Abstract
The mechanism of how soil carbon pools change when coniferous forests are converted into conifer-broadleaf mixed forests is poorly understood. In this study, the impact of additional carbon inputs on soil organic carbon mineralization and microbial communities was evaluated. In a microcosm incubation experiment, three types of 13C-labeled litter (Pinustabulaeformis (PT), Robiniapseudoacacia (RP), and a mixture of PT and RP (1:1, PR)) were added in to top (0–20 cm) and deep (60–80 cm) soil collected from a Chinese pine plantation. The priming effect (PE) and specific microbial groups involved in PE were studied. PT and RP addition to topsoil induced a negative PE. In deep soil, the decomposition rates of soil organic matter (SOM) after adding PT and mixture increased by 16.6% and 34.6% compared to those without litter. The addition of RP with a lower C:N ratio had a stronger negative PE than adding PT or mixture. Moreover, the PE in deep soil was more intense after all litter additions. In topsoil, the litter-derived carbon was mainly incorporated into 16:0, 18:1ω9c, and 18:1ω7c fatty acids. In conclusion, the addition of broadleaf litter into coniferous plantations might be beneficial for enhancing deep soil carbon stocks.
1. Introduction
Forest ecosystems occupy approximately one-third of the world’s land area and store approximately 383 Pg of C in their soils to a depth of 1 m [1,2]. The changes in forest soil organic carbon (SOC) pool have important impacts on global carbon cycling. In recent years, global warming or the change of land use type increased the input of fresh organic carbon to soil, which might affect soil carbon dynamics. Exogenous organic matter inputs can cause rapid changes in native soil organic matter, which is described as the ‘priming effect’(PE) [3]. A positive PE can accelerate SOC decomposition and decrease the content of SOC, while a negative PE induces the opposite [4]. Plant litter quantities and qualities are crucial factors regulating SOC decomposition. Chao et al. [5] demonstrated that litter chemistry controls the magnitude and direction of the PE. Litter hemicellulose and tannin concentrations play important roles in the early stage of decomposition. Lignin and the ratio of lignin:N showed a negative correlation with the PE in the late phase of decomposition. Increasing fresh litter addition resulted in losses of soil organic carbon in agroecosystems [6]. Hick et al. [7] revealed that the complexity of the carbon substrate had a significant impact on the soil PE by affecting microbial carbon-use efficiency in montane forest and lowland soils. The conversion of land use types, such as farmland to forest, pure forest to mixed forest, and broadleaf forest to coniferous forestalter the quantities and qualities of external plant residues and thus cause changes in SOC pools. In addition, forest litter decomposition was important for nutrient return to the soil. Coniferous litter was broken down more slowly than broad leaf litter because of higher lignin concentration and C/N ratio. In recent years, litter mixing experiments (e.g., litters characterized by low and high decomposability) were carried out in order to evaluate the mixture effects on decomposition rates and soil carbon dynamics. The synergistic [8], antagonistic [9], neutral effects [10] were observed because of complex interactions among litter chemical componentand specific soil characteristics. Especially, in arid and semi-arid region, plant species might change frequently because of water shortage, which changed litter composition and chemical component. Litter decomposition rates were essential for nutrient return to soil. There was little known about how litter species diversity affects soil carbon dynamics.
Deep soils are considered a major organic carbon pool in terrestrial ecosystems, containing over half of the total soil C [11]. Small changes in the organic carbon pool in deep soils might have an important impact on the overall soil carbon dynamics and ultimately affect the global carbon cycle [12]. It was reported that theslow decomposition rate of deep SOC is primarily attributed to alack of fresh C supply [11,13]. Fresh leaf litter addition produced a positive PE on SOC mineralization [12]. Nevertheless, some studies found that excessive labile carbon input might transfer into recalcitrant carbon and neutralize some recalcitrant SOC mineralized by priming and thus be useful for soil carbon storage [14]. Litter qualities affected difference in PE through altering microbial activity in mining recalcitrant soil organic matter held nutrients [15]. Fontaine et al. [11] reported that microorganisms might relieve the impact of global warming (e.g., increase of exogenous organic matter inputs) on deep soil mineralization because of the lack of energy. Many studies were conducted in temperate forest ecosystem [16,17]. However, there was not a uniform conclusion. In particular, in arid and semi-arid area, tree species increased the root length in order to better resist droughts, which alter theinput of fresh C to deep soils. Therefore, the response of deep soil carbon pools to fresh carbon supply after the land use conversion in this region needed to be further studied.
Soil microbial communities were reported to be closely related to soil nutrient availability and regulate the direction and intensity of PEs [18]. Soil ‘microbial N mining’ and ‘preferential substrate utilization’ theory have revealed that during the soil PE, microorganisms alter their community structure and their balance of r- and K-strategists to maximize nutrient acquisition [19]. When the soil nutrient availability is low, microbes usually use new labile carbon as energy to obtain nutrients from soil organic carbon and thus induce a positive PE. Conversely, under high nutrient availability, microbes use more fresh organic matter and induce a negative PE [20]. Current studies have reported that microbial species or taxa carry out different functions based on their characteristics during the decomposition of soil organic matter. Gram-negative bacteria preferentially utilize easily decomposed substrates [21]. Gram-positive bacteria and actinomycetes were reported to be more effective at recalcitrant soil organic matter decomposition [22]. During the decomposition of plant residues, bacteria, especially r-strategists, initially mineralized labile C entering the soil [23], speeding up the turnover of bacterial biomass and inducing a PE. In the later stage of decomposition, fungi and certain groups of bacteria (e.g., K-strategists) were the key players decomposing complex substrates (e.g., lignin and cellulose). In addition, the roles of soil microbial communities are different in different ecosystems. Gram-positive bacteria and fungi are the main regulators of rhizosphere PEs, which belong to a K-strategist classification widely observed in a few temperate grassland studies [24]. Nevertheless, there was no connection between specific microbial groups and the PEs in lowland tropical forest soils [25]. Therefore, the microbial species and their contributions involved in the PE remain to be further studied.
In the present study, we want to examine the changes in soil organic carbon pools and microbial communities in the top (0–20 cm) and deep (60–80 cm) soil layers in response to the addition of different kinds of fresh organic matter by using 13C-labeled leaf litter, to understand how the conversion of coniferous forest to conifer-broadleaf forest impact soil organic matter (SOM) decomposition. We hypothesized that (1) in all detected soil layers, the addition of locust (Robinia pseudoacacia, RP) leaf litter to Chinese pine (Pinus tabulaeformis, PT) plantations would induce a negative PE and accelerate soil organic carbon storage because of the high litter quality. Mixed litter addition showed the same trend through the selective decomposition of locust leaf litter. Chinese pine litter addition would result in the opposite result. (2) The magnitude and intensity of the PE would be regulated by the activities of specific microbial groups, which would vary with the fresh organic matter availability. (3) The PE would be more intense in the topsoil layer than in the deep soil layer because of the higher microbial activity.
