
Do Adjacent Forests Affect the Regeneration of Oak-Hornbeam and Ancient Forest Plant Species in Manor Parks in Poland?



Abstract
1. Introduction
2. Materials and Methods
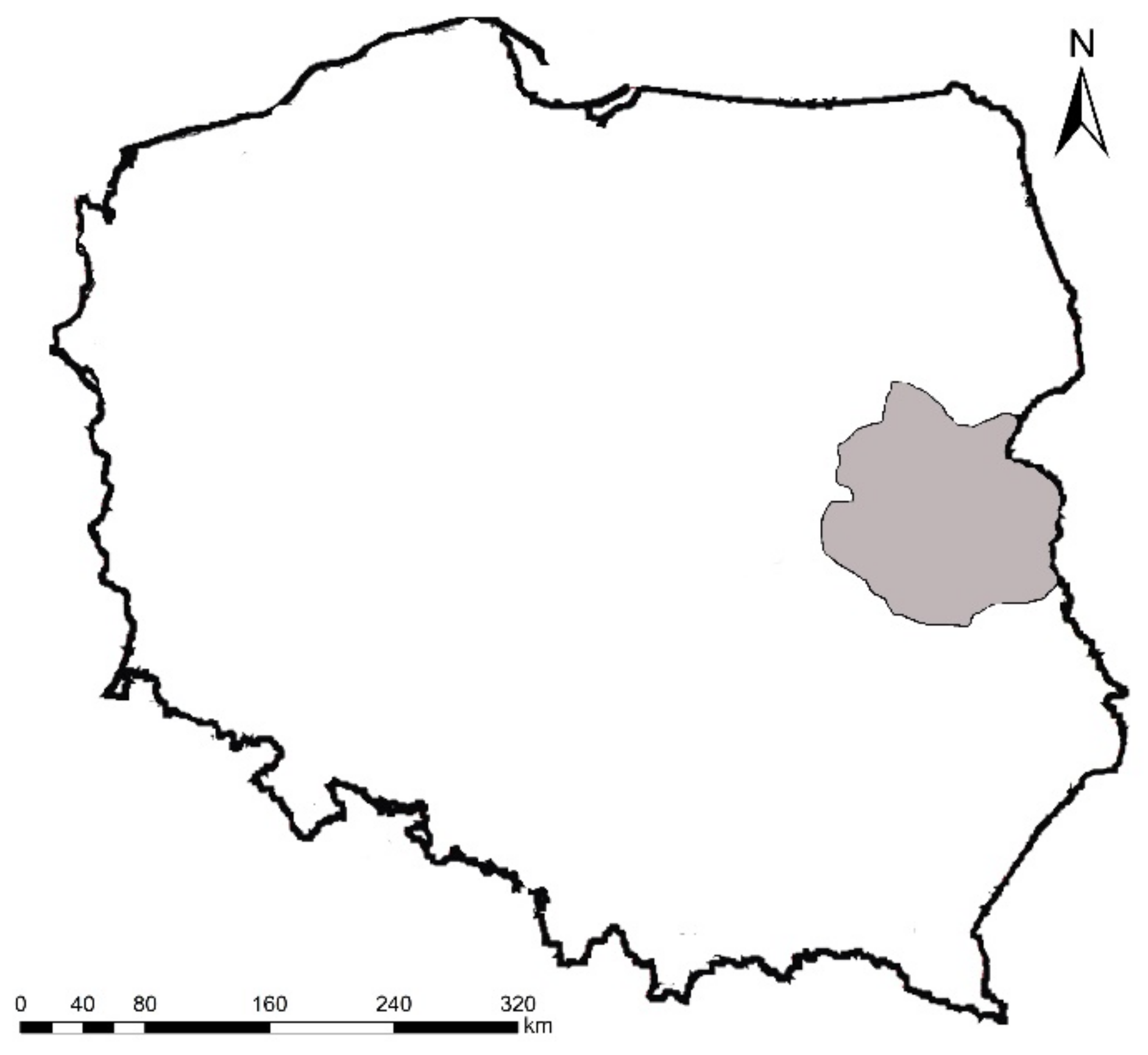
2.1. Study Areas and Field Methods
2.2. Statistical Methods
