Abstract
The genus Quercus (oak), family Fagaceae, comprises around 500 species, being one of the most important and dominant woody angiosperms in the Northern Hemisphere. Nowadays, it is threatened by environmental cues, which are either of biotic or abiotic origin. This causes tree decline, dieback, and deforestation, which can worsen in a climate change scenario. In the 21st century, biotechnology should take a pivotal role in facing this problem and proposing sustainable management and conservation strategies for forests. As a non-domesticated, long-lived species, the only plausible approach for tree breeding is exploiting the natural diversity present in this species and the selection of elite, more resilient genotypes, based on molecular markers. In this direction, it is important to investigate the molecular mechanisms of the tolerance or resistance to stresses, and the identification of genes, gene products, and metabolites related to this phenotype. This research is being performed by using classical biochemistry or the most recent omics (genomics, epigenomics, transcriptomics, proteomics, and metabolomics) approaches, which should be integrated with other physiological and morphological techniques in the Systems Biology direction. This review is focused on the current state-of-the-art of such approaches for describing and integrating the latest knowledge on biotic and abiotic stress responses in Quercus spp., with special reference to Quercus ilex, the system on which the authors have been working for the last 15 years. While biotic stress factors mainly include fungi and insects such as Phytophthora cinnamomi, Cerambyx welensii, and Operophtera brumata, abiotic stress factors include salinity, drought, waterlogging, soil pollutants, cold, heat, carbon dioxide, ozone, and ultraviolet radiation. The review is structured following the Central Dogma of Molecular Biology and the omic cascade, from DNA (genomics, epigenomics, and DNA-based markers) to metabolites (metabolomics), through mRNA (transcriptomics) and proteins (proteomics). An integrated view of the different approaches, challenges, and future directions is critically discussed.
1. Introduction
The genus Quercus (oak), family Fagaceae, comprises around 500 species, being one of the most important and dominant woody angiosperms in the Northern Hemisphere, with relevance from an environmental and economic point of view [1,2]. It includes shrubs and trees, either deciduous or evergreen, which are part of temperate and sub-tropical forests, sub-tropical and tropical savannah, and subtropical woodlands. They are also part of typical agrosilvopastoral systems, such as the Spanish “dehesa” and the Portuguese “montado”, with Quercus ilex and Quercus suber, respectively, being the dominant species [3].
Forest trees are long-lived organisms that have played—since ancient times—an important role in the different human civilizations. They are considered key elements of the Earth’s health and biodiversity, and are associated with our culture and spirituality, providing countless products and services. They must be conserved as unvaluable heritage for future generations. However, humans are taking the opposite direction, with deforestation, clearing, or thinning of forests, which have been constant throughout history. According to Williams’ estimation (reviewed in Seale [4]), humans have destroyed around 1.8 billion Ha over the past 5000 years, with an average of 360 Kha/year. The figures are more dramatic in modern times. Therefore, and according to Food and Agriculture Organization (FAO) [5], since 1990, 420 Mha of forest have been lost, with rates ranging from 16 Mha/year in the 1990s to 10 Mha in the last quinquennium.
Causes such as forest degradation, fragmentation, and loss, as well as anthropogenic ones (overexploitation, fires, and agricultural land conversions, among others), are the main reasons for the high increase in tree mortality and deforestation [6]. Moreover, adverse environmental factors play an important negative role in forest conservation. Such factors, which are termed stresses, are physical, chemical, or biological agents that impose restrictions for the growth and development, and depending on their intensity and duration, they may cause death [7]. Nowadays, the decline syndrome, affecting the genus Quercus and other species, is a good example, and an international concern. Oak decline is a complex syndrome in which several damaging biotic and abiotic agents interact, and together with improper practices, old individuals, and a lack of regeneration, it is causing progressive and massive dieback. The interrelation between abiotic (e.g., drought and high temperature) and biotic stresses (e.g., fungi and insects) is complex, and not properly explained from an experimental point of view. Oak decline diseases are examined in Europe, the Middle East, and North America [8]. Oak trees are predisposed to acute oak decline (AOD) attacks by bacterial species and environmental factors [9,10]. In England, AOD affects mature Quercus robur and Quercus petraea trees, triggering stem bleeds from vertical fissures on trunks and inner bark necrosis caused by gram-negative bacteria such as Gibbsiella quercinecans and Brenneria goodwinii, and, to a lesser extent, Rahnella victoriana and Lonsdalea britannica [11,12,13,14]. On the other hand, the Phytophthora spp. is an oomycete considered to be one of the most lethal pathogens described in oak trees [15,16]. In Spain and Portugal, the synergy between Phytophthora cinnamomi Rands and long drought periods is increasing Q. ilex and Q. suber mortality [17,18,19]. Other pathogens affecting oaks, such as Phytium spiculum Paul and Biscogniauxia mediterranea (De Not) Kuntze, are also considered to be part of the oak decline syndrome [20,21].
Extreme environmental conditions in a climate change scenario represent an issue of great relevance that will determine species survival and migration, as well as the forest composition, structure, and functionality [22,23]. Simulation models predict an increase in both temperature and the frequency of severe drought episodes [24,25], or in northern latitudes the increased wetness, which will determine forest vulnerability. How can we afford the conservation of forest and forest resources under the present and future environmental restrictions? There must be political, technical, and scientific solutions, all of which must be integrated, for obtaining sustainable management, conservation, and rational exploitation. In the 21st century, biotechnology is taking a pivotal role for facing this challenge and finding solutions. Biotechnology is defined as the use of techniques based on the knowledge of living beings, especially at the molecular level. There are different strategies, including classical genetic improvement programs, genetic engineering, searches for agrochemicals, and the exploitation of biodiversity. The latter strategy is considered to be the feasible alternative at short term in the current state of knowledge that can be used with Quercus, as the genus comprises non-domesticated and recalcitrance species with a long biological cycle. In this direction, a main objective is to characterize the phenotypic diversity of the species in terms of growth characteristics, productivity, and stress tolerance, among others, as a step prior to the selection of elite or plus genotypes that can be used in reforestation, conservation, and breeding programs.
This manuscript intends to be a review about the use of classical biochemistry and modern omics techniques (genomics, transcriptomics, proteomics, and metabolomics) employed so far to study the response of Quercus spp. to biotic and abiotic stresses (Table 1), and to understand how the genotype plus environment and epigenetics contribute to tolerant or resistant phenotypes. The genetic and epigenetic factors of healthy or diseased individuals remain poorly investigated and are thus almost unknown in Quercus spp. The current selection of elite trees in reforestation, conservation, and breeding programs requires the use of molecular analysis related to growth and productivity, as well as to responses to environmental factors. Plants genetic variability can be analyzed following the biological information according to the Central Dogma of Molecular Biology and the omic cascade in eukaryotes, as can be observed in Q. ilex (Figure 1). Central Dogma was enunciated by Crick [26] and define the relation between DNA, RNA and proteins, but nowadays different disciplines are interpreted their own “Central Dogma” including the relation with the omics cascade [27,28,29,30] to the final output, the phenotype. This information can be determined at different hierarchical levels of organization from individuals to populations and species by using different approximations and parameters, such as classical biochemistry (isozymes and secondary metabolites), and molecular biology, including the modern omics techniques (genomics, transcriptomics, proteomics, and metabolomics), together with DNA-based markers and epigenetic techniques.

Table 1.
Molecular stress studies carried out on the genus Quercus in the last 5 years. This table also includes those studies carried out at genomic and proteomic levels that are not under stress conditions to indicate the current status in this genus.
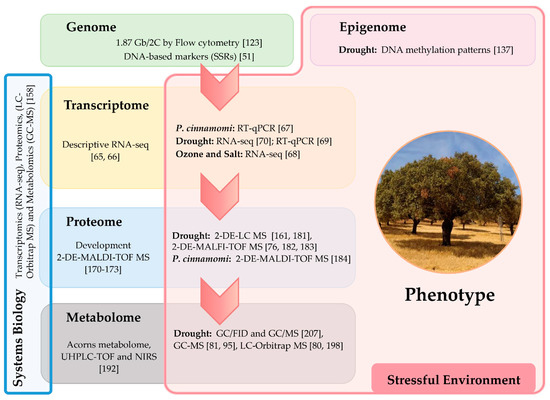
Figure 1.
Central Dogma of Molecular Biology and omic cascade in Quercus ilex to characterize the responses to biotic and abiotic stresses using molecular approaches. The studies analyzed under stress in the different omics are located within the red “stressful environment” box. Abbreviations: Simple Sequence Repeats, SSRs; RNA sequencing, RNA-seq; Reverse Transcription-quantitative PCR, RT-qPCR; 2-dimensional gel electrophoresis, 2-DE; Matrix-Assisted Laser Desorption/Ionization, MALDI; time-of-flight, TOF; Mass Spectrometry, MS; Liquid Chromatography, LC; Ultra High-Performance Liquid Chromatography, UHPLC; near infrared spectroscopy, NIRS; Gas Chromatography, GC; and Flame Ionization Detector, FID.
2. Classical Biochemistry
A wide variety of biochemical studies have been conducted to study the mechanisms of the stress response in Quercus spp., with most of them being focused on abiotic stresses, especially drought. Stress affects many cellular processes, such as photosynthesis, signaling, primary and secondary metabolism, responses to oxidative damage, and osmoprotection.
Stressful conditions produce changes in the photosynthetic apparatus. The analysis of the pigment content, accompanied by photosynthetic measures, has been extensively researched in species of the genus Quercus. These analyses are useful for inferring possible degradation by oxidative damage (with chlorophylls being one of the targets), as well as photoprotection via xanthophyll cycle pigments. The photoprotective mechanisms involved in responses to abiotic stresses, such as drought and cold, have been reviewed for evergreen (e.g., Q. suber, Quercus coccifera, and Q. ilex) and deciduous (e.g., Q. robur and Quercus pubescens) Quercus spp. [34]. Both deciduous and evergreen Quercus species occurring in contrasting climates implement different photoprotective mechanisms under drought stress, in which energy dissipation via xanthophyll cycles plays an important role. Studies on Q. pubescens [91] and Quercus brantii [31] reported a decrease in the pigment content (Chla, Chlb, and carotenoids) suggesting impairment of the light-harvesting complex and a decline in the dissipation processes of thermal energy. Ramírez-Valiente et al. [92] studied the photoprotective mechanisms in four live oak species (Quercus series Virentes) in response to different climatic conditions (drought and low temperatures). By studying pigment accumulation, the authors showed that Quercus fusiformis and Quercus oleoides increase the xanthophyll de-epoxidation state under water stress conditions when photoprotection is needed, while anthocyanins are used under cold stress in species with a limited freezing tolerance, such as Quercus virginiana and Quercus geminate. The performance of the photosynthetic machinery in response to an intense summer stress period was studied in three Mediterranean oaks (Q. ilex, Q. coccifera, and Q. suber). The photo-protection mechanisms were different in the three species. Only Q. suber exhibited a loss of chlorophyll, while the photosynthetic pigment composition remained unchanged in Q. coccifera and Q. ilex, despite interconversions within the xanthophyll cycle [93]. Similar results were reported by San-Eufrasio et al. [38] in Q. ilex under severe drought stress, with no significant changes in the photosynthetic pigment content suggesting little or no damage to the photosynthetic apparatus. Both Pb and Cd contamination have also been reported to increase the pigment content in Q. ilex to counteract the loss of photosynthetic surface area due to particle deposits in these contaminated environments [35].
The oxidative damage produced by stress has been studied in Quercus spp. by classical biochemical techniques such as reactive oxygen species (ROS) content analysis (e.g., H2O2 and O2−), lipid peroxidation (using the malondialdehyde (MDA) content), and the activity of antioxidant enzymes (superoxide dismutase, SOD; peroxidases, POD; catalase, CAT; ascorbate peroxidase, APX; glutathione reductase, GR; polyphenol oxidase activity, PPO; dehydroascorbate reductase, DR; and monodehydroascorbate reductase, MR). In Quercus spp., increased levels of ROS (mainly O2• or H2O2) have been observed in response to abiotic (drought, O3, pollutants (Pb and Cd), and heat) and biotic (as charcoal disease caused by the fungal pathogens Biscogniauxia mediterranea and Obolarina persica) stresses [31,33,39,91,94], being more intense when these stresses are combined (drought and charcoal disease) [39,91]. However, a reduction in ROS in stresses with enhanced ROS scavenging such as pCO2 has also been observed [40]. ROS cause the autocatalytic peroxidation of membrane lipids, which can be determined by the MDA content. Increased levels of superoxide radicals, hydrogen peroxide, and MDA were observed in several Quercus species in response to drought [31], drought and heat [91], drought and O3 [32,33], and drought and charcoal disease (which causes the drying of parts of the canopy and ultimately leads to local or systemic necrosis of the tree) [39]. The antioxidant enzymes SOD and POD increased levels in Q. brantii in response to drought [31], Q. pubescens in response to drought and heat [91], and Quercus infectoria and Quercus libani in response to drought and charcoal disease [39]. Under severe drought stress, Q. ilex showed increased levels of APX, GR, and CAT [31,95], with the latter also being increased in response to pollutants [35].
The content of sugars and amino acids, in primary metabolism, and the phenolic content, as well as the key enzyme activity involved in this metabolic pathway, in secondary metabolism, have been widely analysed in this genus. Under drought conditions, the content of sugars and amino acids (mainly proline) was increased in Q. brantii, improving the osmoprotection and preventing water loss [31]. The combination of this abiotic stress with charcoal disease in Q. libani and Q. infectoria, or O3 in Quercus cerris, also increased the content of these biomolecules [39]. Nevertheless, the content of sugars and amino acids was not altered in Q. ilex and Q. suber [38]. Secondary metabolism is normally stimulated under stress in the genus Quercus, with an increase in the total phenolic and flavonoid content or phenylalanine ammonia lyase (PAL) activity under drought or both drought and charcoal disease [31,38,39,95]. However, Q. suber did not show an increase of these compounds under drought or UV conditions [41].
A signaling process analysis by phytohormones has also been performed in this genus. An increase in the content of abscisic acid (ABA), considered a key hormone that mediates plant responses to adverse environmental stimuli, has been described for the genus Quercus under drought conditions, causing stomatal closure or premature leaf senescence [36,37,96]. The content of ABA, salicylic acid (SA), jasmonic acid (JA), and ethylene (ET) was determined in Q. ilex in response to drought and O3, with all of them reaching high concentrations at the initial stage of O3 treatment, demonstrating a spatial and functional correlation between these compounds [36]. However, the JA levels reached higher values when drought and O3 treatments were combined.