2. Materials and Methods
2.1. Site Description and Soil Sampling
The experimental site was located in the Huai Ping forest region, Shaanxi Province, China, which belongs to the hilly gully area of the Loess Plateau (107°56′–108°200′ E, 34°29′–34°59′ N) (Figure S1). This area features a warm temperate continental monsoon climate. The average annual rainfall is 602 mm, with over 50% falling between July and September. The annual mean temperature is 10.8 °C. The soil is cinnamon soil (Hapludalfs in the USDA classification). The soil properties are described in Table 1. The forests in this region were pure stands planted in the 1960s, which are mainly composed of Chinese pine (PT) and locust (RP). With the development of Chinese pine plantation, greater soil degradation and more pests were found in the pure forest than in a mixed forest. In recent years, broadleaf tree species (e.g., locust) have been introduced into pure forests.

Table 1.
Physicochemical properties of soil and litter used for incubation.
In September 2018, soil samples were collected from 40-year-old Chinese pine plantations. Three 20 m × 20 m plots were selected randomly, and three profiles (1 m width and 1 m depth) were placed in each plot. In each profile, soil samples were obtained from the topsoil (0–20 cm) and deep soil (60–80 cm) using a soil corer (5 cm in diameter). Samples of the same layers in each profile were mixed into a composite sample in each plot. In all, there were nine composite soil samples in the same layer in this study. Each soil sample was divided into two parts after manually removing the visible roots, stones, litter, and debris. One part was stored at 4 °C before the soil incubation experiment and analysis of the enzyme activities and microbial biomass began (about two weeks). The other part was air dried for analysis of the soil properties.
2.2. Litter Properties
The litter samples were oven-dried at 65 °C until constant weight, grinded and passed through a 0.25 mm sieve before subsequent analysis. The litter organic carbon content was measured via the oil-bathK2Cr2O7 titration method [26]. Litter total nitrogen content was determined by the Kjeldahl method after heat digestion with 98% H2SO4 [27]. Litter lignin content was measured through the acid detergent digestion method and then calculated based on the weight differences [28]. The cellulose content was determined by the anthronecolorimetric method after acid detergent digestion (1:8:2 (v/v/v) of nitric acid: acetic acid: water) [29].
2.3. Soil Incubation and CO2 Analysis
The incubation experiment consisted of eight treatments with three replications (based on the above replicate forest plots): (1) topsoil with Chinese pine leaf litter (PTT); (2) topsoil with labeled locust leaf litter (RPT); (3) topsoil with litter mixtures of Chinese pine and locust (PRT); (4) deep soil with Chinese pine leaf litter (PTD); (5) deep soil with locust leaf litter (RPD); (6) deep soil with litter mixtures of Chinese pine and locust (PRD); (7) topsoil without litter; and (8) deep soil without litter. The leaf litters used in the incubation experiment were 13Clabeled via pulse labeling with 13C-enriched CO2 over six months in a growth chamber as described by Lyu et al. [30]. The physicochemical properties of the soil and litter are presented in Table 1.
Fifty grams of fresh soil (oven-dry equivalent, sieved to 2 mm) was added into a 500 mL Mason jar and preincubated for seven days at 20 °C with soil moisture at 50% of the field water-holding capacity to allow microbes to adapt to the environment. After preincubation, 1 g of 13C-labeled litter (oven dried at 60 °C for 48 h and grounded to pass a 1 mm sieve) was mixed with soil uniformly.According to the average field capacity, the soil moisture was adjusted to 60% of the maximum water-holding capacity with deionized water and remained at this level during the whole incubation period. The alkali absorption method was used to trap CO2 evolved from the soil. First, one glass vial containing 20 mL of 0.5 M NaOH solution and another vial containing 10 mL of distilled water were placed in each Mason jar. Then, the jars were sealed with airtight caps. Three Mason jars with only NaOH solution and distilled water added were used as controls to trap CO2 from the air in each jar. The CO2 trapped by NaOH solution was measured after 1, 3, 7, 14, 22, 30, 42, and 56 days by titration with 0.25 M HCl after precipitating all carbonates with 5 mL of 0.5 M BaCl2 [6]. The solution was filtered and heated at 50 °C for 24 h to obtain the BaCO3 precipitate. The δ13C abundance of the trapped CO2 was detected by isotope ratio mass spectrometry (Finnigan MAT-253, Thermo Electron Inc., Bremen, Gemany).
2.4. PLFA and 13C PLFA Analysis
The soil microbial communities were analyzed using phospholipid fatty acids (PLFAs) at the end of incubation. PLFA analysis was performed according to the methods described by White et al. [31] with modification. Eight grams of dry-weight equivalent fresh soil was extracted using a solution of chloroform, methanol, and citrate buffer in a ratio of 1.0:2.0:0.8 (v/v/v). Phospholipids were separated on a silicic acid column (Waters Inc., Milford, MA, USA) and then methylated to fatty acid methyl esters (FAMEs). The FAMEs were quantitatively analyzed by gas chromatography (Agilent 6850N, Agilent Technologies Inc., Santa Clara, CA, USA). The amount of each PLFA was calculated according to the internal standard (19:0). The 15:0i, 15:0a, 16:0i, 16:1ω7c, 16:0, 17:0i, 17:0a, 18:1ω5c, and 18:0 PLFAs were considered bacterial markers [32]. The sums of 14:0i, 15:0i, 15:0a, 16:0i, 17:0i, and 17:0a PLFAs and 14:1ω5c, 15:1ω6c, 16:1ω7c, cy17:0, cy19:0, 15:03OH, 16:12OH, 16:1ω9c, 18:1ω5c, 18:1ω7c and cy18:0 PLFAs represented gram-positive bacteria and gram-negative bacteria, respectively [33]. The 18:2ω6c, 18:3ω6c, 18:1ω9c, and 20:1ω9c PLFAs were used to represent fungi-derived fatty acids [34]. The 16:1ω5c fatty acid is used to indicate arbuscular mycorrhizal fungi (AMF) [35]. The concentrations and 13C abundances of the FAMEs were analyzed by isotope ratio mass spectrometry [36]. The proportion of C (Pi) from labeled litter in each PLFA was calculated by a mass balance approach [12]:
where δ13Ct and δ13Cc represent the δ13C enrichment of individual PLFAs in the soil samples with and without litter, respectively. δ13Cl is the δ13C enrichment of labeled CO2.
Pi = (δ13Ct − δ13Cc)/(δ13Cl − δ13Cc)
2.5. Analyses of Other Soil Properties
Soil organic carbon and total nitrogen were detected by the potassium dichromate oxidation method and Kjeldahl digestion method, respectively [37,38]. The content of soil labile organic carbon (LOC) was assayed with 333 mM potassium permanganate (KMnO4) solution followed by spectrophotometry [39]. Soil recalcitrant carbon was determined by acid hydrolysis [40]. The carbon lability was expressed as the ratio of labile carbon content to non-labile carbon content [41], which was useful for evaluating the stability of soil organic carbon. The higher the ratio, the less stable the soil carbon pool. Soil microbial biomass carbon was measured by the chloroform fumigation-extraction method following Wu et al. [42].
