
Genetic Diversity and Association Analysis among Germplasms of Diospyros kaki in Zhejiang Province Based on SSR Markers


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Simple Sequence Repeat (SSR) Genotyping
2.3. Data Analysis
3. Results
3.1. Characterization of Morphological Traits Persimmon Germplasms
3.2. Characterization of Microsatellite Marker Polymorphism
3.3. Establishment of Genetic Information in Germplasms Populations
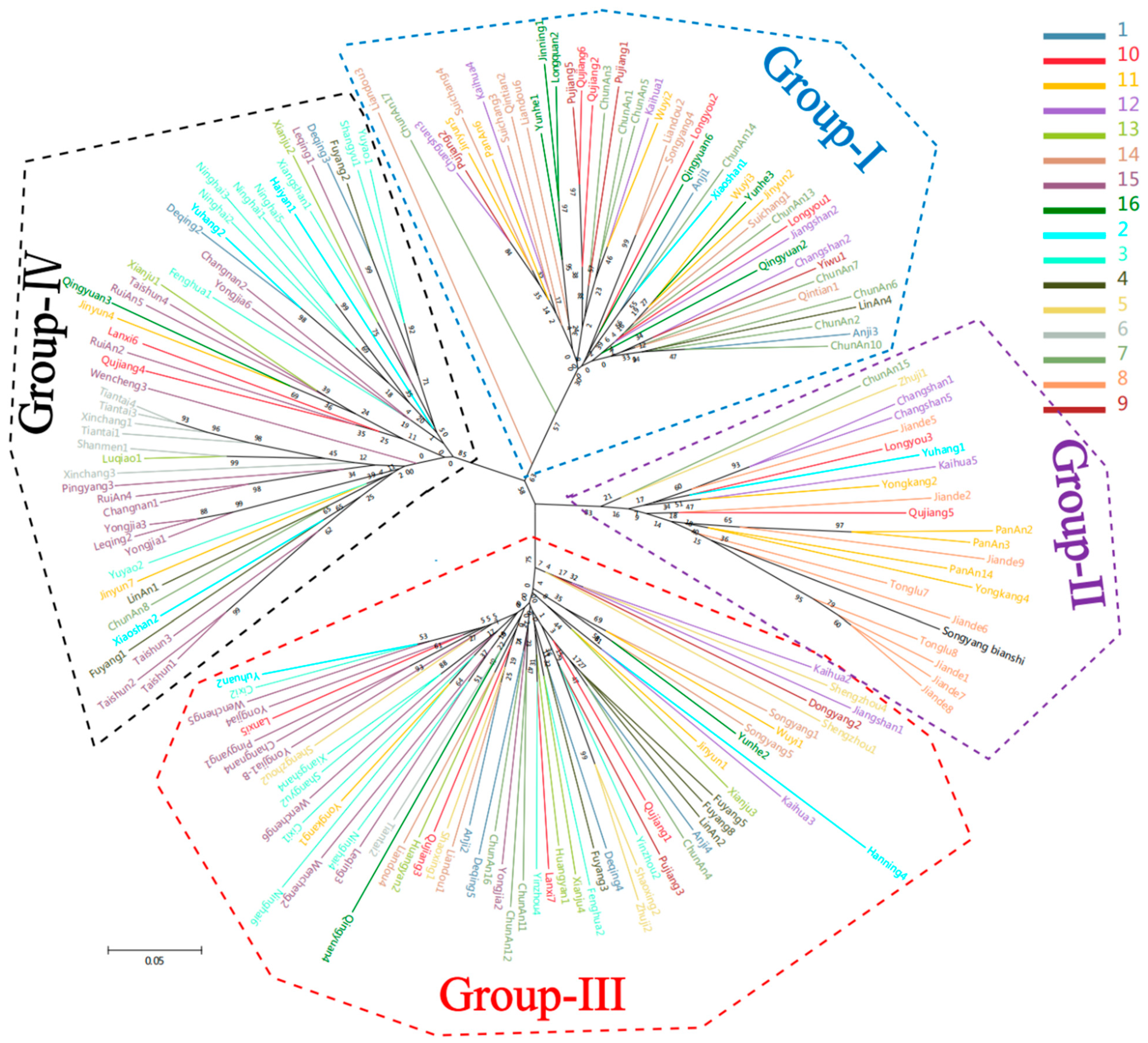
3.4. Establishment of Genetic Information in Clusters
3.5. Association Mapping
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duangjai, S.; Wallnöfer, B.; Samuel, R.; Munzinger, J.; Chase, M.W. Generic delimitation and relationships in Ebenaceae sensu lato: Evidence from six plastid DNA regions. Am. J. Bot. 2006, 93, 1808–1827. [Google Scholar] [CrossRef]
- Luo, Z.R.; Wang, R.Z. Persimmon in China: Domestication and traditional utilizations of genetic resources. Adv. Hortic. Sci. 2008, 22, 239–243. [Google Scholar]
- Yonemori, K.; Sugiura, A.; Yamada, M. Persimmon genetics and breeding. Plant Breed. Rev. 2000, 19, 191–225. [Google Scholar]
- Tang, D.L.; Zhang, Q.L.; Xu, L.Q.; Guo, D.L.; Luo, Z.R. Number of species and geographical distribution of Diospyros L. (Ebenaceae) in China. Hortic. Plant J. 2018, 5, 59–69. [Google Scholar] [CrossRef]
- Wang, R.Z.; Yang, Y.; Li, G.C. Chinese persimmon germplasm resources. Acta Hortic. 1997, 436, 43–50. [Google Scholar]
- Yang, Y.; Yang, T.T.; Jing, Z.B. Genetic diversity and taxonomic studies of date plum (Diospyros lotus L.) using morphological traits and SCoT markers. Biochem. Syst. Ecol. 2015, 61, 253–259. [Google Scholar] [CrossRef]
- Guo, D.L.; Luo, Z.R. Genetic relationships of some PCNA persimmons (Diospyros kaki Thunb.) from China and Japan revealed by SRAP analysis. Genet. Resour. Crop. Evol. 2006, 53, 1597–1603. [Google Scholar] [CrossRef]
- Du, X.Y.; Zhang, Q.L.; Luo, Z.R. Development of retrotransposon primers and their utilization for germplasm identification in Diospyros spp. (Ebenaceae). Tree Genet. Genomes 2009, 5, 235–245. [Google Scholar] [CrossRef]
- Yonemori, K.; Honsho, C.; Kitajima, A.; Aradhya, M.; Giordani, E.; Bellini, E.; Parfitt, D.E. Relationship of European persimmon (Diospyros kaki Thunb.) cultivars to Asian cultivars, characterized using AFLPs. Genet. Resour. Crop. Evol. 2008, 55, 81–89. [Google Scholar] [CrossRef]
- Parfitt, D.E.; Yonemori, K.; Honsho, C.; Nozaka, M.; Kanzaki, S.; Sato, A.; Yamada, M. Relationships among Asian persimmon cultivars, astringent and non-astringent types. Tree Genet. Genomes 2015, 51, 1–9. [Google Scholar] [CrossRef]
- Liang, Y.Q.; Han, W.J.; Sun, P.; Liang, J.; Wuyun, T.N.; Li, F.D.; Fu, J.M. Genetic diversity among germplasms of Diospyros kaki based on SSR markers. Sci. Hortic. 2015, 186, 180–189. [Google Scholar] [CrossRef]
- Guan, C.F.; Zhang, P.X.; Hu, C.Q.; Chachar, S.; Riaz, A.; Wang, R.Z.; Yang, Y. Genetic diversity, germplasm identification and population structure of Diospyros kaki Thunb. from different geographic regions in china using SSR markers. Sci. Hortic. 2019, 251, 233–240. [Google Scholar] [CrossRef]
- Zhao, X.M.; Gong, B.C.; Wu, K.Y.; Chen, H.X.; Lv, X.; Wang, N. Research on quantitative classification of native persimmon varieties in Zhejiang province. For. Res. 2012, 25, 77–87. (In Chinese) [Google Scholar]
- Zhao, X.M.; Gong, B.C.; Wu, K.Y.; Jiang, X.B.; Deng, Q.E.; Xiong, C.Y.; Zheng, X.F.; Chen, W.H. Variation analysis of fruit nutrients of native persimmon in Zhejiang province. J. Northwest. A. F. Univ. (Nat. Sci. Ed.) 2015, 43, 125–133. (In Chinese) [Google Scholar]
- Sun, W.M.; Xu, Y.; Gong, B.C.; Wu, K.Y. Study on fruit shape diversity of persimmon germplasm resources in Zhejiang province. For. Res. 2019, 32, 42–50. (In Chinese) [Google Scholar]
