A Genetic Linkage Map of BC2 Population Reveals QTL Associated with Plant Architecture Traits in Lagerstroemia


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Phenotypic Data Collection
2.3. DNA Extraction and Molecular Marker Development
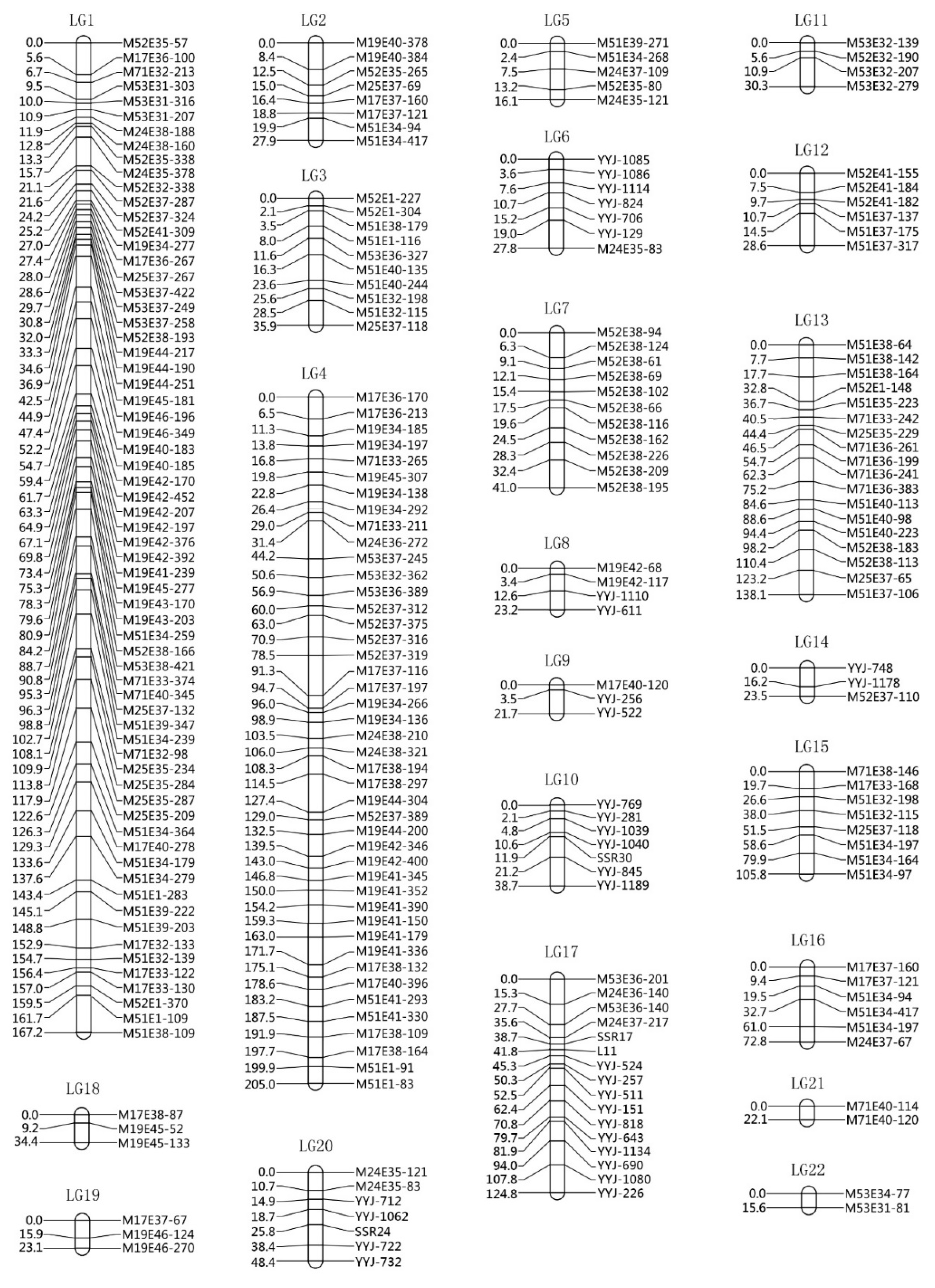
2.4. Data Analysis and Map Construction
2.5. QTL Analysis
3. Results
3.1. Phenotype Data Analysis
3.2. Polymorphisms Revealed by Molecular Markers
3.3. Linkage Analysis and Map Construction
3.4. QTL Mapping
4. Discussion
4.1. Genetic Mapping Is an Effective Way of Gene Mapping
4.2. Mapping Population and Molecular Markers
4.3. Genetic Linkage Map
4.4. QTL Mapping for Plant Architecture Traits
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, B.; Smith, S.M.; Li, J. Genetic regulation of shoot architecture. Annu. Rev. Plant Biol. 2018, 69, 437–468. [Google Scholar] [CrossRef]
- Mencuccini, M. Dwarf trees, super-sized shrubs and scaling: Why is plant stature so important? Plant Cell Environ. 2015, 38, 1–3. [Google Scholar] [CrossRef]
- Li, S.; Zheng, T.; Zhuo, X.; Li, Z.; Wang, J.; Cheng, T.; Zhang, Q. Isolation of the crape myrtle decreased apical dominance gene lfidad2 and characterization of its function in the control of axillary branching. Sci. Hortic. 2020, 262, 109055. [Google Scholar] [CrossRef]
- Peng, J.; Richards, D.E.; Hartley, N.M.; Murphy, G.P.; Devos, K.M.; Flintham, J.E.; Beales, J.; Fish, L.J.; Worland, A.J.; Pelica, F.; et al. ‘Green revolution’ genes encode mutant gibberellin response modulators. Nature 1999, 400, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Jin, Y.; Liu, W.; Li, F.; Fang, J.; Yin, Y.; Qian, Q.; Zhu, L.; Chu, C. Dwarf and low-tillering, a new member of the gras family, plays positive roles in brassinosteroid signaling in rice. Plant J. 2009, 58, 803–816. [Google Scholar] [CrossRef]
- Lu, F.; Ammiraju, J.S.; Sanyal, A.; Zhang, S.; Song, R.; Chen, J.; Li, G.; Sui, Y.; Song, X.; Cheng, Z.; et al. Comparative sequence analysis of monoculm1-orthologous regions in 14 oryza genomes. Proc. Natl. Acad. Sci. USA 2009, 106, 2071–2076. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Lin, Z.; Li, H.; Li, X.; Li, J.; Wang, Y.; Zhang, X.; Zhu, Z.; Zhai, W.; Wang, X.; et al. Tac1, a major quantitative trait locus controlling tiller angle in rice. Plant J. 2007, 52, 891–898. [Google Scholar] [CrossRef]
- Li, S.; Zheng, T.; Zhuo, X.; Li, Z.; Li, L.; Li, P.; Qiu, L.; Pan, H.; Wang, J.; Cheng, T.; et al. Transcriptome profiles reveal that gibberellin-related genes regulate weeping traits in crape myrtle. Hortic. Res. 2020, 7, 54. [Google Scholar] [CrossRef]