Many biochemical studies have been conducted on Quercus in response to abiotic stress, most of them aimed at studying changes in the photosynthetic apparatus and oxi-dative damage on leaves. Global biochemical analysis has been used to make inferences on biological aspects, but it is currently complemented by the application of specific techniques for the identification of compounds, as well as the massive analysis of proteins and metabolites. Studies that use these techniques beyond the global vision have been detailed in the omics layers in this review.
3. DNA-Based Markers
Following the Central Dogma of Molecular Biology, the first molecular layer is the DNA. DNA-based markers are widely used to estimate genetic diversity and germplasm identification. Initially, isozyme markers contributed to tree breeding programs [97]. However, currently, the use of DNA-based markers has achieved a high impact in forest tree programs. These markers can be grouped into hybridization-based DNA markers (RFLP), PCR-based DNA (e.g., sequence-characterized amplified regions (SCAR), amplified fragment length polymorphism (AFLP), random amplified polimorphic DNA (RAPD), simple-sequence repeats (SRR), and inter simple sequence repeats (ISSRs)), and DNA chip and sequencing-based DNA markers (e.g., single nucleotide polymorphism (SNP)) [98]. To characterize the response to stresses in the genus Quercus, investigations have mainly been performed with SSRs and SNPs in leaf tissue (reviewed in Müller and Gailing [99]). The drought tolerance has been widely studied with SSR markers in the genus Quercus [56,59,60,100]. Nagamitsu et al. [50] used nuclear SSRs in Quercus dentata and Quercus mongolica var. crispula to determine their adaptability to different environmental conditions in two different ecotypes, either oak coastal or inland forests in northern Japan, reporting a positive correlation between these DNA markers and environmental conditions. For Q. ilex, there is a study in which different Andalusian regions were determined by using a set of SSR markers (both nuclear and chloroplast) [51]. The eastern Q. ilex populations showed a higher level of drought tolerance than the western ones [101]. For Quercus aquifolioides, patterns of 381 SNPs from 65 candidate genes were analysed in relation to adaptation to climate change were analyzed [102]. Out of these SNPs, twelve and two genes were shown to be associated with drought and temperature, respectively. Candidate genes associated with environmental adaptation have also been reported in Q. suber, Quercus lobata, Q. oleoides, Q. petraea, and Q. robur by using SNPs [52,54,55,57,58,61,103]. For example, SNP 158 and SNP 168 located on a trehalase and peroxidase, respectively, play a role in drought stress [61]. Further, SNP markers have been used to position and localize putative candidate genes for drought tolerance on two genetic linkage maps of Q. robur and Q. petraea, with these being the first mapped genes on linkage maps of Quercus spp. [104].
Additionally, ISSR markers were used to determine the level of tolerance to oak dieback phenomena in Q. brantii [105]. These DNA-based markers provided useful information in the screening of oak seedlings to be used correctly in the restoration of damaged stands. A total of seven ISSR markers were associated with dieback phenomena. Metal toxicity, which is considered to be a major abiotic stressor, has been analysed in two-generation Quercus rubra populations using the RAPD marker system [106]. It was determined that the amount of bioavailable metal did not modify the genetic variation in Q. rubra, suggesting that two generations cannot be enough to cause change in the total DNA. The identification of quantitative trait loci (QTL) for tolerance-related traits is a useful tool for a future search of putative candidate genes for biotic and abiotic tolerance [107,108,109]. In Q. robur clones, five regions containing QTL specific to hypoxia responses were defined, which suggested waterlogging responses from a genetic point of view [108].
The potential of DNA-based markers is far from being exploited in the genus Quercus. So far, the majority of studies developed in these forest species have been related to drought; however, the large battery of SSRs and SNPs already described as well as new batteries of these markers might give relevant information in response to other stresses [110,111].
4. Genomics
The genome is the complete set of genetic information within a single cell of an organism. Genomics focuses on the study and cataloguing of all the genes that an organism possesses, as well as their organization, structure, and function. Research on forest tree genomics has fallen behind that of model and crop systems. However, genomic research on forest trees has progressed significantly with the advent of next-generation sequencing technologies (NGS). Until the sequencing of the genome of black cottonwood (Populus trichocarpa), which was selected for its small genome size, no reference genome sequence was available in forest species [112]. Thereafter, the list of forest tree genomes has been increased with the sequencing of Picea abies [113], Picea glauca [114], Eucalyptus grandis [115], and Pinus tadea [116], producing more than 50 genomes of tree species that are available in the NCBI Genome database [117,118]. In the genus Quercus, three species have been sequenced (Q. robur [46,47], Q. lobata [45], and Q. suber [48]). The oak genome size was initially estimated by Feulgen cytophotometry [119] and then, by flow cytometry [120,121,122]. Focusing only on those oak species already sequenced, Q. robur and Q. suber displayed a 2C nuclear DNA of 1.88 and 1.91 pg with a 38.9 and 39.7 GC% content, respectively. Information on the genome of Q. lobata was published at later [45], being currently the version 3.0 of valley oak genome available at https://valleyoak.ucla.edu/genomic-resources/ (accessed on 1 March 2021). In addition, all Quercus spp. are formed of 12 pairs of chromosomes (2n = 2x = 24). By using NGS, the genome size of Q. robur, Q. lobata, and Q. suber was determined to be 1.45 Gb/2C, 1.15 Gb/2C, and 1.90 Gb/2C, respectively [45,46,47,48]. Progress on the Q. ilex genome is ongoing, datasets have been generated by the single-molecule real-time (SMRT) technology called PacBio. The genome datasets have been submitted to NCBI’s BioProject repository (ID: 687489), but they are not yet public. The estimated genome size of Q. ilex, by flow cytometry, was approximately 1860 Mb/2C with a total length of 1.87 Gb/2C and, as expected, it is formed by 12 pairs of chromosomes [120,123,124,125].
The relevance of forest species genome sequencing is related to the huge genetic diversity observed to identify key genes and alleles controlling a large number of important traits. Much effort has been devoted to elucidating genomic backgrounds to enhance the response to biotic and abiotic stresses. In Gene Ontology (GO) term enrichment analysis in Q. robur, gene products involved in “response to biotic stimulus (GO:0009607)” and “defense response (GO:0006952)” were included in the best supported GO terms [47].
The chloroplast and mitochondrial genomes have been sequenced and annotated in the genus Quercus [42,43,44,49,126,127,128]. The whole mitochondrial genome has been only sequenced in Quercus variabilis [49]. The chloroplast genome has been described in several Quercus species (Table 1). For example, a study published in 2016 compared the chloroplast genomes of five Quercus spp. from China (Quercus baronii, Quercus dolicholepis, Q. variabilis, Quercus aliena, and Q. aliena var. acuteserrata) [42]. After analyzing nucleotide substitutions, indels, and repeats, a total of 19 variable regions were identified that could provide plastid markers for taxonomic, phylogenetic, and stress studies within this genus [42].
The availability of the sequenced genomes facilities the access to essential information about genes and gene products with their functions, transcript levels, cis-acting regulatory elements and alternative splicing patterns [129], as well as characterize novel phenotypes using a combination of whole-genome resequencing, linkage maps and microarrays shedding some light to the gene expression changes and introducing new SNPs related to interest traits [130].
5. Epigenomics
Epigenetics is the study of all processes (heritable or not) that modify gene expression or activity of transposable elements without modifying the DNA sequence [131]. Epigenetics is characterized as a fundamental mechanism that contributes to phenotypic variation (gene regulation, development, the stress response, phenotypic diversity, and evolution). Epigenetic modifications (DNA methylation, histone modifications, and microRNA regulation) play an important role in the expression of stress-induced genes, contributing to a plant’s potential to adapt to stresses [132,133]. DNA methylation has been widely assessed in plants by the silencing of gene expression [134]. Cytosine methylation, common form of post-replicative DNA modification seen in both bacteria and eukaryotes, occurs at CG, CHG and CHH sites, where H can be A, T, or C [134,135]. Moreover, other epigenetic mechanisms are considered in other layers of the central dogma such as histone modification in proteomics and RNA methylation in transcriptomics [29], although they have been less studied. Epigenetic variation contributes the phenotypic plasticity, adaptative capacity, and ability to persist in variable environments, which could be relevant for long-lived organisms with complex life cycles, such as forest trees. The epigenetic variation analysis of trees has gained much interest in recent years, recently reviewed in Amaral et al. [131]; however, few studies have investigated epigenetic variations related to climate adaptation in forest trees, in general, and in the genus Quercus, in particular. A high-quality genome sequence assembly in the sequencing of forest tree genomes may contribute to an increase in forest epigenetic studies. To date, the effect of biotic and abiotic stress at an epigenetic level has been studied in few Quercus spp., so the epigenetic changes that may occur under stress conditions remain largely unknown in this genus. Correia et al. [136] stated that epigenetic mechanisms such as DNA methylation and histone H3 acetylation are essential for the acclimation and survival of 8-month-old Q. suber plants at high temperature. In Q. ilex, Rico et al. [137] reported changes in the DNA methylation patterns in adult individuals exposed to drought for 12 years, indicating that these changes are associated with drought stress acclimation. DNA methylation variations associated with climate gradients have also been described in Q. lobata mature leaves and seedlings [53,62,138]. Browne et al. [62] reported a reduced in the genome-wide methylation induced by 5-Azacytidine in 5-month-old seedlings of the tree, Q. lobata, suggesting that removal of DNA methylation could affect plant adaptation to environmental change. As a first approach to inherited epigenetic analysis in Quercus, we have evaluated the global DNA methylation patterns in two individuals of Q. ilex located in Aldea de Cuenca (Córdoba, Andalusia, Spain) using the methylation sensitive amplification polymorphism (MSAP) technique, which is one of the most used methods for assessing DNA methylation changes in plants [139]. Different methylation patterns have preliminary been found between individuals and developmental stages (adult trees and seedlings). Our current results have also unveiled specific DNA methylation differences that could correspond to relatively stable changes and be involved in the development of epigenetic memory of the stress.
Recently, the existence of epigenetic priming, or individual stress memory triggering natural plant defenses [140,141], has been described in forest trees [133,142]. Initially, this term was associated with biotic stress (e.g., immunity against pathogens), but this phenomenon has also now been related to abiotic stress [143,144]. Most studies carried out to determine the epigenetic priming have been developed for the genera Pinus, Picea, and Populus [142,145,146,147]. For the genus Quercus, there is a lack of priming studies focused on biotic and abiotic stress from an epigenetic point of view. Recently, studies based on applying chemical elicitors, such as benzothiadiazole (BTH), have been applied to Q. ilex seeds and seedlings to induce epigenetic memory of defense responses (priming) that may increase resistance to abiotic (drought) or biotic (P. cinnamomi) future stress, but the results have not yet been published.
Currently, the interest by the tree epigenome is increasing with an emergent need to explore epigenetic diversity in response to stresses [131]. To date, the methodology applied to the genus Quercus provides overall information about global DNA methylation patterns associated to high temperature and drought. However, it is required a step forward in the epigenome knowledge with the identification of genes encoding epigenetic regulators (e.g., DNA methylases) and stress-induced changes in the expression of these genes to explain the contribution of epigenetic modifications to tree plasticity [148].
6. Transcriptomics
The second molecular layer within the Central Dogma of Molecular Biology in eukaryotes is the RNA. The transcriptome is the set of all RNA molecules, including messenger RNA (mRNA), ribosomal RNA (rRNA), transfer RNA (tRNA), small nuclear RNAs (snRNAs), small nucleolar RNAs (snoRNAs), and other noncoding RNA (ncRNA), produced in one cell or a population of cells [149]. The transcriptome, unlike the genome, is modifiable under different conditions (such as development stage, tissue, and environmental changes), being a promising layer for exploring the stress response. The techniques used to determine the gene expression are grouped into targeted (reverse transcription-quantitative PCR (RT-qPCR) and microarray) and untargeted (RNA sequencing (RNA-seq)) transcriptomic studies.
A targeted transcriptomic study is a useful tool for studying processes of interest, being of great utility, even after the arrival of NGS technologies. In Quercus spp., targeted transcriptomics studies started with RT-qPCR or microarrays (as used by Spieß et al. [150] in Q. robur under drought). There are recent studies of RT-qPCR in important processes such as cork formation in Q. suber under drought (e.g., the study of gene QsMYB1 by Almeida et al. [151]), and the different expression of aquaporins in short-term waterlogging in Q. petrea and Q. robur (tolerant) [152] or under biotic stress produced by P. cinnamomi in the expression of defense genes in Q. suber [153] or tannin synthesis genes in Q. ilex [67]. In these studies, it is important to highlight the analysis of the alternative splicing produced by stress, showing how allelic gene variants play a fundamental role in the response to stress. Almeida et al. [151] observed the preference for the QsMYB1.1 variant upon heat stress, while in drought, QsMYB1.2 was the predominant one. In the same sense, Gallardo et al. [67] observed how variant 2 of shikimate dehydrogenase (SDH2) was more active under mechanical defoliation.
NGS was developed to study the genome, allowing the analysis of RNA through the sequencing of complementary DNA (cDNA) [154]. This non-targeted method, called RNA-Seq, has clear advantages over previous approaches (such as microarrays) and has revolutionized our understanding of the complex and dynamic nature of the transcriptome [149]. For these reasons, this review is focused on them. For the genus Quercus, in recent years, thanks to the affordability of these analyses, numerous studies of non-targeted transcriptomic analysis have been conducted. RNA-seq analyses of the genus Quercus were initiated in 2010 in Q. petrea and Q. robur [155], using buds, leaves, and wood. In these analyses, a high enrichment in the molecular function “response to stress” was observed, being the second one with the highest number of transcripts (10.8%), showing a similar percentage to that in the analysis of the Q. robur DF159 clone [156]. The Q. ilex transcriptome [65,66] (analyzed in a mix of embryos, leaves, and roots) also exhibited an enrichment in “response to stress”, being the gene ontology category with the highest number of transcripts (5405 transcripts) in the Biological Processes. However, a transcriptomic analysis of more than 21 types of tissue (covering genes expressed in multiple tissues, developmental stages, and stress conditions) carried out in Q. suber showed a low enrichment of this biological process, being below 3% of those transcripts in “response to stress” [157]. These descriptive transcriptomic studies are of great help for the identification and creation of specific protein databases for Quercus non-sequenced species [158,159], achieving better protein identification than using generalist databases.
