Abstract
Control of seed germination and dormancy is important in seed plant adaptation and evolution. When studying seed dormancy of Quercus species, we observed a substantially delayed shoot emergence following a fast root emergence in Quercus chungii F.P.Metcalf. Since epicotyl physiological dormancy (PD) has not been reported in Quercus section Cyclobalanopsis, we examined seed morphology and germination in Q. chungii and aimed to document epicotyl PD in the seeds. The embryo was fully developed in fresh ripe seeds. The elongating cotyledonary petiole pushed the embryo axis out of the seed during germination, which differed from observations in other Quercus species. Shoots emerged from seeds with developing roots after 3 months of warm stratification (35/25 °C), reaching the highest percentage of shoot emergence in seeds after 5 months. Seeds were recalcitrant and displayed a yet unreported epicotyl PD type, for which we propose the formula Cnd(root) ‒ Cp’’ 2b(shoot). Early emergence and development of the root system in Q. chungii seeds with epicotyl PD appears to be a mechanism to maintain a constant water supply to the shoot during plumule development and emergence. Our documentation of seed germination will provide guidance for the conservation and restoration of this species from seeds.
1. Introduction
Seeds are responsible for perpetuating the species genome. Seeds represent the most vulnerable stage in the life cycle of many plant species [1,2]. In general, the environmental sensitivity provided by dormancy appears to be a key trait in the diversification of seed plants. Indeed, seed dormancy is critical for mediating the interaction between the environment and vulnerable seedlings [3]. Thus, the accurate identification of physical and physiological dormancy mechanisms occurring in seeds is the basis for understanding their adaptation and evolution.
Quercus L. (oaks, ca 430 species) is one of the most widely distributed woody genera in the northern hemisphere, consisting of two subgenera and eight sections [4,5]. Currently, 41% of oaks are considered species of conservation concern [6]. The dispersal units of Quercus species are the fruits (acorns) that are reportedly recalcitrant, except for one, which was categorized as intermediate [7,8,9]. The knowledge of seed germination and dormancy mechanisms in oaks is fundamental to our understanding of their natural regeneration and our ability to ensure their conservation by implementing efficient nursery practices for producing seedlings [10]. Quercus species can be grouped according to the types of seed dormancy: (1) the seeds in Quercus section Lobatae (red oaks) and Protobalanus commonly undergo physiological dormancy (PD), requiring a period of cold stratification to promote rapid and uniform germination; (2) most species in section Quercus (white oaks), Virentes, and Ilex produce nondormant seeds; (3) in the seeds of some species of section Quercus and Cerris, the radicle is nondormant, whereas the epicotyl undergoes PD, requiring cold stratification for both breaking the dormancy and germination; (4) the seed dormancy types in section Cyclobalanopsis are still unclear (Table 1) [11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39].

Table 1.
Seed dormancy classes in different Quercus subgenera and sections.
There have been many studies on seed germination in Quercus section Cyclobalanopsis, but most of them failed to follow the standard protocol [12,13,14]. To facilitate the dormancy classification, Baskin and Baskin (2014) suggested focusing on the following criteria: (1) embryo morphology, (2) whether the embryo grows prior to germination, (3) water-permeability of the dispersal unit, (4) ability (or lack thereof) of seeds to germinate within approximately four weeks, and (5) whether the radicle and the shoot emerge at more or less the same time, or whether the shoot emergence is delayed by several weeks after the radicle emergence [34] (p. 41). However, the water-permeability of the dispersal unit and the time gap between radicle and shoot emergence are often overlooked [40,41,42].
Quercus chungii F. P. Metcalf (Cyclobalanopsis chungii in Flora of China) is a timber oak species endemic to South China (Jiangxi, Fujian, Hunan, Guangdong, Guangxi etc.) at elevations ranging from 200 to 800 m a.s.l., and evergreen trees, up to 15 meters high [43]. Owing to the high quality of the wood, natural forest resources of Q. chungii have suffered overexploitation [15,44]. The seeds of Q. chungii are difficult to store while their germination is easily triggered, and there are difficulties to successfully establish the seedlings [44,45,46]. We failed to determine the dormancy class for Q. chungii seeds based on the published research because only the radicle germination has been examined, whereas the shoot emergence has been neglected (Table 1) [15]. Due to the difficulty of natural renewal of Q. chungii, it was hypothesized that the seeds of this species undergo epicotyl dormancy. Furthermore, it is necessary for seeds to be tested across a range of germination temperatures to elucidate ecological attributes of seed dormancy break and germination requirements.
The primary objectives of this study were to identify the dormancy class for Q. chungii acorns by determining their dormancy break and germination requirements, and to test the effect of temperature on shoot emergence. The results were expected to provide the knowledge base for explaining the seedling developmental patterns and their effect on the reproductive success of Q. chungii.