- Yu, J.; Buckler, E.S. Genetic association mapping and genome organization of maize. Curr. Opin. Biotechnol. 2006, 17, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Abdurakhmonov, I.Y.; Abdukarimov, A. Application of association mapping to understanding the genetic diversity of plant germplasm resources. Int. J. Plant. Genom. 2008, 574927. [Google Scholar] [CrossRef] [PubMed]
- Ambreen, H.; Kumar, S.; Kumar, A.; Agarwal, M.; Jagannath, A.; Goel, S. Association mapping for important agronomic traits in safflower (Carthamus tinctorius L.) core collection using microsatellite markers. Front. Plant. Sci. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.J.; Yu, J.W.; Xue, A.G.; Fan, S.L.; Song, M.Z.; Pang, C.Y.; Pei, W.F.; Yu, S.X.; Zhu, J. Dissecting genetic network of fruit branch traits in upland cotton by association mapping using SSR markers. PLoS ONE 2017, 12, e0162815. [Google Scholar] [CrossRef]
- Zheng, T.C.; Qin, B.; Li, S.Z.; Cai, M.; Zhang, Q.X. Screening of applicable SSR molecular markers linked to creeping trait in crape myrtle. Forests 2019, 10, 429. [Google Scholar] [CrossRef]
- Liao, G.L.; Li, Z.Y.; Huang, C.H.; Zhong, M.; Tao, J.J.; Qu, X.Y.; Chen, L.; Xu, X.X. Genetic diversity of inner quality and SSR association analysis of wild kiwifruit (Actinidia eriantha). Sci. Hortic. 2019, 248, 241–247. [Google Scholar] [CrossRef]
- Breria, C.M.; Hsieh, C.H.; Yen, J.Y.; Nair, R.; Schafleitner, R. Population structure of the world vegetable center mungbean mini core collection and genome-wide association mapping of loci associated with variation of seed coat luster. Trop Plant. Biol. 2019, 13, 1–12. [Google Scholar] [CrossRef]
- Ali, N.; Li, D.; Eltahawy, M.S.; Abdulmajid, D.; Bux, L.; Liu, E.; Dang, X.; Hong, D. Mining of favorable alleles for seed reserve utilization efficiency in Oryza sativa by means of association mapping. BMC Genet. 2020, 21, 4. [Google Scholar] [CrossRef]
- Liu, N.; Cheng, F.Y. Association mapping for yield traits in Paeonia rockii based on SSR markers within transcription factors of comparative transcriptome. BMC Plant. Biol. 2020, 20, 245. [Google Scholar] [CrossRef] [PubMed]
- Akagi, T.; Tao, R.; Tsujimoto, T.; Kono, A.; Yonemori, K. Fine genotyping of a highly polymorphic astringency-linked locus reveals variable hexasomic inheritance in persimmon (Diospyros kaki Thunb.) cultivars. Tree Genet. Genomes 2012, 8, 195–204. [Google Scholar] [CrossRef]
- Onoue, N.; Kobayashi, S.; Kono, A.; Sato, A. SSR-based molecular profiling of 237 persimmon (Diospyros kaki Thunb.) germplasms using an astringency-linked marker. Tree Genet. Genomes 2018, 14, 28. [Google Scholar] [CrossRef]
- Pei, X.; Zhang, Q.L.; Guo, D.Y.; Luo, Z.R. Effectiveness of the RO2 marker for the identification of non-astringency trait in Chinese PCNA persimmon and its possible segregation ratio in hybrid F1 population. Sci. Hortic. 2013, 150, 227–231. [Google Scholar] [CrossRef]
- Akagi, T.; Kajita, K.; Kibe, T.; Morimura, H.; Tsujimoto, T.; Nishiyama, S.; Kawai, T.; Yamane, H.; Tao, R. Development of molecular markers associated with sexuality in Diospyros lotus L. and their application in D. kaki Thunb. J. Jpn. Soc. Hortic. Sci. 2013, 83, 214–221. [Google Scholar] [CrossRef]
- Zhang, P.X.; Yang, S.C.; Liu, Y.F.; Zhang, Q.L.; Xu, L.Q.; Luo, Z.R. Validation of a male-linked gene locus (OGI) for sex identification in persimmon (Diospyros kaki Thunb.) and its application in F1 progeny. Plant. Breed. 2016, 135, 721–727. [Google Scholar] [CrossRef]
- Tong, Y.W.; Lewis, B.J.; Zhou, W.M.; Mao, C.R.; Wang, Y.; Zhou, L.; Yu, D.P.; Dai, L.M.; Qi, L. Genetic Diversity and Population Structure of Natural Pinus koraiensis Populations. Forests 2020, 11, 39. [Google Scholar] [CrossRef]