Comparative non-targeted transcriptomics analysis under stress allows the global transcriptomics response to be deepened, with numerous examples within the genus Quercus. Le-provost et al. [73] studied in seedlings the response to flooding in Q. petraea and Q. robur, and showed that Q. robur was more tolerant due to the activation of suberin biosynthetic, ABA, and ethylene pathways related to the formation of adaptive structures (hypertrophied lenticels and adventitious roots), and the rapid upregulation of the fermentation pathway, which helps to maintain energy production and overcome hypoxia in this tolerant species [73]. Under drought, a non-targeted analysis on leaf of different Q. lobata populations demonstrated how seedlings from different locations respond to different environmental conditions [71,72]. Under drought, the populations overexpressed the same pathways (e.g., abiotic stress or senescence processes) [71], although the authors observed how some gene expression, such as heat shock protein (HSP) 2-like chaperones (related to memory to stress) or metabolism-related proteins (e.g., phosphoglycerate kinase), were population-dependent. In the same line, Mead et al. [72] observed differences between populations adapted to warmer climatic conditions under drought, where gene expression for ribosomal proteins and for chloroplast and mitochondrial structures were differentially increased, while gene expression for DNA replication were differentially decreased between populations. This may be due to the fact that the response pathways are different among populations adapted to combined stress than those that are usually only under one stress (drought) [72]. Another application in transcriptional study is to analyze adaptive introgression using genes associated with environmental factors. In this sense, Oney-Birol et al. [64] analyzed the hybridization and introgression in adult trees of Quercus engelmannii, Quercus berberidifolia, and Quercus cornelius-mulleri using allelic variables in drought genes. Out of 139 drought candidate genes, eleven were selected as possible drought-associated species-specific adaptation genes, such as cellulose synthase and HSP 70kDa protein 10. Within European oaks, the transcriptomic response in drought has also been studied in the seedling roots of Q. suber [75] and seedling leafs of Q. robur, Q. pubescences, and Q. ilex (from less to more tolerant) [70]. Magalhães et al. [75] observed how the activation of the signaling response mediated by ABA- and ABF-dependent genes was of key importance under drought for Q. suber. Furthermore, Madritsch et al. [70] confirmed in 9 year-old plants that, under drought, there are species-specific responses to drought. In Q. robur, the important pathways were the ROS scavenging machinery, ABA-mediated gene expression, and cellular osmotic adjustment, as well as the initiation of the catabolic pathway, possibly due to high stress with the onset of senescence. In Q. pubescens, processes such as ROS scavenging or minor catabolic processes with a decrease in photosynthesis were detected, but heat-shock proteins and lignification pathways were expressed. The more tolerant Q. ilex also presented overexpression of the ROS pathway, although it differentially overexpressed genes relating to a new cell wall remodeling pathway [70]. The exposure to O3 (alone or in combination with other stresses) has also been reported in the Quercus spp. Soltani et al. [74] analyzed the exposure to three doses of O3 (150 ppb, 225 ppb, and 300 ppb) in Q. rubra seedlings. Exposure to the highest concentrations resulted in the activation of defense gene expression by altering the metabolic pathways of carbohydrates, amino acids, lipids, and terpenoids (such as the preferred biosynthesis for mono-, di-, and tetraterpenes under stress), as well as in modifying key processes, such as photosynthesis and the ATP production pathway. Furthermore, oxidative stress caused by O3 produced an increase in ROS which the plant faces with the increase of detoxification pathways, as seen in the increase in glutathione pathway overexpression [74]. In Q. ilex seedlings, Natali et al. [68] studied the combined effect of salt and O3, as well as in individual cases, showing unique responses in terms of the expression in single or combined stress situations with treatment-specific expressed transcripts (such as sucrose-phosphate synthase only being overexpressed in combined stress). The transcriptomic data showed common stress molecular responses (such as ROS scavenging or signaling by phytohormones), but treatment-specific differences in expression in key pathways, such as photosynthesis, cell wall remodeling, and sugar and lipid metabolism. In conclusion, the impact of saline stress was more severe than with O3 alone, but in combination, it had a strong effect on gene overexpression and suppression. Finally, from the RNA-seq analysis, it is possible to observe the selection of genes that do not vary their expression throughout the stress. These genes, considered as candidate reference genes, could improve normalization of RT-qPCR over classical reference genes. The comparative transcriptome analysis obtained among Q. robur, Q. pubescences, and Q. ilex [70] selected specific candidates (such as serine/threonine-protein phosphatase PP1 for Q. ilex) to be used for reference genes for gene expression normalization in a RT-qPCR analysis of each Quercus [69].
The affordability of RNA-seq has allowed a significant advance in the analysis of stress in the Quercus genus, being predominant the studies in drought. These results have usually been obtained in seedlings, with few examples in adult trees, but of great importance for the advancement of research. Moreover, the analysis of the splicing variables that occur under different stresses is a field that is beginning to be studied in this genus, where the progress to global non-targeted analysis is positioned as a promising field of study.
7. Proteomics
The transcriptome is translated into functional proteins that ultimately determine what happens in a living system. Proteomics intends to study the whole proteome or the sum of all proteins from an organism, tissue, and cell. Proteins undergo different changes (post-translational modifications, proteolysis, recycling, or multi-complex formation), which are key to the regulation of cellular processes [160]. As environmentally-sensitive layers, proteomic analysis is a powerful tool for the discovery of the key molecular mechanisms involved in stress. Proteomics has been scarcely used for research on forest species and even less so in the genus Quercus. Within this genus, to the best of our knowledge, holm oak (Q. ilex) is the most studied at the proteomic level and, to a lesser extent, other species, such as Q. suber and Q. rubra. Proteomics studies have been carried out in Q. ilex with a methodological optimization purpose [161,162,163,164,165], proteotyping or proteome descriptive characterization [158,166,167,168,169], and seed maturation and germination [170,171,172,173]. Proteomics studies in Q. suber have mainly addressed the cork quality [174,175] and somatic embryogenesis [176,177]. In addition, proteomics is emerging as a promising tool for identifying allergens in the pollen of Quercus spp. [178,179]. Most proteomic studies on the responses to stresses have been performed in Q. ilex, most being limited to the use of the gel-based (one or two-dimensional gel electrophoresis) coupled to MALDI-TOF MS (Matrix-Assisted Laser Desorption/Ionization time-of-flight mass spectrometry), and to a lesser extent, shotgun gel-free (LC-MSMS) platforms. All of the proteomics studies performed in Q. ilex have been reviewed in Rey et al. [123,180], with many research in abiotic and biotic stresses related to decline syndromes (drought and P. cinnamomi infestation). The first proteomics works in Q. ilex were focused on the variation of the leaf proteome between provenances in response to drought [161]. Further studies were addressed to characterize and select those genotypes with high levels of tolerance to drought [76,181,182,183] and resistance to P. cinnamomi [184]. Overall, differences in tolerance were related in Q. ilex at the provenance level, and a general tendency to decrease proteins of photosynthesis and ATP synthesis upon water withholding was observed [182]. In response to P. cinnamomi, two Q. ilex provenances were studied [184]. Physiological and proteomic analyses revealed differential responses of the provenances studied, with a tendency to decrease proteins of photosynthesis, amino acid metabolism, and stress-related proteins, mainly in the most susceptible one. However, some proteins related to starch biosynthesis, glycolysis, and stress-related peroxiredoxin were increased upon inoculation. The authors concluded that holm oak’s response to P. cinnamomi resembles the drought-stress response [184].
Besides the aforementioned works carried out in Q. ilex, only one proteomic work has been performed for Q. robur to study the effect of one season of drought stress, simulating the conditions of a dry summer for young oak trees [185]. Phenotypical and proteomics (Two-Dimensional Difference Gel Electrophoresis, 2D-DIGE) results revealed that plants were initially able to cope with the stress, but prolonged drought periods produced an accumulation of osmoactive compounds (such as mono- and di-saccharides) in the foliar tissue, adjusting their metabolism for some time, with the cost of reducing growth and productivity. It is also worth mentioning in this section the importance of ectomycorrhizal symbiosis in forest species. The protein profile of Q. suber in response to ectomycorrhizae formation was studied using proteomics and biochemical approaches [77]. The comparative analysis of mycorrhizal and nonmycorrhizal plants revealed no differences at the foliar level. However, an increased allocation of carbohydrates from the plant to the fungus to sustain the symbiosis was observed in roots. In addition, protein unfolding, the attenuation of defense reactions, increased nutrient mobilization from the plant–fungus interface (N and P), cytoskeleton rearrangements, and induction of plant cell wall loosening for fungal root accommodation in colonized roots were also suggested. These results corroborate the potential of mycorrhizal inoculation to improve the cork oak forest resistance capacity to cope with future climate change [77].
As a future prospect, proteomics has great potential in plant biological research in general, and in the genus Quercus in particular, which has been little exploited thus far. Most of proteomics studies have been carried out in Q. ilex, with stress studies being mostly limited to leaves using gel-based approach. Through the use of state-of-the-art proteomics techniques, together with a deep analysis conducted through subcellular fractionation, the study of post-translational modifications (PTMs), and target proteomic techniques (selected/multiple-reaction monitoring (SRM/MRM), parallel-reaction monitoring (PRM) or sequential window acquisition of all theoretical mass spectra (SWATH)), a range of possibilities will emerge that will allow for deeper knowledge on the molecular mechanisms underlying defense responses in the genus Quercus against biotic and abiotic stresses.
8. Metabolomics
Metabolomics can be considered as the omics layer the closest to the phenotype, being the last layer within the omic cascade. Metabolomics is also one of the most environmentally-sensitive layers, which allows a snapshot of any stress situation to be obtained [186,187,188,189]. Therefore, in the renewed search for biomarkers for stress tolerance, it is one of the most promising omics for the discovery of candidates [190,191] and useful for the identification of nutraceutical and bioactive acorn compounds [192], as possible food products for our diet in this genus. Metabolomics stress research was started with a classical biochemical analysis of total compounds within the different plant organisms, as has already been described above. These global biochemical analyses were completed with a targeted metabolomic analysis focused on the identification of compounds of interest [193].
The metabolomic analysis of phenolic compounds is a relevant field within the genus Quercus. These compounds have multiple protective functions against environmental stress [89], being important in the synthesis of the cell wall under temperature stress [194]. In recent years, within the phenolic compounds, target studies aimed at analyzing the production of tannins (condensed and hydrolysable) in different stress conditions. By using LC-MS, Top et al. [89] determined the molecular composition of condensed and hydrolysable tannins produced under a high temperature and drought in Q. rubra. In this study, plants grown under a single stress (high temperature or drought) produced more tannins per unit of leaf mass than those grown under favorable conditions, being higher for the combination of these stresses. Additionally, within this targeted study of the metabolome, volatile organic compounds (VOCs) emitted by oaks were also analyzed by using GS-MS. In trees, the emission of isoprenoids is widespread, with the genus Quercus being unique in evolved species emitting isoprene, monoterpene, or non-isoprenoids [195]. Oaks from the western hemisphere (Quercus alba, Q. lobata, and Q. rubra) and two from the eastern hemisphere (Q. robur, Q. pubescens) mainly emit isoprene, whereas some oaks from Mediterranean environments (Q. ilex, Q. coccifera, and Q. suber) emit monoterpenes [195,196]. The emission of isoprenoids helps protect against oxidative stress in plants, as protection against high temperatures [197] or in interaction with other organisms for different reasons (herbivory, pollination, or communication with other plants) [198]. The study of these emissions ranges from the investigation of seasonal emissions variations [87,199,200,201] to the change in emissions under stress conditions [202]. The emission of isoprenoids under drought stress within the genus Quercus is controversial [87,95]. In most studies reported, emissions under a slight stress were increased, but were reduced under extreme drought [87,203]. Despite this, studies developed in adults trees of Q. pubescens have identified an increase in isoprene emissions in situations of mild water stress [86,200], as well as a reduction in them [204,205]. Furthermore, conditions of a high temperature have a stimulating effect on the release of carbon from isoprene synthesis and mitochondrial respiration. However, this effect was attenuated under drought [82]. In adult trees of Q. ilex, the emission of terpenes has mainly been analyzed under drought using GS-MS [81,95]. The temperature is an incentive for the emission of terpenes in this species, although it is very sensitive to drought, only displaying an increase of particular terpene emissions at moderate stress, and a slight reduction in the total amount of terpenes emitted [95]. The increase occurred for the main terpenes, such as limonene [81,95] and α-pinene [81]. In prolonged drought stress, Q. suber adult trees reduced the total terpenoid emission, remaining increased in some specific terpenoids, such as γ-terpinene [83]. This may be due to its concrete importance in the adaptation to stress [83,206]. In this context, Q. suber trees under biotic stress by Cerambyx welensii modified the monoterpene emission profile, emitting more limonene as opposed to the non-stressed profile dominated by the pinene type [85]. The emissions in Q. suber do not greatly depend on leaf water potential or air temperature [83] as in Q. ilex [207]. A comparison of emissions in drought versus flooding has also been studied in Q. robur [208]. According to the authors, the total VOC emissions were decreased in drought and increased in flooding, although the isoprene emissions were not altered in both stresses. The VOC emission induction against herbivory has recently been reviewed for trees [209]. For example, Volf et al. [90] showed an increase in the emission of 45 VOCs, mainly terpenes (4 mono- and 21-sesquiterpenes), which produce a decrease in caterpillars’ appetite for methyl jasmonate-treated branches, as well as a possible attraction of predators in Q. robur.
In recent years, the advent of massive data analysis has allowed metabolomic analysis from a global perspective, which has permitted a more in-depth understanding of the metabolome in the presence of different stresses. Among the platforms used, LC-MS coupled to an LTQ Orbitrap XL is one of the most frequently used, permitting the analysis of a high number of compounds. Rivas-Ubach et al. [198] determined an increase in the flavonoid concentration under drought, as well as in the total content of soluble sugars, in Q. ilex adult trees. The accumulation of these compounds in drought may be due to their role as antioxidants (flavonoids) and in osmotic regulation (soluble sugars), making leaves more attractive to folivores [198]. Additionally, small-sized Q. ilex trees showed a greater carbohydrate accumulation and terpene concentration than larger trees in drought [79]. The response to drought in Q. ilex seedlings has also been studied at the level of root exudates, due to its impact on the plant, other neighboring plants, and soil properties, being mainly identified compounds of primary and secondary metabolism [80]. The root metabolism under drought was analyzed in Q. rubra and Q. alba seedlings and it was reported that the amount of lignins was not modified in the fine roots, although it did increase in others part of the plants as in leaves [78]. Therefore, adaptation to drought was facilitated in part by changes in the content and composition of tannins [78]. In Q. rubra trees, Susavella et al. [210] also analyzed the change in the metabolic production and resorption in leaves against single (high temperature or drought) or combined stresses. The metabolomic profile of the single stresses was not comparable to the combined one, showing an increase of the amino acid content in both, but with different key amino acids per treatment [210] (as spartic acid, methionine and lysine under combined stress). The combined stress of moderate drought and CO2 was studied in Quercus pyrenaica seedlings by Aranda et al. [88], who only observed significant differences in recovery. This study differs from those performed in Q. rubra, where the concentration of amino acids increased in drought [210]. This denotes the possible species-specific response within the genus Quercus.
