
Short-Term N-Fertilization Differently Affects the Leaf and Leaf Litter Chemistry of the Dominant Species in a Mediterranean Forest under Drought Conditions


 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Sampling
2.4. Chemical Analyses
2.5. Statistical Analyses
3. Results
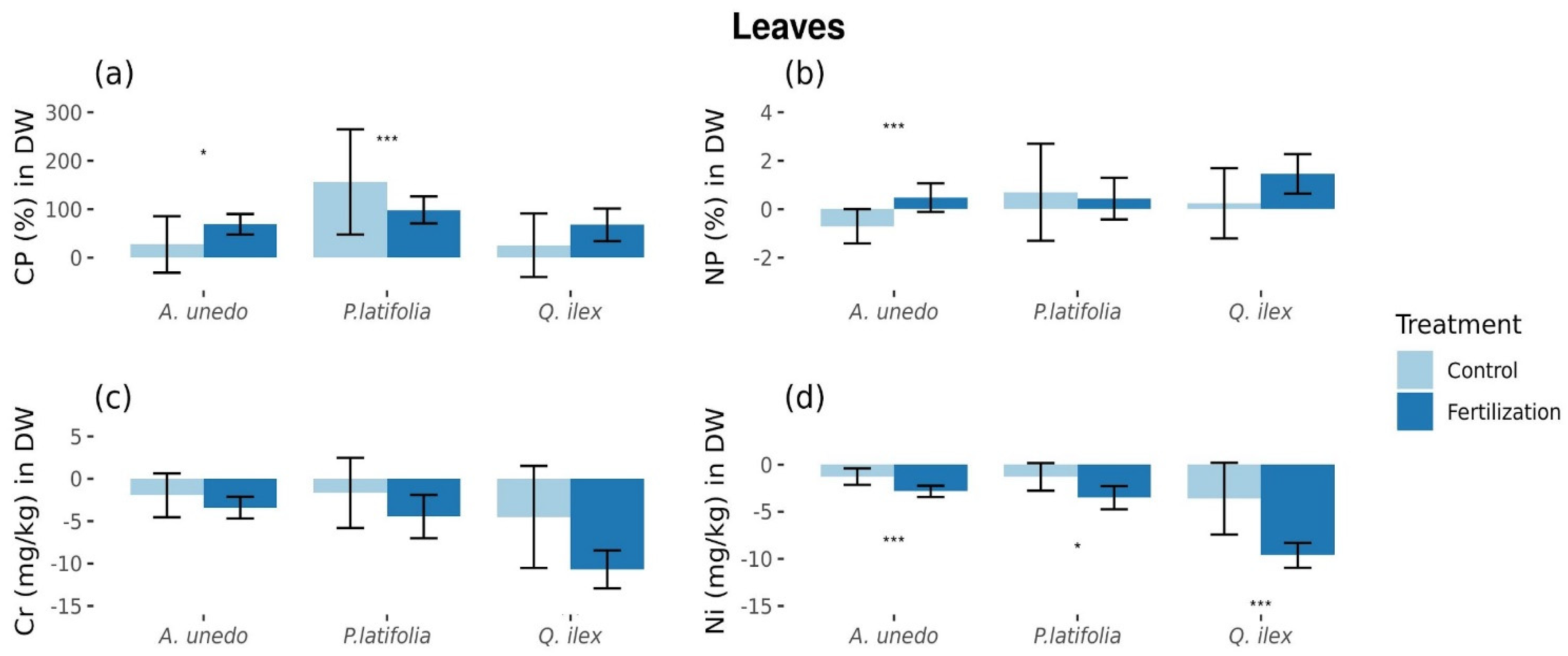
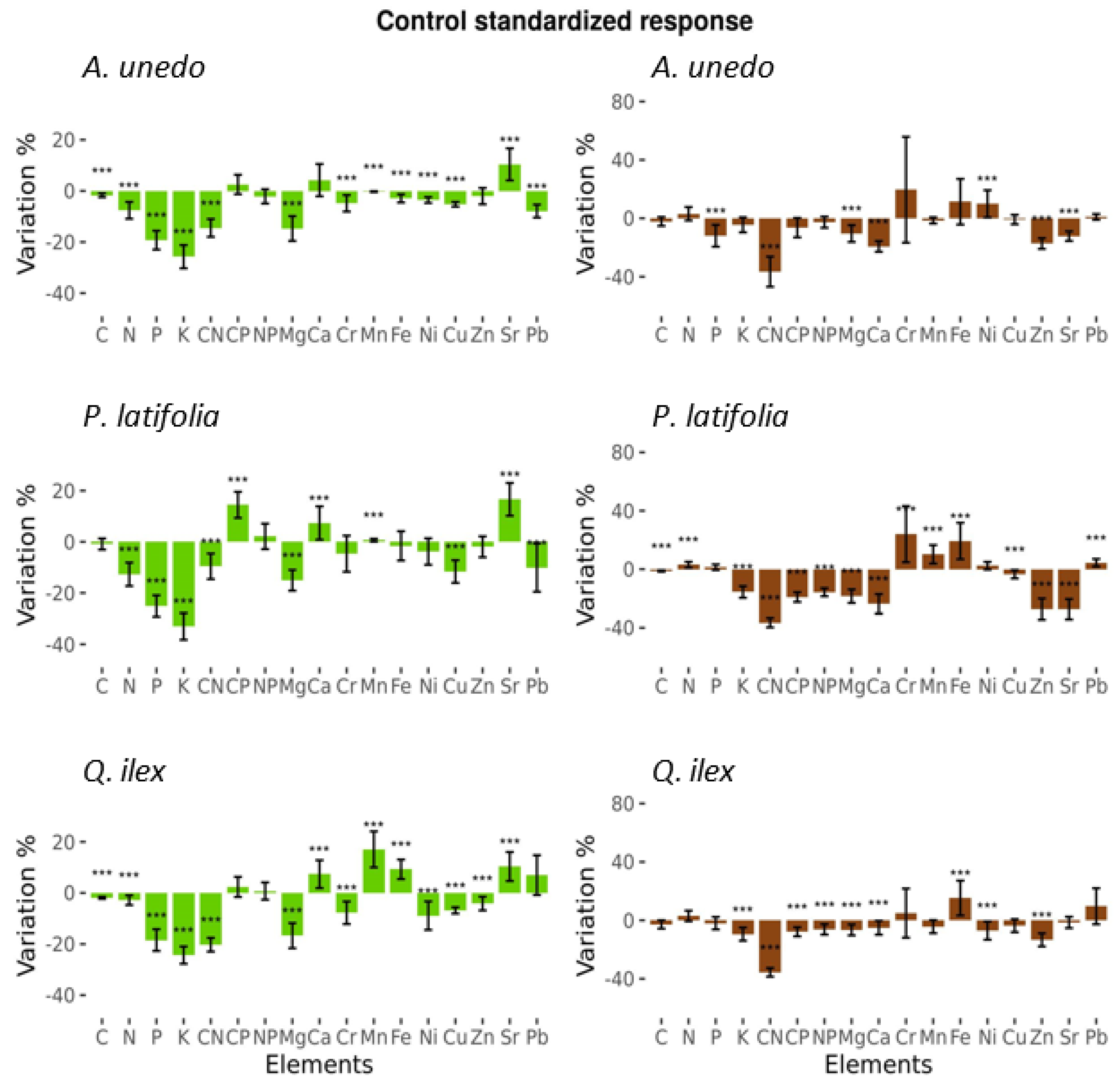
3.1. Fertilization Effects on Foliar Element Concentrations
3.2. Fertilization Effects on Leaf Litter Element Concentrations
3.3. Foliar Nutrient Resorption
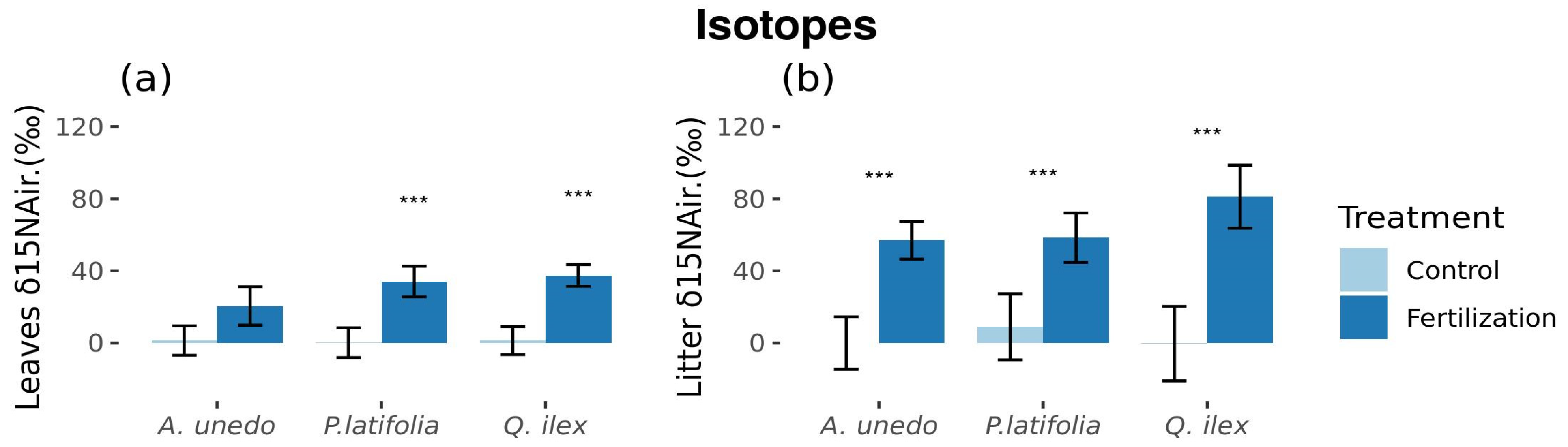
3.4. Fertilizer N-Uptake
3.5. Species Differences in Leaf and Leaf Litter Element Concentrations
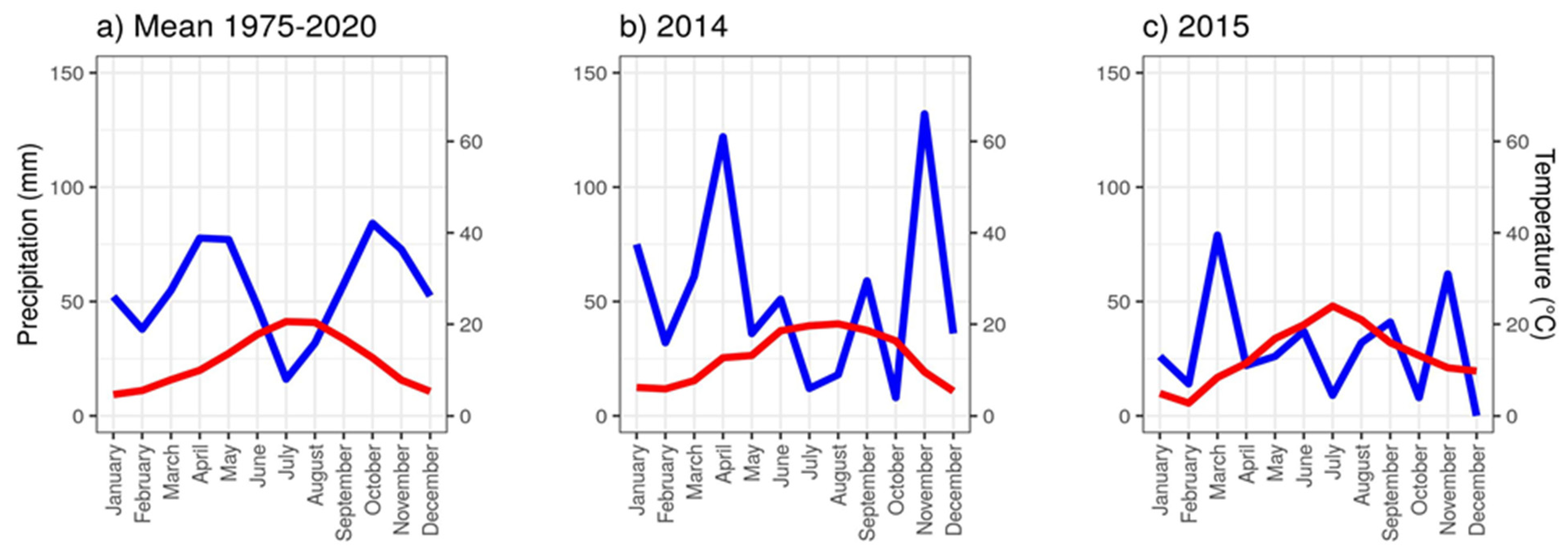
3.6. Drought Effects
4. Discussion
4.1. Interannual Differences
4.2. Macro-Elements
4.3. Micro-Elements
4.4. Nutrient Uptake
4.5. N limited Ecosystem
4.6. Biogeochemical Niche Differentiation
4.7. Species Differences in Response to N Fertilization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Peoples:, M.B.; Herridge, D.F.; Ladha, J.K. Biological nitrogen fixation: An efficient source of nitrogen for sustainable agricultural production? Plant Soil 1995, 174, 3–28. [Google Scholar] [CrossRef]
- Foth, H. Chapter 12: Plant-Soil Macronutrient Relations. In Fundamentals of Soil Science, 8th ed.; John Wiley Company: New York, NY, USA, 1990; pp. 186–209. [Google Scholar]
- Brady, N.; Weil, R. Nutrient cycles and soil fertility. In Elements of the Nature and Properties of Soils, 3rd ed.; Anthony, V.R., Ed.; Pearson Education Inc.: Upper Saddle River, NJ, USA, 2010; pp. 396–420. [Google Scholar]
- Galloway, J.N.; ATownsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the Nitrogen Cycle: Recent Trends, Questions, and Potential Solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; Van Der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hisinger, P.; Llusia, J.; et al. Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef] [PubMed]
- Penuelas, J.; Janssens, I.; Ciais, P.; Obersteiner, M.; Sardans, J. Anthropogenic global shifts in biospheric N and P concentrations and ratios and their impacts on biodiversity, ecosystem productivity, food security, and human health. Glob. Chang. Biol. 2020, 26, 1962–1985. [Google Scholar] [CrossRef] [PubMed]
- Engardt, M.; Simpson, D.; Schwikowski, M.; Granat, L. Deposition of sulphur and nitrogen in Europe 1900–2050. Model calculations and comparison to historical observations. Tellus B Chem. Phys. Meteorol. 2017, 69, 1328945. [Google Scholar] [CrossRef]