- Zuzana, B.; Jiri, K.; Jakub, D.; Jan, B.; Dagmar, Z.; Vaclav, J.; Milan, L. Genetic structure of norway spruce ecotypes studied by SSR markers. Forests 2020, 11, 110. [Google Scholar]
- Shi, X.M.; Wen, Q.; Cao, M.; Guo, X.; Xu, L.A. Genetic diversity and structure of natural Quercus variabilis population in china as revealed by microsatellites markers. Forests 2017, 8, 495. [Google Scholar] [CrossRef]
- Zhejiang Climate. Available online: http://zj.weather.com.cn/hyqx/index.shtml (accessed on 16 February 2020).
- State forestry administration of the People’s Republic of China. Technical Regulation of Cultivation for Japanese Persimmon (Diospyros kaki Thunb); LY/T 1887-2010 [S]; Standards Press of China: Beijing, China, 2010; pp. 06–10. [Google Scholar]
- Ross, I.; Robert, G. R: A Language for Data Analysis and Graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Soriano, J.M.; Pecchioli, S.; Romero, C.; Vilanova, S.; Llácer, G.; Giordani, E.; Badenes, M.L. Development of microsatellite markers in polyploid persimmon (Diospyros kaki Lf) from an enriched genomic library. Mol. Ecol. Notes 2006, 6, 368–370. [Google Scholar] [CrossRef]
- Ruan, X.F.; Gabi, K.; Yang, Y. Isolation of Microsatellite in Diospyros kaki by magnetic beads. J. Northwest. A. F. Univ. (Nat. Sci. Ed.) 2008, 36, 97–102. [Google Scholar]
- Guo, D.L. Establishment of Several Molecular Markers and Analysis of Genetic Relationships in Diospyros Linn. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2006. [Google Scholar]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef]
- Wang, Y.; Rashid, M.A.R.; Li, X.; Yao, C.; Lu, L.; Bai, J.; Li, Y.; Xu, N.; Yang, Q.; Zhang, L.; et al. Collection and Evaluation of Genetic Diversity and Population Structure of Potato Landraces and Varieties in China. Front. Plant. Sci. 2019, 10, 139. [Google Scholar] [CrossRef]
- Baytar, A.A.; Erdogan, O.; Frary, A.; Frary, A.; Doganlar, S. Molecular diversity and identification of alleles for Verticillium wilt resistance in elite cotton (Gossypium hirsutum L.) germplasm. Euphytica 2017, 213, 31. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Alderfasi, A.; Romdhane, W.B.; Seleiman, M.F.; EI-Said, R.A.; Al-Doss, A. Morphological and Genetic Diversity within Salt Tolerance Detection in Eighteen Wheat Genotypes. Plants 2020, 9, 287. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.B.; Singh, B.; Singh, R.K. Development of potential dbEST-derived microsatellite markers for genetic evaluation of sugarcane and related cereal grasses. Ind. Crop. Prod. 2019, 128, 38–47. [Google Scholar] [CrossRef]
- Singh, R.B.; Mahenderakar, M.D.; Jugran, A.K.; Singh, R.K.; Srivastava, R.K. Assessing genetic diversity and population structure of sugarcane cultivars, progenitor species and genera using microsatellite (SSR) markers. Gene 2020, 753, 144800. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.C.; Boyle, J. POPGENE, the user-friendly shareware for population genetic analysis. Mol. Biol. Biotechnol. 1997, 434, 724–731. [Google Scholar]
- Nagy, S.; Poczai, P.; Cernák, I.; Gorji, A.M.; Hegedűs, G.; Taller, J. PICcalc: An online program to calculate polymorphic information content for molecular genetic studies. Biochem. Genet. 2012, 50, 670–672. [Google Scholar] [CrossRef]
- Smouse, P.E.; Banks, S.C.; Peakall, R. Converting quadratic entropy to diversity: Both animals and alleles are diverse, but some are more diverse than others. PLoS ONE 2017, 12, e0185499. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Price, A.L.; Patterson, N.J.; Plenge, R.M.; Weinblatt, M.E.; Shadick, N.A.; Reich, D. Principal components analysis corrects for stratification in genome-wide association studies. Nat. Genet. 2006, 38, 904–909. [Google Scholar] [CrossRef]