The activities of β-glucosidase, cellulase and alkaline phosphatase were evaluated using the methods of Guan [43]. For each sample, 5 g of soil was used to determine the alkaline phosphatase activity. One gram of soil was used to determine the β-glucosidase (GLU) and cellulase activities. The substrates were p-nitrophenyl-β-D-glucoside, disodium phenylphosphate and carboxymethyl cellulose. Controls contained distilled water instead of substrates. All three enzyme activities were detected spectrophotometrically at 400, 540, and 510 nm, respectively and expressed based on the soil dry weight. The activity of N-acetylglucosaminidase was evaluated via fluorometric microplate techniques [44]. Briefly, 1.0 g of fresh soil was added to 125 mL of 50 mM sodium acetate buffer (pH 6.0) and homogenized with a vortex mixer. The 200 μL suspension was transferred to a 96-well black plate, and then 50 μL of substrate solution (4-MUB-N-acetyl-β-D-glucosaminide) was added. The negative control included 200 μL of buffer and 50 μL of substrate solution. The sample control included 200 μL of sample suspension and 50 μL of buffer. The quench control included 200 μL of sample suspension and 50 μL of standard. The microplate was incubated for 4 h at 25 °C in darkness. The fluorescence was detected by a fluorescence microplate reader (Infinite M200 Pro, Tecan Inc., Männedorf, Switzerland) at 460 nm.
2.6. PE Calculation
According to the method of Luo et al. [45], the PE (mg C kg−1 soil) induced by litter addition was calculated as follows:
where α is the percentage of leaf litter-derived C in respired CO2 released from the NaOH solution. is the 13C abundance in the respired CO2 from amended soil C. is the 13C abundance in the respired CO2 from control soil C. A13Sis the 13C abundance of litter.
Rr(mg·kg−1) = αRT
Rs = RT − Rr
PE = Rs − RC
RT is the amount of accumulated CO2 emitted from soil with added leaf litter. Rr is the amount of CO2 released from leaf litter-derived C. Rs is the amount of CO2 from soil in the treatment with added leaf litter. Rc is the amount of CO2 derived from the control soil.
2.7. Statistical Analyses
Repeated measures analysis of variance (ANOVA) was used to analyze the temporal impacts of litter addition on SOM mineralization (CO2 emissions and partitioning) and PE throughout theincubation. One-way ANOVA was conducted to analyze the impacts of litter addition on the soil microbial communities and microbial biomass carbon in each detected soil layer at the end of incubation. For all ANOVAs, the least significant difference (LSD) test was used to compare the means at a significance level of p < 0.05. The Pearson correlation analysis was used to evaluate the relationship between the 13C-labled fungal PLFA and soil PE (SPSS version 19.0, IBM, Armonk, NY, USA). The differences in soil microbial communities among treatments were evaluated by principal component analysis. The data of soil microbial communities was standardized by Hellinger. The permutational multivariate analysis of variance (PERMANOVA) analysis was used to evaluate the dissimilaritysignificances. Redundancy analysis (RDA) was carried out to assess the impacts of soil properties on the soil microbial community structure. Before RDA analysis, the data of soil properties and soil microbial community structure were standardized via scale and Hellinger, respectively. Anova with 999 permutations were used to test the significance. Principal component analysis and redundancy analysis were conducted via ‘prcomp’ and ‘rda’ functionin vegan package in R version 2.9 (R Foundation for Statistical Computing, Vienna, Austria), respectively.
3. Results
3.1. Soil Organic Carbon Pool
As shown in Table 2, leaf litter addition had no influence on the soil LOC in the topsoil but significantly increased the labile and recalcitrant carbon in the deep soil. In the topsoil, the amount of recalcitrant carbon in the locust, Chinese pine, and mixed litter treatments was 14.87%, 21.06%, and 15.22% higher, respectively, than that in the control soil. In the deep soil, the highest values of soil labile and recalcitrant carbon were observed in the Chinese pine and locust litter treatments, respectively. In addition, there were no significant differences in the carbon lability among all treatments in the topsoil. However, in the deep soil, the carbon lability after Chinese pine litter addition was significantly higher than that in the other treatments.
Table 2.
Effects of litter addition on soil organic carbon pools at the end of incubation.
3.2. Soil Organic Matter Mineralization
Litter addition significantly increased the cumulative CO2 flux in the top and deep soil, and the highest values appeared with locust litter addition (Figure 1). In the topsoil, the total cumulative CO2 flux in the locust litter addition treatment increased by 55.47%, 19.05%, and 352.04% compared with the fluxes in the Chinese pine litter addition, mixture litter addition, and no-addition treatments, respectively (Figure 1a). In deep soil, the total cumulative CO2 flux in the locust litter addition treatment increased by 132.48%, 31.64%, and 936.75% compared with the fluxes in the Chinese pine litter addition, mixture litter addition, and no-addition treatments, respectively (Figure 1b). At the end of incubation, the total cumulative CO2 flux emitted by topsoil was significantly higher than that emitted by deep soil in all treatments. However, increase in the total cumulative CO2 flux in topsoil (190.76%–352.04%) was significantly lower than that in deep soil (345.94%–936.75%), which indicated that the impact of litter addition on soil carbon mineralization was more obvious in deep soil than in topsoil.
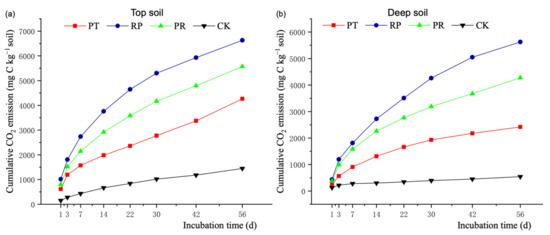
Figure 1.
Effects of litter additions on cumulative CO2 emissions during 56 days of incubation in top (a) and deep (b) soil. PT: soil with Chinese pine leaf litter; RP: soil with locust leaf litter; PR: soil with the litter mixtures of Chinese pine and locust; CK: soil without litter.Significant differences between trends were shown in Table S2a (topsoil) and Table S2b (deep soil).
3.3. Soil- and Litter-Derived CO2-C and the PE
During 56 days of incubation, the released CO2-C was mainly derived from litter (Figure 2). Leaf litter type was an important factor affecting the CO2-C distribution. In the topsoil, during 14 days of incubation, locust and mixed leaf litter addition significantly increased the release of litter-derived CO2-C (Figure 2b,c). Then, the amount of mineralized C mainly derived from locust and mixed leaf litter decomposition decreased. These results indicated that the two kinds of leaf litter gradually induced native SOC-derived CO2 production during days 15–56. However, the amount of CO2 derived from the addition of Chinese pine leaf litter peaked on days 42–56 (Figure 2a). In the deep soil, the release of litter-derived CO2-C after locust and mixed litter addition showed a similar trend with those in the topsoil (Figure 2e,f). In the Chinese pine and mixed litter treatment, the total amount of CO2 was mainly derived from litter during days 1–7 of incubation. Subsequently, CO2 emissions derived from native SOC gradually increased (Figure 2d).