- Schmitz, A.; Sanders, T.G.M.; Bolte, A.; Bussotti, F.; Dirnböck, T.; Johnson, J.; Peñuelas, J.; Pollastrini, M.; Prescher, A.-K.; Sardans, J.; et al. Responses of forest ecosystems in Europe to decreasing nitrogen deposition. Environ. Pollut. 2019, 244, 980–994. [Google Scholar] [CrossRef] [PubMed]
- Slootweg, J.; Posch, M.; Hettelingh, J.P. Modelling and Mapping the Impacts of Atmospheric Deposition of Nitrogen and Sulphur: CCE Status Report 2015; Coordination Centre for Effects, National Institute for Public Health and the Environment: Bilthoven, The Netherlands, 2015. [Google Scholar]
- Pardo, L.H.; Fenn, M.E.; Goodale, C.L.; Geiser, L.H.; Driscoll, C.T.; Allen, E.B.; Baron, J.S.; Bobbink, R.; Bowman, W.D.; Clark, C.M.; et al. Effects of nitrogen deposition and empirical nitrogen critical loads for ecoregions of the United States. Ecol. Appl. 2011, 21, 3049–3082. [Google Scholar] [CrossRef]
- Fenn, M.E.; Allen, E.B.; Weiss, S.B.; Jovan, S.; Geiser, L.H.; Tonnesen, G.S.; Johnson, R.F.; Rao, L.E.; Gimeno, B.S.; Yuan, F.; et al. Nitrogen critical loads and management alternatives for N-impacted ecosystems in California. J. Environ. Manag. 2010, 91, 2404–2423. [Google Scholar] [CrossRef]
- Rao, L.E.; Allen, E.B. Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts. Oecologia 2010, 162, 1035–1046. [Google Scholar] [CrossRef]
- Rao, L.E.; Allen, E.B.; Meixner, T. Risk-based determination of critical nitrogen deposition loads for fire spread in southern California deserts. Ecol. Appl. 2010, 20, 1320–1335. [Google Scholar] [CrossRef]
- Gilliam, F.S. Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition. J. Ecol. 2006, 94, 1176–1191. [Google Scholar] [CrossRef]
- Friedrich, U.; Von Oheimb, G.; Kriebitzsch, W.U.; Schleßelmann, K.; Weber, M.S.; Härdtle, W. Nitrogen deposition increases susceptibility to drought—Experimental evidence with the perennial grass Molinia caerulea (L.) Moench. Plant Soil 2012, 353, 59–71. [Google Scholar] [CrossRef]
- Ochoa-Hueso, R.; Allen, E.B.; Branquinho, C.; Cruz, C.; Dias, T.; Fenn, M.E.; Manrique, E.; Pérez-Corona, M.E.; Sheppard, L.J.; Stock, W.D. Nitrogen deposition effects on Mediterranean-type ecosystems: An ecological assessment. Environ. Pollut. 2011, 159, 2265–2279. [Google Scholar] [CrossRef] [PubMed]
- Eisenlord, S.; Zak, D. Simulated Atmospheric Nitrogen Deposition Alters Actinobacterial Community Composition in Forest Soils. Soil Biol. Biochem. 2009, 74, 1157–1166. [Google Scholar] [CrossRef]
- Bontemps, J.D.; Hervé, J.C.; Leban, J.M.; Dhôte, J.F. Nitrogen footprint in a long-term observation of forest growth over the twentieth century. Trees 2011, 25, 237–251. [Google Scholar] [CrossRef]
- Schulte-Uebbing, L.; De Vries, W. Global-scale impacts of nitrogen deposition on tree carbon sequestration in tropical, temperate, and boreal forests: A meta-analysis. Glob. Chang. Biol. 2018, 24, e416–e431. [Google Scholar] [CrossRef]