- Manel, S.; Schwartz, M.K.; Luikart, G.; Taberlet, P. Landscape genetics combining landscape ecology and population genetics. Trends Ecol. Evol. 2003, 18, 189–197. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Earl, D.A.; Vonholdt, B.M. Structure Harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef]
- Francis, R.M. Pophelper: An R package and web app to analyse and visualize population structure. Mol. Ecol. Resour. 2017, 17, 27–32. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Wright, S. Genetic structure of populations. Br. Med. J. 1950, 2, 36. [Google Scholar] [PubMed]
- Quian-Ulloa, R.; Stange, C. Carotenoid Biosynthesis and Plastid Development in Plants: The Role of Light. Int. J. Mol. Sci. 2021, 22, 1184. [Google Scholar] [CrossRef]
- Mpai, S.; Sivakumar, D. Stimulation of Light-Emitting Diode Treatment on Defence System and Changes in Mesocarp Metabolites of Avocados Cultivars (Hass and Fuerte) during Simulated Market Shelf Conditions. Agronomy 2020, 10, 1654. [Google Scholar] [CrossRef]
- Peavey, M.; Goodwin, I.; McClymont, L. The Effects of Canopy Height and Bud Light Exposure on the Early Stages of Flower Development in Prunus persica (L.) Batsch. Plants 2020, 9, 1073. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.; Kalcsits, L. Water Deficit Timing Affects Physiological Drought Response, Fruit Size, and Bitter Pit Development for ‘Honeycrisp’ Apple. Plants 2020, 9, 874. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.B.; Gabira, M.M.; Prado, D.Z.; Uesugi, G.; Simões, D.; Silva, M.R. Influence of Mean Leaf Angles and Irrigation Volumes on Water Capture, Leaching, and Growth of Tropical Tree Seedlings. Forests 2020, 11, 1198. [Google Scholar] [CrossRef]
- Anna, K.-I.; Sylwia, Ł.; Marcin, Z.; Ewa, S.-O.; Wojtan, B. Variability of Leaf Wetting and Water Storage Capacity of Branches of 12 Deciduous Tree Species. Forests 2020, 11, 1158. [Google Scholar] [CrossRef]
- Arenas-Navarro, M.; García-Oliva, F.; Terrazas, T.; Torres-Miranda, A.; Oyama, K. Leaf Habit and Stem Hydraulic Traits Determine Functional Segregation of Multiple Oak Species along a Water Availability Gradient. Forests 2020, 11, 894. [Google Scholar] [CrossRef]
- Kim, C.-B.; Kim, Y.S.; Choi, H.T.; Kim, J.; Kim, S.; Cha, S.; Gao, G.-L.; Bao, Y.-F.; Son, Y.; Kwon, J.; et al. Sand Dune Height Increases Water Use Efficiency at the Expense of Growth and Leaf Area in Mongolian Pine Growing in Hulunbeier Steppe, Inner Mongolia, China. Forests 2019, 10, 558. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar]
- Luo, C.; Zhang, F.; Zhang, Q.L.; Guo, D.Y.; Luo, Z.R. Characterization and comparison of EST-SSR and TRAP markers for genetic analysis of the Japanese persimmon Diospyros kaki. Genet. Mol. Res. 2013, 12, 2841–2851. [Google Scholar] [CrossRef]
- Naval, M.D.M.; Zuriaga, E.; Pecchioli, S.; Llácer, G.; Giordani, E.; Badenes, M.L. Analysis of genetic diversity among persimmon cultivars using microsatellite markers. Tree Genet. Genomes 2010, 6, 677–687. [Google Scholar] [CrossRef][Green Version]
- Deng, L.B.; Liang, Q.Z.; He, X.H.; Luo, C.; Chen, H.; Qin, Z.S. Investigation and Analysis of Genetic Diversity of Diospyros Germplasms Using SCoT Molecular Markers in Guangxi. PLoS ONE 2015, 10, e0136510. [Google Scholar] [CrossRef]
- Jump, A.S.; Hunt, J.M.; Martínez-Izquierdo, J.A.; Peñuelas, J. Natural selection and climate change: Temperature-linked spatial and temporal trends in gene frequency in Fagus sylvatica. Mol. Ecol. 2006, 15, 3469–3480. [Google Scholar] [CrossRef]
- Kelly, C.K.; Chase, M.W.; De Bruijn, A.; Fay, M.F.; Woodward, F.I. Temperature-based population segregation in birch. Ecol. Lett. 2003, 6, 87–89. [Google Scholar] [CrossRef]
- Bradbury, D.; Smithson, A.; Krauss, S.L. Signatures of diversifying selection at EST-SSR loci and association with climate in natural Eucalyptus populations. Mol. Ecol. 2013, 22, 5112–5129. [Google Scholar] [CrossRef]