- Virk, P.S.; Khush, G.S.; Peng, S. Breeding to enhance yield potential of rice at irri: The ideotype approach. Int. Rice Res. Notes 2004, 29, 5–9. [Google Scholar]
- Clark, R.M.; Wagler, T.N.; Quijada, P.; Doebley, J. A distant upstream enhancer at the maize domestication gene tb1 has pleiotropic effects on plant and inflorescent architecture. Nat. Genet. 2006, 38, 594–597. [Google Scholar] [CrossRef]
- Begna, S.; Ri, H.; Dwyer, L.; Dw, S.; Smith, D. Variability among maize hybrids differing in canopy architecture for above-ground dry matter and grain yield. Maydica 2000, 45, 135–141. [Google Scholar]
- Baldi, P.; Wolters, P.J.; Komjanc, M.; Viola, R.; Velasco, R.; Salvi, S. Genetic and physical characterisation of the locus controlling columnar habit in apple (malus × domestica borkh.). Mol. Breed. 2012, 31, 429–440. [Google Scholar] [CrossRef]
- Harrison, N.; Harrison, R.J.; Barber-Perez, N.; Cascant-Lopez, E.; Cobo-Medina, M.; Lipska, M.; Conde-Ruiz, R.; Brain, P.; Gregory, P.J.; Fernandez-Fernandez, F. A new three-locus model for rootstock-induced dwarfing in apple revealed by genetic mapping of root bark percentage. J. Exp. Bot. 2016, 67, 1871–1881. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Zhang, D.; Ma, J.; Mao, J.; Zhang, B.; Han, M. Isolation and expression analysis of mdabcb19 gene and its promoter from dwarfing apple rootstock. Acta Hortic. 2017, 44, 409–421. [Google Scholar]
- Trifonova, A.; Savova, D.; Ivanova, K. Agrobacterium-mediated transformation of the apple cultivar granny smith. In Progress in Temperate Fruit Breeding; Springer: Dordrecht, The Netherlands, 1994; Volume 1, pp. 343–347. [Google Scholar]
- Holefors, A.; Xue, Z.-T.; Welander, M. Transformation of the apple rootstock m26 with the rola gene and its influence on growth. Plant Sci. 1998, 136, 69–78. [Google Scholar] [CrossRef]
- Xue, Z.-T.; Holefors, A.; Welander, M. Intron splicing in 5′ untranslated region of the rola transcript in transgenic apple. J. Plant Physiol. 2008, 165, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Niu, L.; Chagné, D.; Cui, G.; Pan, L.; Foster, T.; Zhang, R.; Zeng, W.; Wang, Z. Fine mapping of the temperature-sensitive semi-dwarf (tssd) locus regulating the internode length in peach (prunus persica). Mol. Breed. 2016, 36, 20. [Google Scholar] [CrossRef]
- Foster, T.M.; Ledger, S.E.; Janssen, B.J.; Luo, Z.; Drummond, R.S.M.; Tomes, S.; Karunairetnam, S.; Waite, C.N.; Funnell, K.A.; van Hooijdonk, B.M.; et al. Expression of mdccd7 in the scion determines the extent of sylleptic branching and the primary shoot growth rate of apple trees. J. Exp. Bot. 2018, 69, 2379–2390. [Google Scholar] [CrossRef]