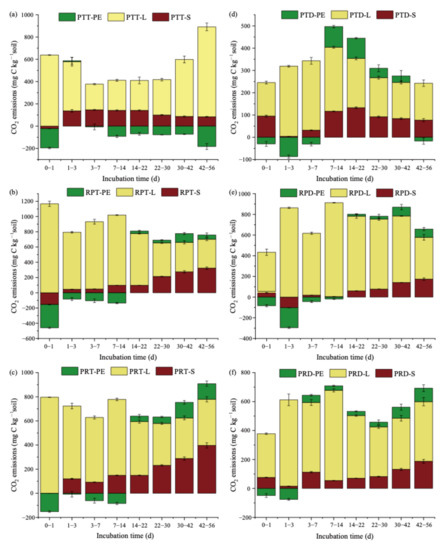
Figure 2.
Partitioning of C sources in CO2 under litter additions during 56 days of incubation. PTT: topsoil with Chinese pine leaf litter (a); RPT: topsoil with locust leaf litter (b); PRT: topsoil with the litter mixtures of Chinese pine and locust (c); PTD: deep soil with Chinese pine leaf litter (d); RPD: deep soil with locust leaf litter (e); PRD: deep soil with the litter mixtures of Chinese pine and locust (f); S: the amount of CO2 from the soils; L: the amount of CO2 from the litter; PE: priming effect.Significant differences about partitioning of C sources in CO2 under litter additions were shown in Table S3a (topsoil) and Table S3b (deep soil).
All litter addition treatments induced a PE in the measured soil layers. However, the direction and extent of the PE depended on the litter characteristics (as shown in Table 1) and soil layer. In the topsoil layer, all treatments showed negative PEs during the initial stage of 14 days. Then, positive effects were observed in the locust and mixed litter addition treatments. The extent was higher in the mixed litter addition treatment and stimulated the decomposition of native soil organic matter more intensively. However, at the end of incubation, the cumulative negative priming in the locust litter addition treatment (−389.0 mg C kg−1 soil) demonstrated that the negative PE during the initial stage of incubation balanced the positive PE at the later stage and promoted soil carbon storage (Figure 3). In the deep soil layer, all litter treatments maintained similar PEs as the top layer during incubation. However, the direction and magnitude of cumulative priming at the end of the incubation were different from those in the top layer. The cumulative PE induced by locust, Chinese pine, and mixed litter addition was −191.7 mg C kg−1 soil, 89.9 mg C kg−1 soil, and 187.8 mg C kg−1 soil, respectively (Figure 3). Compared to that in the top layer, the response of soil organic matter mineralization in the deep layer was more sensitive to litter addition.
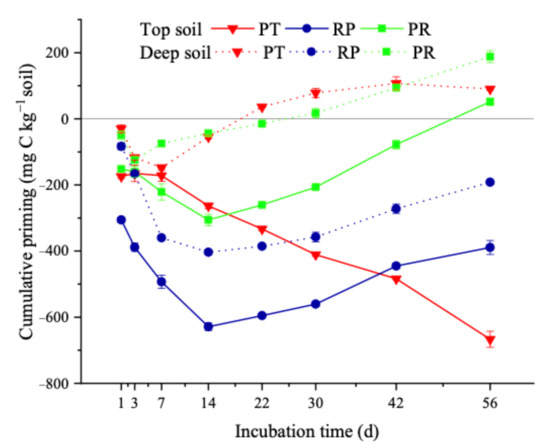
Figure 3.
Effects of litter additions on the cumulative PEs during 56 days of incubation. PT: soil with Chinese pine leaf litter; RP: soil with locust leaf litter; PR: soil with the litter mixtures of Chinese pine and locust.Significant differences between trends were shown in Table S4a (top soil) and Table S4b (deep soil).
3.4. Soil Microbial Communities
In the topsoil layer, leaf litter addition significantly affected the soil microbial communities, especially causing an increase in the total PLFAs, bacterial PLFAs, fungal PLFAs, and G−PLFAs compared to those in the control soils (Table 3). The highest total PLFAs, bacterial PLFAs, and G−PLFAs were observed after mixed leaf litter addition. Fungal PLFAs in the locust litter treatment were highest. However, there was no significance in G+PLFAs or AMF among treatments. In addition, the ratio of fungi to bacteria (F:B) increased markedly in all leaf litter treatments. However, the ratio of G+ to G− (G+:G−) showed a reverse trend (Figure 4). In the deep soil, the total concentration about PLFAs was very low and only four PLFAs might meet the demand of isotope analysis. Therefore, we did not analyze the data about PLFAs in the deep soil.
Table 3.
Effects of litter addition on the soil microbial community composition at the end of incubation.
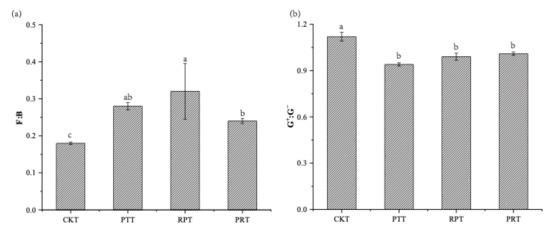
Figure 4.
The ratios of fungi to bacteria (F:B) (a) and gram-positive bacteria to gram-negative bacteria (G+:G−) (b) over 56 days of incubation under Chinese pine, locust and mixed leaf litters addition in topsoil. CKT: topsoil without litter; PTT: topsoil with Chinese pine leaf litter; RPT: topsoil with locust leaf litter; PRT: topsoil with the litter mixtures of Chinese pine and locust. Different lowercase letters indicate significant differences among treatments at p < 0.05 level.
In topsoil layer, the 16:0, 18:1ω9c, and 18:1ω7c fatty acids assimilated a larger proportion of litter-derived carbon in all litter addition treatments. The percentages of litter-derived C within these three PLFAs were 66.70%–86.63%, 68.04%–91.38%, and 53.39%–64.67%, respectively, which illustrated that the bacterial marker 16:0, fungal marker 18:1ω9c, and G− marker 18:1ω7c involved in litter decomposition (Figure 5).
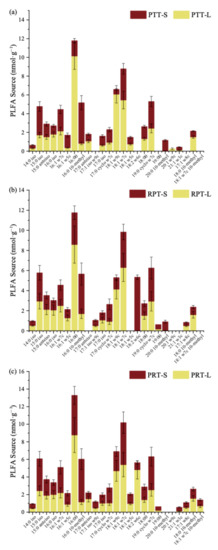
Figure 5.
Sources of soil and of litters to PLFAs content over 56 days of incubationunder Chinese pine (a), locust (b) and mixed (c) leaf litters addition in topsoil.PTT: topsoil with Chinese pine leaf litter; RPT: topsoil with locust leaf litter; PRT: topsoil with the litter mixtures of Chinese pine and locust; S: carbon derived from soil organic carbon; L: carbon derived from leaf litter addition.
The principal component analysis (PCA) results showed that the microbial community structures differed significantly among the litter addition treatments (Figure 6). Principal components 1 and 2 explained 70.27%and 17.62% of the total variance, respectively. Component 1 was mainly associated with 18:1ω7c, 17:0i, 16:1ω5c, 18:1ω9c, 16:0 and 18:0 fatty acids. In component 2, 16:1ω5c, 17:0i, 10me 18:0, 16:1ω7c, 18:1ω9c and 17:1ω8c fatty acids showed high factor weights. In addition, redundancy analysis (RDA) indicated that the first two axes explained 51.59% of the total variation in microbial communities. Soiltotal nitrogen, total organic carbon and pH were the most important factors influencing the microbial community composition (Figure 7).