2. Materials and Methods
2.1. Sample Site and Seed Collection
Acorns of Q. chungii were collected from ten trees in one population of Gaotai Mountain (24.703N, 116.134E, Jiao-Ling County, Guangdong Province, China) on October 26, 2014, at an elevation of 211 m a.s.l. The area is a subtropical evergreen broad-leaved forest where the minimum and maximum temperatures are 10 and 35 °C in January and July, respectively. The annual mean temperature is 21 °C, and the mean annual precipitation is approximately 1500 mm, mostly recorded between June and September [45]. We directly sent the collected acorns to Shanghai Chenshan Botanical Garden to start the experiment. Full and robust acorns were chosen by flotation and disinfected with 1% potassium permanganate solution for 30 min, then washed three times with distilled water. The size and weight of 30 selected acorns were measured. Several acorns were used for anatomical observation. Some of the acorns were directly used in the seed germination experiment. The remaining acorns were enclosed to zip lock bags and stored at seed cryogenic storage cabinet with the temperature of 4 °C.
2.2. Fruit and Seed Size and Mass
The dimensions (breadth and length) of 30 acorns (fruits) and seeds (pericarps were removed manually) were measured using a Vernier caliper (Guanglu, Guilin, China). Then, both acorns and seeds were weighed, oven-dried for 2 days at 103 °C, and weighed again to obtain a constant mass.
2.3. Morphological and Anatomical Analysis
Photographs were taken of acorns and seeds (Camera: Nikon D90, Tokyo, Japan). Acorns were dissected lengthwise with a razor blade along the long axis, and the seed embryos were observed and photographed. The embryo and seed length were measured in longitudinally sectioned seeds, and the embryo axis to embryo (Ea:Em) ratio was calculated [47]. Photographs were taken of the germinating seeds every week until full seedling development.
2.4. Cotyledonary Petiole Growth and Root Emergence
The Q. chungii acorns started germinating rapidly, even during the transportation from the field to the laboratory [48]. We used room temperature (22 °C) to test the uniformity of the time for germination initiation. Three batches of 20 acorns each were incubated as replicates at room temperature for 30 days. Acorns were planted in seedling bags (25 × 20 cm) filled with vermiculite. The development and elongation of the cotyledonary petiole and root were investigated using temperature-controlled incubators set at 15/5, 20/10, and 25/15°C temperature regimes under a uniform 12/12 h light/dark (day/night) regime; the photoperiod coincided with the 12 h high-temperature half of the diurnal cycle. Each experiment included 8 acorns as replicates.
Incubated acorns were checked at 2-d intervals for 28 d. The length of the cotyledonary petiole plus the root (LoCR) was measured with a Vernier caliper. The seeds incubated at 25/15 were only measured for 16 d because their roots grew too long, and it was difficult to take them out for measurement. The length of the cotyledonary petiole (LoC) was measured from the cotyledon to the swollen part of the root (Figure 1E,F). The length of the root (LoR) was equal to LoCR minus LoC.

Figure 1.
Fruit and seed morphology photographs of Q. chungii seed germination. A, fruit; B, seed; C,D, start of seed germination (C, one seed in one fruit; D, two seeds in one fruit); E, morphology of seeds; F, elongation of cotyledonary petiole and embryo axis (red circle); G, seed morphology after cotyledonary petiole had protruded through the fruit coat; H, shoot germination; I, seedling. In each panel, a separate scale bar of 1 cm is shown.
2.5. Effect of Temperature on the Release of Shoot Dormancy in Root Emerged Seeds
To ensure the uniformity of germination conditions, we cultivated 800 seeds at room temperature (20–25 °C) until the cotyledonary petiole plus root growth reached more than 30 mm in length. Then, seeds were transferred to one of five different temperature regimes: 15/5, 20/10, 25/15, 30/20, or 35/25 °C (12/12 h), under a 12 h photoperiod. Three replicates consisting of 20 acorns each were used per treatment. Shoot germination was evaluated weekly over the first two months and once per month thereafter, until the 8th month.
After 8 months of incubation, no shoot emergence was observed in the 15/5 or 20/10 °C temperature treatments. At the beginning of the 9th month, seeds were transferred to an incubator set at 30/20 °C. Shoot germination was evaluated weekly during the first month. Then, it was evaluated once per month until the 12th month.
2.6. Statistical Analysis
Mean (±SE) fruit size, fruit weight, seed mass, seed weight, LoCR, LoC, LoR, and germination percentage were calculated. Root and shoot emergence percentages were determined using the following equations:
Root emergence percentage (%) = (number of germinating seeds/number of seeds per sample) × 100
Shoot emergence percentage (%) = (number of emerged shoots/number of emerged radicles) × 100
Finally, Tukey’s HSD test and t test were used to test for the effects of different treatments on seed germination.
3. Results
3.1. Fruit Morphology during Seed Germination
Fruits of Q. chungii were generally oblatoid, composed of a woody epicarp, a brown fibrotic endocarp, and one or two seeds (Figure 1A–D). The Q. chungii embryo filled the whole inside of the acorn (Figure 1B,D). The embryo axis was small, spherosome–shaped, and inserted into the cotyledonary petiole near the germination pore (Figure 1E,F). The cotyledonary petiole was found adjacent to the seed coat and inserted into the cotyledon (Figure 1E–I). The cotyledonary petiole carrying the embryo axis would break through the pericarp at the junction between the main pericarp and the scar. Germination was detected as the protrusion of the cotyledonary petiole overcoming the resistance of the adjacent tissues and dislocating the pericarp (Figure 1C,D). Dislodgment of the pericarp was caused by the elongation of the cotyledonary petiole (Figure 1C,D). The root began to emerge through the distal end of the petiole and formed an intumescent tubular structure (Figure 1C,D,G,H). Under appropriate conditions, the shoot emerged from the tubular structure (Figure 1H, I).