- Fenn, M.E.; Poth, M.A.; Aber, J.D.; Baron, J.S.; Bormann, B.T.; Johnson, D.W.; Lemly, A.D.; McNulty, S.G.; Ryan, D.F.; Stottlemyer, R. Nitrogen Excess in North American Ecosystems: Predisposing Factors, Ecosystem Responses, and Management Strategies. Ecol. Appl. 1998, 8, 706–733. [Google Scholar] [CrossRef]
- Baron, J.; Rueth, H.; Wolfe, A.; Nydick, K.R.; Allstott, E.J.; Minear, J.T.; Moraska, B. Ecosystem Responses to Nitrogen Deposition in the Colorado Front Range. Ecosystems 2000, 3, 352–368. [Google Scholar] [CrossRef]
- Duquesnay, A.; Dupouey, J.L.; Clement, A.; Ulrich, E.; Le Tacon, F. Spatial and temporal variability of foliar mineral concentration in beech (Fagus sylvatica) stands in northeastern France. Tree Physiol. 2000, 20, 13–22. [Google Scholar] [CrossRef]
- Kowalenko, C.G. The effect of nitrogen and boron fertilizer applications on Willamette red raspberry growth, and on applied and other nutrients in the plant and soil over two growing seasons. Can. J. Plant Sci. 2005, 86, 213–225. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, B.; Davis, M.; Sardans, J.; Peñuelas, J.; Billings, S. Long-term nitrogen deposition linked to reduced water use efficiency in forests with low phosphorus availability. New Phytol. 2016, 210, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, M.; Marchetto, A.; Arisci, S.; Bussotti, F.; Calderisi, M.; Carnicelli, S.; Cecchini, G.; Fabbio, G.; Bertini, G.; Matteucci, G.; et al. On the tracks of Nitrogen deposition effects on temperate forests at their southern European range—An observational study from Italy. Glob. Chang. Biol. 2014, 20, 3423–3438. [Google Scholar] [CrossRef]
- Veresoglou, S.D.; Peñuelas, J.; Fischer, R.; Rautio, P.; Sardans, J.; Merilä, P.; Tabakovic-Tosic, M.; Rillig, M.C. Exploring continental-scale stand health e N: P ratio relationships for European forests. New Phytol. 2014, 202, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, Q.; Han, X. Soil Bacterial Communities Respond to Mowing and Nutrient Addition in a Steppe Ecosystem. PLoS ONE 2013, 8, e84210. [Google Scholar] [CrossRef]
- Sardans, J.; Penuelas, J. Plant-soil interactions in Mediterranean forest and shrublands: Impacts of climatic change. Plant Soil 2013, 365, 1–33. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Filella, I.; Estiarte, M.; Llusià, J.; Ogaya, R.; Carnicer, J.; Bartrons, M.; Rivas-Ubach, A.; Grau, O.; et al. Assessment of the impacts of climate change on Mediterranean terrestrial ecosystems based on data from field experiments and long-term monitored field gradients in Catalonia. Environ. Exp. Bot. 2018, 152, 49–59. [Google Scholar] [CrossRef]