- Dardick, C.; Callahan, A.; Horn, R.; Ruiz, K.B.; Zhebentyayeva, T.; Hollender, C.; Whitaker, M.; Abbott, A.; Scorza, R. Ppetac1 promotes the horizontal growth of branches in peach trees and is a member of a functionally conserved gene family found in diverse plants species. Plant J. 2013, 75, 618–630. [Google Scholar] [CrossRef]
- Sun, L.; Wang, Y.; Yan, X.; Cheng, T.; Ma, K.; Yang, W.; Pan, H.; Zheng, C.; Zhu, X.; Wang, J.; et al. Genetic control of juvenile growth and botanical architecture in an ornamental woody plant, prunus mume sieb. Et zucc. As revealed by a high-density linkage map. BMC Genet. 2014, 15 (Suppl. 1), S1. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, K.; Hou, D.; Cai, J.; Zhang, Q.; Cheng, T.; Pan, H.; Yang, W. Genome-wide discovery of DNA polymorphisms in mei (prunus mume sieb. Et zucc.), an ornamental woody plant, with contrasting tree architecture and their functional relevance for weeping trait. Plant Mol. Biol. Rep. 2017, 35, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Sun, R.; Hou, X.; Zheng, H.; Zhang, F.; Zhang, Y.; Liu, B.; Liang, J.; Zhuang, M.; Liu, Y.; et al. Subgenome parallel selection is associated with morphotype diversification and convergent crop domestication in brassica rapa and brassica oleracea. Nat. Genet. 2016, 48, 1218–1224. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Meng, R.; Pan, H.-T.; Gao, Y.-K.; Sun, M.; Song, P.; Wang, X.-F.; Zhang, Q.-X. Isolation and characterization of microsatellite markers from lagerstroemia caudata (lythraceae) and cross-amplification in other related species. Conserv. Genet. Resour. 2010, 2, 89–91. [Google Scholar] [CrossRef]
- Wang, J.; Dai, X.; Chen, Y.; Yang, Y.; Zhang, X.; Li, S.; Yin, T. Genomic sequencing using 454 pyrosequencing and development of an ssr primer database for lagerstroemia indica l. Plant Omics 2015, 8, 17–23. [Google Scholar]
- Ye, Y.; Cai, M.; Ju, Y.; Jiao, Y.; Feng, L.; Pan, H.; Cheng, T.; Zhang, Q. Identification and validation of snp markers linked to dwarf traits using slaf-seq technology in lagerstroemia. PLoS ONE 2016, 11, e0158970. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Feng, L.; Liang, X.; Liu, T.; Cai, M.; Cheng, T.; Wang, J.; Zhang, Q.; Pan, H. Characterization, validation, and cross-species transferability of newly developed est-ssr markers and their application for genetic evaluation in crape myrtle (Lagerstroemia spp.). Mol. Breed. 2019, 39, 26. [Google Scholar] [CrossRef]