VOCs emission is well studied in the genus under different stresses in adult trees, predominantly drought. However, in non-targeted metabolomics (more recently used), the majority of studies are in seedlings. The studies show a great variability in the emission or accumulation of metabolites within the genus under stress, showing the existence of key compounds specific for species and stress. This validates the metabolome as one of the most promising molecular layers in the search for markers of tolerance to different stresses in the Quercus genre.
9. Integration and Systems Biology
The current trend in molecular analysis is to go one step beyond the mere analysis of single and independent omics layers using a Systems Biology approach. System biology is the study of biological systems whose behavior cannot be reduced to the linear sum of the functions of their parts [211]. For this purpose, computational modelling of molecular systems and an integrative interpretation of the different omics layers (such as transcriptomics, proteomics or metabolomics) are used [212]. This approach can potentially provide a more complete picture of the stress response in plants by exposing the interconnection between the different molecular levels and provided a global vision of the stress. Moreover, it is possible to improve the robustness of the molecular analysis obtained under stress. The Systems Biology approach has been carried out through a global integration of the different omics layers, as well as with classical biochemistry, physiological, or phenotypic data. This approach has been little studied in the genus Quercus, as in general in trees, although a few studies have already been performed in other forest species, such as Pinus spp. [191,213,214,215]. The first contact with this approach in the genus Quercus was the integration of different omics layers (transcriptomics, proteomics, and metabolomics) for the most complete network reconstruction of metabolic pathways in Q. ilex spp. [158]. The Q. ilex metabolic network included carbohydrate and energy metabolism, amino acid metabolism, lipid metabolism, nucleotide metabolism, and the biosynthesis of secondary metabolites, with the tricarboxylic acid cycle being the pathway most represented. Under stress, Almeida et al. [84] analyzed the response to drought in Q. suber seedlings by means of an integrated analysis of the metabolome, together with physiological parameters, showing different metabolomic profiles under drought and post rehydration. This study highlights the fundamental role of secondary metabolism in the different processes (in short- and long-term drought stress and recovery), showing specific metabolites for each moment, and proposing key metabolites in the different parts of the processes.
10. Conclusions and Future Perspectives
The present review is, to our knowledge, together with Kremer et al. [216], the only manuscripts that deal with molecular studies developed in the genus Quercus. This kind of studies started with the development of an EST catalogue performed in Q. petraea and Q. robur to be used in ecosystem genomics studies twenty years ago [155]. It is remarkable that the first whole genome sequencing of a species included in the genus Quercus, Q. robur, has been carried out five year ago [46], in comparison to the first genome sequencing in plants, A. thaliana carried out 20 years ago in 2000 [217], or in forest trees, Populus trichocarpa in 2006 [112]. To date, massive analysis methodologies have been developed (genomics, epigenomics, transcriptomics, proteomics, and metabolomic) implemented with other information (such as classical biochemistry or physiological data) that have helped to understand key concepts such as stress in the biology of forest species [129,218].
The integration of research molecular techniques in the study of Quercus spp. will support the most advances in the direction of Systems Biology contributing to our molecular stress understanding as has been performed in other forest species such as poplar or pine [213,214,219]. The available information on this genus is disperse and fragmented (as in general for the majority of forest species [218]), so that coordinated and cooperative multidisciplinary research consortiums, including ecologists, ecophysiologists, geneticists, forestry engineers, and molecular biologist, should be pursued in the future to understand, monitor, and predict functional genetic diversity. The molecular interpretation in the genus Quercus, as in other non-model species, has been challenging due to the absence of reference genotypes. The high molecular variability observed as well as other methodological aspects that complicate experimentation (such as the complexity in obtaining clones, seed recalcitrance or its long life cycle [220]) have contributed to the lack of molecular information in this genus. So, there is an imperative need to build consensus reference populations for each species in all countries [221], as well as establishing defined phenotypes for comparison and integration of data.
Nevertheless, the molecular techniques used are starting to give answers to processes related to development, germination, and genes or gene products related to environmental resilience and productivity, as well as elucidating biosynthetic pathways and regulatory networks. In comparison to the development of molecular approaches in model plant species such as A. thaliana, Oryza sativa or P. trichocarpa, the genus Quercus is still in its infancy. Quercus stress studies have focused on simple experimental systems in seedlings analyzing the response in a single organ (leaves) under controlled conditions. This allows us to establish partial and fixed frames, from which it is difficult to extrapolate to natural environments. So, the combination of controlled conditions with outdoor experiments across environmental gradients (e.g., provenances of different environments) will give us a more accurate view of the responses [218] triggered in the face of multiple stresses. Multiple stresses providing a more real understanding of how they occur in nature (as occurs for oak decline syndrome) and how responses differ from those observed by the same stresses separately.
The availability of a higher number of sequenced genomes will allow characterizing and cataloguing the natural biodiversity, as well as developing molecular markers based on DNA and epigenetic marks to establish correlations between variability and the responses to stresses and resilience. In the most environmentally sensitive omics layers (transcriptomics, proteomics, and metabolomics), key putative genes can be identified, and individual genes can be functionally validated by using model organisms, such as Arabidopsis. Therefore, the use all the described molecular approaches will facilitate the identification of those elite genotypes that, after clonal propagation, can be used in reforestation, conservation, and improvement programs as well as to mitigate the effect of climate change.
Author Contributions
Conceptualization, M.E. and J.V.J.-N.; writing—original draft preparation, M.E., M.Á.C. and M.-D.R.; writing—review and editing, M.E., M.Á.C., J.V.J.-N. and M.-D.R.; supervision, J.V.J.-N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grant ENCINOMICS-2 PID2019-10908RB-100 from the Spanish Ministry of Science, Innovation and Universities. M.E., M.Á.C., and M.-D.R. are grateful for awards of Juan de la Cierva-Formación (FJCI-2017-31613), Ramón y Cajal (RYC-2017-23706), and Juan de la Cierva-Incorporación (IJC2018-035272-I) contracts by the Spanish Ministry of Science, Innovation and Universities, respectively.
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nixon, K. Infrageneric classification of Quercus (Fagaceae) and typification of sectional names. Ann. For. Sci. 1993, 50, 25s–34s. [Google Scholar] [CrossRef]
- Manos, P.; Doyle, J.; Nixon, K. Phylogeny, biogeography, and processes of molecular differentiation in Quercus subgenus Quercus (Fagaceae). Mol. Phylogenet. Evol. 1999, 12, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Correia, T.; Azeda, C. Public policies creating tensions in Montado management models: Insights from farmers’ representations. Land Use Policy 2017, 64, 76–82. [Google Scholar] [CrossRef][Green Version]
- Seale, D. Michael Williams: Deforesting the Earth: From prehistory to global crisis, an abridgment. J. Agric. Environ. Ethics 2011, 24, 673–686. [Google Scholar] [CrossRef]
- FAO; UNEP. The State of the World’s Forests 2020: Forests, Biodiversity and People; FAO: Rome, Italy, 2020. [Google Scholar]
- Grantham, H.S.; Duncan, A.; Evans, T.D.; Jones, K.R.; Beyer, H.L.; Schuster, R.; Walston, J.; Ray, J.C.; Robinson, J.G.; Callow, M.; et al. Anthropogenic modification of forests means only 40% of remaining forests have high ecosystem integrity. Nat. Commun. 2020, 11, 5978. [Google Scholar] [CrossRef]
- Wardlaw, T.; Mitchell, P.; Pinkard, L. Combined stresses in forests. In Combined Stresses in Plants; Mahalingam, R., Ed.; Springer: Cham, Switzerland, 2015; pp. 223–244. ISBN 978-3-319-07898-4. [Google Scholar]
- Denman, S.; Doonan, J.; Ransom-Jones, E.; Broberg, M.; Plummer, S.; Kirk, S.; Scarlett, K.; Griffiths, A.R.; Kaczmarek, M.; Forster, J.; et al. Microbiome and infectivity studies reveal complex polyspecies tree disease in Acute Oak Decline. ISME J. 2018, 12, 386–399. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.; Jeger, M.; Kirk, S.; Xu, X.; Denman, S. Spatial and temporal patterns in symptom expression within eight woodlands affected by Acute Oak Decline. For. Ecol. Manag. 2016, 360, 97–109. [Google Scholar] [CrossRef]
- Brown, N.; Vanguelova, E.; Parnell, S.; Broadmeadow, S.; Denman, S. Predisposition of forests to biotic disturbance: Predicting the distribution of Acute Oak Decline using environmental factors. For. Ecol. Manag. 2018, 407, 145–154. [Google Scholar] [CrossRef]
- Denman, S.; Webber, J. Oak declines: New definitions and new episodes in Britain. Q. J. For. 2009, 103, 285–290. [Google Scholar]
- Denman, S.; Brown, N.; Kirk, S.; Jeger, M.; Webber, J. A description of the symptoms of Acute Oak Decline in Britain and a comparative review on causes of similar disorders on oak in Europe. Forestry 2014, 87, 535–551. [Google Scholar] [CrossRef]
- Brady, C.; Arnold, D.; McDonald, J.; Denman, S. Taxonomy and identification of bacteria associated with acute oak decline. World J. Microbiol. Biotechnol. 2017, 33, 143. [Google Scholar] [CrossRef]
- Brown, N.; Jeger, M.; Kirk, S.; Williams, D.; Xu, X.; Pautasso, M.; Denman, S. Acute Oak Decline and Agrilus biguttatus: The Co-occurrence of stem bleeding and D-shaped emergence holes in Great Britain. Forests 2017, 8, 87. [Google Scholar] [CrossRef]
- Brasier, C. Phytophthora cinnamomi and oak decline in southern Europe. Environmental constraints including climate change. Ann. For. Sci. 1996, 53, 347–358. [Google Scholar] [CrossRef]
- Gentilesca, T.; Camarero, J.; Colangelo, M.; Nolè, A.; Ripullone, F. Drought-induced oak decline in the western Mediterranean region: An overview on current evidences, mechanisms and management options to improve forest resilience. iForest Biogeosci. For. 2017, 10, 796–806. [Google Scholar] [CrossRef]
- Ruiz-Gómez, F.; Pérez-de-Luque, A.; Sánchez-Cuesta, R.; Quero, J.; Cerrillo, R.M.N. Differences in the response to acute drought and Phytophthora cinnamomi rands infection in Quercus ilex L. seedlings. Forests 2018, 9, 634. [Google Scholar] [CrossRef]
- Ruiz-Gómez, F.; Pérez-de-Luque, A.; Navarro-Cerrillo, R. The involvement of Phytophthora root rot and drought stress in Holm oak Decline: From ecophysiology to microbiome influence. Curr. For. Rep. 2019, 5, 251–266. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Natalini, F.; Cardillo, E. Holm oak death is accelerated but not sudden and expresses drought legacies. Sci. Total Environ. 2021, 754, 141793. [Google Scholar] [CrossRef]
- Luque, J.; Parladé, J.; Péra, J. Pathogenicity of fungi isolated from Quercus suber in Catalonia (NE Spain). For. Pathol. 2000, 30, 247–263. [Google Scholar] [CrossRef]
- Romero, M.A.; Sánchez, J.E.; Jiménez, J.J.; Belbahri, L.; Trapero, A.; Lefort, F.; Sanchez, M.E. New Pythium taxa causing root rot on Mediterranean Quercus species in south-west Spain and Portugal. J. Phytopathol. 2007, 155, 289–295. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Collins, W.; Fry, M.M.; Yu, H.; Yu, H.; Fuglestvedt, J.; Shindell, D.; West, J. Global and regional temperature-change potentials for near-term climate forcers. Atmos. Chem. Phys. 2013, 13, 2471–2485. [Google Scholar] [CrossRef]