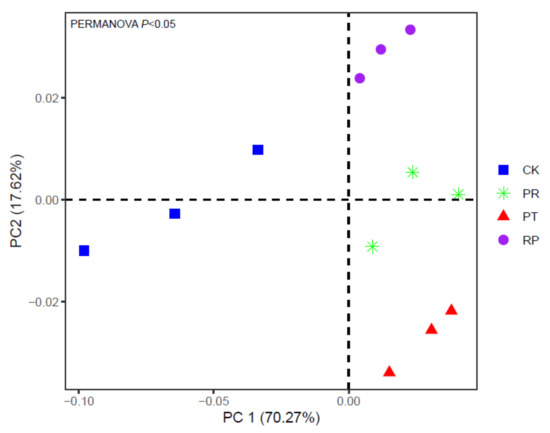
Figure 6.
PCA of the total microbial community composition and the PLFA levels over 56 days of incubation under Chinese pine, locust and mixed leaf litter addition treatments in topsoil. CK: topsoil without litter; PT: topsoil with Chinese pine leaf litter; RP: topsoil with locust leaf litter; PR: topsoil with the litter mixtures of Chinese pine and locust.
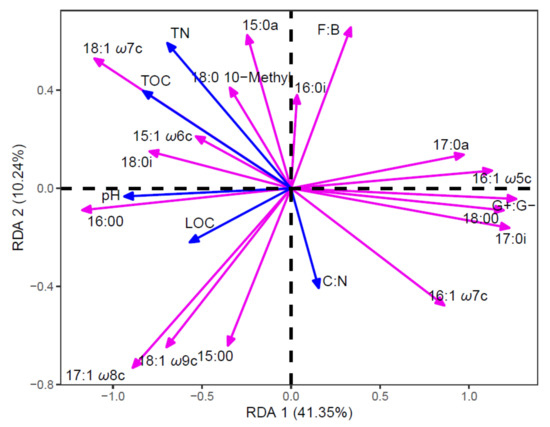
Figure 7.
Ordination plots of the redundancy analysis to identify the relationships among the soil microbial community structure of the control, Chinese pine, locust and mixed leaf litters addition (red arrows) treatments and the soil properties (blue arrows).
3.5. Microbial Biomass Carbon
Leaf litter addition significantly increased soil microbial biomass carbon and had a greater influence on microbial carbon biomass carbon in the deep soil layer than in the topsoil layer (Figure 8). In the topsoil layer, the biomass carbon was significantly higher in the leaf litter treatments than in the control. The microbial biomass carbon was 21.13%, 10.52%, and 131.15% higher in the mixed litter treatment than in the Chinese pine treatment, locust litter treatment and control soil, respectively. In the deep soil layer, leaf litter addition also led to an increase in microbial biomass carbon. However, the litter type had no significant impact on the microbial biomass carbon.
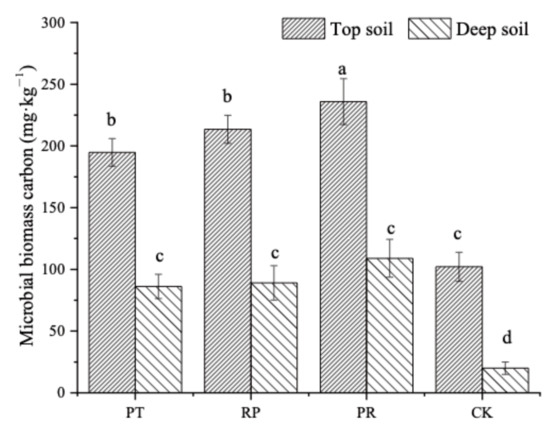
Figure 8.
Soil microbial biomass carbon content under Chinese pine, locust and mixed leaf litters addition treatments in top and deep soil. PT: Chinese pine leaf litter; RP: locust leaf litter; PR: litter mixtures of Chinese pine and locust; CK: without litter. Different lowercase letters indicate significant differences among treatments at p < 0.05 level.
3.6. Soil Enzyme Activities
Leaf litter addition significantly increased all detected enzyme activities, which had a greater influence on the deep soil layer than on the topsoil layer (Table 4). The enzyme activities after leaf litter addition mainly depended on the litter type and the soil layer. The β-glucosidase activity of the broadleaf litter (e.g., locust) treatment in the topsoil layer was significantly higher than that in the deep soil layer. However, the phosphatase and N-acetylglucosaminidase activities after the same litter addition showed reverse trends. In the coniferous leaf litter (e.g., Chinese pine) treatment, all detected enzyme activities were significantly higher in the topsoil layer than in the deep soil layer. Mixed leaf litter addition caused all detected enzyme activities, except the activities of cellulose-related enzymes, to increase greatly in the deep soil layer.
Table 4.
Effects of litter addition on soil enzyme activities at the end of incubation.
4. Discussion
Leaf litter addition significantly increased the cumulative CO2 flux in the top and deep soil, which was caused by litter decomposition and soil mineralization (Figure 1 and Figure 2). In the topsoil layer, the release of litter-derived CO2-C in the locust litter treatment increased rapidly during 14 days of incubation and then decreased gradually (Figure 2). The slow decomposition indicated that the gradual reduction of labile components and concomitant increase in recalcitrant components in the remaining residues. Our results showed that in the topsoil layer, during all incubation stages (a total of 56 days), the negative PE induced by locust litter amendment was −388.99 mg C kg−1 soil (Figure 3). In particular, during the 14 days of incubation, the amount of mineralized C mainly derived from litter decomposition was 2.38 times higher than that induced by Chinese pine litter (Figure 2). This result agreed with previous studies [5,46] and confirmed the ‘preferential substrate utilization’ hypothesis [47]. However, the Chinese pine leaf litter amendment induced a negative PE. The intensity of the PE became stronger with the extension of the incubation time. This was inconsistent with our first hypothesis. There might be a home field advantage in the process of litter decomposition. In addition, the functional breadth hypothesis assumes that decomposer communities from recalcitrant litter environments have a wider functional ability, which degrades more litter species than functional narrow communities [48]. Therefore, we speculated that the soil microbial communities distributed in Chinese pine plantations had wider functional breadth and decomposed coniferous litter more easily, resulting in a negative PE and promoting soil carbon storage. The mixed litter addition caused the native soil organic carbon mineralization more intensely compared to the single litter treatment. It was reported that the slow decomposing species might have a synergistic effect on fast decomposing ones through carbon priming [49]. We deduced that locust litter (a high quality litter, fast decomposing species) was decomposed easily in Chinese pine plantation soils, which lead to the increase of microbial community size and thus showed higher decomposing ability. When the microbial communities were limited by the carbon sources from the locust litter, the added Chinese pine litter (a low quality litter, slow decomposing species) might induce carbon priming to enhance the decomposing ability of soil microbial community. However, we did not fully confirm the mechanism of the synergistic effect of the litter mixture because the differences about species mass loss was not measured.