- Ye, Y.J.; Liu, Y.; Cai, M.; He, D.; Shen, J.S.; Ju, Y.Q.; Bian, X.M.; Pan, H.T.; Zhang, Q.X. Screening of molecular markers linked to dwarf trait in crape myrtle by bulked segregant analysis. Genet. Mol. Res. 2015, 14, 4369–4380. [Google Scholar] [CrossRef]
- He, D.; Liu, Y.; Cai, M.; Pan, H.T.; Zhang, Q.X.; Wang, X.Y.; Wang, X.J. Genetic diversity of lagerstroemia (lythraceae) species assessed by simple sequence repeat markers. Genet. Mol. Res. 2012, 11, 3522–3533. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.-J.; Wu, J.-Y.; Feng, L.; Ju, Y.-Q.; Cai, M.; Cheng, T.-R.; Pan, H.-T.; Zhang, Q.-X. Heritability and gene effects for plant architecture traits of crape myrtle using major gene plus polygene inheritance analysis. Sci. Hortic. 2017, 225, 335–342. [Google Scholar] [CrossRef]
- Ju, Y.; Feng, L.; Wu, J.; Ye, Y.; Zheng, T.; Cai, M.; Cheng, T.; Wang, J.; Zhang, Q.; Pan, H. Transcriptome analysis of the genes regulating phytohormone and cellular patterning in lagerstroemia plant architecture. Sci. Rep. 2018, 8, 15162. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.R.; Gao, Y.K.; Xu, L.J.; Zhang, Q.X. Genetic diversity of aquilegia (ranunculaceae) species and cultivars assessed by aflps. Genet. Mol. Res. 2011, 10, 817–827. [Google Scholar] [CrossRef]
- He, D.; Liu, Y.; Cai, M.; Pan, H.; Zhang, Q.; Debener, T. The first genetic linkage map of crape myrtle (lagerstroemia) based on amplification fragment length polymorphisms and simple sequence repeats markers. Plant Breed. 2014, 133, 138–144. [Google Scholar] [CrossRef]
- Liu, Y.; He, D.; Cai, M.; Tang, W.; Li, X.Y.; Pan, H.T.; Zhang, Q.X. Development of microsatellite markers for lagerstroemia indica (lythraceae) and related species. Appl. Plant Sci. 2013, 1, 1200203. [Google Scholar] [CrossRef]
- Zhang, E.; Wang, P.; Li, Y.; Wang, S.; Li, L.; Yang, R. Development and utility of est-ssr markers in lagerstroemia indica. North. Hortic. 2016, 40, 107–111. [Google Scholar]
- Wang, X.; Dean, D.; Wadl, P.; Hadziabdic, D.; Scheffler, B.; Rinehart, T.; Cabrera, R.; Trigiano, R. Development of microsatellite markers from crape myrtle (Lagerstroemia L.). HortScience 2010, 45, 842–844. [Google Scholar] [CrossRef]
- Kosambi, D.D. The estimation of map distances from recombination values. Ann. Eugen. 1944, 12, 172–175. [Google Scholar] [CrossRef]
- Liu, R.-H.; Meng, J.-L. MapDraw: A microsoft excel macro for drawing genetic linkage maps based on given genetic linkage data. Yi Chuan Hered. 2003, 25, 317–321. [Google Scholar]