- Carreira, J.A.; Sánchez-Vázquez, F.; Niell, F.X. Short-termand small-scale patterns of post-fire regeneration in a semi-ariddolomitic basin of Southern Spain. Acta Oecol. 1992, 13, 241–253. [Google Scholar]
- Henkin, Z.; Seligman, N.G.; Kafkafi, U.; Noy-Meir, I. Effective growing days: A simple predictive model of re-sponse of herbaceous plant growth in a Mediterranean ecosystem to variation in rainfall and phosphorus availability. J. Ecol. 1998, 86, 137–148. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Tree stoichiometry and growth. Glob. Ecol. Biogeogr. 2013, 22, 494–507. [Google Scholar] [CrossRef]
- Peñuelas, J.; Fernández-Martínez, M.; Ciais, P.; Jou, D.; Piao, S.; Obersteiner, M.; Vicca, S.; Janssens, I.A.; Sardans, J. The bioelements, the elementome, and the biogeochemical niche. Ecology 2019, 100, e02652. [Google Scholar] [CrossRef]
- Hu, M.; Peñuelas, J.; Sardans, J.; Sun, Z.; Wilson, B.J.; Huang, J.; Zhu, Q.; Tong, C. Stoichiometry patterns of plant organ N and P in coastal herbaceous wetlands along the East China Sea: Implications for biogeochemical niche. Plant Soil 2018, 431, 273–288. [Google Scholar] [CrossRef]
- Bobbink, B.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.M.; Morefield, P.E.; Gilliam, F.S.; Pardo, L.H. Estimated losses of plant biodiversity in the United States from historical N deposition (1985–2010). Ecology 2013, 94, 1441–1448. [Google Scholar] [CrossRef] [PubMed]
- Soons, M.B.; Hefting, M.M.; Dorland, E.; Lamers, L.P.M.; Versteeg, C.; Bobbink, R. Nitrogen effects on plant species richness in herbaceous communities are more widespread and stronger than those of phosphorus. Biol. Conserv. 2017, 212, 390–397. [Google Scholar] [CrossRef]
- Escarre, J.; Lepart, J.; Sans, X.; Sentuc, J.; Gorse, V. Effects of herbivory on the growth and reproduction of Picris hieracioides in the Mediterranean region. J. Veg. Sci. 1999, 10, 101–110. [Google Scholar] [CrossRef]
- Ogaya, R.; Peñuelas, J. Tree growth, mortality, and above-ground biomass accumulation in a holm oak forest under a five-year experimental field drought. Plant Ecol. 2007, 189, 291–299. [Google Scholar] [CrossRef]
- Tretiach, M. Photosynthesis and transpiration of evergreen Mediterranean and deciduous trees in an ecotone during a growing season. Acta Oecol. Int. J. Ecol. 1993, 14, 341–360. [Google Scholar]
- Lloret, F.; Siscart, D. Demographic drought effects on holm oak populations. Cuad. Soc. Esp. Cien. Forest. 1995, 2, 77–81. [Google Scholar]