3.2. Quantitative Characteristics of Fruit and Seeds
The fruits of Q. chungii were 18.81 ± 1.39 mm in length and 16.13 ± 1.42 mm in width. Their calculated length width ratio was 1.17. Seeds were 15.63 ± 1.21 mm in length and 13.66 ± 1.19 mm in width with a calculated length width ratio of 1.14. The embryo axis was 1.52 mm in length and 1.21 mm in width, and its calculated length width ratio was 1.26. The Ea:Em ratio was 0.10. The estimated volume ratio of embryo and seed was 0.08%. The mean fruit fresh and dry weights were 2.91 ± 0.70 g and 1.74 ± 0.34 g, respectively. Seeds had a fresh weight of 2.03 ± 0.42 g and a dry weight of 1.21 ± 0.24 g. By calculation, the moisture content of acorn and seed was 40.20% and 40.39%.
3.3. Effects of Temperature on Root Emergence
Seed germination started 4 days after sowing, at which time 40% of the seeds had started germinating. Eight days after sowing, the percentage of germinating seeds reached 85% (Figure 2). The mean length of cotyledonary petioles was 24.84 ± 3.40 mm. When the length of the cotyledonary petiole plus the root exceeded 25 mm, the emerging radicle was visible (Figure 1). There were significant differences in the meantime of root emergence among temperature regimes. Root emergence occurred at 4, 9, and 16 days after the start of germination under the 25/15, 20/10, and 15/5 °C temperature regimes, respectively (Figure 3). There was no gap between the growth of the cotyledonary petiole and that of the radicle. The root grew faster under the 25/15 °C regime than under the other two regimes (Figure 4). Under the 20/10 and 15/5 °C regimes, the root grew very slowly, and the root length did not significantly change within one week (Figure 4).
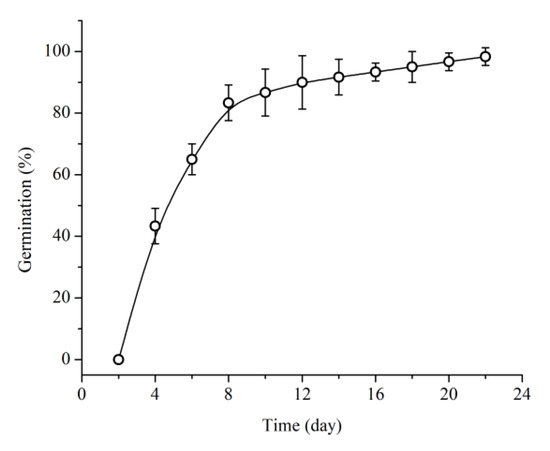
Figure 2.
Percentage of cumulative germination of Quercus chungii seeds initiating germination at room temperature.
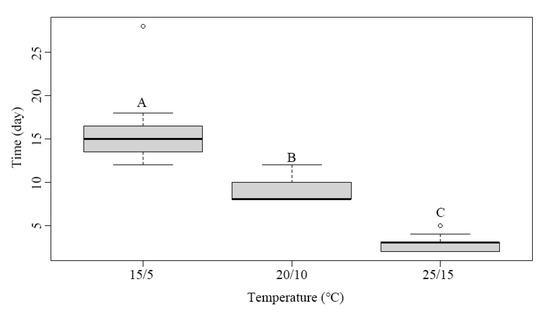
Figure 3.
Mean time (days) needed for root emergence under different temperature conditions. Different uppercase letters indicate significant differences between the incubation temperature regimes (Tukey’s HSD test, p = 0.05).
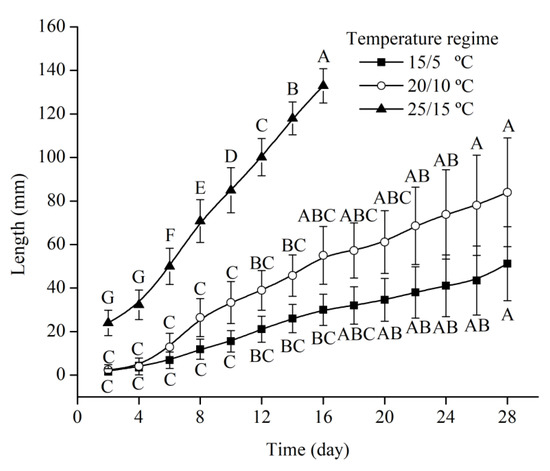
Figure 4.
Length of elongated cotyledonary petiole and root of Quercus chungii under three temperature regimes. Different uppercase letters indicate significant differences between the different time points under the same incubation temperature regime (Tukey’s HSD test, p = 0.05).