- Xing, Z.; Tan, F.; Hua, P.; Sun, L.; Xu, G.; Zhang, Q. Characterization of the main effects, epistatic effects and their environmental interactions of qtls on the genetic basis of yield traits in rice. Theor. Appl. Genet. 2002, 105, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Xing, Y.; Mao, H.; Lu, T.; Han, B.; Xu, C.; Li, X.; Zhang, Q. Gs3, a major qtl for grain length and weight and minor qtl for grain width and thickness in rice, encodes a putative transmembrane protein. Theor. Appl. Genet. 2006, 112, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Vendramin, E.; Pea, G.; Dondini, L.; Pacheco, I.; Dettori, M.T.; Gazza, L.; Scalabrin, S.; Strozzi, F.; Tartarini, S.; Bassi, D.; et al. A unique mutation in a myb gene cosegregates with the nectarine phenotype in peach. PLoS ONE 2014, 9, e90574. [Google Scholar] [CrossRef] [PubMed]
- Peltier, D.; Farcy, E.; Dulieu, H.; Berville, A. Origin, distribution and mapping of rapd markers from wildpetunia species inpetunia hybrida hort lines. Theor. Appl. Genet. 1994, 88, 637–645. [Google Scholar] [CrossRef]
- Abe, H.; Nakano, M.; Nakatsuka, A.; Nakayama, M.; Koshioka, M.; Yamagishi, M. Genetic analysis of floral anthocyanin pigmentation traits in asiatic hybrid lily using molecular linkage maps. Theor. Appl. Genet. 2002, 105, 1175–1182. [Google Scholar] [CrossRef]
- Sun, L.; Yang, W.; Zhang, Q.; Cheng, T.; Pan, H.; Xu, Z.; Zhang, J.; Chen, C. Genome-wide characterization and linkage mapping of simple sequence repeats in mei (prunus mume sieb. Et zucc.). PLoS ONE 2013, 8, e59562. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Luo, L.; Pan, H.; Guo, X.; Wan, H.; Zhang, Q. Filling gaps with construction of a genetic linkage map in tetraploid roses. Front. Plant Sci. 2014, 5, 796. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; van Silfhout, A.; Shahin, A.; Egberts, R.; Beers, M.; van der Velde, A.; van Houten, A.; van Tuyl, J.M.; Visser, R.G.F.; Arens, P. Genetic mapping and qtl analysis of botrytis resistance in gerbera hybrida. Mol. Breed. 2017, 37, 13. [Google Scholar] [CrossRef]
- Du, Q.; Pan, W.; Xu, B.; Li, B.; Zhang, D. Polymorphic simple sequence repeat (ssr) loci within cellulose synthase (ptocesa) genes are associated with growth and wood properties in populus tomentosa. New Phytol. 2013, 197, 763–776. [Google Scholar] [CrossRef]
- Behrend, A.; Borchert, T.; Spiller, M.; Hohe, A. Aflp-based genetic mapping of the “bud-flowering” trait in heather (calluna vulgaris). BMC Genet. 2013, 14, 64. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.M.; Feng, F.; Sui, X.; Han, S. Genetic linkage maps of pinus koraiensis sieb. Et zucc. Based on aflp markers. Afr. J. Biotechnol. 2010, 9, 5659–5664. [Google Scholar]