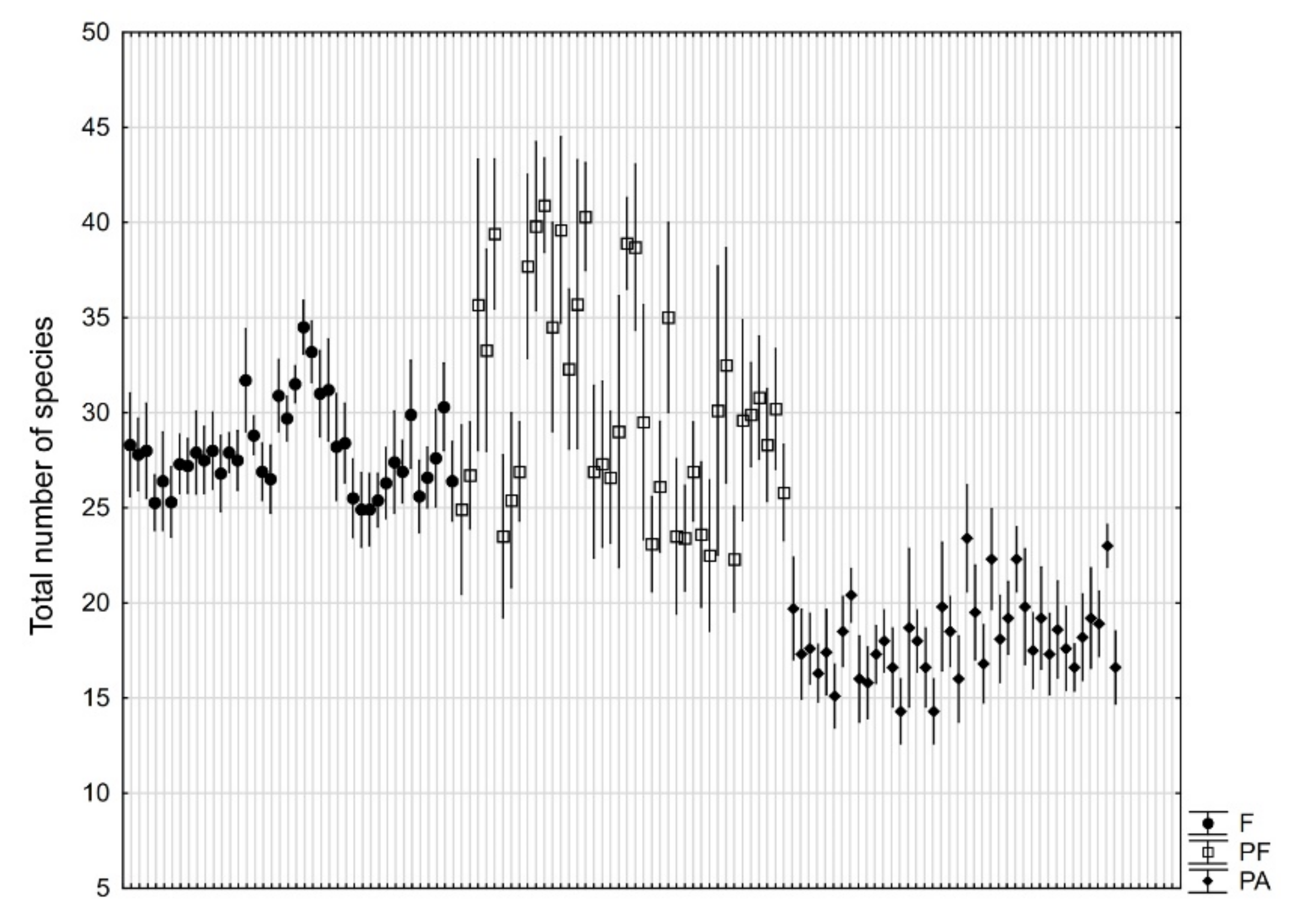
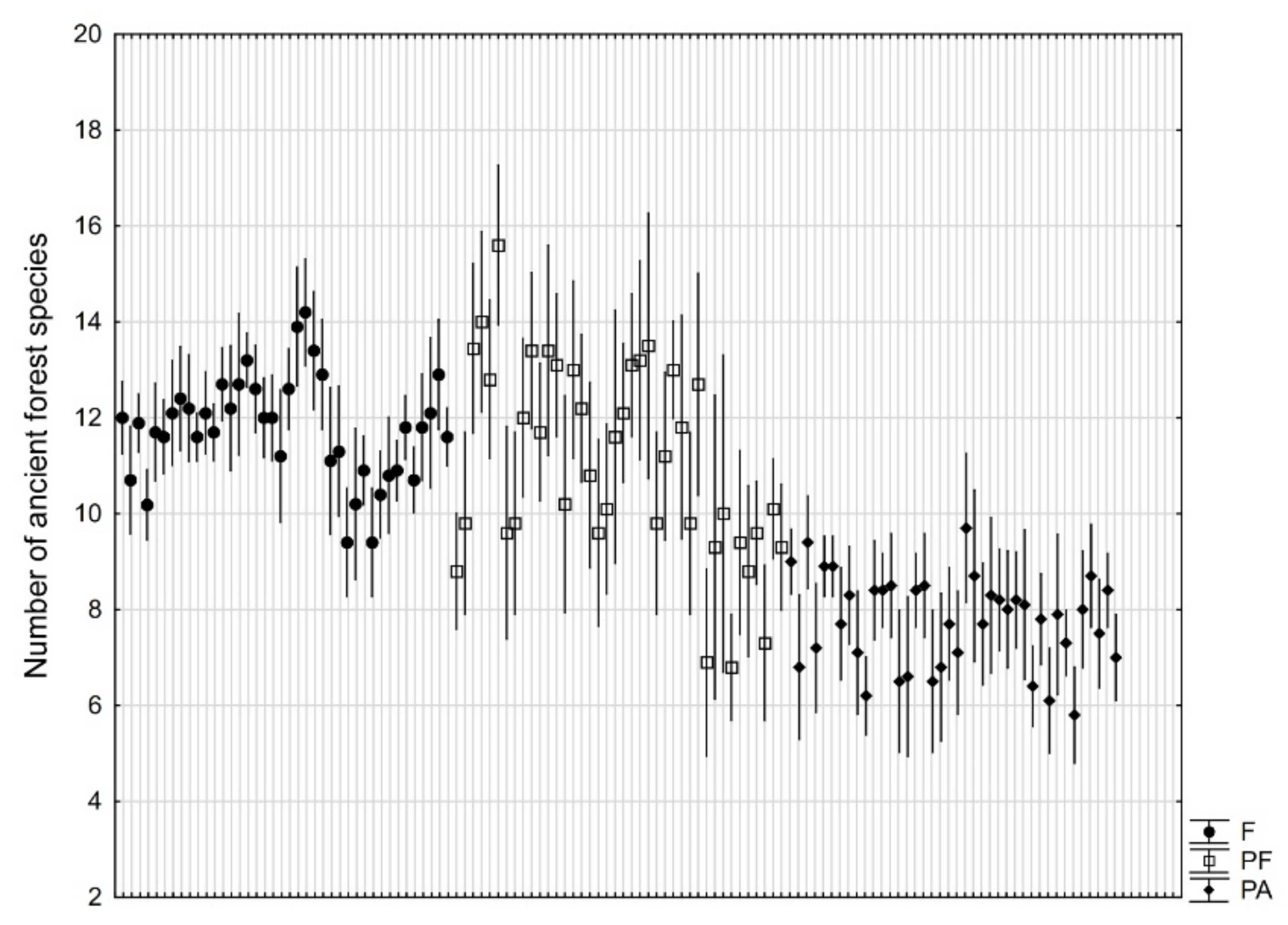
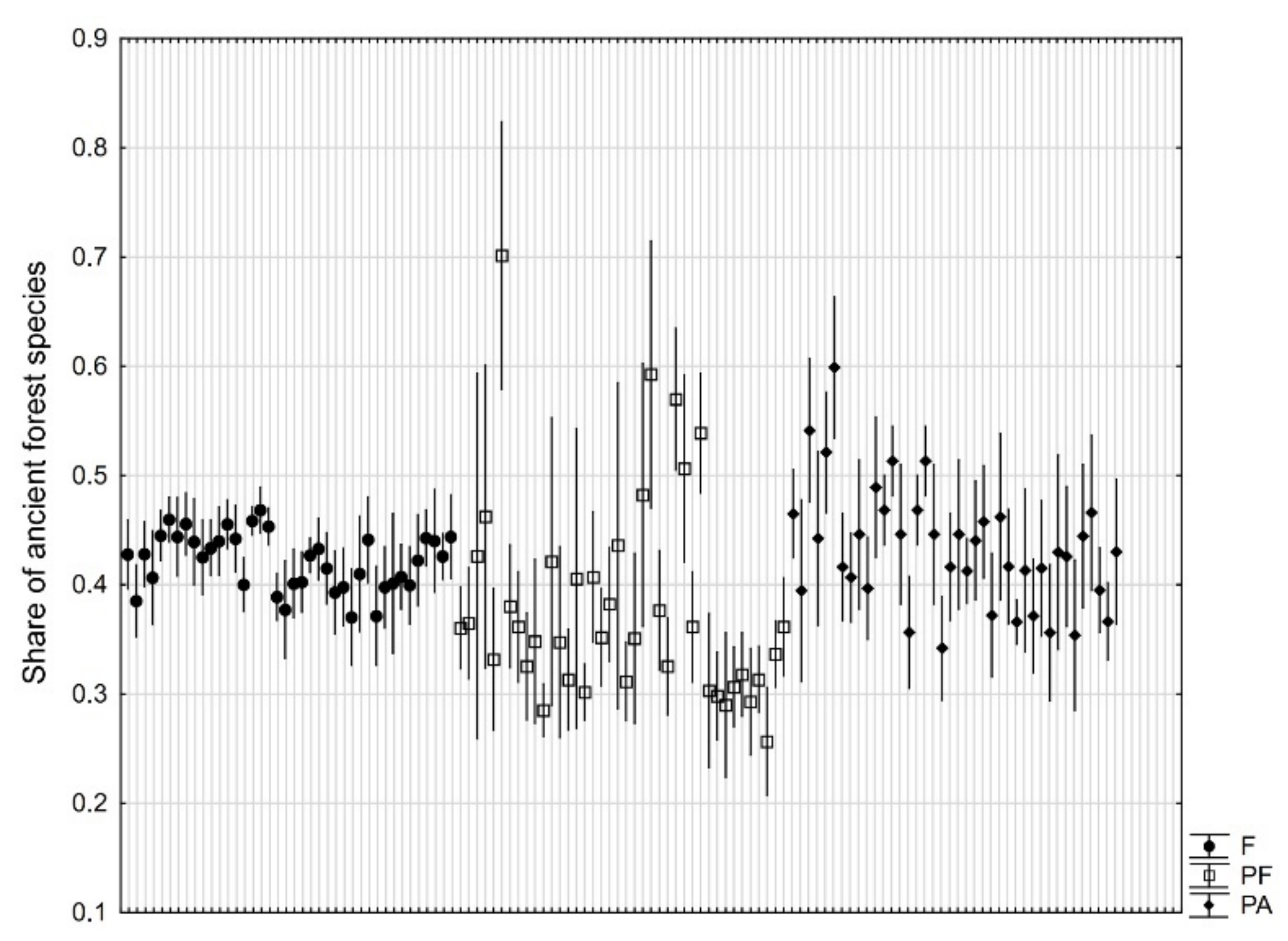
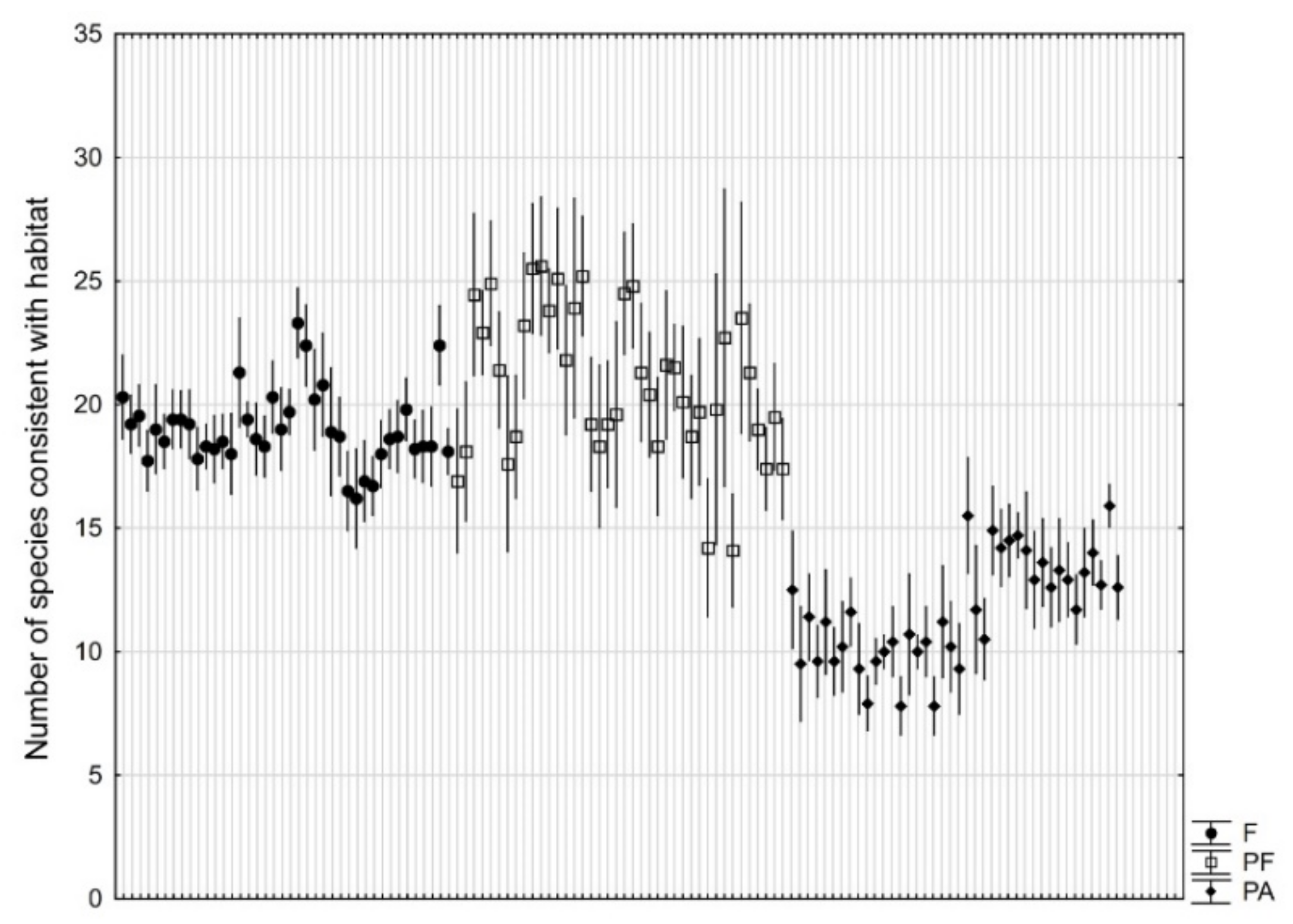
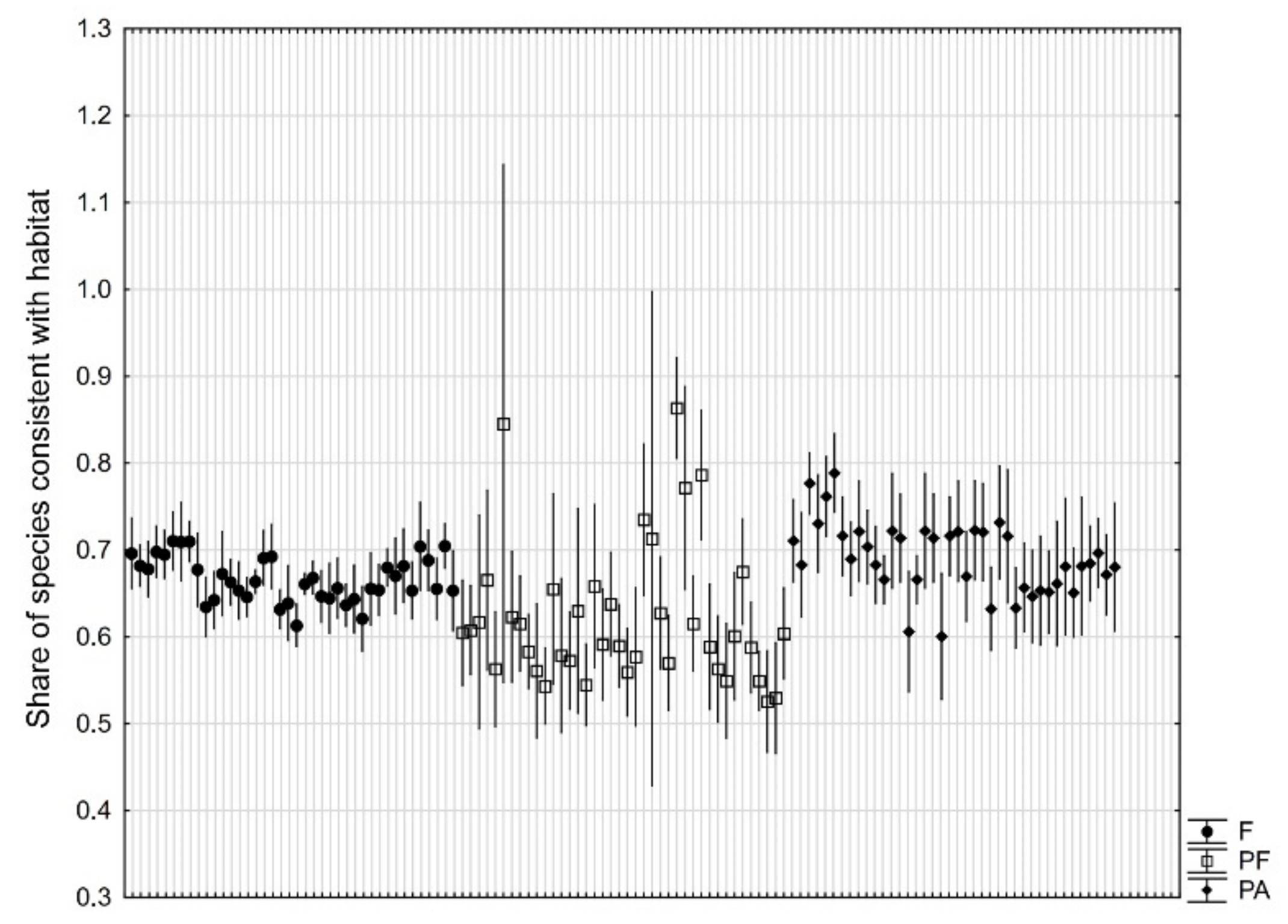
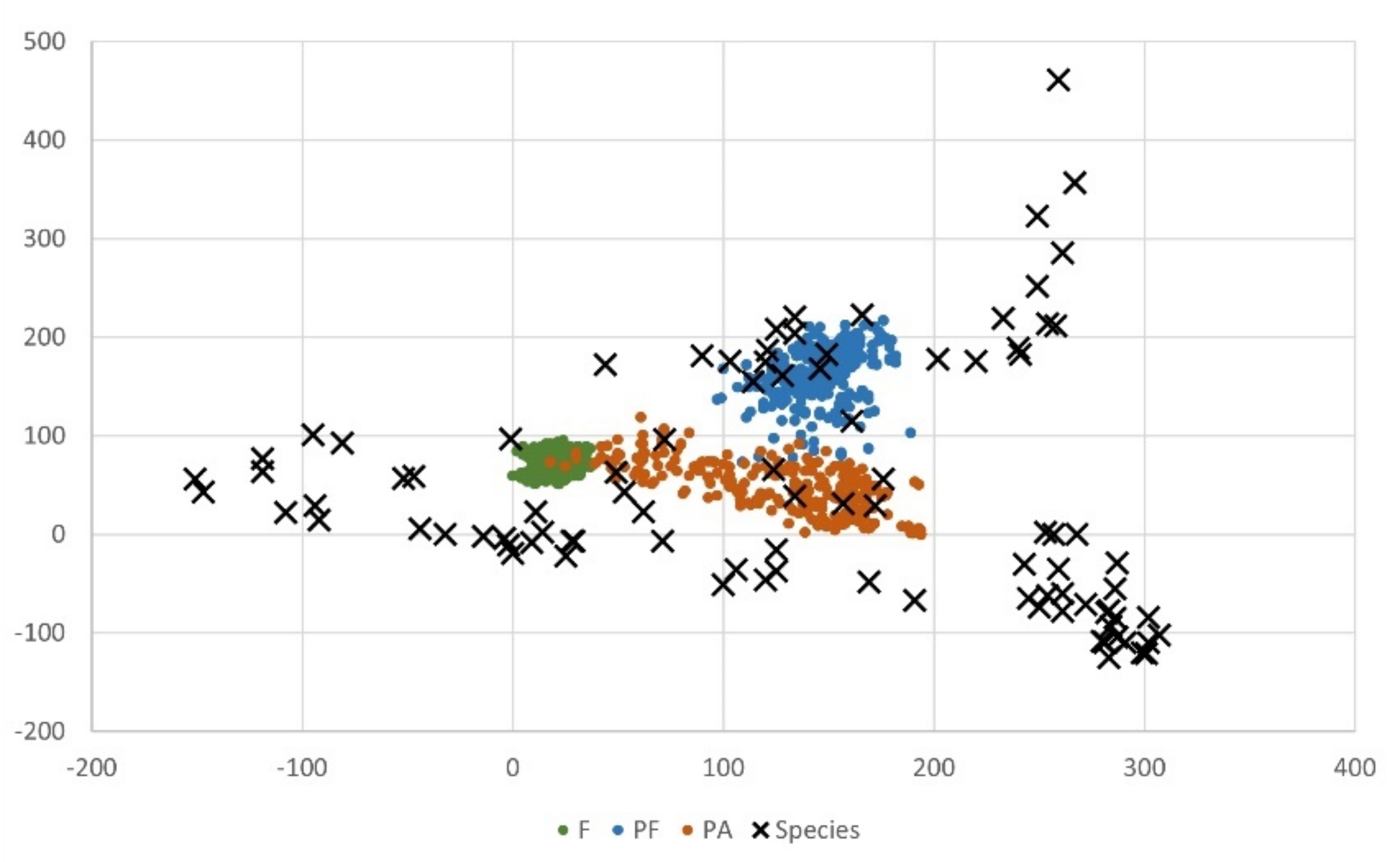
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latowski, K.; Zieliński, J. Parki wiejskie—wybrane zagadnienia geobotaniczne i kulturowe. In Szata Roślinna Wielkopolski i Pojezierza Południowopomorskiego, Przewodnik Sesji Terenowej 52. Zjazdu PTB, 24−28 Września 2001; Wojterska, M., Ed.; Bogucki Wyd. Naukowe: Poznań, Poland, 2001; pp. 291–304. [Google Scholar]
- Fabiańska, P. Rola parków podworskich w krajobrazie wsi. In Krajobraz i Ogród Wiejski, t. 1, Nowe Idee i Metody w Architekturze Krajobrazu; Janecki, J., Borkowski, Z., Eds.; Wydawnictwo KUL: Lublin, Poland, 2004; pp. 165–175. [Google Scholar]
- International Council on Monuments and Sites. Historic gardens (The Florence Charter 1981). Available online: https://www.icomos.org/charters/gardens_e.pdf (accessed on 17 January 2020).