3.4. Effects of Temperature on Epicotyl Dormancy Release in Seeds with the Emerging Radicle
Shoot emergence did not occur under the 20/10 and 15/5 °C temperature regimes during 8 months of incubation, while it started under the 35/25 °C regime after 2 months of incubation. After 3 months of incubation, the percentages of shoot emergence were significantly higher under the 35/25 and 30/20 °C regimes than under the other conditions. The shoot emergence was greater than 60% under the 25/15, 30/20, and 35/25 °C regimes after 4 months of incubation. There was no significant difference among these three temperature regimes from 4 to 8 months of incubation. The percentage of shoot emergence was nearly or higher than 80% after 5 months of incubation (Table 2).
Table 2.
Percentage of cumulative shoot emergence (mean ± SD) in Quercus chungii seeds under different temperature regimes.
Shoot emergence in seeds transferred from the 20/10 °C to the 30/20 °C regime nearly reached 30% after 1 month, and it was significantly higher than in seeds transferred from the 15/5 °C to the 30/20 °C regime. Shoot emergence in seeds transferred from 15/5 °C or 20/10 °C to 30/20 °C was more than 66% or 80%, respectively, after 4 months of incubation (Table 3).
Table 3.
Percentage of cumulative shoot emergence (mean ± SD) of Quercus chungii transferred to 30/20 °C after 8 months under 15/5 °C or 20/10 °C conditions.
4. Discussion
The acorns of Quercus species consist of a single seed contained within a hard pericarp [9]. In Q. chungii, there are occasionally two seeds in one acorn (Figure 1D). During field surveys over the last decade, we observed this phenomenon in many oak species; indeed, we found it particularly widespread among species that inhabit alpine environments (e.g., Quercus pannosa). In most Quercus species, the embryo axis (germination pore or hypocotyl root axis) sits at the apex region underneath the remaining stylar parts of the pericarp (apex) [9,39,49], whereas in Q. chungii, the embryo axis points to the opposite direction that sits at the scar region (Figure 1C,D).
Fruit structure plays an important role in protecting seeds from predation and against physiological alterations caused by storage [50,51]. The germination pore is the most critical part of the fruit because death of the embryo axis due to desiccation prevents germination [9]. The germination pore of Q. chungii fruits resides at the scar (Figure 1C), which is a key route for water flow into and out of the fruit in Quercus [9]. Thus, the embryo axis of Q. chungii fruits may easily die because of rapid desiccation. An assessment of the relationship between the storage process and seed germination of Q. chungii fruits suggested that they are recalcitrant [48], which we confirmed in our study based on the moisture content of fresh seeds. This is consistent with reports about other Quercus species indicating that Quercus seeds are recalcitrant [7]. Thus, recalcitrant properties of Quercus seeds are a common phenomenon.
If the seed has a differentiated and fully developed embryo, it indicates that there is no morphological seed dormancy [34] (p. 43). All the acorns of Quercus have fully developed embryos filling the whole seeds, which contain two big cotyledons and the embryo axis [9]. A morphological analysis of the Q. chungii fruit identified the fully developed embryo, in which the embryo axis is much smaller than the cotyledons. It indicates that morphological dormancy does not exist in Quercus species.
Along with Q. chungii, other members of Quercus section Cyclobalanopsis, e.g., Q. fleuryi, Q. glauca, and Q. multinervis, are common species in subtropical evergreen broad-leaved forests in East and South China [43]. However, there is not much information available on the germination of these species. According to Xia et al. (2015), Q. fleuryi, Q. glauca, and Q. multinervis seeds germinate (root emergence) very slowly. The time to reach 50% germination is at least 40 days, and typically, it takes 2–3 months (Table 1) [12]. In contrast, Wang et al. (2009) observed that the root emergence of Q. chungii is very fast, occurring even during harvesting and transportation [48]. This study confirmed in laboratory experiments that Q. chungii seeds were nondormant for the radicle. Early germination of white oak (Quercus section Quercus) seeds is widely viewed as an evolutionary strategy to escape rodent predation [52,53]. Q. chungii seeds have the same pattern of root emergence as white oak seeds, but it differs from that in the seeds of other species of the Quercus section Cyclobalanopsis in East and South China, implying a different ecological adaptive strategy for the local environment and the rodent predation. Firstly, the nondormant root in Q. chungii seeds appears to be related to the position of the embryo axis and rapid water loss (i.e., desiccation). Secondly, similar to that in white oaks, the rapid root emergence minimizes the risk of predation by rodents.
During the onset of seed germination in Q. chungii, we observed a special structure termed the elongated cotyledonary petiole. This normally occurring structure has been well studied in palms that have differentiated, underdeveloped embryos [51,54,55,56]. However, the elongated cotyledonary petiole in the fully developed seed was only reported in Q. virginiana more than one hundred years ago [39]. Our study reports the second example in Q. chungii belonging to the same genus. At the initiation of germination, the elongation of the cotyledonary petiole pushes the embryo axis on the outside of the seed to an approximate length of 25 mm. Then, the radicle begins to develop from the tip of the cotyledonary petiole. Finally, the shoot emerges from the juncture between cotyledonary petiole and the taproot base. Typically, there are two short cotyledonary petioles matching the pair of cotyledons [53,57]. However, the elongated cotyledonary petiole of Q. chungii is one tubular structure. Thus, we observed hypogeal and remote-tubular germination in Q. chungii, which was recorded for the first time in Quercus.