- Müller, M.; Nelson, C.D.; Gailing, O. Analysis of Environment-Marker Associations in American Chestnut. Forests 2018, 9, 695. [Google Scholar] [CrossRef]
- Chen, Z.; Wu, B.; Li, J.; Zhao, T.; Zhou, X.; Zhang, Y. Germination of the seeds and growth of the seedlings of Cinnamomum camphora (L.) Presl. Plant. Species Biol. 2004, 19, 55–58. [Google Scholar] [CrossRef]
- Doh, E.J.; Kim, J.H.; Oh, S.E.; Lee, G. Identification and monitoring of Korean medicines derived from Cinnamomum spp. by using ITS and DNA marker. Genes Genom. 2017, 39, 101–109. [Google Scholar] [CrossRef]
- Panetta, F.D. Seedling emergence and seed longevity of the tree weeds Celtis sinensis and Cinnamomum camphora. Weed Res. 2010, 41, 83–95. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, J.H.; Zhao, Y.Q.; Wang, Y.; Wang, X.M.; Liu, W.D.; Shi, J.S. Variation of EST-SSR molecular markers among provenances of Chinese fir. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2014, 57, 1–8. [Google Scholar]
- Kitamura, K.; Uchiyama, K.; Ueno, S.; Ishizuka, W.; Tsuyama, I.; Goto, S. Geographical Gradients of Genetic Diversity and Differentiation among the Southernmost Marginal Populations of Abies sachalinensis Revealed by EST-SSR Polymorphism. Forests 2020, 11, 233. [Google Scholar] [CrossRef]
- Zeng, Y.F.; Wang, W.T.; Liao, W.J.; Wang, H.F.; Zhang, D.Y. Multiple glacial refugia for cool-temperate deciduous trees in northern East Asia: The Mongolian oak as a case study. Mol. Ecol. 2015, 24, 5676–5691. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Hassanpouraghdam, M.B.; Paliyath, G.; Farmani, B. The language of calcium in postharvest life of fruits, vegetables and flowers. Sci. Hortic. 2012, 144, 102–115. [Google Scholar] [CrossRef]
- Katayama-Ikegami, A.; Yonemori, K.; Sato, A.; Yamada, M.; Habu, T.; Kitajima, A. Relationship between astringency type and fruit shape in progenies of Chinese PCNA persimmon. Hortic. Res. 2013, 12, 29–34. (In Japanese) [Google Scholar] [CrossRef]
- Maeda, H.; Akagi, T.; Tao, R. Quantitative characterization of fruit shape and its differentiation pattern in diverse persimmon (Diospyros kaki) cultivars. Sci. Hortic. 2018, 228, 41–48. [Google Scholar] [CrossRef]
- Yamamura, H.; Naito, R. GA-like substances in immature fruit of kaki (Diospyros kaki L.). J. Jpn. Soc. Hort. Sci. 1973, 42, 1–6. [Google Scholar] [CrossRef][Green Version]
- Maeda, H.; Akagi, T.; Onoue, N.; Kono, A.; Tao, R. Evolution of Lineage-Specific Gene Networks Underlying the Considerable Fruit Shape Diversity in Persimmon. Plant. Cell Physiol. 2019, 60, 2464–2477. [Google Scholar] [CrossRef]
- Resende, R.T.; Resende, M.D.; Silva, F.F.; Azevedo, C.F.; Takahashi, E.K.; Silva-Junior, O.B.; Grattapaglia, D. Regional heritability mapping and genome-wide association identify loci for complex growth, wood and disease resistance traits in Eucalyptus. New Phytol. 2017, 213, 1287–1300. [Google Scholar] [CrossRef]
- Cao, K.; Zhou, Z.; Wang, Q.; Guo, J.; Zhao, P.; Zhu, G.; Fang, W.; Chen, C.; Wang, X.; Wang, X.; et al. Genome-wide association study of 12 agronomic traits in peach. Nat. Commun. 2016, 7, 13246. [Google Scholar] [CrossRef] [PubMed]
- Farneti, B.; Di Guardo, M.; Khomenko, I.; Cappellin, L.; Biasioli, F.; Velasco, R.; Costa, F. Genome-wide association study unravels the genetic control of the apple volatilome and its interplay with fruit texture. J. Exp. Bot. 2017, 68, 1467–1478. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.M. Post-floral perianth functionality: Contribution of persistent sepals to seed development in Helleborus foetidus (Ranunculaceae). Am. J. Bot. 2005, 92, 1486–1491. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pop No. | Full Name of Population | Abbreviation | Full Name of Climatic Factor |