In our study, we found that leaf litter addition induced more obvious soil carbon mineralization and PEs in the deep soil layer than in the topsoil layer (Figure 1 and Figure 3). This result suggested that the soil organic carbon in deep soil would turnover faster than that in topsoil in response to fresh organic matter input. The main reasons for higher levels of carbon mineralization and priming in deep soil might be as follows: (1) Based on the ‘microbial nitrogen mining’hypothesis, lower nitrogen availability in soils facilitates soil microorganisms to decompose recalcitrant organic matter to obtain nitrogen [50]. The lower content of available nitrogen in deep soil (topsoil: 11.69 mg·kg−1; deep soil: 5.39 mg·kg−1) might enhance microbial nitrogen mining and induce a higher PE. (2) Topsoil is rich in easily utilized substrates (e.g., litter, rhizodeposition, and roots) [11] and is thus suitable for the growth of microorganisms. Therefore, the soil microorganisms did not mineralize more soil organic carbon in the topsoil.
The higher ratio of fungi to bacteria after litter addition suggested changes in microbial substrate utilization (Figure 4a), which might induce the succession and competition between r- (fast-growing microorganisms) and K-strategists (slow-growing microorganisms) and thus determined the magnitude and intensity of the PE [51]. Through 13C-PLFA analysis, we found that the litter-derived PLFA composition was different among the three litter treatments (based on the percentage of litter-derived 13C in the PLFAs that exceeded 50%, Table S1). This result suggested that the litter type might be the main factor that caused the transformation of the soil microbial community and substrate utilization. In the three litter treatments, the carbon derived from litter was mainly incorporated into 18:1ω9c fungal PLFAs (PTT: 91.38%, RPT: 78.11%, and PRT: 68.04%), implying that the exogenous organic carbon was mainly utilized by fungi, which was in accordance with previous studies [12,52,53]. At the same time, this result reflected the important role of fungi in litter decomposition. The Pearson correlation analysis showed a negative relation between the 13C-labeled fungal PLFAs and the soil PE (r = −0.372). It might be that fungi are better able to grow in nutrient-poor environments than bacteria [54] and thus induced less intense positive soil PEs. In this study, we detected the response of soil microbial communities only after litter addition in the topsoil layer, allowing for their activity to reach detectable levels. The magnitude and intensity of the soil PE were obviously different between the top and deep soil layers. Therefore, how soil microorganisms affect the soil PE in the deep soil layer requires further research.
Leaf litter addition promoted microbial activity and boosted enzyme production [55]. In the present study, we found that the β-glucosidase, phosphatase, and cellulase activities in the locust litter treatment were significantly higher than those in the Chinese pine litter treatment (Table 4). However, the SOM decomposition rate increased by 41.7% in the topsoil. According to previous studies, high-quality residue additions decreased specific enzyme activities because of microbial saturation with substrate [56]. If the soil microorganisms are in a state of starvation or nutrient limitation, they might secrete specific enzymes to obtain energy from native soil organic matter, resulting in higher soil enzyme activities [57]. Therefore, our results were inconsistent with these reports, potentially because of the higher contents of easily dissolved substances and lower contents of recalcitrant compounds, such as ligninsand tannins, in the leaves of broadleaf species [58]. The input of labile substrates promoted microbial growth and activity, in turn alleviating the spatial inaccessibility to SOM through their expansion to new areas [59] and thus speeding up the decomposition of SOM.
5. Conclusions
Different kinds of leaf litter addition significantly affected the magnitude and direction of priming effect in all detected soils. In top soil, Chinese pine (PT, a low quality litter with higher lignin concentration and C:N ratio) and locustlitter (RP, a high quality litter) addition induced a negative PE and reduced the losses of soil organic carbon by 46.1% and 26.9%, respectively. However, PT addition gave the primed CO2 of 89.9 mg C kg−1 soil evolved from the deep soil, indicating coniferous leaf litter input accelerated the decomposition of SOM. The input of locust (broadleaf) litter decreased the release of CO2-C derived from SOC. The magnitude of the PE in deep soil was more intensive than that in topsoil. Fungi and G- bacteria were essential for decomposing litter and incorporating new litter C in to soil microbes (average: 69.0% and 55.7%).Soil microbial communities regulated the mineralization and immobilization of SOM through litter quality control. In summary, we speculated that the introduction of broadleaf species into coniferous plantations might be beneficial for enhancing deep soil carbon stocks. However, the effects of litter transplantation among broadleaf and coniferous forests andlitter addition amount on soil PE need further research.
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4907/12/2/170/s1, Figure S1. Location of the experiment site in the Loess Plateau of China. Table S1: The percentage distribution of litter-derived 13C among PLFAs over 56 days incubation under Chinese pine, locust and mixed leaf litters addition in topsoil. Table S2a. Effects of litter additions on cumulative CO2 emissions during 56 days of incubation in topsoil. Table S2b. Effects of litter additions on cumulative CO2 emissions during 56 days of incubation in deep soil. Table S3a. Partitioning of C sources in CO2 under litter additions during 56 days of incubation in topsoil. Table S3b. Partitioning of C sources in CO2 under litter additions during 56 days of incubation in deep soil. Table S4a. Effects of litter additions on the cumulative PEs during 56 days of incubation in topsoil. Table S4b. Effects of litter additions on the cumulative PEs during 56 days of incubation in deep soil.