However, there was a significant delay in shoot emergence after the radicle emerged in the germination experiments. The shoot remained dormant for at least 2 months after radicle emergence under laboratory light and temperature conditions. The delayed shoot emergence clearly demonstrated that Q. chungii has a type of epicotyl physiological dormancy (PD). Epicotyl PD has been reported in three different sections of Quercus, (1) Q. alba and Q. prinus in section Quercus, (2) Q. ilicifolia in section Lobatae, and (3) Q. suber in section Cerris (Table 1). Epicotyl PD has also been described in several other woody families of Clusiaceae (Platonia insignis Mart.) [58,59], Fabaceae (Brownea coccinea Jacq., Cynometra cauliflora Linn. Attenuates, and Humboldtia laurifolia Vahl) [60,61], Lecythidaceae (Lecythis ampla Miers) [62], and Oleaceae (Chionanthus retusus Lindl.et Paxt.) [63,64].
Which type of epicotyl PD occurs in Q. chungii seeds? In temperate white oaks, the root is ND, whereas the epicotyl PD requires a long cold period to break deep PD; Baskin and Baskin (2008 and 2014) [34,65] defined the dormancy type and suggested the formula Cnd(root) ‒ C3a(shoot). Unlike in these temperate oaks, the epicotyl in seeds of Q. chungii require a warm stratification (3–5 months) in spring and/or early summer to trigger shoot emergence. Compared with the other formula of epicotyl PD in Clusiaceae [58,59], Fabaceae [60,61], Lecythidaceae [62], and Oleaceae [63,64] summarized by Baskin and Baskin (2014) [34] (p. 61), we suggest a new formula, Cnd(root)‒Cp’’ 2b(shoot), to describe dormancy in seeds of Q. chungii. In the formula, C is PD; nd, nondormant; 2, intermediate PD; 3, deep PD; a, cold treatment is required; b, warm treatment is required; p’’, plumule growth occurs outside the seed (the force of elongated cotyledonary petiole), and there is a delay in the start of growth after root emergence. We further suggest that this type of seed dormancy should be considered a subtype of the epicotyl PD.
5. Conclusions
Quercus chungii is mainly distributed in the montane lowland forests of East and South China, dominated by the subtropical monsoon system with a distinct wet summer season and a dry winter season. Recalcitrant seed survival and germination depend on a constant water supply to the embryo [34]. Perhaps early emergence and development of the root system in Q. chungii with epicotyl PD is a mechanism to maintain a constant water supply to the shoot during plumule development and emergence.
Quercus chungii is an important commercial woody species in East China. The distribution area of Q. chungii dropped sharply during the last century, and it has been treated as a rare and endangered species in the Fujian province [46]. Thus, the information on seed germination of this species in our study will be of considerable value to conservation biologists and restoration ecologists who want to propagate the species from seeds.
Author Contributions
Conceptualization, Y.-G.S. and X.-L.D.; methodology, X.-Q.S. and Y.-G.S.; software, X.-Q.S. and B.-J.G.; formal analysis, X.-Q.S.; investigation, Y.-G.S.; resources, Y.-G.S. and B.-J.G.; data curation, X.-Q.S. and Y.-G.S.; writing—original draft preparation, X.-Q.S. and Y.-G.S.; writing—review and editing, X.-L.D. and G.K.; visualization, B.-J.G.; supervision, Y.-G.S. and X.-L.D.; project administration, Y.-G.S. and G.K.; funding acquisition, Y.-G.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant number 31901217 and Shanghai Municipal Administration of Forestation and City Appearances, grant number G142424.
Acknowledgments
We thank Carol C. Baskin for pointing out the significant error for the first version of this manuscript. We also thank Cheng Du for his help on how to use the Latin name correctly.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hanley, M.E.; Lamont, B.B.; Fairbanks, M.M.; Rafferty, C.M. Plant structural traits and their role in anti-herbivore defence. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 157–178. [Google Scholar] [CrossRef]
- Keeley, J.E. Seed germination and life history syndromes in the California chaparral. Bot. Rev. 1991, 57, 81–116. [Google Scholar] [CrossRef]
- Willis, C.G.; Baskin, C.C.; Baskin, J.M.; Auld, J.R.; Venable, D.L.; Cavender-Bares, J.; Donohue, K.; De Casas, R.R. The NESCent Germination Working Group The evolution of seed dormancy: Environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytol. 2014, 203, 300–309. [Google Scholar] [CrossRef]
- Denk, T.; Grimm, G.W.; Manos, P.S.; Deng, M.; Hipp, A.L. An Updated Infrageneric Classification of the Oaks: Review of Previous Taxonomic Schemes and Synthesis of Evolutionary Patterns. In Oaks Physiological Ecology. Exploring the Functional Diversity of Genus; Quercus, L., Gil-Pelegrín, E., Peguero-Pina, J.J., Sancho-Knapik, D., Eds.; Springer: Gewerbestrasse, Switzerland, 2017; Volume 7, pp. 13–38. [Google Scholar]
- Fazan, L.; Song, Y.-G.; Kozlowski, G. The Woody Planet: From Past Triumph to Manmade Decline. Plants 2020, 9, 1593. [Google Scholar] [CrossRef]
- Carrero, C.; Jerome, D.; Beckman, E.; Byrne, A.; Coombes, A.J.; Deng, M.; Rodriguez, A.G.; Sam, H.V.; Khoo, E.; Nguyen, N.; et al. The Red List of Oaks 2020; The Morton Arboretum: Lisle, IL, USA, 2020; pp. 11–12. [Google Scholar]
- Royal Botanic Gardens Kew, Seed Information Database (SID). Available online: http://data.kew.org/sid (accessed on 30 April 2020).