- Panaiotis, C.; Loiser, R.; Paradis, G. Dating natural gaps in the holm oak forest (Quercus ilex L.) in Fango MAB Reserve (Corsica) by reading rings of maquis components datation des trouées naturelles en forêt de chênes verts (Quercus ilex L.) dans la réserve MAB du Fango (Corse) par la lecture des cernes des essences du maquis. Ann. Sci. For. 1995, 52, 477–487. [Google Scholar] [CrossRef]
- Ogaya, R.; Liu, D.; Barbeta, A.; Peñuelas, J. Stem Mortality and Forest Dieback in a 20-Years Experimental Drought in a Mediterranean Holm Oak Forest. Front. For. Glob. Chang. 2020, 2, 89. [Google Scholar] [CrossRef]
- Ogaya, R.; Peñuelas, J.; Martínez-Vilalta, J.; Mangirón, M. Effect of drought on diameter increment of Quercus ilex, Phillyrea latifolia, and Arbutus unedo in a holm oak forest of NE Spain. For. Ecol. Manag. 2003, 180, 175–184. [Google Scholar] [CrossRef]
- Ogaya, R.; Penuelas, J. Contrasting foliar responses to drought in Quercus ilex and Phillyrea latifolia. Biol. Plant. 2006, 50, 373–382. [Google Scholar] [CrossRef]
- Porter, J. Evaluation of washing procedures for pollution analysis of Ailanthus altissima leaves. Environ. Pollut. Ser. B Chem. Phys. 1986, 12, 195–202. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Bartoń, K. MuMIn: Multi-Model Inference. R Package Version 1.43.17. 2020. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 6 March 2021).
- Sardans, J.; Peñuelas, J.; Estiarte, M.; Prieto, P. Warming and drought alter C and N concentration, allocation and accumulation in a Mediterranean shrubland. Glob. Chang. Biol. 2008, 14, 2304–2316. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Ogaya, R. Drought-induced changes in C and N stoichiometry in a Quercus ilex Mediterranean forest. For. Sci. 2008, 54, 513–522. [Google Scholar]
- Sardans, J.; Peñuelas, J. Increasing drought decreases phosphorus availability in an evergreen Mediterranean forest. Plant Soil 2004, 267, 367–377. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Drought changes phosphorus and potassium accumulation patterns in an evergreen Mediterranean forest. Funct. Ecol. 2007, 21, 191–201. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Prieto, P.; Estiarte, M. Drought and warming induced changes in P and K concentration and accumulation in plant biomass and soil in a Mediterranean shrubland. Plant Soil. 2008, 306, 261–271. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Ogaya, R. Drough’s impact on Ca, Fe, Mg, Mo and S concentration and accumulation patterns in the plants and soil of a Mediterranean evergreen Quercus ilex forest. Biogeochemistry 2008, 87, 49–69. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Prieto, P.; Estiarte, M. Changes in Ca, Fe, Mg, Mo, Na, and S content under warming and drought. J. Geophys. Res. 2008, 113, G03039. [Google Scholar] [CrossRef]