- Crick, F. Central dogma of molecular biology. Nature 1970, 227, 561–563. [Google Scholar] [CrossRef]
- Di Leo, A.; Claudino, W.; Colangiuli, D.; Bessi, S.; Pestrin, M.; Biganzoli, L. New strategies to identify molecular markers predicting chemotherapy activity and toxicity in breast cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2007, 18 (Suppl. S1), xii8–xii14. [Google Scholar] [CrossRef]
- Patti, G.J.; Yanes, O.; Siuzdak, G. Innovation: Metabolomics: The apogee of the omics trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269. [Google Scholar] [CrossRef]
- Fu, Y.; Dominissini, D.; Rechavi, G.; He, C. Gene expression regulation mediated through reversible m6A RNA methylation. Nat. Rev. Genet. 2014, 15, 293–306. [Google Scholar] [CrossRef]
- Srivastava, S. Emerging insights into the metabolic alterations in aging using metabolomics. Metabolites 2019, 9, 301. [Google Scholar] [CrossRef] [PubMed]
- Jafarnia, S.; Akbarinia, M.; Hosseinpour, B.; Sanavi, S.A.M.M.; Salami, S.A. Effect of drought stress on some growth, morphological, physiological, and biochemical parameters of two different populations of Quercus brantii. iForest Biogeosci. For. 2018, 11, 212–220. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Remorini, D.; Pellegrini, E.; Landi, M.; Massai, R.; Nali, C.; Guidi, L.; Lorenzini, G. Variations in physiological and biochemical traits of oak seedlings grown under drought and ozone stress. Physiol. Plant. 2016, 157, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Cotrozzi, L.; Remorini, D.; Pellegrini, E.; Guidi, L.; Lorenzini, G.; Massai, R.; Nali, C.; Landi, M. Cross-talk between physiological and metabolic adjustments adopted by Quercus cerris to mitigate the effects of severe drought and realistic future ozone concentrations. Forests 2017, 8, 148. [Google Scholar] [CrossRef]
- García-Plazaola, J.L.; Hernández, A.; Fernández-Marín, B.; Esteban, R.; Peguero-Pina, J.J.; Verhoeven, A.; Cavender-Bares, J. Photoprotective mechanisms in the Genus Quercus in response to winter cold and summer drought in oaks physiological ecology. In Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L. Tree Physiology; Gil-Pelegrín, E., Peguero-Pina, J., Sancho-Knapik, D., Eds.; Springer: Cham, Switzerland, 2017; Volume 7, pp. 361–391. ISBN 978-3-319-69098-8. [Google Scholar]
- Arena, C.; Santorufo, L.; Cataletto, P.R.; Memoli, V.; Scudiero, R.; Maisto, G. Eco-physiological and antioxidant responses of Holm oak (Quercus ilex L.) leaves to Cd and Pb. Water Air Soil Pollut. 2017, 228, 459. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Pellegrini, E.; Guidi, L.; Landi, M.; Lorenzini, G.; Massai, R.; Remorini, D.; Tonelli, M.; Trivellini, A.; Vernieri, P.; et al. Losing the warning signal: Drought compromises the cross-talk of signaling molecules in Quercus ilex exposed to ozone. Front. Plant Sci. 2017, 8, 1020. [Google Scholar] [CrossRef] [PubMed]
- Peguero-Pina, J.J.; Mendoza-Herrer, O.; Gil-Pelegrín, E.; Sancho-Knapik, D. Cavitation limits the recovery of gas exchange after severe drought stress in Holm Oak (Quercus ilex L.). Forest 2018, 9, 443. [Google Scholar] [CrossRef]
- San-Eufrasio, B.; Sánchez-Lucas, R.; López-Hidalgo, C.; Guerrero-Sanchez, V.M.; Castillejo, M.; Maldonado-Alconada, A.M.; Jorrín-Novo, J.V.; Rey, M.-D. Responses and differences in tolerance to water shortage under climatic dryness conditions in seedlings from Quercus spp. and Andalusian Q. ilex populations. Forests 2020, 11, 707. [Google Scholar] [CrossRef]
- Ghanbary, E.; Tabari Kouchaksaraei, M.; Zarafshar, M.; Bader, K.-F.M.; Mirabolfathy, M.; Ziaei, M. Differential physiological and biochemical responses of Quercus infectoria and Q. libani to drought and charcoal disease. Physiol. Plant. 2020, 168, 876–892. [Google Scholar] [CrossRef]
- Arab, L.; Seegmueller, S.; Kreuzwieser, J.; Eiblmeier, M.; Rennenberg, H. Atmospheric pCO(2) impacts leaf structural and physiological traits in Quercus petraea seedlings. Planta 2019, 249, 481–495. [Google Scholar] [CrossRef]
- Díaz-Guerra, L.; Llorens, L.; Bell, T.L.; Font, J.; González, J.A.; Verdaguer, D. Physiological, growth and root biochemical responses of Arbutus unedo and Quercus suber seedlings to UV radiation and water availability before and after aboveground biomass removal. Environ. Exp. Bot. 2019, 168, 103861. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, T.; Duan, D.; Yang, J.; Feng, L.; Zhao, G. Comparative analysis of the complete chloroplast genomes of five Quercus species. Front. Plant Sci. 2016, 7, 959. [Google Scholar] [CrossRef]
- Liu, X.; Chang, E.; Liu, J.; Huang, Y.; Wang, Y.; Yao, N.; Jiang, Z. Complete chloroplast genome sequence and phylogenetic analysis of Quercus bawanglingensis Huang, Li et Xing, a vulnerable oak tree in China. Forests 2019, 10, 587. [Google Scholar] [CrossRef]
- Pang, X.; Liu, H.; Wu, S.; Yuan, Y.; Li, H.; Dong, J.; Liu, Z.; An, C.; Su, Z.; Li, B. Species identification of oaks (Quercus L., Fagaceae) from gene to genome. Int. J. Mol. Sci. 2019, 20, 5940. [Google Scholar] [CrossRef]
- Sork, V.; Fitz-Gibbon, S.; Puiu, D.; Crepeau, M.; Gugger, P.F.; Sherman, R.M.; Stevens, K.; Langley, C.; Pellegrini, M.; Salzberg, S. First draft assembly and annotation of the genome of a California Endemic oak Quercus lobata Née (Fagaceae). G3 Genes Genomes Genet. 2016, 6, 3485–3495. [Google Scholar] [CrossRef]
- Plomion, C.; Aury, J.; Amselem, J.; Alaeitabar, T.; Barbe, V.; Belser, C.; Bergès, H.; Bodénès, C.; Boudet, N.; Boury, C.; et al. Decoding the oak genome: Public release of sequence data, assembly, annotation and publication strategies. Mol. Ecol. Resour. 2016, 16, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Plomion, C.; Aury, J.; Amselem, J.; Leroy, T.; Murat, F.; Duplessis, S.; Faye, S.; Francillonne, N.; Labadie, K.; Provost, G.L.; et al. Oak genome reveals facets of long lifespan. Nat. Plants 2018, 4, 440–452. [Google Scholar] [CrossRef]
- Ramos, A.M.; Usie, A.; Barbosa, P.; Barros, P.M.; Capote, T.; Chaves, I.; Simoes, F.; Abreu, I.; Carrasquinho, I.; Faro, C.; et al. The draft genome sequence of cork oak. Sci. Data 2018, 5, 180069. [Google Scholar] [CrossRef] [PubMed]
- Bi, Q.; Li, D.; Zhao, Y.; Wang, M.; Li, Y.; Liu, X.; Wang, L.; Yu, H. Complete mitochondrial genome of Quercus variabilis (Fagales, Fagaceae). Mitochondrial DNA Part B 2019, 4, 3927–3928. [Google Scholar] [CrossRef]
- Nagamitsu, T.; Shimizu, H.; Aizawa, M.; Nakanishi, A. An admixture of Quercus dentata in the coastal ecotype of Q. mongolica var. crispula in northern Hokkaido and genetic and environmental effects on their traits. J. Plant Res. 2019, 132, 211–222. [Google Scholar] [CrossRef]
- Fernandez i Martí, A.; Romero-Rodríguez, C.; Navarro-Cerrillo, R.; Abril, N.; Jorrín-Novo, J.; Dodd, R. Population genetic diversity of Quercus ilex subsp. ballota (Desf.) Samp. reveals divergence in recent and evolutionary migration rates in the Spanish dehesas. Forests 2018, 9, 337. [Google Scholar] [CrossRef]
- Sork, V.L.; Squire, K.; Gugger, P.F.; Steele, S.E.; Levy, E.D.; Eckert, A.J. Landscape genomic analysis of candidate genes for climate adaptation in a California endemic oak, Quercus lobata. Am. J. Bot. 2016, 103, 33–46. [Google Scholar] [CrossRef]
- Gugger, P.F.; Fitz-Gibbon, S.; PellEgrini, M.; Sork, V.L. Species-wide patterns of DNA methylation variation in Quercus lobata and their association with climate gradients. Mol. Ecol. 2016, 25, 1665–1680. [Google Scholar] [CrossRef]
- Browne, L.; Wright, J.W.; Fitz-Gibbon, S.; Gugger, P.F.; Sork, V.L. Adaptational lag to temperature in valley oak (Quercus lobata) can be mitigated by genome-informed assisted gene flow. Proc. Natl. Acad. Sci. USA 2019, 116, 25179–25185. [Google Scholar] [CrossRef]
- Meireles, J.E.; Beulke, A.; Borkowski, D.S.; Romero-Severson, J.; Cavender-Bares, J. Balancing selection maintains diversity in a cold tolerance gene in broadly distributed live oaks. Genome 2017, 60, 762–769. [Google Scholar] [CrossRef]
- Ramírez-Valiente, J.A.; Deacon, N.J.; Etterson, J.; Center, A.; Sparks, J.P.; Sparks, K.; Longwell, T.; Pilz, G.; Cavender-Bares, J. Natural selection and neutral evolutionary processes contribute to genetic divergence in leaf traits across a precipitation gradient in the tropical oak Quercus oleoides. Mol. Ecol. 2018, 27, 2176–2192. [Google Scholar] [CrossRef]
- Rellstab, C.; Zoller, S.; Walthert, L.; Lesur, I.; Pluess, A.R.; Graf, R.; Bodénès, C.; Sperisen, C.; Kremer, A.; Gugerli, F. Signatures of local adaptation in candidate genes of oaks (Quercus spp.) with respect to present and future climatic conditions. Mol. Ecol. 2016, 25, 5907–5924. [Google Scholar] [CrossRef]
- Truffaut, L.; Chancerel, E.; Ducousso, A.; Dupouey, J.L.; Badeau, V.; Ehrenmann, F.; Kremer, A. Fine-scale species distribution changes in a mixed oak stand over two successive generations. New Phytol. 2017, 215, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Lind-Riehl, J.F.; Gailing, O. Adaptive variation and introgression of a CONSTANS-like gene in North American red oaks. Forests 2017, 8, 3. [Google Scholar] [CrossRef]
- Khodwekar, S.; Gailing, O. Evidence for environment-dependent introgression of adaptive genes between two red oak species with different drought adaptations. Am. J. Bot. 2017, 104, 1088–1098. [Google Scholar] [CrossRef] [PubMed]
- Pina-Martins, F.; Baptista, J.; Pappas, G.J.; Paulo, O.S. New insights into adaptation and population structure of cork oak using genotyping by sequencing. Glob. Change Biol. 2019, 25, 337–350. [Google Scholar] [CrossRef]
- Browne, L.; Mead, A.; Horn, C.; Chang, K.; Celikkol, Z.A.; Henriquez, C.L.; Ma, F.; Beraut, E.; Meyer, R.S.; Sork, V.L. Experimental DNA demethylation associates with changes in growth and gene expression of oak tree seedlings. G3 Genes Genomes Genet. 2020, 10, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Inácio, V.; Barros, P.M.; Costa, A.; Roussado, C.; Gonçalves, E.; Costa, R.; Graça, J.; Oliveira, M.M.; Morais-Cecílio, L. Differential DNA methylation patterns are related to phellogen origin and quality of Quercus suber cork. PLoS ONE 2017, 12, e0169018. [Google Scholar] [CrossRef]
- Oney-Birol, S.; Fitz-Gibbon, S.; Chen, J.-M.; Gugger, P.F.; Sork, V.L. Assessment of shared alleles in drought-associated candidate genes among southern California white oak species (Quercus sect. Quercus). BMC Genet. 2018, 19, 88. [Google Scholar] [CrossRef]
- Guerrero-Sanchez, V.M.; Maldonado-Alconada, A.M.; Amil-Ruiz, F.; Verardi, A.; Jorrín-Novo, J.V.; Rey, M.-D. Ion Torrent and lllumina, two complementary RNA-seq platforms for constructing the Holm oak (Quercus ilex) transcriptome. PLoS ONE 2019, 14, e0210356. [Google Scholar] [CrossRef]
- Guerrero-Sanchez, V.M.; Maldonado-Alconada, A.M.; Amil-Ruiz, F.; Jorrin-Novo, J. V Holm oak (Quercus ilex) transcriptome. De novo sequencing and assembly analysis. Front. Mol. Biosci. 2017, 4, 70. [Google Scholar] [CrossRef]
- Gallardo, A.; Morcuende, D.; Solla, A.; Moreno, G.; Pulido, F.; Quesada, A. Regulation by biotic stress of tannins biosynthesis in Quercus ilex: Crosstalk between defoliation and Phytophthora cinnamomi infection. Physiol. Plant. 2019, 165, 319–329. [Google Scholar] [CrossRef]
- Natali, L.; Vangelisti, A.; Guidi, L.; Remorini, D.; Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pellegrini, E.; Trivellini, A.; Vernieri, P.; et al. How Quercus ilex L. saplings face combined salt and ozone stress: A transcriptome analysis. BMC Genom. 2018, 19, 872. [Google Scholar] [CrossRef]
- Kotrade, P.; Sehr, E.M.; Wischnitzki, E.; Brüggemann, W. Comparative transcriptomics-based selection of suitable reference genes for normalization of RT-qPCR experiments in drought-stressed leaves of three European Quercus species. Tree Genet. Genomes 2019, 15, 38. [Google Scholar] [CrossRef]
- Madritsch, S.; Wischnitzki, E.; Kotrade, P.; Ashoub, A.; Burg, A.; Fluch, S.; Brüggemann, W.; Sehr, E.M. Elucidating drought stress tolerance in European oaks through cross-species transcriptomics. G3 Genes Genomes Genet. 2019, 9, 3181–3199. [Google Scholar] [CrossRef]
- Gugger, P.F.; Peñaloza-Ramírez, J.M.; Wright, J.W.; Sork, V.L. Whole-transcriptome response to water stress in a California endemic oak, Quercus lobata. Tree Physiol. 2017, 37, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Mead, A.; Peñaloza Ramirez, J.; Bartlett, M.K.; Wright, J.W.; Sack, L.; Sork, V.L. Seedling response to water stress in valley oak (Quercus lobata) is shaped by different gene networks across populations. Mol. Ecol. 2019, 28, 5248–5264. [Google Scholar] [CrossRef] [PubMed]