- Xia, K.; Daws, M.I.; Hay, F.R.; Chen, W.-Y.; Zhou, Z.-K.; Pritchard, H.W. A comparative study of desiccation responses of seeds of Asian Evergreen Oaks, Quercus subgenus Cyclobalanopsis and Quercus subgenus Quercus. S. Afr. J. Bot. 2012, 78, 47–54. [Google Scholar] [CrossRef]
- Xia, K.; Daws, M.I.; Stuppy, W.; Zhou, Z.-K.; Pritchard, H.W. Rates of Water Loss and Uptake in Recalcitrant Fruits of Quercus Species Are Determined by Pericarp Anatomy. PLoS ONE 2012, 7, e47368. [Google Scholar] [CrossRef]
- Dey, D.C. Sustaining Oak Forests in Eastern North America: Regeneration and Recruitment, the Pillars of Sustainability. For. Sci. 2014, 60, 926–942. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Q.M.; Liu, G.Q.; Liu, Y.; Hou, L.Y.; Li, G.L. Inhibitory mechanism of seed germination of Quercus acutissima. Sci. Silva. Sin. 2012, 48, 164–170. [Google Scholar]
- Xia, K.; Daws, M.I.; Zhou, Z.-K.; Pritchard, H.W. Habitat-linked temperature requirements for fruit germination in Quercus species: A comparative study of Quercus subgenus Cyclobalanopsis (Asian evergreen oaks) and Quercus subgenus Quercus. S. Afr. J. Bot. 2015, 100, 108–113. [Google Scholar] [CrossRef]
- Merouani, H.; Branco, C.; Almeida, M.H.; Pereira, J.S. Effects of acorn storage duration and parental tree on emergence and physiological status of Cork oak (Quercus suber L.) seedlings. Ann. For. Sci. 2001, 58, 543–554. [Google Scholar] [CrossRef]
- Li, Q.S.; Chen, J.; Deng, M.; Zhou, C.L.; Shen, J. Seed germination and seedling establishment of the endangered oak Quercus austrocochinchinensis. Seed 2016, 35, 4–12. [Google Scholar]
- Huang, Y.R.; Zhuang, K.; Wu, P.F.; Ma, X.Q.; Lai, X.L.; Tang, W.M. Seed germination and growth characteristics of Cyclobalanopsis chungii. Chinese J. Ecol. 2017, 36, 1251–1258. [Google Scholar]
- Zhou, Y. Seed germination of Cyclobalanopsis glaucoides. J. Plant Physiol. 2003, 39, 325–326. [Google Scholar]
- Chen, X.B.; Zhang, X. Effects of storage temperature on seed germination and emergence of Quercus aquifolioides. Guizhou Agric. Sci. 2015, 43, 129–131. [Google Scholar]
- Rao, P.B. Effects of Environmental Factors on Germination and Seedling Growth in Quercus floribunda and Cupressus torulosa, Tree Species of Central Himalaya. Ann. Bot. 1988, 61, 531–540. [Google Scholar] [CrossRef]
- Rao, P.B. Effect of temperature on seed germination of certain woody species of central Himalaya. Adv. Biosci. 1992, 11, 41–52. [Google Scholar]
- Viswanath, S.; Singh, R.P.; Thapliyal, R.C. Seed germination patterns in a Himalayan moist temperate forest. Trop. Ecol. 2002, 43, 265–273. [Google Scholar]
- Saklani, K.P.; Singh, B.; Bhatt, B.P. Influence of altitude on seed and seedling characteristics in Quercus leucotrichophora A. Camus. ex. Bahadur. Silvae Genet. 2012, 61, 36–43. [Google Scholar] [CrossRef]
- Griffin, J.R. Oak Regeneration in the Upper Carmel Valley, California. Ecology 1971, 52, 862–868. [Google Scholar] [CrossRef]
- Matsuda, K.; McBride, J.R. Germination Characteristics of Selected California Oak Species. Am. Midl. Nat. 1989, 122, 66. [Google Scholar] [CrossRef]
- Nyandiga, C.O.; McPherson, G.R. Germination of two warm-temperate oaks, Quercusemoryi and Quercusarizonica. Can. J. For. Res. 1992, 22, 1395–1401. [Google Scholar] [CrossRef]
- Peterson, J.K. Mechanisms Involved in Delayed Germination of Quercus nigra L. Seeds. Ann. Bot. 1983, 52, 81–92. [Google Scholar] [CrossRef]
- Hawkins, T.S. Regulating acorn germination and seedling emergence in Quercus pagoda (Raf.) as it relates to natural and artificial regeneration. New For. 2019, 50, 425–436. [Google Scholar] [CrossRef]
- Hawkins, T.S. The Influence of Dormancy Break Requirements on Germination and Viability Responses to Winter Submergence in Acorns of Three Bottomland Red Oak (Sect. Lobatae) Species. For. Sci. 2019, 65, 556–561. [Google Scholar] [CrossRef]
- Hopper, G.M.; Smith, D.W.; Parrish, D.J. Germination and seedling growth of northern red oak: Effects of stratification and pericarp removal. For. Sci. 1985, 31, 31–39. [Google Scholar]