- Brunet, J.; Valtinat, K.; Mayr, M.L.; Felton, A.; Lindbladh, M.; Bruun, H.H. Understory succession in post-agricultural oak forests: Habitat fragmentation affects forest specialists and generalists differently. For. Ecol. Manag. 2011, 262, 1863–1871. [Google Scholar] [CrossRef]
- Fijałkowski, D.; Kseniak, M. Parki Wiejskie Lubelszczyzny. Stan, Ochrona i Rewaloryzacja Biocenotyczna; PWNL: Warsaw, Poland, 1982. [Google Scholar]
- Vellend, M.; Baeten, L.; Myers-Smith, I.H.; Elmendorf, S.C.; Beauséjour, R.; Brown, C.D. Global meta-analysis reveals no net change in local-scale plant biodiversity over time. Proc. Natl. Acad. Sci. USA 2013, 110, 19456–19459. [Google Scholar] [CrossRef]
- Chmielewski, T.J. Pojezierze Łęczycko-Włodawskie: Przekształcenia Struktury Ekologicznej Krajobrazu i Uwarunkowania Zagospodarowania Przestrzennego; Monografie Komitetu Inżynierii Środowiska PAN: Lublin, Poland, 2001. [Google Scholar]
- Convention on Biological Diversity. Available online: https://www.cbd.int/doc/legal/cbd-en.pdf (accessed on 12 March 2021).
- Liira, J.; Lõhmus, K.; Tuisk, E. Old manor parks as potential habitats for forest flora in agricultural landscape of Estonia. Biol. Conserv. 2012, 146, 144–154. [Google Scholar] [CrossRef]
- Alberdi, I.S.; Condés, S.; Martínez-Millán, J. Review of monitoring and assessing ground vegetation biodiversity in national forest inventories. Environ. Monit. Assess. 2010, 164, 649–676. [Google Scholar] [CrossRef]
- Del Vecchio, S.; Slaviero, A.; Fantinato, E.; Buffa, G. The use of plant community attributes to detect habitat quality in coastal environments. AoB Plants 2016, 8, plw040. [Google Scholar] [CrossRef]
- Wulf, M. Plant species as indicators as ancient woodland in north-western Germany. J. Veg. Sci. 1997, 8, 635–642. [Google Scholar] [CrossRef]
- Wulf, M. Preference of plant species for woodlands with differing habitat continuities. Flora 2003, 198, 444–460. [Google Scholar] [CrossRef]
- Verheyen, K.; Bossuyt, B.; Honnay, O.; Hermy, M. Herbaceous plant community structure of ancient and recent forest in two contrasting forest types. Basic Appl. Ecol. 2003, 4, 537–546. [Google Scholar] [CrossRef]
- Kolb, A.; Diekmann, M. Effects of environment, habitat configuration, and forest continuity on the distribution of forest plant species. J. Veg. Sci. 2004, 15, 199–208. [Google Scholar] [CrossRef]
- Stefańska-Krzaczek, E.; Kacki, Z.; Szypuła, B. Coexistence of ancient forest species as an indicator of high species richness. For. Ecol. Manag. 2016, 365, 12–21. [Google Scholar] [CrossRef]
- Lalechère, E.; Jabot, F.; Archaux, F.; Deffuant, G. Non-equilibrium Plant metapopulation dynamics challenge the concept of ancient/recent forest species. Ecol. Model. 2017, 366, 48–57. [Google Scholar] [CrossRef]
- Webb, J.C.; Goodenough, A.E. Questioning the reliability of “ancient” woodland indicators: Resilience to interruptions and persistence following deforestation. Ecol. Indic. 2018, 84, 354–363. [Google Scholar] [CrossRef]
- Dzwonko, Z.; Gawroński, S. The role woodlands fragments, soil types, and dominant species in secondary succession on the western Carpatian foothills. Vegetatio 1994, 111, 149–160. [Google Scholar] [CrossRef]
- Bossuyt, B. Plant species and soil dynamics across ancient-recent forest ecotones: Consequences for ecological restoration. Ph.D. Thesis, KU Leuven, Leuven, Belgium, 2001. [Google Scholar]
- Mortelliti, A.; Amori, G.L.; Boitani, L. The role of habitat quality in fragmented landscapes: A conceptual overview and prospectus for future research. Oecologia 2010, 163, 535–1547. [Google Scholar] [CrossRef] [PubMed]
- Aggemyr, E.; Cousins, S.A.O. Landscape structure and land use history influence changes in island plant composition after 100 years. J. Biogeogr. 2010, 39, 1645–1656. [Google Scholar] [CrossRef]
- Hill, J.L.; Curran, P.J. Species composition in fragmented forest: Conservation implications of changing forest area. Appl. Geogr. 2001, 21, 157–174. [Google Scholar] [CrossRef]
- Hill, J.L.; Curran, P.J. Area, shape, and isolation of tropical forest fragments: Effects on tree species diversity and implications for conservations. J. Biogeogr. 2003, 30, 1391–1403. [Google Scholar] [CrossRef]
- Closset-Kopp, D.; Wasof, S.; Decocq, G. Using process-based indicator species to evaluate ecological corridors in fragmented landscape. Biol. Conserv. 2016, 201, 152–159. [Google Scholar] [CrossRef]
- Matuszkiewicz, W. Przewodnik do Oznaczania Zbiorowisko Roślinnych Polski; Wydawnictwo Naukowe PWN: Warsaw, Poland, 2012. [Google Scholar]
- Żarska, B. Zmiany roślinności w strefie brzegowej lasów. Ochr. Śr. Zasobów Nat. 1997, 12, 105–115. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie, 2nd ed.; Springer: Wien, Austria, 1951. [Google Scholar]
- Mirek, Z.; Zając, M.; Zając, A.; Piękoś-Mirkowa, H. Vascular Plants of Poland: A Checklist; Instytut Botaniki im. Władysława Szafera Polskiej Akademii Nauk, Komitet Badań Naukowych: Cracow, Poland, 1995. [Google Scholar]
- Hermy, M.; Honnay, O.; Firbank, L.; Grashof-Bokdam, C.; Lawesson, J.E. An ecological comparison between ancient and other forest plant species of Europe and the implications for conservation. Biol. Conserv. 1999, 91, 9–22. [Google Scholar] [CrossRef]