|---|---|---|---|
| Pop 1 | Hill north of Hang Jia Hu Plain | SprPR | Precipitation in spring (March to May) |
| Pop 2 | Hang Jia Hu Plain | SumPR | Precipitation in summer (June to August) |
| Pop 3 | Ningshao Plain | AutPR | Precipitation in autumn (September to November) |
| Pop 4 | Western Hang Jia Hu Plain | WinPR | Precipitation in winter (December to February) |
| Pop 5 | Eastern Zhejiang Hill | SprSR | Solar radiation in spring (March to May) |
| Pop 6 | Tiantai Mountain | SumSR | Solar radiation in summer (June to August) |
| Pop 7 | Thousand Island Lake | AutSR | Solar radiation in autumn (September to November) |
| Pop 8 | Xin’an River | WinSR | Solar radiation in winter (December to February) |
| Pop 9 | Eastern Qujing Basin | SprMT | Mean temperature in spring (March to May) |
| Pop 10 | JingQu Basin | SumMT | Mean temperature in summer (June to August) |
| Pop 11 | The eastern hills of JingQu Basin | AutMT | Mean temperature in autumn (September to November) |
| Pop 12 | Xianxia Mountain and Qian Li Gang | WinMT | Mean temperature in winter (December to February) |
| Pop 13 | Wenhuang Plain | ||
| Pop 14 | Songgu Basin and Lishui Basin | ||
| Pop 15 | Yandang Mountain and Wenrui Plain | ||
| Pop 16 | Cave mountain range |
| Pop No. | FCR (mg/100 g) | FCC (mg/100 g) | FWC (%) | FLD | FAR | SW (mm) | SL (mm) | LAR | NLV |
|---|---|---|---|---|---|---|---|---|---|
| Pop 1 | 0.89 ± 0.26 a,b,c | 7.81 ± 2.45 a,b,c,d | 78.35 ± 2.14 b,c,d,e | 50.65 ± 8.26 b,c | 0.93 ± 0.18 a,b | 11.27 ± 1.3 a,b | 20.75 ± 4.35 b,c,d,e | 1.74 ± 0.2 a | 3.61 ± 0.26 e |
| Pop 2 | 0.88 ± 0.39 a,b,c | 8.71 ± 1.92 a,b,c | 77.11 ± 6.23 c,d,e | 49.87 ± 10.26 c | 0.86 ± 0.24 b | 11.19 ± 1.83 a,b | 18.15 ± 3.59 e | 1.74 ± 0.13 a | 4.22 ± 0.73 b,c,d |
| Pop 3 | 0.75 ± 0.15 a,b,c,d | 8.97 ± 1.81 a,b | 83.39 ± 1.51 a | 53.68 ± 9.51 a,b,c | 0.91 ± 0.25 a,b | 11.66 ± 2.62 a,b | 22.06 ± 3.79 a,b,c,d | 1.66 ± 0.22 a | 4.91 ± 0.28 a |
| Pop 4 | 1.18 ± 0.29 a,b | 7.66 ± 2.94 a,b,c,d | 79.44 ± 2.72 b,c,d | 55.66 ± 8.72 a,b,c | 1.03 ± 0.15 a,b | 12.1 ± 2.03 a | 20.1 ± 1.62 c,d,e | 1.77 ± 0.14 a | 4.41 ± 0.4 b,c,d |
| Pop 5 | 0.67 ± 0.07 c,d | 6.46 ± 1.73 b,c,d | 78.89 ± 3.17 b,c,d,e | 49.57 ± 6.8 c | 1.03 ± 0.25 a,b | 11.63 ± 4.65 a,b | 21.08 ± 3.96 a,b,c,d,e | 1.65 ± 0.07 a | 4.6 ± 0.93 a,b,c |
| Pop 6 | 0.91 ± 0.33 a,b,c | 7.71 ± 1.28 a,b,c,d | 80.66 ± 3.06 a,b | 61.62 ± 8.24 a | 1.16 ± 0.27 a | 10.69 ± 1.22 a,b | 24.55 ± 3.58 a | 1.6 ± 0.12 a,b | 4.19 ± 0.81 b,c,d |
| Pop 7 | 0.77 ± 0.19 a,b,c,d | 6.19 ± 3.26 c,d | 75.94 ± 3.96 e | 55.79 ± 8.87 a,b,c | 1.08 ± 0.18 a,b | 9.75 ± 1.11 b | 22.06 ± 2.76 a,b,c,d | 1.77 ± 0.23 a | 4.08 ± 0.49 d,e |
| Pop 8 | 0.43 ± 0.09 d | 5.34 ± 2.03 d | 80.4 ± 1.57 a,b | 52.56 ± 8.23 a,b,c | 0.96 ± 0.17 a,b | 10.08 ± 0.82 b | 19.79 ± 3.75 c,d,e | 1.64 ± 0.2 a | 3.61 ± 0.31 e |
| Pop 9 | 0.79 ± 0.18 a,b,c,d | 6.42 ± 2.62 b,c,d | 77.91 ± 1.2 b,c,d,e | 56.04 ± 13.46 a,b,c | 1.12 ± 0.36 a | 10.15 ± 1.13 b | 19.1 ± 4.86 d,e | 1.68 ± 0.11 a | 4.15 ± 0.42 c,d |
| Pop 10 | 0.69 ± 0.14 c,d | 7.18 ± 2.24 a,b,c,d | 80.32 ± 2.47 a,b,c | 47.49 ± 9.55 c | 0.92 ± 0.2 a,b | 11.09 ± 1.4 a,b | 21.08 ± 2.84 a,b,c,d,e | 1.65 ± 0.17 a | 3.95 ± 0.15 d,e |
| Pop 11 | 0.85 ± 0.34 a,b,c,d | 8.75 ± 3.02 a,b,c | 78.49 ± 4.18 b,c,d,e | 48.14 ± 9.67 c | 1.04 ± 0.26 a,b | 10.48 ± 1.67 a,b | 19.45 ± 3.19 c,d,e | 1.7 ± 0.17 a | 4.45 ± 0.42 a,b,c,d |
| Pop 12 | 1.19 ± 1.38 a | 7.04 ± 1.59 a,b,c,d | 76.23 ± 2.65 d,e | 50.59 ± 6.66 b,c | 1.12 ± 0.14 a | 10.14 ± 1.23 b | 19.41 ± 1.9 c,d,e | 1.61 ± 0.13 a,b | 4.68 ± 0.48 a,b |
| Pop 13 | 0.74 ± 0.17 b,c,d | 6.97 ± 0.99 a,b,c,d | 80.01 ± 0.48 b,c | 60.82 ± 11.49 a,b | 1.17 ± 0.34 a | 10.85 ± 1.29 a,b | 23.86 ± 2.48 a,b | 1.61 ± 0.17 a,b | 4.43 ± 0.33 b,c,d |