- Olson, D.F., Jr.; Quercus, L. Oak. In Seeds of Woody Plants in the United States; Agriculture Handbook No., 450; Schopmeyer, C.S., Ed.; Forest Service: Washington, DC, USA, 1974; pp. 692–703. [Google Scholar]
- Chandler, A.F. Glacier point restoration project. Comb. Proc. Int. Plant. Prop. Soc. 2000, 50, 585–587. [Google Scholar]
- Farmer, R.E., Jr. Epicotyl dormancy in white and chestnut oaks. For. Sci. 1977, 23, 329–332. [Google Scholar]
- Liu, Q.; Lan, Q.Y.; Tan, Y.H.; Shen, Y.X.; Wen, B. A preliminary study on seed germination of dominant plants from a Karst landscape in Bijie, Guizhou. Acta Botanical Yunnanica 2010, 32, 539–546. [Google Scholar]
- Sopp, D.F.; Salac, S.S.; Sutton, P.K. Germination of Gambel oak seed. Tree Planter’s Notes 1977, 28, 4–5. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Elsevier/Academic Press: San Diego, CA, USA, 2014; pp. 41–863. [Google Scholar]
- Garrison, W.J.; Augspurger, C.K. Double- and single-seeded acorns of Bur oak (Quercus macrocarpa): Frequency and some ecologial consequences. Bull. Torrey Bot. Club 1983, 110, 154–160. [Google Scholar] [CrossRef]
- Corbineau, F.; Dacher, F.; Come, D. Effects of cold storage duration of acorns and of germination temperature on seedling development in sessile oak. Rev. For. 2001, 53, 32–43. [Google Scholar]
- Jastrzębowski, S.; Ukalska, J. Dynamics of epicotyl emergence of Quercus robur from different climatic regions is strongly driven by post-germination temperature and humidity conditions. Dendrobiology 2019, 81, 73–85. [Google Scholar] [CrossRef]
- Center, A.; Etterson, J.R.; Deacon, N.J.; Cavender-Bares, J. Seed production timing influences seedling fitness in the tropical live oak Quercus oleoides of Costa Rican dry forests. Am. J. Bot. 2016, 103, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Lewis, I.M. The seedling of Querucus virginiana. Plant World 1911, 14, 119–123. [Google Scholar]
- Atwater, B.R. Germination, dormancy and morphology of the seeds of herbaceous ornamental plants. Seed Sci. Technol. 1980, 8, 523–573. [Google Scholar]
- Joley, D.B.; Maddox, D.M.; Schoenig, S.E.; Mackey, B.E. Parameters afferting germinability and seed bank dynamics in dimorphic achenes of Centaurea solstitialis in California. Can. J. Bot. 2003, 81, 993–1007. [Google Scholar] [CrossRef]
- Timmermans, B.G.H.; Vos, J.; Van Nieuwburg, J.; Stomph, T.J.; Van Der Putten, P.E.L. Germination rates of Solanum sisymbriifolium: Temperature response models, effects of temperature fluctuations and soil water potential. Seed Sci. Res. 2007, 17, 221–231. [Google Scholar] [CrossRef]
- Huang, C.C.; Chang, Y.T.; Bartholomew, B. Fagaceae. In Flora of China; Wu, C.Y., Raven, P.H., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 1999; Volume 4, pp. 380–400. (In English) [Google Scholar]
- Lai, H.Y.; Wang, Y.F.; Wu, K.; Ma, X.Q.; Wu, P.F. Effect of exogenous hormones on germination and antioxidant enzyme activity of seeds of Cyclobalanopsis chungii. Chinese J. Ecol. 2017, 36, 382–388. [Google Scholar]
- Huang, Y.R.; Ma, X.Q.; Zhuang, K.; Liu, M.X.; Huang, D.D. Seed Rain and Soil Seed Bank of Cyclobalanopsis chungii Forest in Minqing, Fujian Province. J. Trop. Subtrop. Bot. 2010, 18, 68–74. [Google Scholar]
- Wang, Y.F.; Wu, P.F.; Wang, R.W.; Ma, X.Q.; Zhou, X.H. Community characteristics of Cyclobalanopsis chungii forest in Mingqing nature reserve. J. Fujian Agric. For. Univ. Nat. Sci. Ed. 2011, 1, 39–44. [Google Scholar]
- Vandelook, F.; Bolle, N.; Van Assche, J.A. Multiple environmental signals required for embryo growth and germination of seeds of Selinum carvifolia (L.) L. and Angelica sylvestris L. (Apiaceae). Seed Sci. Res. 2007, 17, 283–291. [Google Scholar] [CrossRef]
- Wang, Y.F.; Ma, X.Q.; Liu, A.Q.; Huang, Y.R.; Huang, J.G. Effects of storage time and environmental factors on seed germination of Cyclobalanopsis chungii. J. Southwest For. Univ. 2009, 29, 28–31. [Google Scholar]