- Le Provost, G.; Lesur, I.; Lalanne, C.; Da Silva, C.; Labadie, K.; Aury, J.M.; Leple, J.C.; Plomion, C. Implication of the suberin pathway in adaptation to waterlogging and hypertrophied lenticels formation in pedunculate oak (Quercus robur L.). Tree Physiol. 2016, 36, 1330–1342. [Google Scholar]
- Soltani, N.; Best, T.; Grace, D.; Nelms, C.; Shumaker, K.; Romero-Severson, J.; Moses, D.; Schuster, S.; Staton, M.; Carlson, J.; et al. Transcriptome profiles of Quercus rubra responding to increased O(3) stress. BMC Genom. 2020, 21, 160. [Google Scholar] [CrossRef]
- Magalhães, A.P.; Verde, N.; Reis, F.; Martins, I.; Costa, D.; Lino-Neto, T.; Castro, P.H.; Tavares, R.M.; Azevedo, H. RNA-seq and gene network analysis uncover activation of an ABA-dependent signalosome during the cork oak root response to drought. Front. Plant Sci. 2016, 6, 1195. [Google Scholar] [CrossRef]
- Simova-Stoilova, L.P.; López-Hidalgo, C.; Sanchez-Lucas, R.; Valero-Galvan, J.; Romero-Rodríguez, C.; Jorrin-Novo, J.V. Holm oak proteomic response to water limitation at seedling establishment stage reveals specific changes in different plant parts as well as interaction between roots and cotyledons. Plant Sci. 2018, 276, 1–13. [Google Scholar] [CrossRef]
- Sebastiana, M.; Martins, J.; Figueiredo, A.; Monteiro, F.; Sardans, J.; Peñuelas, J.; Silva, A.; Roepstorff, P.; Pais, M.; Coelho, A. Oak protein profile alterations upon root colonization by an ectomycorrhizal fungus. Mycorrhiza 2016, 27, 109–128. [Google Scholar] [CrossRef]
- Suseela, V.; Tharayil, N.; Orr, G.; Hu, D. Chemical plasticity in the fine root construct of Quercus spp. varies with root order and drought. New Phytol. 2020, 228, 1835–1851. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Ubach, A.; Barbeta, A.; Sardans, J.; Guenther, A.; Ogaya, R.; Oravec, M.; Urban, O.; Peñuelas, J. Topsoil depth substantially influences the responses to drought of the foliar metabolomes of Mediterranean forests. Perspect. Plant Ecol. Evol. Syst. 2016, 21, 41–54. [Google Scholar] [CrossRef]
- Gargallo-Garriga, A.; Preece, C.; Sardans, J.; Oravec, M.; Urban, O.; Peñuelas, J. Root exudate metabolomes change under drought and show limited capacity for recovery. Sci. Rep. 2018, 8, 12696. [Google Scholar] [CrossRef]
- Mu, Z.; Llusià, J.; Liu, D.; Ogaya, R.; Asensio, D.; Zhang, C.; Peñuelas, J. Seasonal and diurnal variations of plant isoprenoid emissions from two dominant species in Mediterranean shrubland and forest submitted to experimental drought. Atmos. Environ. 2018, 191, 105–115. [Google Scholar] [CrossRef]
- Rodríguez-Calcerrada, J.; Rodrigues, A.M.; Perdiguero, P.; António, C.; Atkin, O.K.; Li, M.; Collada, C.; Gil, L. A molecular approach to drought-induced reduction in leaf CO(2) exchange in drought-resistant Quercus ilex. Physiol. Pantarum 2018, 162, 394–408. [Google Scholar] [CrossRef] [PubMed]
- Haberstroh, S.; Kreuzwieser, J.; Lobo-do-Vale, R.; Caldeira, M.C.; Dubbert, M.; Werner, C. Terpenoid emissions of two Mediterranean woody species in response to drought stress. Front. Plant Sci. 2018, 9, 1071. [Google Scholar] [CrossRef]
- Almeida, T.; Pinto, G.; Correia, B.; Gonçalves, S.; Meijón, M.; Escandón, M. In-depth analysis of the Quercus suber metabolome under drought stress and recovery reveals potential key metabolic players. Plant Sci. 2020, 299, 110606. [Google Scholar] [CrossRef]
- Sánchez-Osorio, I.; López-Pantoja, G.; Tapias, R.; Pareja-Sánchez, E.; Domínguez, L. Monoterpene emission of Quercus suber L. highly infested by Cerambyx welensii Küster. Ann. For. Sci. 2019, 76, 98. [Google Scholar] [CrossRef]
- Saunier, A.; Ormeño, E.; Wortham, H.; Temime-Roussel, B.; Lecareux, C.; Boissard, C.; Fernandez, C. Chronic drought decreases anabolic and catabolic BVOC emissions of Quercus pubescens in a Mediterranean forest. Front. Plant Sci. 2017, 8, 71. [Google Scholar] [CrossRef]
- Genard-Zielinski, A.-C.; Boissard, C.; Ormeño, E.; Lathière, J.; Reiter, I.M.; Wortham, H.; Orts, J.-P.; Temime-Roussel, B.; Guenet, B.; Bartsch, S.; et al. Seasonal variations of Quercus pubescens isoprene emissions from an in natura forest under drought stress and sensitivity to future climate change in the Mediterranean area. Biogeosciences 2018, 15, 4711–4730. [Google Scholar] [CrossRef]
- Aranda, I.; Cadahía, E.; Fernández de Simón, B. Leaf ecophysiological and metabolic response in Quercus pyrenaica Willd seedlings to moderate drought under enriched CO2 atmosphere. J. Plant Physiol. 2020, 244, 153083. [Google Scholar] [CrossRef]
- Top, S.M.; Preston, C.M.; Dukes, J.S.; Tharayil, N. Climate influences the content and chemical composition of foliar tannins in green and senesced tissues of Quercus rubra. Front. Plant Sci. 2017, 8, 423. [Google Scholar] [CrossRef]
- Volf, M.; Weinhold, A.; Seifert, C.L.; Holicová, T.; Uthe, H.; Alander, E.; Richter, R.; Salminen, J.-P.; Wirth, C.; van Dam, N.M. Branch-localized induction promotes efficacy of volatile defences and herbivore predation in trees. J. Chem. Ecol. 2021, 47, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Contran, N.; Günthardt-Goerg, M.S.; Kuster, T.M.; Cerana, R.; Crosti, P.; Paoletti, E. Physiological and biochemical responses of Quercus pubescens to air warming and drought on acidic and calcareous soils. Plant Biol. 2013, 15 (Suppl. S1), 157–168. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Valiente, J.A.; Koehler, K.; Cavender-Bares, J. Climatic origins predict variation in photoprotective leaf pigments in response to drought and low temperatures in live oaks (Quercus series Virentes). Tree Physiol. 2015, 35, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Peguero-Pina, J.J.; Sancho-Knapik, D.; Morales, F.; Flexas, J.; Gil-Pelegrin, E. Differential photosynthetic performance and photoprotection mechanisms of three Mediterranean evergreen oaks under severe drought stress. Funct. Plant Biol. 2009, 36, 453–462. [Google Scholar] [CrossRef]
- Ebadzad, G.; Medeira, C.; Maia, I.; Martins, J.; Cravador, A. Induction of defence responses by cinnamomins against Phytophthora cinnamomi in Quercus suber and Quercus ilex subs. rotundifolia. Eur. J. Plant Pathol. 2015, 143, 705–723. [Google Scholar] [CrossRef]
- Nogués, I.; Llusià, J.; Ogaya, R.; Munné-Bosch, S.; Sardans, J.; Peñuelas, J.; Loreto, F. Physiological and antioxidant responses of Quercus ilex to drought in two different seasons. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2014, 148, 268–278. [Google Scholar]
- Loewenstein, N.J.; Pallardy, S.G. Influence of a drying cycle on post-drought xylem sap abscisic acid and stomatal responses in young temperate deciduous angiosperms. New Phytol. 2002, 156, 351–361. [Google Scholar] [CrossRef]
- Hamrick, J.; Godt, M.; Sherman-Broyles, S.L. Factors influencing levels of genetic diversity in woody plant species. New For. 1992, 6, 95–124. [Google Scholar] [CrossRef]
- Gupta, P.; Varshney, R.; Sharma, P.C.; Ramesh, B. Molecular markers and their applications in wheat breeding. Plant Breed. 1999, 118, 369–390. [Google Scholar] [CrossRef]
- Müller, M.; Gailing, O. Abiotic genetic adaptation in the Fagaceae. Plant Biol. 2019, 21, 783–795. [Google Scholar] [CrossRef]
- Homolka, A.; Schueler, S.; Burg, K.; Fluch, S.; Kremer, A. Insights into drought adaptation of two European oak species revealed by nucleotide diversity of candidate genes. Tree Genet. Genomes 2013, 9, 1179–1192. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.; Gómez, F.R.; Cabrera-Puerto, R.J.; Sánchez-Cuesta, R.; Rodríguez, G.; Pérez, J. Growth and physiological sapling responses of eleven Quercus ilex ecotypes under identical environmental conditions. For. Ecol. Manag. 2018, 58–69. [Google Scholar] [CrossRef]
- Du, F.; Wang, T.; Wang, Y.; Ueno, S.; de Lafontaine, G. Contrasted patterns of local adaptation to climate change across the range of an evergreen oak, Quercus aquifolioides. Evol. Appl. 2020, 13, 2377–2391. [Google Scholar] [CrossRef]
- Modesto, I.S.; Miguel, C.; Pina-Martins, F.; Glushkova, M.; Veloso, M.; Paulo, O.; Batista, D. Identifying signatures of natural selection in cork oak (Quercus suber L.) genes through SNP analysis. Tree Genet. Genomes 2014, 10, 1645–1660. [Google Scholar] [CrossRef]
- Porth, I.; Scotti-Saintagne, C.; Barreneche, T.; Kremer, A.; Burg, K. Linkage mapping of osmotic stress induced genes of oak. Tree Genet. Genomes 2005, 1, 31–40. [Google Scholar] [CrossRef]
- Shamari, A.-R.; Mehrabi, A.-A.; Maleki, A.; Rostami, A. Association analysis of tolerance to dieback phenomena and trunk form using ISSR markers in Quercus brantii. Cell. Mol. Biol. 2018, 64, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Makela, M.; Michael, P.; Thériault, G.; Nkongolo, K. High genetic variation among closely related red oak (Quercus rubra) populations in an ecosystem under metal stress: Analysis of gene regulation. Genes Genom. 2016, 38, 967–976. [Google Scholar] [CrossRef]
- Casasoli, M.; Derory, J.; Morera-Dutrey, C.; Brendel, O.; Porth, I.; Guehl, J.-M.; Villani, F.; Kremer, A. Comparison of quantitative trait loci for adaptive traits between oak and chestnut based on an expressed sequence tag consensus map. Genetics 2006, 172, 533–546. [Google Scholar] [CrossRef]
- Parelle, J.; Zapater, M.; Scotti-Saintagne, C.; Kremer, A.; Jolivet, Y.; Dreyer, E.; Brendel, O. Quantitative trait loci of tolerance to waterlogging in a European oak (Quercus robur L.): Physiological relevance and temporal effect patterns. Plant. Cell Environ. 2007, 30, 422–434. [Google Scholar] [CrossRef]
- Brendel, O.; Le Thiec, D.; Scotti, C.; Bodénès, C.; Kremer, A.; Guehl, J. Quantitative trait loci controlling water use efficiency and related traits in Quercus robur L. Tree Genet. Genomes 2008, 4, 263–278. [Google Scholar] [CrossRef]
- Wang, L.; Du, H.; Li, T.; Wuyun, T. De novo transcriptome sequencing and identification of genes related to salt stress in Eucommia ulmoides Oliver. Trees 2018, 32, 151–163. [Google Scholar] [CrossRef]
- Ranade, S.S.; Ganea, L.-S.; Razzak, A.M.; García Gil, M.R. Fungal infection increases the rate of somatic mutation in Scots pine (Pinus sylvestris L.). J. Hered. 2015, 106, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar]
- Nystedt, B.; Street, N.R.; Wetterbom, A.; Zuccolo, A.; Lin, Y.-C.; Scofield, D.G.; Vezzi, F.; Delhomme, N.; Giacomello, S.; Alexeyenko, A.; et al. The Norway spruce genome sequence and conifer genome evolution. Nature 2013, 497, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Birol, I.; Raymond, A.; Jackman, S.D.; Pleasance, S.; Coope, R.; Taylor, G.A.; Yuen, M.M.S.; Keeling, C.I.; Brand, D.; Vandervalk, B.P.; et al. Assembling the 20 Gb white spruce (Picea glauca) genome from whole-genome shotgun sequencing data. Bioinformatics 2013, 29, 1492–1497. [Google Scholar] [CrossRef]
- Myburg, A.; Grattapaglia, D.; Tuskan, G.; Hellsten, U.; Hayes, R.D.; Grimwood, J.; Jenkins, J.; Lindquist, E.; Tice, H.; Bauer, D.; et al. The genome of Eucalyptus grandis. Nature 2014, 510, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Neale, D.B.; Wegrzyn, J.L.; Stevens, K.A.; Zimin, A.V.; Puiu, D.; Crepeau, M.W.; Cardeno, C.; Koriabine, M.; Holtz-Morris, A.E.; Liechty, J.D.; et al. Decoding the massive genome of loblolly pine using haploid DNA and novel assembly strategies. Genome Biol. 2014, 15, R59. [Google Scholar] [CrossRef] [PubMed]
- Sow, M.D.; Allona, I.; Ambroise, C.; Conde, D.; Fichot, R.; Gribkova, S.; Jorge, V.; Le-Provost, G.; Pâques, L.; Plomion, C.; et al. Epigenetics in forest trees: State of the art and potential implications for breeding and management in a context of climate change. In Plant Epigenetics Coming of Age for Breeding Applications; Mirouze, M., Bucher, E., Gallusci, P.B.T.-A., Eds.; Academic Press: Cambridge, MA, USA, 2018; Volume 88, pp. 387–453. ISBN 0065-2296. [Google Scholar]
- Wegrzyn, J.; Falk, T.; Grau, E.; Buehler, S.; Ramnath, R.; Herndon, N. Cyberinfrastructure and resources to enable an integrative approach to studying forest trees. Evol. Appl. 2020, 13, 228–241. [Google Scholar] [CrossRef]
- Olszewska, M.; Osiecka, R. Relationship between 2C DNA content, systematic position and level of DNA endoreplication during differentiation of root parenchyma in dicot shrubs and trees-comparison with herbaceous sp. Biochem. Physiol. Pflanz. 1984, 179, 641–657. [Google Scholar] [CrossRef]
- Zoldoš, V.; Papeš, D.; Brown, S.; Panaud, O.; Šiljak-Yakovlev, S. Genome size and base composition of seven Quercus species: Inter- and intra-population variation. Genome 1998, 41, 162–168. [Google Scholar] [CrossRef]
- Kremer, A.; Casasoli, M.; Barreneche, T.; Bodenes, C.; Sisco, P.; Kubisiak, T.; Scalfi, M.; Leonardi, S.; Bakker, E.; Buiteveld, J.; et al. Fagaceae Trees. In Genome Mapping and Molecular Breeding in Plants; Springer: Cham, Switzerland, 2007; Volume 7, pp. 161–187. [Google Scholar]