Author Contributions
X.Y. and M.Z. conceived and designed the experiment; S.-X.F., L.-G.D. and M.Z. completed the experiment; J.-W.Z., X.-M.J. and Y.W. analyzed the experimental data; X.Y. and M.Z. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by the National Natural Science Foundation of China (31670629), National Key Research and Development Program of China (2016YFC0501706-1), China Postdoctoral Science Foundation (2016M60082), Tang Scholar Program of Northwest A&F University, and Fundamental Research Funds for the Central Universities (2452018132).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bloom, A.A.; Exbrayat, J.F.; van Velde, I.R.; Feng, L.; Williams, M. The decadal state of the terrestrial carbon cycle: Global retrievals of terrestrial carbon allocation, pools, and residence times. Proc. Natl. Acad. Sci. USA 2016, 113, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.M.; Dijkstra, F.A.; Wang, P.; Zhu, B.; Cheng, W.X. Rhizosphere priming effects on soil carbon and nitrogen dynamics among tree species with and without intraspecific competition. New Phytol. 2018, 218, 1036–1048. [Google Scholar] [CrossRef] [PubMed]
- Kuzyakov, Y.; Friedel, J.K.; Stahr, K. Review of mechanisms and quantification of priming effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Liu, E.K.; Wang, J.B.; Zhang, Y.Q.; Angers, D.A.; Yan, C.R.; Oweis, T.; He, W.Q.; Liu, Q.; Chen, B.Q. Priming effect of 13C-labelled wheat straw in no-tillage soil under drying and wetting cycles in the Loess Plateau of China. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chao, L.; Liu, Y.Y.; Freschet, G.T.; Zhang, W.D.; Yu, X.; Zheng, W.H.; Guan, X.; Yang, Q.P.; Chen, L.C.; Dijkstra, F.A.; et al. Litter carbon and nutrient chemistry control the magnitude of soil priming effect. Funct. Ecol. 2019, 33, 876–888. [Google Scholar] [CrossRef]
- Shahzad, T.; Anwar, F.; Hussain, S.; Mahmood, F.; Arif, M.S.; Sahar, A.; Nawaz, M.F.; Perveen, N.; Sanaullah, M.; Rehman, K.; et al. Carbon dynamics in surface and deep soil in response to increasing litter addition rates in an agro-ecosystem. Geoderma 2019, 333, 1–9. [Google Scholar] [CrossRef]
- Hick, L.C.; Meri, P.; Nottingham, A.T.; Reay, D.S.; Stott, A.W.; Salinas, N.; Whitaker, J. Carbon and nitrogen inputs differentially affect priming of soil organic matter in tropical lowland and montane soils. Soil Biol. Biochem. 2019, 129, 212–222. [Google Scholar] [CrossRef]
- Liu, J.; Liu, X.Y.; Song, Q.N.; Compson, Z.G.; LeRoy, C.J.; Luan, F.G.; Wang, H.; Hu, Y.L.; Yang, Q.P. Synergistic effects: A common theme in mixed-species litter decomposition. New Phytol. 2020, 227, 757–765. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Y.J.; Zou, J.W.; Siemann, E. Decomposition of Phragmitesaustralis litter retarded by invasive Solidagocanadensis in mixtures: An antagonistic non-additive effect. Sci. Rep. 2014, 4, 1–8. [Google Scholar]
- Scheibe, A.; Gleixner, G. Influence of litter diversity on dissolved organic matter release and soil carbon formation in a mixed beech forest. PLoS ONE 2014, 9, 1–21. [Google Scholar] [CrossRef][Green Version]
- Fontaine, S.; Barot, S.; Barré, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.K.; Wang, Y.P.; Wang, S.L.; He, T.X.; Liu, L. Fresh carbon and nitrogen inputs alter organic carbon mineralization and microbial community in forest deep soil layers. Soil Biol. Biochem. 2014, 72, 145–151. [Google Scholar] [CrossRef]
- Fontaine, S.; Barot, S. Size and functional diversity of microbe populations control plant persistence and long-term soil carbon accumulation. Ecol. Lett. 2005, 8, 1075–1087. [Google Scholar] [CrossRef]
- Wang, H.; Xu, W.; Hu, G.; Dai, W.; Jiang, P.; Bai, E. The priming effect of soluble carbon inputs in organic and mineral soils from a temperate forest. Oecologia 2015, 178, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Hartley, I.P.; Hopkins, D.W.; Sommerkorn, M.; Wookey, P.A. The response of organic matter mineralisation to nutrient and substrate additions in sub-arctic soils. Soil Biol. Biochem. 2010, 42, 92–100. [Google Scholar] [CrossRef]
- Norris, C.E.; Quideau, S.A.; Oh, S.W. Microbial utilization of double-labeled aspen litter in boreal aspen and spruce soils. Soil Biol. Biochem. 2016, 100, 9–20. [Google Scholar] [CrossRef]
- Jílková, V.; Jandová, K.; Cajthaml, T.; Devetter, M.; Kukla, J.; Starý, J.; Vacířová, A. Organic matter decomposition and carbon content in soil fractions as affected by a gradient of labile carbon input to a temperate forest soil. Biol. Fertil. Soils 2020, 56, 411–421. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Turner, B.L.; Stott, A.W.; Tanner, E.V.J. Nitrogen and phosphorus constrain stable and labile carbon turnover in lowland tropical forest soils. Soil Biol. Biochem. 2015, 80, 26–33. [Google Scholar] [CrossRef]
- Yu, G.C.; Zhao, H.B.; Chen, J.; Zhang, T.L.; Cai, Z.L.; Zhou, G.Y.; Li, Z.J.; Qiu, Z.J.; Wu, Z.M. Soil microbial community dynamics mediate the priming effects caused by in situ decomposition of fresh plant residues. Sci. Total Environ. 2020, 737, 1–13. [Google Scholar] [CrossRef]
- Whitaker, J.; Ostle, N.; McNamara, N.P.; Nottingham, A.T.; Scott, A.W.; Bardgett, R.D.; Salinas, N.; Ccahuana, A.J.Q.; Meri, P. Microbial carbon mineralization in tropical lowland and montane forest soils of Peru. Front. Microbiol. 2014, 5, 1–14. [Google Scholar] [CrossRef]
- Herman, D.J.; Firestone, M.K.; Nuccio, E.; Hodge, A. Interactions between an arbuscularmycorrhizal fungus and a soil microbial community mediating litter decomposition. FEMS Microbiol. Ecol. 2012, 80, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Liang, C.; Bodè, S.; Huygens, D.; Boeckx, P. Phospholipid 13C stable isotopic probing during decomposition of wheat residues. Appl. Soil Ecol. 2016, 98, 65–74. [Google Scholar] [CrossRef]
- Fontaine, S.; Mariotti, A.; Abbadie, L. The priming effect of organic matter: A question of microbial competition? Soil Biol. Biochem. 2003, 35, 837–843. [Google Scholar] [CrossRef]
- DeVries, D.A.; Peter, J. Women on display: The effect of portraying the self online on women’s self-objectification. Comput. Hum. Behav. 2013, 29, 1483–1489. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Turner, B.L.; Chamberlain, P.M.; Stott, A.W.; Tanner, E.V.J. Priming and microbial nutrient limitation in lowland tropical forest soils of contrasting fertility. Biogeochemistry 2012, 111, 219–237. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis; Page, A.L., Miller, R.H., Keeney, R.D., Eds.; American Society of Agronomy and Soil Science Society of American: Madison, WI, USA, 1982; pp. 101–129. [Google Scholar]
- Gallaher, R.N.; Weldon, C.O.; Boswell, F.C. A semiautomated procedure for total nitrogen in plant and soil samples. Soil Sci. Soc. Am. J. 1976, 40, 887–889. [Google Scholar] [CrossRef]
- Dranski, J.A.L.; Malavasi, U.C.; Malavasi, M.D. Relationship between lignin content and quality of Pinustaeda seedlings. Rev. Arvore 2015, 39, 905–913. [Google Scholar] [CrossRef]