- Plett, D.C.; Ranathunge, K.; Melino, V.J.; Kuya, N.; Uga, Y.; Kronzucker, H.J. The intersection of nitrogen nutrition and water use in plants: New paths toward improved crop productivity. J. Exp. Bot. 2020, 71, 4452–4468. [Google Scholar] [CrossRef]
- Barak, P.; Jobe, B.O.; Krueger, A.R.; Peterson, L.A.; Laird, D.A. Effects of long-term soil acidification due to nitrogen fertilizer inputs in Wisconsin. Plant Soil 1997, 197, 61–69. [Google Scholar] [CrossRef]
- Lu, X.K.; Mao, Q.G.; Gilliam, F.S.; Luo, Y.Q.; Mo, J.M. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Glob. Chang. Biol. 2014, 20, 3790–3801. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.K.; Vitousek, P.M.; Mao, Q.G.; Gilliam, F.S.; Luo, Y.Q.; Zhou, G.Y.; Zou, X.; Bai, E.; Scanlon, T.M.; Hou, E.; et al. Plant acclimation to long-term high nitrogen deposition in an N-rich tropical forest. Proc. Natl. Acad. Sci. USA 2018, 115, 5187–5192. [Google Scholar] [CrossRef] [PubMed]
- Fairfax, J.; Lepp, N. Effect of simulated ‘acid rain’ on cation loss from leaves. Nature 1975, 255, 324–325. [Google Scholar] [CrossRef]
- Sardans, J.; Urbina, I.; Grau, O.; Asensio, L.; Ogaya, R.; Peñuelas, J. Long-term drought decreases ecosystem C and nutrient storage in a Mediterranean holm oak forest. Environ. Exp. Bot. 2020, 177, 104135. [Google Scholar] [CrossRef]
- Peñuelas, J.; Matamala, R. Changes in N and S Leaf Content, Stomatal Density and Specific Leaf Area of 14 Plant Species during the Last Three Centuries of CO2 Increase. J. Exp. Bot. 1990, 41, 1119–1124. [Google Scholar] [CrossRef]
- Mayor, X.; Rodà, F. Effects of irrigation and fertilization on stem diameter growth in a Mediterranean holm oak forest. For. Ecol. Manag. 1994, 68, 119–126. [Google Scholar] [CrossRef]
- Ogaya, R.; Peñuelas, J. Climate Change Effects in a Mediterranean Forest Following 21 Consecutive Years of Experimental Drought. Forests 2021, 12, 306. [Google Scholar] [CrossRef]
- Sardans, J.; Janssens, I.A.; Alonso, R.; Veresoglou, S.D.; Rillig, M.C.; Sanders, T.G.; Carnicer, J.; Filella, I.; Farré-Armengol, G.; Peñuelas, J. Biogeochemical niche of European forest trees. Glob. Ecol. Biogeogr. 2015, 24, 240–255. [Google Scholar] [CrossRef]
- Sardans, J.; Vallicrosa, H.; Zuccarini, P.; Farré-Armengol, G.; Fernández-Martínez, M.; Peguero, G.; Gargallo-Garriga, A.; Ciais, P.; Janssens, I.A.; Obersteiner, M.; et al. Empirical support for the biogeochemical niche hypothesis in forest trees. Nat. Ecol. Evol. 2021, 5, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Capitanio, R.; Carcaillet, C. Post-fire Mediterranean vegetation dynamics and diversity: A discussion of succession models. For. Ecol. Manag. 2008, 255, 431–439. [Google Scholar] [CrossRef]
