% Other 11% BF 3% | SM 81% WB 15% AE 4% | YB 93% WB 7% |
| 3 | BF 53% SM 21% BA 26% | EH 100% | WP 83% BF 17% | SM 93% YB 5% BF 2% | YB 87% BF 13% |
| 4 | -- | -- | WP 100% | SM 92% BC 5% YB 1% WB 1% BF 1% | -- |
| 5 | -- | -- | WP 100% | SM 100% | -- |
| Horizon | Parameter | Balsam Fir | Eastern Hemlock | White Pine | Sugar Maple | Yellow Birch | p Value |
|---|---|---|---|---|---|---|---|
| L | Bulk Density (kg m−3) | 31 (15) | 34 (10) | 18 (4) | 39 (14) | 12 (5) | 0.468 |
| Mass (kg m−2) | 0.2 (0.1) | 0.2 (0.1) | 0.5 (0.1) | 0.2 (0.1) | 0.1 (<0.1) | 0.163 | |
| pH | 4.8 ab (0.2) | 4.5 a (0.2) | 4.5 a (<0.1) | 5.0 b (0.1) | 5.2 b (0.3) | <0.001 | |
| %OM | 92.4 ab (0.3) | 95.2 a (0.1) | 94.4 ab (1.0) | 87.3 b (1.5) | 91.6 ab (0.4) | <0.05 | |
| FH | Bulk Density (kg m−3) | 116 (22) | 54 (8) | 41 (16) | 89 (18) | 89 (16) | 0.060 |
| Mass (kg m−2) | 7.6 a (1.9) | 8.4 a (1.9) | 1.8 b (0.7) | 1.8 b (0.4) | 3.2 ab (0.9) | <0.01 | |
| pH | 4.4 (<0.1) | 4.4 (0.1) | 4.3 (0.1) | 4.9 (0.1) | 4.3 (0.1) | 0.055 | |
| %OM | 82.8 (7.5) | 93.3 (1.2) | 82.5 (4.5) | 75.5 (2.6) | 76.1 (13.5) | 0.238 | |
| Ah | Bulk Density (kg m−3) | 813 ab (127) | 793 ab (50) | 434 a (104) | 524 ab (51) | 915 b (100) | <0.05 |
| Mass (kg m−2) | 52.3 ab (11.2) | 70.0 a (19.3) | 21.7 b (5.2) | 26.2 b (2.5) | 50.8 ab (8.2) | <0.01 | |
| pH | 4.1 ab (0.3) | 3.7 a (<0.1) | 3.9 ab (0.2) | 4.7 b (0.1) | 4.2 ab (0.1) | <0.05 | |
| %OM | 5.0 a (2.0) | 4.1 a (0.9) | 20.1 b (0.3) | 12.5 ab (3.0) | 5.9 a (1.0) | <0.01 | |
| Bm1 | Bulk Density (kg m−3) | 976 (212) | 944 (29) | 699 (84) | 803 (49) | 1018 (114) | 0.309 |
| Mass (kg m−2) | 66.3 (16.6) | 81.2 (35.5) | 55.9 (7.5) | 64.3 (3.9) | 61.1 (5.6) | 0.761 | |
| pH | 4.6 a (0.1) | 4.1 b (<0.1) | 4.1 b (0.1) | 4.8 a (0.1) | 4.5 ab (<0.1) | <0.001 | |
| %OM | 6.3 (2.5) | 8.5 (2.4) | 11.9 (2.0) | 8.3 (3.0) | 7.5 (1.1) | 0.610 | |
| Bm2 | Bulk Density (kg m−3) | 1157 (121) | 990 (64) | 830 (72) | 953 (227) | 890 (165) | 0.677 |
| Mass (kg m−2) | 309.0 (32.3) | 270.3 (17.4) | 266.8 (63.5) | 266.9 (40.3) | 232.5 (20.1) | 0.812 | |
| pH | 4.7 ab (0.1) | 4.2 a (<0.1) | 4.3 ab (0.1) | 4.8 b (0.1) | 4.7 ab (0.1) | <0.05 | |
| %OM | 5.6 (1.8) | 8.9 (1.0) | 7.8 (1.7) | 6.8 (1.4) | 5.2 (0.2) | 0.818 |
| Oxide | Balsam Fir | Eastern Hemlock | White Pine | Sugar Maple | Yellow Birch | p Value |
|---|---|---|---|---|---|---|
| Units = % of soil | ||||||
| SiO2 | 66.8 (1.8) | 64.7 (1.7) | 62.4 (3.5) | 60.7 (4.6) | 59.7 (2.6) | 0.535 |
| Al2O3 | 13.0 (0.2) | 13.2 (0.2) | 13.1 (0.3) | 13.2 (3.4) | 13.1 (0.4) | 0.987 |
| Fe2O3 | 3.8 bc (0.3) | 3.5 c (0.3) | 5.6 ab (0.8) | 3.5 c (0.2) | 5.8 a (0.2) | <0.001 |
| MgO | 0.9 (<0.1) | 0.9 (<0.1) | 1.1 (0.2) | 1.1 (0.1) | 1.1 (0.1) | 0.113 |