| Pop 14 | 0.84 ± 0.22 a,b,c,d | 8.71 ± 3.2 a,b,c | 76.66 ± 4.29 d,e | 54.49 ± 9.39 a,b,c | 1.13 ± 0.22 a | 9.8 ± 1 b | 21.02 ± 2.82 a,b,c,d,e | 1.61 ± 0.2 a,b | 4.09 ± 0.4 d,e |
| Pop 15 | 0.81 ± 0.16 a,b,c,d | 9.18 ± 2.49 a | 80.94 ± 2.21 a,b | 57.25 ± 11.72 a,b,c | 1.09 ± 0.28 a,b | 11.21 ± 1.62 a,b | 22.85 ± 3.99 a,b,c | 1.72 ± 0.16 a | 4.42 ± 0.47 b,c,d |
| Pop 16 | 0.9 ± 0.41 a,b,c | 6.74 ± 2.55 a,b,c,d | 78.72 ± 2.79 b,c,d,e | 49.36 ± 12.42 c | 1 ± 0.2 a,b | 9.92 ± 1.07 b | 21.09 ± 3.02 a,b,c,d,e | 1.44 ± 0.12 b | 3.66 ± 0.55 e |
| All | 0.82 ± 0.42 | 7.74 ± 2.62 | 79.16 ± 3.66 | 53.39 ± 10.18 | 1.03 ± 0.24 | 10.74 ± 1.84 | 21.21 ± 3.57 | 1.67 ± 0.18 | 4.26 ± 0.58 |
| Min * | 0.32 | 2.61 | 64.13 | 24.9 | 0.63 | 10.11 | 6.46 | 2.97 | 1.11 |
| Max # | 5.32 | 16.01 | 85.63 | 81.56 | 1.74 | 30.98 | 21.98 | 6 | 2.16 |
| Item | SprPR | SumPR | AutPR | WinPR | SprSR | SumSR | AutSR | WinSR | SprMT | SumMT | AutMT | WinMT | Longitude | Latitude | Altitude |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FCR | 0.081 | −0.089 | −0.031 | 0.013 | −0.056 | 0.014 | 0.04 | −0.026 | 0.071 | 0.064 | 0.095 | 0.06 | −0.093 | −0.028 | 0.021 |
| FCC | −0.159 * | 0.122 | 0.190 * | −0.187 * | −0.082 | −0.232 ** | −0.155 * | 0.156 * | −0.142 | 0.037 | 0.079 | 0.125 | 0.250 ** | −0.190 * | −0.004 |
| FWC | −0.248 ** | 0.091 | 0.369 ** | −0.142 | 0.116 | −0.195 ** | −0.270 ** | 0.208 ** | −0.312 ** | −0.018 | 0.061 | 0.003 | 0.410 ** | −0.052 | −0.09 |
| FLD | −0.085 | −0.09 | 0.127 | −0.225 ** | 0.058 | −0.109 | −0.122 | 0.098 | −0.082 | −0.015 | 0.102 | 0.142 | 0.198 ** | −0.023 | −0.041 |
| FAR | 0.117 | −0.076 | −0.042 | −0.177 * | −0.146 | −0.091 | 0.019 | 0.016 | 0.063 | −0.083 | −0.018 | 0.067 | 0.006 | −0.148 * | 0.071 |
| SW | −0.218 ** | −0.094 | 0.228 ** | −0.04 | 0.161 * | −0.056 | −0.179 * | 0.044 | −0.175 * | 0.13 | 0.07 | 0.026 | 0.233 ** | 0.077 | −0.141 |
| SL | −0.133 | 0.04 | 0.188 * | −0.187 * | 0.004 | −0.171 * | −0.121 | 0.111 | −0.061 | 0.063 | 0.164 * | 0.146 | 0.187 * | −0.118 | −0.075 |
| LAR | −0.218 ** | −0.096 | 0.039 | 0.052 | 0.166 * | 0.145 | −0.055 | −0.133 | −0.05 | 0.14 | 0.041 | −0.075 | −0.011 | 0.217 ** | −0.076 |
| NLV | −0.067 | −0.002 | 0.320** | −0.046 | 0.106 | −0.081 | −0.206 ** | −0.111 | −0.173 * | 0.059 | 0.009 | 0.044 | 0.329 ** | −0.006 | −0.039 |
| Pop No. | Sample Size | Np | Ne | Nei | I | He | PIC |
|---|---|---|---|---|---|---|---|
| Pop 1 | 8 | 190 | 1.5826 | 0.3524 | 0.4836 | 0.3304 | 0.2606 |
| Pop 2 | 7 | 196 | 1.5963 | 0.3655 | 0.4975 | 0.3394 | 0.2681 |
| Pop 3 | 18 | 208 | 1.5932 | 0.3507 | 0.5042 | 0.341 | 0.2707 |
| Pop 4 | 8 | 189 | 1.5624 | 0.3424 | 0.4721 | 0.321 | 0.254 |
| Pop 5 | 7 | 174 | 1.4984 | 0.311 | 0.4275 | 0.2887 | 0.2299 |
| Pop 6 | 7 | 151 | 1.3241 | 0.2242 | 0.3236 | 0.2082 | 0.1725 |
| Pop 7 | 16 | 204 | 1.5388 | 0.325 | 0.4711 | 0.3149 | 0.2519 |
| Pop 8 | 9 | 143 | 1.384 | 0.2379 | 0.336 | 0.2247 | 0.1799 |
| Pop 9 | 6 | 177 | 1.5078 | 0.3237 | 0.4392 | 0.2967 | 0.2367 |
| Pop 10 | 12 | 210 | 1.6287 | 0.3718 | 0.5232 | 0.3563 | 0.2813 |
| Pop 11 | 15 | 215 | 1.6224 | 0.3694 | 0.5268 | 0.3571 | 0.2831 |
| Pop 12 | 11 | 197 | 1.565 | 0.341 | 0.4814 | 0.3255 | 0.2586 |
| Pop 13 | 7 | 178 | 1.5251 | 0.3252 | 0.4449 | 0.3019 | 0.2395 |
| Pop 14 | 14 | 191 | 1.5446 | 0.3243 | 0.4624 | 0.3127 | 0.2482 |
| Pop 15 | 25 | 208 | 1.6113 | 0.3527 | 0.5076 | 0.3456 | 0.2725 |
| Pop 16 | 9 | 199 | 1.5281 | 0.3293 | 0.466 | 0.311 | 0.2495 |
| Mean | 11.19 | 189.37 | 1.5383 | 0.3279 | 0.4604 | 0.3109 | 0.2473 |
| Pop | P7 | P16 | P9 | P14 | P1 | P10 | P4 | P2 | P11 | P12 | P8 | P5 | P3 | P13 | P15 | P6 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P7 | 0 | |||||||||||||||
| P16 | 0.0826 | 0 | ||||||||||||||