- Dzwonko, Z.; Loster, S. Wskaźnikowe gatunki roślin starych lasów i ich znaczenie dla ochrony przyrody i kartografii roślinności. Typologia zbiorowisk i kartografia roślinności w Polsce. Pr. Geogr. 2001, 178, 119–132. [Google Scholar]
- Xue-Kun Song, P.; Tan, M. Marginal models for longitudinal continuous proportional data. Biometrics 2000, 56, 496–502. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Van der Maarel, E. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 1979, 39, 97–114. [Google Scholar] [CrossRef]
- Graae, B.J.; Oakland, R.H. Influence of historical, geographical and environmental variables on understory composition and richness in Danish forest. J. Veg. Sci. 2004, 15, 465–474. [Google Scholar] [CrossRef]
- Mac Arthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Naff, T.; Kolk, J. Colonization credit of post-agricultural forest patches in NE Germany. Biol. Conserv. 2015, 182, 155–163. [Google Scholar] [CrossRef]
- Fornal-Pieniak, B.; Ollik, M.; Schwerk, A. Impact of surroundings landscape structure on formation of plant species. Appl. Ecol. Environ. Res. 2018, 16, 6483–6497. [Google Scholar] [CrossRef]
- Damschen, E.I.; Baker, D.V.; Bohrer, G.; Nathan, R.; Orrock, J.L.; Turner, J.R.; Brudvig, L.A.; Haddad, N.M.; Levey, D.J.; Tewksbury, J.J. How fragmentation and corridors affect wind dynamics and seed dispersal in open habitats. Proc. Natl. Acad. Sci. USA 2014, 111, 3484–3489. [Google Scholar] [CrossRef] [PubMed]
- Król, S. Problemy synantropizacji lasów a penetracja antropofitów dendroflory. Sylwan 2002, 146, 75–89. [Google Scholar] [CrossRef]
- Eriksson, O. Regional dynamics of plants: A review of evidence of remnant, source-sink, and metapopulations. Oikos 1996, 77, 248–258. [Google Scholar] [CrossRef]
- Liira, J.; Sepp, T.; Parrest, O. The forest structure and ecosystem quality in conditions of anthropogenic disturbance along productivity gradient. For. Ecol. Manag. 2007, 250, 34–46. [Google Scholar] [CrossRef]
- Desender, K.; Ervynck, A.; Tack, G. Beetle diversity and historical ecology of woodlands in Flanders. Belg. J. Zool. 1999, 129, 139–155. [Google Scholar]
- Kalda, R.; Kalda, O.; Lõhmus, K.; Liira, J. Multi-scale ecology of woodland bat the role of species pool, landscape complexity and stand structure. Biodivers. Conserv. 2014, 24, 337–353. [Google Scholar] [CrossRef]
- De Keersmaeker, L.; Martens, L.; Verheyen, K.; Hermy, M.; de Schrijver, A.; Lust, N. Impact of soil fertility and isolation on diversity of herbaceosus woodland species colonizig afforestations in Muizen forest (Belgium). For. Ecol. Manag. 2004, 188, 291–297. [Google Scholar] [CrossRef]
- Matuszkiewicz, J.M.; Kowalska, A.; Solon, J.; Degórski, M.; Kozłowska, A.; Roo-Zielińska, E.; Zawiska, I.; Wolski, J. Long-term evolution models of post-agricultural forests. Pr. Geogr. 2013, 240, 1–318. [Google Scholar]
- Łaska, G. Ecological consequences of deforestation and afforestation on a post-arable land: Changes in the composition and structure of plant communities and transformations of oak-hornbeam habitats and soil. Ecol. Quest. 2014, 20, 9–21. [Google Scholar] [CrossRef]
- Higgs, E.; Falk, D.A.; Guerrini, A.; Hall, M.; Harris, J.; Hobbs, R.J.; Jackson, S.T.; Rhemtulla, J.M.; Throop, W. The changing role of history in restoration ecology. Front. Ecol. Environ. 2014, 12, 499–506. [Google Scholar] [CrossRef]
- Evers, C.R.; Wardropper, C.B.; Branoff, B.; Granek, E.F.; Hirsch, S.L.; Link, T.E.; Olivero-Lora, S.; Wilson, C. The ecosystem services and biodiversity of novel ecosystems: A literature review. Glob. Ecol. Conserv. 2017, 13, e00362. [Google Scholar] [CrossRef]
- Andersen, J.; Rowcliffe, J.M.; Cowlishaw, G. Does the matrix matter? A forest primate in a complex agricultural landscape. Biol. Conserv. 2007, 135, 212–222. [Google Scholar] [CrossRef]
- Jamoneau, A.; Chabrerie, O.; Closset-Kopp, D.; Decocq, G. Fragmentation alters beta-diversity patterns of habitat specialists within forest metacommunities. Ecography 2012, 35, 124–133. [Google Scholar] [CrossRef]
- Hermy, M.; Verheyen, K. Legacies of the past in the present-day forest biodiversity: A review of past land-use effects on forest plant species composition and diversity. Ecol. Res. 2007, 22, 361–371. [Google Scholar] [CrossRef]
- Closset-Kopp, D.; Hattab, T.; Decocq, G. Do drivers of forestry vehicles also drive herb layer changes (1970–2015) in a temperate forest with contrasting habitat and management conditions? J. Ecol. 2019, 107, 1439–1456. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Characteristic | Species Score for Axis 1 | Species Score for Axis 2 |
|---|---|---|---|
| Species characteristic for nature reserves (F) | |||
| Oxalis acetosella | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −151 | 56 |
| Melampyrum nemorosum | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −147 | 43 |
| Hedera helix | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −119 | 77 |
| Moehringia trinervia | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −119 | 64 |
| Asarum europaeum | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −108 | 22 |
| Melica nutans | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −95 | 101 |