- Haanstra, J.P.W.; Wye, C.; Verbakel, H.; Meijer-Dekens, F.; van den Berg, P.; Odinot, P.; van Heusden, A.W.; Tanksley, S.; Lindhout, P.; Peleman, J. An integrated high-density rflp-aflp map of tomato based on two lycopersicon esculentum×l. Pennellii f2 populations. Theor. Appl. Genet. 1999, 99, 254–271. [Google Scholar] [CrossRef]
- Lu, J.-J.; Zhao, H.-Y.; Suo, N.-N.; Wang, S.; Shen, B.; Wang, H.-Z.; Liu, J.-J. Genetic linkage maps of dendrobium moniliforme and d. Officinale based on est-ssr, srap, issr and rapd markers. Sci. Hortic. 2012, 137, 1–10. [Google Scholar] [CrossRef]
- Khush, G.S. Green revolution: Preparing for the 21st century. Genome 1999, 42, 646–655. [Google Scholar] [CrossRef]
- Kenis, K.; Keulemans, J. Study of tree architecture of apple (malus × domestica borkh.) by qtl analysis of growth traits. Mol. Breed. 2007, 19, 193–208. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Q.; Cheng, T.; Yang, W.; Pan, H.; Zhong, J.; Huang, L.; Liu, E. High-density genetic map construction and identification of a locus controlling weeping trait in an ornamental woody plant (prunus mume sieb. Et zucc). DNA Res. 2015, 22, 183–191. [Google Scholar] [CrossRef]
- Du, Q.; Yang, X.; Xie, J.; Quan, M.; Xiao, L.; Lu, W.; Tian, J.; Gong, C.; Chen, J.; Li, B.; et al. Time-specific and pleiotropic quantitative trait loci coordinately modulate stem growth in populus. Plant Biotechnol. J. 2019, 17, 608–624. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Trait | Mean | Max | Min | SD | Variance | Skewness | Kurtosis | CV |
|---|---|---|---|---|---|---|---|---|
| Plant height/cm | 31.1 | 72.0 | 8.1 | 11.4 | 130.8 | 0.39 | 0.7 | 36.70% |
| Crown width/cm | 44.1 | 86.0 | 10.3 | 16.6 | 274.1 | −0.4 | 0.1 | 37.6% |
| Primary lateral branch height/cm | 6.5 | 22.0 | 0.2 | 4.6 | 21.2 | 1.74 | 1.47 | 70.80% |
| Internode length/mm | 22.5 | 34.6 | 8.0 | 5.1 | 26.0 | 0.41 | −0.24 | 22.70% |
| Linkage Group | Markers Number | Number of Gaps Greater than 20 cm | Total (cm) | Max (cm) | Min (cm) | Average (cm) |
|---|---|---|---|---|---|---|
| LG1 | 66 | 0 | 167.2 | 5.8 | 0.4 | 2.53 |
| LG2 | 8 | 0 | 27.9 | 8.4 | 1.1 | 3.49 |
| LG3 | 10 | 0 | 35.9 | 7.4 | 1.4 | 3.59 |
| LG4 | 44 | 0 | 205.0 | 12.9 | 1.3 | 4.66 |
| LG5 | 5 | 0 | 16.1 | 5.7 | 2.4 | 3.22 |
| LG6 | 7 | 0 | 27.8 | 8.8 | 3.1 | 3.97 |
| LG7 | 11 | 0 | 41.0 | 8.6 | 2.1 | 3.73 |
| LG8 | 4 | 0 | 23.2 | 10.6 | 3.4 | 5.80 |