- Gratani, L.; Catoni, R.; Varone, L. Morphological, anatomical and physiological leaf traits of Q. ilex, P. latifolia, P. lentiscus, and M. communis and their response to Mediterranean climate stress factors. Bot. Stud. 2013, 54, 35. [Google Scholar] [CrossRef] [PubMed]
- Barbeta, A.; Mejía-Chang, M.; Ogaya, R.; Voltas, J.; Dawson, T.E.; Peñuelas, J. The combined effects of a long-term experimental drought and an extreme drought on the use of plant-water sources in a Mediterranean forest. Glob. Chang. Biol. 2015, 21, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Vilalta, J.; Prat, E.; Oliveras, I.; Piñol, J. Hydraulic properties of roots and stems of nine woody species from a Holm oak forest in NE Spain. Oecologia 2002, 133, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, L.; Mereu, S.; Brunetti, C.; Ferdinando, M.D.; Ferrini, F.; Manes, F.; Salvatori, E.; Marzuoli, R.; Gerosa, G.; Tattini, M. Photosynthetic performance and biochemical adjustments in two co-occurring Mediterranean evergreens, Quercus ilex and Arbutus unedo, differing in salt-exclusion ability. Funct. Plant Biol. 2014, 41, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Burucs, Z.; Schmidhalter, U. Effects of foliar fertilization application on the growth and mineral nutrient content of maize seedlings under drought and salinity. Soil Sci. Plant Nutr. 2008, 54, 133–141. [Google Scholar] [CrossRef]
- Tramblay, Y.; Koutroulis, A.; Samaniego, L.; Vicente-Serrano, S.M.; Volaire, F.; Boone, A.; Le Page, M.; Llasat, M.C.; Albergel, C.; Burak, S.; et al. Challenges for drought assessment in the Mediterranean region under future climate scenarios. Earth-Sci. Rev. 2020, 210, 103348. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) Leaves | |||
| Test | Wilks Lambda | F | p-Value |
| 13C | 0.953 | 2.67 | 0.023 |
| 15N | 0.748 | 18.1 | <0.0001 |
| N | 0.989 | 0.615 | 0.69 |
| P | 0.962 | 2.13 | 0.062 |
| K | 0.959 | 2.24 | 0.049 |
| Mg | 0.962 | 2.14 | 0.061 |
| Ca | 0.938 | 3.57 | 0.0038 |
| Mn | 0.742 | 18.7 | <0.0001 |
| Fe | 0.906 | 5.6 | <0.0001 |
| Cu | 0.946 | 3.1 | 0.0098 |
| Zn | 0.941 | 3.36 | 0.0058 |
| season | 0.941 | 1.66 | 0.087 |
| (b) Litter | |||
| Variables | Wilks Lambda | F | p-Value |
| 13C | 0.917 | 4.36 | 0.00081 |
| 15N | 0.655 | 25.3 | <0.00001 |
| N | 0.948 | 2.65 | 0.023 |
| P | 0.827 | 10.1 | <0.00001 |
| K | 0.869 | 7.26 | <0.00001 |
| Mg | 0.959 | 2.07 | 0.07 |
| Ca | 0.85 | 8.51 | <0.00001 |
| Mn | 0.842 | 9.04 | <0.00001 |
| Fe | 0.947 | 2.68 | 0.022 |
| Cu | 0.955 | 2.28 | 0.048 |
| Zn | 0.932 | 3.52 | 0.0043 |