| CaO | 2.4 (0.1) | 2.2 (0.1) | 2.4 (0.1) | 2.5 (0.1) | 2.3 (0.1) | 0.236 |
| Cr2O3 | 0.1 (<0.1) | 0.1 (<0.1) | 0.1 (<0.1) | 0.1 (<0.1) | 0.1 (<0.1) | 0.355 |
| K2O | 2.6 (0.1) | 2.5 (<0.1) | 2.6 (0.2) | 2.5 (0.2) | 2.4 (0.1) | 0.372 |
| Na2O | 3.0 (0.1) | 2.9 (0.1) | 2.8 (0.1) | 2.8 (0.2) | 2.6 (0.1) | 0.115 |
| MnO | 0.5 a (<0.1) | 0.6 a (<0.1) | 0.7 a (<0.1) | 0.6 a (<0.1) | 0.9 b (0.1) | <0.001 |
| TiO2 | 0.1 (<0.1) | 0.1 (<0.1) | 0.2 (0.1) | 0.1 (<0.1) | 0.1 (0.1) | 0.052 |
| P2O5 | 0.2 (<0.1) | 0.1 (<0.1) | 0.2 (0.1) | 0.2 (0.1) | 0.1 (<0.1) | 0.146 |
| V2O5 | 0.1 (<0.1) | 0.1 (<0.1) | 0.1 (<0.1) | 0.1 (<0.1) | 0.1 (<0.1) | 0.061 |
| C/N or Element | Balsam Fir | Eastern Hemlock | White Pine | Sugar Maple | Yellow Birch | p Value | Mean | Median |
|---|---|---|---|---|---|---|---|---|
| Units = years | ||||||||
| C/N–L | 25 a | 37 ab | 40 b | 26 a | 23 a | <0.05 | 30 | 26 |
| C/N–FH | 20 a | 26 b | 26 b | 19 a | 18 a | <0.001 | 22 | 20 |
| Mass | 32 b | 90 c | 19 a | 6.2 a | 12 ab | <0.001 | 32 | 19 |
| C | 26 b | 82 c | 15 b | 3.6 a | 10 ab | <0.001 | 27 | 15 |
| N | 55 b | 105 c | 43 b | 9.9 a | 16 ab | <0.001 | 51 | 55 |
| Ca | 20 ab | 44 b | 17 ab | 4.1 a | 13 ab | <0.05 | 20 | 17 |
| Mg | 18 ab | 26 ab | 34 b | 4.3 a | 6.9 a | <0.01 | 18 | 18 |
| K | 15 ab | 46 b | 22 ab | 2.0 a | 12 ab | <0.05 | 19 | 15 |
| P | 35 ab | 67 b | 12 a | 44 ab | 31 ab | <0.05 | 38 | 35 |
| Al | 624 ab | 182 a | 173 a | 540 ab | 1340 b | <0.01 | 572 | 540 |
| Mn | 34 ab | 78 b | 17 a | 11 a | 15 a | <0.05 | 31 | 17 |
| Zn | 34 ab | 80 b | 24 a | 10 a | 13 a | <0.01 | 32 | 24 |
| Sr | 29 ab | 42 b | 30 ab | 4.6 a | 18 ab | <0.05 | 25 | 29 |
| Cu | 40 ab | 107 b | 17 ab | 7.6 a | 32 ab | <0.05 | 41 | 32 |
| Cd | 62 ab | 158 ab | 242 b | 16 a | 19 a | <0.05 | 99 | 62 |
| Mean | 83 | 82 | 58 | 55 | 127 | |||
| Median | 34 | 76 | 23 | 8.6 | 17 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ott, N.F.J.; Watmough, S.A. Contrasting Litter Nutrient and Metal Inputs and Soil Chemistry among Five Common Eastern North American Tree Species. Forests 2021, 12, 613. https://doi.org/10.3390/f12050613
Ott NFJ, Watmough SA. Contrasting Litter Nutrient and Metal Inputs and Soil Chemistry among Five Common Eastern North American Tree Species. Forests. 2021; 12(5):613. https://doi.org/10.3390/f12050613
Chicago/Turabian StyleOtt, Neil F. J., and Shaun A. Watmough. 2021. "Contrasting Litter Nutrient and Metal Inputs and Soil Chemistry among Five Common Eastern North American Tree Species" Forests 12, no. 5: 613. https://doi.org/10.3390/f12050613
APA StyleOtt, N. F. J., & Watmough, S. A. (2021). Contrasting Litter Nutrient and Metal Inputs and Soil Chemistry among Five Common Eastern North American Tree Species. Forests, 12(5), 613. https://doi.org/10.3390/f12050613