- Favre, J.; Brown, S. A flow cytometric evaluation of the nuclear DNA content and GC percent in genomes of European oak species. Ann. For. Sci. 1996, 53, 915–917. [Google Scholar] [CrossRef][Green Version]
- Rey, M.-D.; Castillejo, M.Á.; Sánchez-Lucas, R.; Guerrero-Sanchez, V.M.; López-Hidalgo, C.; Romero-Rodríguez, C.; Valero-Galván, J.; Sghaier-Hammami, B.; Simova-Stoilova, L.; Echevarría-Zomeño, S.; et al. Proteomics, holm oak (Quercus ilex L.) and other recalcitrant and orphan forest tree species: How do they see each other? Int. J. Mol. Sci. 2019, 20, 692. [Google Scholar] [CrossRef] [PubMed]
- Zoldoš, V.; Papeš, D.; Cerbah, M.; Panaud, O.; Besendorfer, V.; Šiljak-yakovlev, S. Molecular-cytogenetic studies of ribosomal genes and heterochromatin reveal conserved genome organization among 11 Quercus species. Theor. Appl. Genet. 1999, 99, 969–977. [Google Scholar] [CrossRef]
- Ribeiro, T.; Loureiro, J.; Santos, C.; Morais-Cecílio, L. Evolution of rDNA FISH patterns in the Fagaceae. Tree Genet. Genomes 2011, 7, 1113–1122. [Google Scholar] [CrossRef]
- Alexander, L.W.; Woeste, K.E. Pyrosequencing of the northern red oak (Quercus rubra L.) chloroplast genome reveals high quality polymorphisms for population management. Tree Genet. Genomes 2014, 10, 803–812. [Google Scholar] [CrossRef]
- Hu, H.-L.; Zhang, J.-Y.; Li, Y.-P.; Xie, L.; Chen, D.-B.; Li, Q.; Liu, Y.-Q.; Hui, S.-R.; Qin, L. The complete chloroplast genome of the daimyo oak, Quercus dentata Thunb. Conserv. Genet. Resour. 2019, 11, 409–411. [Google Scholar] [CrossRef]
- Hu, H.-L.; Wang, L.-Z.; Yang, J.; Zhang, R.-S.; Li, Q.; Liu, Y.-Q.; Qin, L. The complete chloroplast genome of Quercus fenchengensis and the phylogenetic implication. Mitochondrial DNA Part B 2019, 4, 3066–3067. [Google Scholar] [CrossRef] [PubMed]
- Harfouche, A.; Meilan, R.; Altman, A. Molecular and physiological responses to abiotic stress in forest trees and their relevance to tree improvement. Tree Physiol. 2014, 34, 1181–1198. [Google Scholar] [CrossRef]
- Della Coletta, R.; Qiu, Y.; Ou, S.; Hufford, M.B.; Hirsch, C.N. How the pan-genome is changing crop genomics and improvement. Genome Biol. 2021, 22, 3. [Google Scholar] [CrossRef] [PubMed]
- Amaral, J.; Ribeyre, Z.; Vigneaud, J.; Sow, M.D.; Fichot, R.; Messier, C.; Pinto, G.; Nolet, P.; Maury, S. Advances and promises of epigenetics for forest trees. Forests 2020, 11, 976. [Google Scholar] [CrossRef]
- Gutzat, R.; Scheid, O.M. Epigenetic responses to stress: Triple defense? Curr. Opin. Plant Biol. 2012, 15, 568–573. [Google Scholar] [CrossRef]
- Turgut-Kara, N.; As1kan, B.; Celik, H. Epigenetic memory and priming in plants. Genetica 2020, 148, 47–54. [Google Scholar] [CrossRef]
- Law, J.A.; Jacobsen, S.E. Molecular biology. Dynamic DNA methylation. Science 2009, 323, 1568–1569. [Google Scholar] [CrossRef]
- Johannes, F.; Schmitz, R.J. Spontaneous epimutations in plants. New Phytol. 2019, 221, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- Correia, B.; Valledor, L.; Meijón, M.; Rodriguez, J.L.; Dias, M.C.; Santos, C.; Cañal, M.J.; Rodriguez, R.; Pinto, G. Is the interplay between epigenetic markers related to the acclimation of cork oak plants to high temperatures? PLoS ONE 2013, 8, e53543. [Google Scholar] [CrossRef]
- Rico, L.; Ogaya, R.; Barbeta, A.; Penuelas, J. Changes in DNA methylation fingerprint of Quercus ilex trees in response to experimental field drought simulating projected climate change. Plant Biol. 2014, 16, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Platt, A.; Gugger, P.F.; Pellegrini, M.; Sork, V.L. Genome-wide signature of local adaptation linked to variable CpG methylation in oak populations. Mol. Ecol. 2015, 24, 3823–3830. [Google Scholar] [CrossRef]
- Chwialkowska, K.; Korotko, U.; Kosinska, J.; Szarejko, I.; Kwasniewski, M. Methylation Sensitive Amplification Polymorphism Sequencing (MSAP-Seq)-a method for high-throughput analysis of differentially methylated CCGG sites in plants with large genomes. Front. Plant Sci. 2017, 8, 2056. [Google Scholar] [CrossRef]
- Espinas, N.A.; Saze, H.; Saijo, Y. Epigenetic control of defense signaling and priming in plants. Front. Plant Sci. 2016, 7, 1201. [Google Scholar] [CrossRef] [PubMed]
- Balao, F.; Paun, O.; Alonso, C. Uncovering the contribution of epigenetics to plant phenotypic variation in Mediterranean ecosystems. Plant Biol. 2018, 20 (Suppl. S1), 38–49. [Google Scholar] [CrossRef]
- Yakovlev, I.; Carneros, E.; Lee, Y.; Olsen, J.; Fossdal, C.G. Transcriptional profiling of epigenetic regulators in somatic embryos during temperature induced formation of an epigenetic memory in Norway spruce. Planta 2016, 243, 1237–1249. [Google Scholar] [CrossRef] [PubMed]
- Hilker, M.; Schwachtje, J.; Baier, M.; Balazadeh, S.; Bäurle, I.; Geiselhardt, S.; Hincha, D.; Kunze, R.; Mueller-Roeber, B.; Rillig, M.; et al. Priming and memory of stress responses in organisms lacking a nervous system. Biol. Rev. 2016, 91, 753–761. [Google Scholar] [CrossRef]
- Ramirez-Prado, J.S.; Abulfaraj, A.A.; Rayapuram, N.; Benhamed, M.; Hirt, H. Plant immunity: From signaling to epigenetic control of defense. Trends Plant Sci. 2018, 23, 833–844. [Google Scholar] [CrossRef]
- Carneros, E.; Yakovlev, I.; Viejo, M.; Olsen, J.E.; Fossdal, C.G. The epigenetic memory of temperature during embryogenesis modifies the expression of bud burst-related genes in Norway spruce epitypes. Planta 2017, 246, 553–566. [Google Scholar] [CrossRef]
- Avramova, Z. Defence-related priming and responses to recurring drought: Two manifestations of plant transcriptional memory mediated by the ABA and JA signalling pathways. Plant. Cell Environ. 2019, 42, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Hilker, M.; Schmülling, T. Stress priming, memory, and signalling in plants. Plant. Cell Environ. 2019, 42, 753–761. [Google Scholar] [CrossRef]
- Gallusci, P.; Dai, Z.; Génard, M.; Gauffretau, A.; Leblanc-Fournier, N.; Richard-Molard, C.; Vile, D.; Brunel-Muguet, S. Epigenetics for plant improvement: Current knowledge and modeling avenues. Trends Plant Sci. 2017, 22, 610–623. [Google Scholar] [CrossRef] [PubMed]
- Kukurba, K.R.; Montgomery, S.B. RNA sequencing and analysis. Cold Spring Harb. Protoc. 2015, 2015, 951–969. [Google Scholar] [CrossRef] [PubMed]
- Spieß, N.; Oufir, M.; Matušíková, I.; Stierschneider, M.; Kopecky, D.; Homolka, A.; Burg, K.; Fluch, S.; Hausman, J.; Wilhelm, E. Ecophysiological and transcriptomic responses of oak (Quercus robur) to long-term drought exposure and rewatering. Environ. Exp. Bot. 2012, 77, 117–126. [Google Scholar] [CrossRef]
- Almeida, T.; Pinto, G.; Correia, B.; Santos, C.; Gonçalves, S. Plant Physiology and Biochemistry QsMYB1 expression is modulated in response to heat and drought stresses and during plant recovery in Quercus suber. Plant Physiol. Biochem. 2013, 73, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Rasheed-Depardieu, C.; Parelle, J.; Tatin-Froux, F.; Parent, C.; Capelli, N. Short-term response to waterlogging in Quercus petraea and Quercus robur: A study of the root hydraulic responses and the transcriptional pattern of aquaporins. Plant Physiol. Biochem. 2015, 97, 323–330. [Google Scholar] [CrossRef]
- Ebadzad, G.; Cravador, A. Quantitative RT-PCR analysis of differentially expressed genes in Quercus suber in response to Phytophthora cinnamomi infection. Springerplus 2014, 3, 613. [Google Scholar] [CrossRef][Green Version]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Ueno, S.; Le Provost, G.; Léger, V.; Klopp, C.; Noirot, C.; Frigerio, J.-M.; Salin, F.; Salse, J.; Abrouk, M.; Murat, F.; et al. Bioinformatic analysis of ESTs collected by Sanger and pyrosequencing methods for a keystone forest tree species: Oak. BMC Genom. 2010, 11, 650. [Google Scholar] [CrossRef]
- Tarkka, M.T.; Herrmann, S.; Wubet, T.; Feldhahn, L.; Recht, S.; Kurth, F.; Mailänder, S.; Bönn, M.; Neef, M.; Angay, O.; et al. OakContigDF159.1, a reference library for studying differential gene expression in Quercus robur during controlled biotic interactions: Use for quantitative transcriptomic profiling of oak roots in ectomycorrhizal symbiosis. New Phytol. 2013, 199, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Leal, J.B.; Abreu, I.A.; Alabaca, C.S.; Almeida, M.H.; Almeida, P.; Almeida, T.; Amorim, M.I.; Araujo, S.; Azevedo, H.; Badia, A.; et al. A comprehensive assessment of the transcriptome of cork oak (Quercus suber) through EST sequencing. BMC Genom. 2014, 15, 371. [Google Scholar] [CrossRef]
- López-Hidalgo, C.; Guerrero-Sánchez, V.M.; Gómez-Gálvez, I.; Sánchez-Lucas, R.; Castillejo-Sánchez, M.A.; Maldonado-Alconada, A.M.; Valledor, L.; Jorrín-Novo, J.V. A multi-omics analysis pipeline for the metabolic pathway reconstruction in the orphan species Quercus ilex. Front. Plant Sci. 2018, 9, 935. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Sanchez, V.M.; Maldonado-Alconada, A.M.; Sánchez-Lucas, R.; Rey, M.D. Specific protein database creation from transcriptomics data in nonmodel species: Holm oak (Quercus ilex L.). In Plant Proteomics. Methods in Molecular Biology; Jorrin-Novo, J., Valledor, L., Castillejo, M.A., Rey, M., Eds.; Springer: New York, NY, USA, 2020; Volume 2139, pp. 57–68. ISBN 978-1-0716-0528-8. [Google Scholar]
- Park, O.K. Proteomic studies in plants. J. Biochem. Mol. Biol. 2004, 37, 133–138. [Google Scholar] [CrossRef]
- Jorge, I.; Navarro, R.M.; Lenz, C.; Ariza, D.; Jorrín, J. Variation in the Holm oak leaf proteome at different plant developmental stages, between provenances and in response to drought stress. Proteomics 2006, 6 (Suppl. S1), S207–S214. [Google Scholar] [CrossRef]
- Romero-Rodríguez, M.C.; Pascual, J.; Valledor, L.; Jorrín-Novo, J. Improving the quality of protein identification in non-model species. Characterization of Quercus ilex seed and Pinus radiata needle proteomes by using SEQUEST and custom databases. J. Proteom. 2014, 105, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gálvez, I.; Sánchez-Lucas, R.; San-Eufrasio, B.; Rodríguez de Francisco, L.E.; Maldonado-Alconada, A.M.; Fuentes-Almagro, C.; Castillejo, M.A. Optimizing shotgun proteomics analysis for a confident protein identification and quantitation in orphan plant species: The case of Holm oak (Quercus ilex). In Plant Proteomics. Methods in Molecular Biology; Jorrin-Novo, J., Valledor, L., Castillejo, M.A., Rey, M., Eds.; Springer: New York, NY, USA, 2020; Volume 2139, pp. 157–168. ISBN 978-1-0716-0527-1. [Google Scholar]
- Escandón, M.; Jorrín-Novo, J.V.; Castillejo, M.Á. Application and optimization of label-free shotgun approaches in the study of Quercus ilex. J. Proteom. 2020, 233, 104082. [Google Scholar] [CrossRef]
- Sghaier-Hammami, B.; Castillejo, M.Á.; Baazaoui, N.; Jorrín-Novo, J.V.; Escandón, M. GeLC-Orbitrap/MS and 2-DE-MALDI-TOF/TOF comparative proteomics analysis of seed cotyledons from the non-orthodox Quercus ilex tree species. J. Proteom. 2021, 233, 104087. [Google Scholar] [CrossRef]
- Valero-Galván, J.; Valledor, L.; Navarro-Cerrillo, R.M.; Gil Pelegrín, E.; Jorrín-Novo, J.V. Studies of variability in Holm oak (Quercus ilex subsp. ballota [Desf.] Samp.) through acorn protein profile analysis. J. Proteom. 2011, 74, 1244–1255. [Google Scholar] [CrossRef] [PubMed]
- Valero-Galván, J.; Sghaier-Hammami, B.; Navarro-Cerrillo, R.M.; Jorrín-Novo, J.V. Natural variability and responses to stresses in andalusia Holm oak (Quercus ilex subsp. ballota) populations. In Oak: Ecology, Types and Management; Chuteira, C.A., Grão, A.B., Eds.; Nova Science Publisher: Hauppauge, NY, USA, 2012; pp. 193–206. ISBN 978-1-61942-492-0. [Google Scholar]
- Valero-Galván, J.; González, R.; Valledor, L.; Navarro-Cerrillo, R.M.; Jorrin-Novo, J.V. Proteotyping of Holm oak (Quercus ilex subsp. ballota) provenances through proteomic analysis of acorn flour. Methods Mol. Biol. 2014, 1072, 709–723. [Google Scholar]
- López-Hidalgo, C.; Escandón, M.; Valledor, L.; Jorrin-Novo, J.V. A pipeline for metabolic pathway reconstruction in plant orphan species. In Plant Proteomics. Methods in Molecular Biology; Jorrin-Novo, J., Valledor, L., Castillejo, M.A., Rey, M., Eds.; Springer: New York, NY, USA, 2020; Volume 2139, pp. 367–380. [Google Scholar]
- Romero-Rodríguez, M.C.; Abril, N.; Sánchez-Lucas, R.; Jorrín-Novo, J.V. Multiplex staining of 2-DE gels for an initial phosphoproteome analysis of germinating seeds and early grown seedlings from a non-orthodox specie: Quercus ilex L. subsp. ballota [Desf.] Samp. Front. Plant Sci. 2015, 6, 620. [Google Scholar] [CrossRef]