| Dactylis polygama | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −94 | 29 |
| Anemone ranunculoides | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −92 | 15 |
| Ranunculus lanuginosus | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −81 | 93 |
| Geranium phaeum | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −52 | 57 |
| Viola mirabilis | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −47 | 59 |
| Paris quadrifolia | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −44 | 6 |
| Pulmonaria officinalis | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −32 | 0 |
| Milium effusum | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | −14 | −2 |
| Species characteristic for manor parks surrounded by fields (PA) | |||
| Veronica officinalis | Non-forest species, grass species (Nardo-Callunetea class) | 283 | −125 |
| Impatiens parviflora | Non-forest species, synanthropical species (Artemisietea vulgaris class) | 301 | −121 |
| Galium mollugo | Non-forest species, grass species (Trifolio-Geranietea sanguinei species) | 299 | −120 |
| Geum urbanum | Non-forest species, synanthropical species (Artemisietea vulgaris class) | 281 | −110 |
| Plantago major | Non-forest species, grass species (Molinio-Arrhenatheretea class) | 291 | −110 |
| Lamium maculatum | Non-forest species, synanthropical species (Artemisietea vulgaris class) | 302 | −110 |
| Chelidonium majus | Artemisietea vulgaris | 280 | −108 |
| Lysimachia nummularia | Non-forest species, grass species (Molinio-Arrhenatheretea class) | 287 | −104 |
| Convolvulus arvensis | Agropyretea | 307 | −102 |
| Chaerophyllum aromaticum | Non-forest species, synanthropical species (Artemisietea vulgaris class) | 284 | −93 |
| Dactylis glomerata | Non-forest species, grass species (Molinio-Arrhenatheretea class) | 286 | −85 |
| Rubus caesius | Non-forest species, shrub species (Rhamno-Prunetea class) | 302 | −84 |
| Rubus idaeus | Epilobietea angustifolii | 282 | −79 |
| Peucedanum oreoselinum | Non-forest species, grass species Trifolio-Geranietea sanguinei | 261 | −78 |
| Alliaria petiolata | Non-forest species, synanthropical species (Artemisietea vulgaris class) | 283 | −77 |
| Taraxacum officinale | Non-forest species, grass species (Molinio-Arrhenatheretea class) | 250 | −74 |
| Glechoma hederacea | Artemisietea vulgaris | 272 | −71 |
| Acer campestre | Forest species Querco-Fagetea | 245 | −65 |
| Rosa rugosa | Non-forest species, shrub species (Rhamno-Prunetea class) | 254 | −62 |
| Polygonatum odoratum | Non-forest species, grasses species Trifolo-Geranietea sanguinei | 261 | −60 |
| Prunella vulgaris | Non-forest species, grass species (Molinio-Arrhenatheretea class) | 286 | −55 |
| Aesculus hippocastanum | Associated species | 259 | −35 |
| Sorbus aucuparia | Associated species | 243 | −30 |
| Lamium purpureum | Non-forest species, synanthropical species (Artemisietea vulgaris class) | 287 | −29 |
| Species characteristic for manor parks adjacent to forests (PF) | |||
| Oxalis fontana | Associated species | 220 | 176 |
| Tilia tomentosa | Associated species | 202 | 178 |
| Trisetum flavescens | Non-forest species, grass species (Molinio-Arrhenatheretea class) | 241 | 183 |
| Urtica dioica | Non-forest species, synanthropical species (Artemisietea vulgaris class) | 240 | 189 |
| Sambucus nigra | Non-forest species, synanthropical species (Epilobietea angustifolii class) | 258 | 212 |
| Populus alba | Forest species, not consistent with oak-hornbeam habitat (Salicetea purpureae class) | 254 | 214 |
| Deschampsia flexuosa | Forest species, not consistent with oak-hornbeam habitat (Vaccinio-Piceetea class) | 233 | 219 |
| Galium sylvaticum | Forest species, consistent with oak-hornbeam habitat (Querco-Fageta class, Tilio-Carpinetum association) | 249 | 252 |
| Lamium album | Non-forest species, synanthropical species (Artemisietea vulgaris class) | 261 | 286 |
| Lapsana communis | Non-forest species, synanthropical species (Stellarietea mediae class) | 249 | 323 |
| Cerastium sylvaticum | Non-forest species, synanthropical species (Artemisietea vulgaris class) | 267 | 357 |
| Fragaria vesca | Non-forest species, synanthropical species (Epilobietea angustifolii class) | 259 | 461 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fornal-Pieniak, B.; Ollik, M.; Schwerk, A. Do Adjacent Forests Affect the Regeneration of Oak-Hornbeam and Ancient Forest Plant Species in Manor Parks in Poland? Forests 2021, 12, 538. https://doi.org/10.3390/f12050538
Fornal-Pieniak B, Ollik M, Schwerk A. Do Adjacent Forests Affect the Regeneration of Oak-Hornbeam and Ancient Forest Plant Species in Manor Parks in Poland? Forests. 2021; 12(5):538. https://doi.org/10.3390/f12050538
Chicago/Turabian StyleFornal-Pieniak, Beata, Marcin Ollik, and Axel Schwerk. 2021. "Do Adjacent Forests Affect the Regeneration of Oak-Hornbeam and Ancient Forest Plant Species in Manor Parks in Poland?" Forests 12, no. 5: 538. https://doi.org/10.3390/f12050538
APA StyleFornal-Pieniak, B., Ollik, M., & Schwerk, A. (2021). Do Adjacent Forests Affect the Regeneration of Oak-Hornbeam and Ancient Forest Plant Species in Manor Parks in Poland? Forests, 12(5), 538. https://doi.org/10.3390/f12050538