| season | 0.9 | 2.6 | 0.0045 |
| (a) Leaves | |||||
| PC | AC | AN | QN | PN | |
| QC | M = 3.83 | M = 3.08 | M = 4.40 | M = 2.96 | M = 4.21 |
| F = 6.77 | F = 5.44 | F = 7.69 | F = 5.24 | F = 7.44 | |
| p < 0.0001 | p < 0.0001 | p < 0.0001 | p < 0.0001 | p < 0.0001 | |
| PC | M = 1.03 | M = 1.37 | M = 6.27 | M = 1.76 | |
| F = 1.82 | F = 2.39 | F = 11.09 | F = 3.10 | ||
| p = 0.041 | p = 0.0046 | p < 0.0001 | p = 0.0002 | ||
| AC | M = 1.29 | M = 6.28 | M = 2.31 | ||
| F = 2.26 | F = 11.1 | F = 4.08 | |||
| p = 0.0077 | p < 0.0001 | p < 0.0001 | |||
| AN | M = 5.28 | M = 1.75 | |||
| F = 0.23 | F = 3.07 | ||||
| p < 0.0001 | p = 0.0003 | ||||
| QN | M = 3.64 | ||||
| F = 6.43 | |||||
| p < 0.0001 | |||||
| (b) Litter | |||||
| AC | PC | AN | QN | PN | |
| QC | M = 2.62 | M = 3.41 | M = 4.09 | M = 5.9 | M = 4.03 |
| F = 4.61 | F = 5.21 | F = 7.12 | F = 10.2 | F = 5.77 | |
| p < 0.000001 | p < 0.000001 | p < 0.000001 | p < 0.000001 | p < 0.000001 | |
| AC | M = 3.63 | M = 3.57 | M = 6.50 | M = 7.25 | |
| F = 5.56 | F = 6.21 | F = 11.2 | F = 10.38 | ||
| p < 0.000001 | p < 0.000001 | p < 0.000001 | p < 0.000001 | ||
| PC | M = 5.23 | M = 7.29 | M = 5.39 | ||
| F = 7.93 | F = 10.9 | F = 6.89 | |||
| p < 0.000001 | p < 0.000001 | p < 0.000001 | |||
| AN | M = 3.19 | M = 3.24 | |||
| F = 5.44104 | F = 4.60059 | ||||
| p < 0.000001 | p < 0.000001 | ||||
| QN | M = 2.551809 | ||||
| F = 3.60 | |||||
| p < 0.000001 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vallicrosa, H.; Sardans, J.; Ogaya, R.; Fernández, P.R.; Peñuelas, J. Short-Term N-Fertilization Differently Affects the Leaf and Leaf Litter Chemistry of the Dominant Species in a Mediterranean Forest under Drought Conditions. Forests 2021, 12, 605. https://doi.org/10.3390/f12050605
Vallicrosa H, Sardans J, Ogaya R, Fernández PR, Peñuelas J. Short-Term N-Fertilization Differently Affects the Leaf and Leaf Litter Chemistry of the Dominant Species in a Mediterranean Forest under Drought Conditions. Forests. 2021; 12(5):605. https://doi.org/10.3390/f12050605
Chicago/Turabian StyleVallicrosa, Helena, Jordi Sardans, Romà Ogaya, Pere Roc Fernández, and Josep Peñuelas. 2021. "Short-Term N-Fertilization Differently Affects the Leaf and Leaf Litter Chemistry of the Dominant Species in a Mediterranean Forest under Drought Conditions" Forests 12, no. 5: 605. https://doi.org/10.3390/f12050605
APA StyleVallicrosa, H., Sardans, J., Ogaya, R., Fernández, P. R., & Peñuelas, J. (2021). Short-Term N-Fertilization Differently Affects the Leaf and Leaf Litter Chemistry of the Dominant Species in a Mediterranean Forest under Drought Conditions. Forests, 12(5), 605. https://doi.org/10.3390/f12050605