- Romero-Rodríguez, M.C.; Jorrín-Novo, J.V.; Castillejo, M.A. Toward characterizing germination and early growth in the non-orthodox forest tree species Quercus ilex through complementary gel and gel-free proteomic analysis of embryo and seedlings. J. Proteom. 2019, 197, 60–70. [Google Scholar] [CrossRef]
- Sghaier-Hammami, B.; Redondo-López, I.; Valero-Galvàn, J.; Jorrín-Novo, J.V. Protein profile of cotyledon, tegument, and embryonic axis of mature acorns from a non-orthodox plant species: Quercus ilex. Planta 2016, 243, 369–396. [Google Scholar] [CrossRef] [PubMed]
- Sghaier-Hammami, B.; Hammami, S.B.M.; Baazaoui, N.; Gómez-Díaz, C.; Jorrín-Novo, J.V. Dissecting the seed maturation and germination processes in the Non-Orthodox Quercus ilex species based on protein signatures as revealed by 2-DE coupled to MALDI-TOF/TOF proteomics strategy. Int. J. Mol. Sci. 2020, 21, 4870. [Google Scholar] [CrossRef] [PubMed]
- Ricardo, C.P.P.; Martins, I.; Francisco, R.; Sergeant, K.; Pinheiro, C.; Campos, A.; Renaut, J.; Fevereiro, P. Proteins associated with cork formation in Quercus suber L. stem tissues. J. Proteom. 2011, 74, 1266–1278. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, C.; Wienkoop, S.; de Almeida, J.F.; Brunetti, C.; Zarrouk, O.; Planchon, S.; Gori, A.; Tattini, M.; Ricardo, C.P.; Renaut, J.; et al. Phellem cell-wall components are discriminants of cork quality in Quercus suber. Front. Plant Sci. 2019, 10, 944. [Google Scholar] [CrossRef]
- Gómez, A.; López, J.A.; Pintos, B.; Camafeita, E.; Bueno, M.A. Proteomic analysis from haploid and diploid embryos of Quercus suber L. identifies qualitative and quantitative differential expression patterns. Proteomics 2009, 9, 4355–4367. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Garay, A.; Lopez, J.A.; Camafeita, E.; Bueno, M.A.; Pintos, B. Proteomic perspective of Quercus suber somatic embryogenesis. J. Proteom. 2013, 93, 314–325. [Google Scholar] [CrossRef]
- Huerta-Ocampo, J.Á.; Valenzuela-Corral, A.; Robles-Burgueño, M.D.R.; Guzmán-Partida, A.M.; Hernández-Oñate, M.Á.; Vázquez-Moreno, L.; Pavón-Romero, G.F.; Terán, L.M. Proteomic identification of allergenic proteins in red oak (Quercus rubra) pollen. World Allergy Organ. J. 2020, 13, 100111. [Google Scholar] [CrossRef]
- Pedrosa, M.; Guerrero-Sanchez, V.M.; Canales-Bueno, N.; Loli-Ausejo, D.; Castillejo, M.Á.; Quirce, S.; Jorrin-Novo, J.V.; Rodriguez-Perez, R. Quercus ilex pollen allergen, Que i 1, responsible for pollen food allergy syndrome caused by fruits in Spanish allergic patients. Clin. Exp. Allergy 2020, 50, 815–823. [Google Scholar] [CrossRef]
- Rey, M.D.; Valledor, L.; Castillejo, M.A.; Sánchez-Lucas, R.; López-Hidalgo, C.; Guerrero-Sánchez, V.M.; Colina, F.J.; Escandón, M.; Maldonado-Alconada, A.M.; Jorrín-Novo, J.V. Recent advances in MS-based plant proteomics: Proteomics data validation through integration with other classic and -omics approaches. In Progress in Botany; Cánovas, F., Lüttge, U., Leuschner, C., Risueño, M., Eds.; Springer: Cham, Switzerland, 2019; Volume 81, ISBN 978-3-030-36327-7. [Google Scholar]
- Echevarría-Zomeño, S.; Ariza, D.; Jorge, I.; Lenz, C.; del Campo, A.D.; Jorrín, J.; Navarro, R.M. Changes in the protein profile of Quercus ilex leaves in response to drought stress and recovery. J. Plant Physiol. 2009, 166, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Valero-Galván, J.; González-Fernández, R.; Navarro-Cerrillo, R.; Gil-Pelegrin, E.; Jorrín-Novo, J. Physiological and proteomic analyses of drought stress response in Holm oak provenances. J. Proteome Res. 2013, 12, 5110–5123. [Google Scholar] [CrossRef] [PubMed]
- Simova-Stoilova, L.P.; Romero-Rodríguez, M.C.; Sánchez-Lucas, R.; Navarro-Cerrillo, R.M.; Medina-Aunon, J.A.; Jorrín-Novo, J.V. 2-DE proteomics analysis of drought treated seedlings of Quercus ilex supports a root active strategy for metabolic adaptation in response to water shortage. Front. Plant Sci. 2015, 6, 627. [Google Scholar] [CrossRef]
- Sghaier-Hammami, B.; Valero-Galvàn, J.; Romero-Rodríguez, M.C.; Navarro-Cerrillo, R.M.; Abdelly, C.; Jorrín-Novo, J. Physiological and proteomics analyses of Holm oak (Quercus ilex subsp. ballota [Desf.] Samp.) responses to Phytophthora cinnamomi. Plant Physiol. Biochem. 2013, 71, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Sergeant, K.; Spiess, N.; Renaut, J.; Wilhelm, E.; Hausman, J.F. One dry summer: A leaf proteome study on the response of oak to drought exposure. J. Proteom. 2011, 74, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, H.; O’Gorman, A.; Brennan, L. Metabolomics as a tool in nutritional research. Curr. Opin. Lipidol. 2015, 26, 30–34. [Google Scholar] [CrossRef]
- Patel, S.; Ahmed, S. Emerging field of metabolomics: Big promise for cancer biomarker identification and drug discovery. J. Pharm. Biomed. Anal. 2015, 107, 63–74. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, F.; Deng, Y.; Zhao, Y.; Ren, H. Metagenomic and metabolomic analysis of the toxic effects of trichloroacetamide-induced gut microbiome and urine metabolome perturbations in mice. J. Proteome Res. 2015, 14, 1752–1761. [Google Scholar] [CrossRef]
- Shulaev, V.; Cortes, D.; Miller, G.; Mittler, R. Metabolomics for plant stress response. Physiol. Plant. 2008, 132, 199–208. [Google Scholar] [CrossRef]
- Monteiro, M.S.; Carvalho, M.; Bastos, M.L.; Guedes de Pinho, P. Metabolomics analysis for biomarker discovery: Advances and challenges. Curr. Med. Chem. 2013, 20, 257–271. [Google Scholar] [CrossRef]
- Escandón, M.; Meijón, M.; Valledor, L.; Pascual, J.; Pinto, G.; Cañal, M.J. Metabolome integrated analysis of high-temperature response in Pinus radiata. Front. Plant Sci. 2018, 9, 485. [Google Scholar] [CrossRef]
- López-Hidalgo, C.; Trigueros, M.; Menéndez, M.; Jorrin-Novo, J.V. Phytochemical composition and variability in Quercus ilex acorn morphotypes as determined by NIRS and MS-based approaches. Food Chem. 2021, 338, 127803. [Google Scholar] [CrossRef]
- Schwanz, P.; Polle, A. Differential stress responses of antioxidative systems to drought in pendunculate oak (Quercus robur) and maritime pine (Pinus pinaster) grown under high CO2 concentrations. J. Exp. Bot. 2001, 52, 133–143. [Google Scholar] [PubMed]
- Chaves, I.; Passarinho, J.A.P.; Capitão, C.; Chaves, M.M.; Fevereiro, P.; Ricardo, C.P.P. Temperature stress effects in Quercus suber leaf metabolism. J. Plant Physiol. 2011, 168, 1729–1734. [Google Scholar] [CrossRef] [PubMed]
- Loreto, F. Distribution of isoprenoid emitters in the Quercus genus around the world: Chemo-taxonomical implications and evolutionary considerations based on the ecological function of the trait. Perspect. Plant Ecol. Evol. Syst. 2002, 5, 185–192. [Google Scholar] [CrossRef]
- Loreto, F.; Bagnoli, F.; Fineschi, S. One species, many terpenes: Matching chemical and biological diversity. Trends Plant Sci. 2009, 14, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Llusià, J. Linking photorespiration, monoterpenes and thermotolerance in Quercus. New Phytol. 2002, 155, 227–237. [Google Scholar] [CrossRef]
- Rivas-Ubach, A.; Gargallo-Garriga, A.; Sardans, J.; Oravec, M.; Mateu-Castell, L.; Pérez-Trujillo, M.; Parella, T.; Ogaya, R.; Urban, O.; Peñuelas, J. Drought enhances folivory by shifting foliar metabolomes in Quercus ilex trees. New Phytol. 2014, 202, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Keenan, T.; Niinemets, Ü.; Sabate, S.; Gracia, C.; Peñuelas, J. Seasonality of monoterpene emission potentials in Quercus ilex and Pinus pinea: Implications for regional VOC emissions modeling. J. Geophys. Res. Atmos. 2009, 114. [Google Scholar] [CrossRef]
- Genard-Zielinski, A.-C.; Boissard, C.; Fernandez, C.; Kalogridis, C.; Lathière, J.; Gros, V.; Bonnaire, N.; Ormeño, E. Variability of BVOC emissions from a Mediterranean mixed forest in southern France with a focus on Quercus pubescens. Atmos. Chem. Phys. 2015, 15, 431–446. [Google Scholar] [CrossRef]
- Llusià, J.; Peñuelas, J.; Seco, R.; Filella, I. Seasonal changes in the daily emission rates of terpenes by Quercus ilex and the atmospheric concentrations of terpenes in the natural park of Montseny, NE Spain. J. Atmos. Chem. 2012, 69, 215–230. [Google Scholar] [CrossRef]
- Loreto, F.; Schnitzler, J.-P. Abiotic stresses and induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef]
- Llusia, J.; Roahtyn, S.; Yakir, D.; Rotenberg, E.; Seco, R.; Guenther, A.; Peñuelas, J. Photosynthesis, stomatal conductance and terpene emission response to water availability in dry and mesic Mediterranean forests. Trees 2016, 30, 749–759. [Google Scholar] [CrossRef]
- Brüggemann, N.; Schnitzler, J. Comparison of isoprene emission, intercellular isoprene concentration and photosynthetic performance in water-limited oak (Quercus pubescens Willd. and Quercus robur L.) saplings. Plant Biol. 2002, 4, 456–463. [Google Scholar] [CrossRef]
- Rodríguez-Calcerrada, J.; Buatois, B.; Chiche, E.; Shahin, O.; Staudt, M. Leaf isoprene emission declines in Quercus pubescens seedlings experiencing drought—Any implication of soluble sugars and mitochondrial respiration? Environ. Exp. Bot. 2013, 85, 36–42. [Google Scholar] [CrossRef]
- Llusià, J.; Peñuelas, J.; Asensio, D.; Munné-Bosch, S. Airborne limonene confers limited thermotolerance to Quercus ilex. Physiol. Plant. 2005, 123, 40–48. [Google Scholar] [CrossRef]
- Lavoir, A.-V.; Staudt, M.; Schnitzler, J.; Landais, D.; Massol, F.; Rocheteau, A.; Rodríguez, R.; Zimmer, I.; Rambal, S. Drought reduced monoterpene emissions from the evergreen Mediterranean oak Quercus ilex: Results from a throughfall displacement experiment. Biogeosciences 2009, 6, 1167–1180. [Google Scholar] [CrossRef]
- Bourtsoukidis, E.; Kawaletz, H.; Radacki, D.; Schütz, S.; Hakola, H.; Hellén, H.; Noe, S.; Mölder, I.; Ammer, C.; Bonn, B. Impact of flooding and drought conditions on the emission of volatile organic compounds of Quercus robur and Prunus serotina. Trees 2014, 28, 193–204. [Google Scholar] [CrossRef]
- Faiola, C.; Taipale, D. Impact of insect herbivory on plant stress volatile emissions from trees: A synthesis of quantitative measurements and recommendations for future research. Atmos. Environ. X 2020, 5, 100060. [Google Scholar] [CrossRef]
- Suseela, V.; Tharayil, N.; Xing, B.; Dukes, J.S. Warming and drought differentially influence the production and resorption of elemental and metabolic nitrogen pools in Quercus rubra. Glob. Change Biol. 2015, 21, 4177–4195. [Google Scholar] [CrossRef] [PubMed]
- Palsson, B. Systems Biology. Constraint-Based Reconstruction and Analysis; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Breitling, R. What is systems biology? Front. Physiol. 2010, 1, 9. [Google Scholar] [CrossRef]
- Escandón, M.; Valledor, L.; Pascual, J.; Pinto, G.; Cañal, M.J.; Meijón, M. System-wide analysis of short-term response to high temperature in Pinus radiata. J. Exp. Bot. 2017, 68, 3629–3641. [Google Scholar] [CrossRef]
- Pascual, J.; Cañal, M.J.; Escandón, M.; Meijón, M.; Weckwerth, W.; Valledor, L. Integrated physiological, proteomic, and metabolomic analysis of Ultra Violet (UV) stress responses and adaptation mechanisms in Pinus radiata. MCP 2017, 16, 485–501. [Google Scholar] [PubMed]
- Meijón, M.; Feito, I.; Oravec, M.; Delatorre, C.; Weckwerth, W.; Majada, J.; Valledor, L. Exploring natural variation of Pinus pinaster Aiton using metabolomics: Is it possible to identify the region of origin of a pine from its metabolites? Mol. Ecol. 2016, 25, 959–976. [Google Scholar] [CrossRef] [PubMed]
- Kremer, A.; Abbott, A.G.; Carlson, J.E.; Manos, P.S.; Plomion, C.; Sisco, P.; Staton, M.E.; Ueno, S.; Vendramin, G.G. Genomics of Fagaceae. Tree Genet. Genomes 2012, 8, 583–610. [Google Scholar] [CrossRef]
- Initiative, T.A.G. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [PubMed]
- Estravis-Barcala, M.; Mattera, M.G.; Soliani, C.; Bellora, N.; Opgenoorth, L.; Heer, K.; Arana, M.V. Molecular bases of responses to abiotic stress in trees. J. Exp. Bot. 2020, 71, 3765–3779. [Google Scholar] [CrossRef]
- Weighill, D.; Tschaplinski, T.J.; Tuskan, G.A.; Jacobson, D. Data Integration in Poplar: ‘Omics Layers and Integration Strategies. Front. Genet. 2019, 10, 874. [Google Scholar] [CrossRef]
- Martínez, M.T.; Corredoira, E.; Vieitez, A.; Cernadas, M.J.; Montenegro, R.; Ballester, A.; Vieitez, F.J.; José, M.C.S. Micropropagation of mature Quercus ilex L. trees by axillary budding. Plant Cell Tissue Organ Cult. 2017, 131, 499–512. [Google Scholar] [CrossRef]
- Plomion, C.; Bastien, C.; Bogeat-Triboulot, M.-B.; Bouffier, L.; Déjardin, A.; Duplessis, S.; Fady, B.; Heuertz, M.; Gac, A.; Provost, G.; et al. Forest tree genomics: 10 achievements from the past 10 years and future prospects. Ann. For. Sci. 2016, 73, 77–103. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).