| LG9 | 3 | 0 | 21.7 | 18.2 | 3.5 | 7.23 |
| LG10 | 7 | 0 | 38.7 | 17.5 | 1.3 | 5.53 |
| LG11 | 4 | 0 | 30.3 | 19.4 | 5.3 | 7.58 |
| LG12 | 6 | 0 | 28.6 | 14.1 | 1.0 | 4.77 |
| LG13 | 18 | 0 | 138.1 | 15.1 | 2.1 | 7.67 |
| LG14 | 3 | 0 | 23.5 | 16.2 | 6.9 | 7.83 |
| LG15 | 8 | 1 | 105.8 | 25.9 | 7.38 | 13.23 |
| LG16 | 6 | 1 | 72.8 | 28.3 | 9.4 | 12.13 |
| LG17 | 16 | 0 | 124.8 | 17.0 | 2.2 | 7.80 |
| LG18 | 3 | 1 | 34.4 | 25.2 | 9.2 | 11.47 |
| LG19 | 3 | 0 | 23.1 | 15.9 | 7.2 | 7.70 |
| LG20 | 7 | 0 | 48.4 | 12.6 | 3.9 | 6.91 |
| LG21 | 2 | 1 | 22.1 | 22.1 | 22.1 | 11.05 |
| LG22 | 2 | 0 | 15.6 | 15.6 | 15.6 | 7.80 |
| Average | 11 | 0.75 | 57.8 | 15.1 | 5.1 | 6.80 |
| Trait | QTL Loci | Linkage Group | Marker Interval | Position | LOD Threshold | Variation Explained |
|---|---|---|---|---|---|---|
| Plant height | qPH-1-1 | LG1 | M19E44-251-M19E45-181 | 38.2 | 4.2 | 32.36 |
| qPH-4-1 | LG4 | M52E37-319-M17E37-116 | 84.6 | 3.2 | 16.18 | |
| qPH-17-1 | LG17 | YYJ-1134-YYJ-690 | 83.2 | 2.9 | 12.73 | |
| Crown width | qCW-4-1 | LG4 | M19E34-138-M19E34-292 | 23.1 | 3.6 | 18.07 |
| Primary lateral branch height | qPLBH-1-1 | LG1 | M19E44-251-M19E45-181 | 40.9 | 3.4 | 20.59 |
| qPLBH-7-1 | LG7 | M52E38-116-M52E38-162 | 20.7 | 2.9 | 15.34 | |
| Internode length | qIL-1-1 | LG1 | M25E35-209-M51E34-364 | 123.8 | 3.1 | 14.86 |
| qIL-20-1 | LG20 | YYJ-1062-SSR24 | 22.6 | 2.5 | 9.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Ye, Y.; Feng, L.; Zhang, Y.; Lin, Q.; Liu, J.; Cai, M.; Wang, J.; Cheng, T.; Zhang, Q.; et al. A Genetic Linkage Map of BC2 Population Reveals QTL Associated with Plant Architecture Traits in Lagerstroemia. Forests 2021, 12, 322. https://doi.org/10.3390/f12030322
Zhou Y, Ye Y, Feng L, Zhang Y, Lin Q, Liu J, Cai M, Wang J, Cheng T, Zhang Q, et al. A Genetic Linkage Map of BC2 Population Reveals QTL Associated with Plant Architecture Traits in Lagerstroemia. Forests. 2021; 12(3):322. https://doi.org/10.3390/f12030322
Chicago/Turabian StyleZhou, Yang, Yuanjun Ye, Lu Feng, Ye Zhang, Qifang Lin, Jieru Liu, Ming Cai, Jia Wang, Tangren Cheng, Qixiang Zhang, and et al. 2021. "A Genetic Linkage Map of BC2 Population Reveals QTL Associated with Plant Architecture Traits in Lagerstroemia" Forests 12, no. 3: 322. https://doi.org/10.3390/f12030322
APA StyleZhou, Y., Ye, Y., Feng, L., Zhang, Y., Lin, Q., Liu, J., Cai, M., Wang, J., Cheng, T., Zhang, Q., & Pan, H. (2021). A Genetic Linkage Map of BC2 Population Reveals QTL Associated with Plant Architecture Traits in Lagerstroemia. Forests, 12(3), 322. https://doi.org/10.3390/f12030322