- Hou, X.G.; Yi, X.F.; Yang, Y.Q.; Liu, W.J. Acorn germination and seedling survival of Q. variabilis: Effects of cotyledon excision. Ann. For. Sci. 2010, 67, 711. [Google Scholar] [CrossRef]
- Pinheiro, C.U.B. Germination strategies of palms: The case of Schippia concolor Burret in Belize. Brittonia 2002, 53, 519–527. [Google Scholar] [CrossRef]
- Neves, S.D.C.; Ribeiro, L.M.; Da Cunha, I.R.G.; Pimenta, M.A.S.; Mercadante-Simões, M.O.; Lopes, P.S.N. Diaspore structure and germination ecophysiology of the babassu palm (Attalea vitrivir). Flora Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 68–78. [Google Scholar] [CrossRef]
- Fox, J.F. Adaptation of gray squirrel behavior to autumn germination by white oak acorns. Evolution 1982, 36, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.F.; Yang, Y.Q.; Curtis, R.; Bartlow, A.W.; Agosta, S.J.; Steele, M.A. Alternative strategies of seed predator escape by early-germinating oaks in Asia and North America. Ecol. Evol. 2012, 2, 487–492. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. What kind of seed dormancy might palms have? Seed Sci. Res. 2014, 24, 17–22. [Google Scholar] [CrossRef]
- Ribeiro, L.M.; Oliveira, D.M.T.; Garcia, Q.D.S. Structural evaluations of zygotic embryos and seedlings of the macaw palm (Acrocomia aculeata, Arecaceae) during in vitro germination. Trees 2012, 26, 851–863. [Google Scholar] [CrossRef]
- Silva, R.S.; Ribeiro, L.M.; Mercadante-Simões, M.O.; Nunes, Y.R.F.; Lopes, P.S.N. Seed structure and germination in buriti (Mauritia flexuosa), the Swamp palm. Flora Morphol. Distrib. Funct. Ecol. Plants 2014, 209, 674–685. [Google Scholar] [CrossRef]
- Yi, X.F.; Curtis, R.; Bartlow, A.W.; Agosta, S.J.; Steele, M.A. Ability of chestnut oak to tolerate acorn pruning by rodents. Naturwissenschaften 2013, 100, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.E.U.; Muller, C.H.; Leao, N.V. Cronologia dos eventos morfologicos associados a germinacao esensibilidade ao dessecamento em sementes de bacuri (Platonia insignis Mart.—Clusiaceae). Rev. Brasil. Semen. 1998, 20, 236–240. [Google Scholar]
- Carvalho, J.E.U.; Muller, C.H.; Nascimento, W.M.O. Cronologia dos eventos morfologicos associados a germinacao esensibilidade ao dessecamento em sementes de bacuri (Platonia insignis Mart.—Clusiaceae). Rev. Brasil. Semen. 1998, 20, 475–479. [Google Scholar] [CrossRef]
- Jayasuriya, K.M.G.G.; Wijetunga, A.S.T.B.; Baskin, J.M.; Baskin, C.C. Recalcitrancy and a new kind of epicotyl dormancy in seeds of the understory tropical rainforest tree Humboldtia laurifolia(Fabaceae, Ceasalpinioideae). Am. J. Bot. 2010, 97, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Jayasuriya, K.M.G.G.; Wijetunga, A.S.T.B.; Baskin, J.M.; Baskin, C.C. Physiological epicotyl dormancy and recalcitrant storage behaviour in seeds of two tropical Fabaceae (subfamily Caesalpinioideae) species. AoB PLANTS 2012, 2012, pls044. [Google Scholar] [CrossRef] [PubMed]
- Flores, E.M. Seed Biology. In Tropical Tree Seed Manual; Vozzo, J.A., Ed.; United States Department of Agriculture, Forest Service: Washington, DC, USA, 2002; Volume 721, pp. 13–124. [Google Scholar]
- Chien, C.-T.; Kuo-Huang, L.-L.; Shen, Y.-C.; Zhang, R.; Chen, S.-Y.; Yang, J.-C.; Pharis, R.P. Storage Behavior of Chionanthus retusus Seed and Asynchronous Development of the Radicle and Shoot Apex during Germination in Relation to Germination Inhibitors, Including Abscisic Acid and Four Phenolic Glucosides. Plant Cell Physiol. 2004, 45, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Huang, I.C.; Chii, C.T.; Chen, I.Z.; Hsia, I.S. Effect of fruit maturity, seed scarification and medium composition on germination uniformity of Chionanthus retusus seeds. J. Taiwan Soc. Hort. Sci. 2009, 55, 1–12. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. Some considerations for adoption of Nikolaeva’s formula system into seed dormancy classification. Seed Sci. Res. 2008, 18, 131–137. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).