- Updegraff, D.M. Semimicro determination of cellulose in biological materials. Anal. Biochem. 1969, 32, 420–424. [Google Scholar] [CrossRef]
- Lyu, M.K.; Xie, J.S.; Vadeboncoeur, M.A.; Wang, M.H.; Qiu, X.; Ren, Y.B.; Jiang, M.H.; Yang, Y.S.; Kuzyakov, Y. Simulated leaf litter addition causes opposite priming effects on natural forest and plantation soils. Biol. Fertil. Soils 2018, 54, 925–934. [Google Scholar] [CrossRef]
- White, D.; Stair, J.; Ringelberg, D. Quantitative comparisons of in situ microbial biodiversity by signature biomarker analysis. J. Ind. Microbiol. Biotechnol. 1996, 17, 185–196. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, W.J.; Wei, W.J.; He, Z.L.; Kuzyakov, Y.; Bol, R.; Hu, R.G. Soil organic matter priming and carbon balance after straw addition is regulated by long-term fertilization. Soil Biol. Biochem. 2019, 135, 383–391. [Google Scholar] [CrossRef]
- Qiao, N.; Wang, J.; Xu, X.L.; Shen, Y.X.; Long, X.E.; Hu, Y.H.; Schaefer, D.; Li, S.G.; Wang, H.M.; Kuzyakov, Y. Priming alters soil carbon dynamics during forest succession. Biol. Fertil. Soils 2019, 55, 339–350. [Google Scholar] [CrossRef]
- Frostegård, Å.; Bååth, E. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol. Fertil. Soils 1996, 22, 59–65. [Google Scholar] [CrossRef]
- Olsson, P.A.; Thingstrup, I.; Jakobsen, I.; Bååtha, E. Estimation of the biomass of arbuscularmycorrhizal fungi in a linseed field. Soil Biol. Biochem. 1999, 31, 1879–1887. [Google Scholar] [CrossRef]
- Williams, M.A.; Myrold, D.D.; Bottomley, P.J. Carbon flow from 13C-labeled straw and root residues into the phospholipid fatty acids of a soil microbial community under field conditions. Soil Biol. Biochem. 2006, 38, 759–768. [Google Scholar] [CrossRef]
- Walkley, A.J.; Black, I.A. Estimation of soil organic carbon by the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Bremner, J.M. Nitrogen-Total. In Methods of Soil Analysis: Part III, Chemical Methods, 2nd ed.; Sparks, D.L., Ed.; Soil Science Society America: Madison, WI, USA, 1996; pp. 1085–1122. [Google Scholar]
- DuPont, S.T.; Culman, S.W.; Ferris, H.; Buckley, D.H.; Glover, J.D. No-tillage conversion of harvested perennial grassland to annual cropland reduces root biomass, decreases active carbon stocks, and impacts soil biota. Agric. Ecosyst. Environ. 2010, 137, 25–32. [Google Scholar] [CrossRef]
- Leavitt, S.W.; Follett, R.F.; Paul, E.A. Estimation of slow-and fast-cycling soil organic carbon pools from 6 N HCl hydrolysis. Radiocarbon 1996, 38, 231–239. [Google Scholar] [CrossRef]
- Sainepo, B.M.; Gachene, C.; Karuma, A. Assessment of soil organic fractions and carbon management index under different land use types in Olesharo Catchment, Narok County, Kenya. Carbon Balance Manag. 2018, 13, 1–9. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation extraction-an autoclaved procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Guan, S.Y. Soil Enzyme and Research Methods; Agricultural Press: Beijing, China, 1986. (In Chinese) [Google Scholar]
- Qi, R.M.; Li, J.; Lin, Z.A.; Li, Z.J.; Li, Y.T.; Yang, X.D.; Zhang, J.J.; Zhao, B.Q. Temperature effects on soil organic carbon, soil labile organic carbon fractions, and soil enzyme activities under long-term fertilization regimes. Appl. Soil Ecol. 2016, 102, 36–45. [Google Scholar] [CrossRef]
- Luo, Y.; Durenkamp, M.; De Nobili, M.; Lin, Q.; Brookes, P.C. Short term soil priming effects and the mineralisation of biochar following its incorporation to soils of different pH. Soil Biol. Biochem. 2011, 43, 2304–2314. [Google Scholar] [CrossRef]
- Wang, H.; Boutton, T.W.; Xu, W.H.; Hu, G.Q.; Jiang, P.; Bai, E. Quality of fresh organic matter affects priming of soil organic matter and substrate utilization patterns of microbes. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.R.; Senbayram, M.; Blagodatsky, S.; Myachina, O.; Dittert, K.; Lin, X.; Blagodatskaya, E.; Kuzyakov, Y. Soil C and N availability determine the priming effect: Microbial N mining and stoichiometric decomposition theories. Glob. Chang. Biol. 2014, 20, 2356–2367. [Google Scholar] [CrossRef] [PubMed]
- Keiser, A.D.; Keiser, D.A.; Strickland, M.S.; Bradford, M.A. Disentangling the mechanisms underlying functional differences among decomposer communities. J. Ecol. 2014, 102, 603–609. [Google Scholar] [CrossRef]
- Cuchietti, A.; Marcotti, E.; Gurvich, D.E.; Cingolani, A.M.; Harguindeguy, N.P. Leaf litter mixtures and neighbor effects: Low-nitrogen and high-lignin species increase decomposition rate of high-nitrogen and low-lignin neighbours. Appl. Soil Ecol. 2014, 82, 44–51. [Google Scholar] [CrossRef]
- Mason-Jones, K.; Schmucker, N.; Kuzyakov, Y. Contrasting effects of organic and mineral nitrogen challenge the N-mining hypothesis for soil organic matter priming. Soil Biol. Biochem. 2018, 124, 38–46. [Google Scholar] [CrossRef]
- Wang, Y.M.; Li, M.; Jiang, C.Y.; Liu, M.; Wu, M.; Liu, P.; Li, Z.P.; Uchimiya, M.; Yuan, X.Y. Soil microbiome-induced changes in the priming effects of 13C-labelled substrates from rice residues. Sci. Total Environ. 2020, 726, 1–11. [Google Scholar] [CrossRef]
- Zhang, P.; Tian, X.J.; He, X.B.; Song, F.Q.; Ren, L.L.; Jiang, P. Effect of litter quality on its decomposition in broad leaf and coniferous forest. Eur. J. Soil Biol. 2008, 44, 392–399. [Google Scholar] [CrossRef]
- Li, Z.Q.; Song, M.; Li, D.D.; Ma, L.; Zhao, B.Z.; Zhang, J.B. Effect of long-term fertilization on decomposition of crop residues and their incorporation into microbial communities of 6-year stored soils. Biol. Fertil. Soils 2020, 56, 25–37. [Google Scholar] [CrossRef]
- Magan, N.; Fragoeiro, S.; Bastos, C. Environmental factors and bioremediation of xenobiotics using white rot fungi. Mycobiology 2010, 38, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Blagodatskaya, E.; Kuzyakov, Y. Active microorganisms in soil: Critical review of estimation criteria and approaches. Soil Biol. Biochem. 2013, 67, 192–211. [Google Scholar] [CrossRef]
- Xiao, C.; Guenet, B.; Zhou, Y.; Su, J.; Janssens, I.A. Priming of soil organic matter decomposition scales linearly with microbial biomass response to litter input in steppe vegetation. Oikos 2015, 124, 649–657. [Google Scholar] [CrossRef]
- Blagodatskaya, E.; Khomyakov, N.; Myachina, O.; Bogomolova, I.; Blagodatsky, S.; Kuzyako, Y. Microbial interactions affect sources of priming induced by cellulose. Soil Biol. Biochem. 2014, 74, 39–49. [Google Scholar] [CrossRef]
- Ferreira, V.; Faustino, H.; Raposeiro, P.M.; Gonçalves, V. Replacement of native forests by conifer plantations affects fungal decomposer community structure but not litter decomposition in Atlantic island streams. For. Ecol. Manag. 2017, 389, 323–330. [Google Scholar] [CrossRef]
- Finley, B.K.; Dijkstra, P.; Rasmussen, C.; Schwartz, E.; Mau, R.L.; Liu, X.J.A.; Getel, N.; Hungate, B.A. Soil mineral assemblage and substrate quality effects on microbial priming. Geoderma 2018, 322, 38–47. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).