| P9 | 0.0321 | 0.0597 | 0 | |||||||||||||
| P14 | 0.061 | 0.056 | 0.0462 | 0 | ||||||||||||
| P1 | 0.0679 | 0.0991 | 0.0722 | 0.0749 | 0 | |||||||||||
| P10 | 0.0706 | 0.0743 | 0.0583 | 0.0755 | 0.0436 | 0 | ||||||||||
| P4 | 0.1185 | 0.145 | 0.13 | 0.1284 | 0.0401 | 0.0528 | 0 | |||||||||
| P2 | 0.1066 | 0.116 | 0.1279 | 0.1189 | 0.0446 | 0.0608 | 0.0665 | 0 | ||||||||
| P11 | 0.0708 | 0.0762 | 0.0494 | 0.0551 | 0.0547 | 0.0458 | 0.0981 | 0.0867 | 0 | |||||||
| P12 | 0.0665 | 0.0744 | 0.0414 | 0.0447 | 0.0851 | 0.0612 | 0.1468 | 0.1144 | 0.0289 | 0 | ||||||
| P8 | 0.3013 | 0.336 | 0.3243 | 0.3002 | 0.3095 | 0.2351 | 0.3348 | 0.3023 | 0.1947 | 0.2296 | 0 | |||||
| P5 | 0.1791 | 0.1943 | 0.1727 | 0.161 | 0.0974 | 0.0918 | 0.1222 | 0.1725 | 0.1279 | 0.1542 | 0.3337 | 0 | ||||
| P3 | 0.1508 | 0.1592 | 0.169 | 0.1611 | 0.0486 | 0.0743 | 0.0567 | 0.0496 | 0.1182 | 0.1653 | 0.3095 | 0.1294 | 0 | |||
| P13 | 0.1876 | 0.1981 | 0.2193 | 0.193 | 0.0866 | 0.1044 | 0.0986 | 0.0951 | 0.1583 | 0.2083 | 0.3912 | 0.1511 | 0.0552 | 0 | ||
| P15 | 0.1639 | 0.1539 | 0.1769 | 0.1646 | 0.0787 | 0.0834 | 0.0891 | 0.0628 | 0.1224 | 0.1659 | 0.3101 | 0.1563 | 0.0427 | 0.0664 | 0 | |
| P6 | 0.3144 | 0.3277 | 0.3579 | 0.3233 | 0.2509 | 0.2453 | 0.2305 | 0.2063 | 0.2717 | 0.3362 | 0.497 | 0.3797 | 0.177 | 0.2236 | 0.151 | 0 |
| Cluster | Sample Size | Np | Ne | Nei | I | He | PIC |
|---|---|---|---|---|---|---|---|
| Cluster-I | 70 | 213 | 1.5127 | 0.3041 | 0.4554 | 0.3015 | 0.2422 |
| Cluster-II | 61 | 220 | 1.5493 | 0.3211 | 0.4781 | 0.3188 | 0.2546 |
| Cluster-III | 48 | 198 | 1.5026 | 0.2985 | 0.4433 | 0.2954 | 0.2367 |
| Mean | 59.7 | 210.33 | 1.5215 | 0.3079 | 0.45893 | 0.3052 | 0.2445 |
| Organ | Traits | SSR Locus * | GLM | MLM | ||
|---|---|---|---|---|---|---|
| p-Value | R2 (%) | p-Value | R2 (%) | |||
| Fruit | Fruit crude fiber content | DKMP17-282 | 0.00024404 | 9.2 | 0.00033412 | 9 |
| DK11-197 | 0.00024404 | 8.56 | 0.00216 | 6.5 | ||
| Fruit calcium content | DKMP18-156 | 0.000072152 | 10.19 | 0.01611 | 4.24 | |
| Fruit water content | DKMP13-440 | 0.0004181 | 6.28 | 0.01158 | 4.02 | |
| Fruit longitudinal diameter | DK26-184 | 0.00022922 | 7.42 | 0.01737 | 3.4 | |
| Fruit aspect ratio | MDP18-310 | 7.6122 × 10−7 | 13.53 | 0.00056065 | 7.39 | |
| Seed | Seed width | DKMP7-600 | 0.0001777 | 7.81 | 0.0021 | 6.14 |
| MDP8-288 | 0.000073631 | 8.69 | 0.00046859 | 8.01 | ||
| Seed length | DKMP18-190 | 0.00028986 | 7.94 | 0.01601 | 3.87 | |
| MDP18-310 | 0.000042848 | 10.01 | 0.00603 | 5.06 | ||
| Leaf | Leaf aspect ratio | DKMP15-514 | 0.00020945 | 9.29 | 0.01005 | 4.81 |
| Number of lateral veins | DK11-297 | 0.00013218 | 9.48 | 0.00027251 | 9.51 | |
| DK11-271 | 0.000072327 | 10.18 | 0.00016836 | 10.19 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Cheng, W.; Xiong, C.; Jiang, X.; Wu, K.; Gong, B. Genetic Diversity and Association Analysis among Germplasms of Diospyros kaki in Zhejiang Province Based on SSR Markers. Forests 2021, 12, 422. https://doi.org/10.3390/f12040422
Xu Y, Cheng W, Xiong C, Jiang X, Wu K, Gong B. Genetic Diversity and Association Analysis among Germplasms of Diospyros kaki in Zhejiang Province Based on SSR Markers. Forests. 2021; 12(4):422. https://doi.org/10.3390/f12040422
Chicago/Turabian StyleXu, Yang, Wenqiang Cheng, Chunyan Xiong, Xibing Jiang, Kaiyun Wu, and Bangchu Gong. 2021. "Genetic Diversity and Association Analysis among Germplasms of Diospyros kaki in Zhejiang Province Based on SSR Markers" Forests 12, no. 4: 422. https://doi.org/10.3390/f12040422
APA StyleXu, Y., Cheng, W., Xiong, C., Jiang, X., Wu, K., & Gong, B. (2021). Genetic Diversity and Association Analysis among Germplasms of Diospyros kaki in Zhejiang Province Based on SSR Markers. Forests, 12(4), 422. https://doi.org/10.3390/f12040422