The Populations and Habitat Preferences of Three Elm Species in Conditions Prevailing on Plains of Poland


Abstract
1. Introduction
- The habitat preferences of elms growing in a given area.
- Whether the occurrence of DED depends on the species of elm and the habitat in which it occurs.
2. Materials and Methods
2.1. Study Area
2.2. Methodology
- Distributions of separate data for all site types identified in a given classification.
- Distributions of data from alluvial (AF) site types only (these habitats are particularly related to elms) in the first classification (based on the topography).
- Distributions of summary data from alluvial (AF) and from oak-hornbeam (OH-F/fresh deciduous forest and fresh mixed deciduous forest) site types (the oak-hornbeam forest potentially has habitats with the largest area in Poland).
- Distributions of summary data from ‘moist’ and from ‘fresh’ site types.
- Distributions of data for slope forest (SF) site and for sum data from other site types.
3. Results
3.1. Results Are Based on Actual Data and Were Prepared on the Basis of the Forest Inventory (without Division into Species)
3.1.1. Number and Area of Forest Plots with Elms
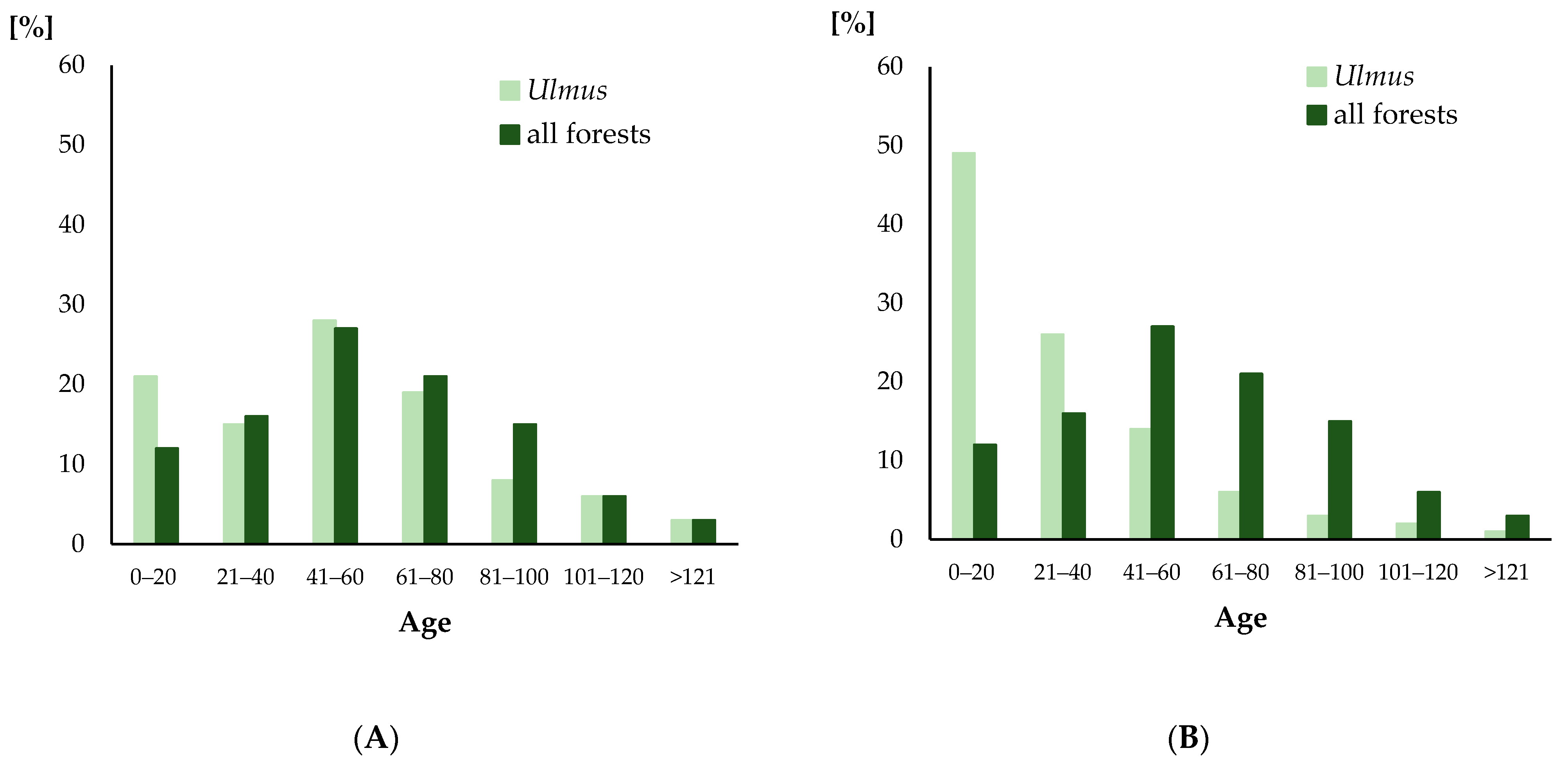
3.1.2. Age Distribution
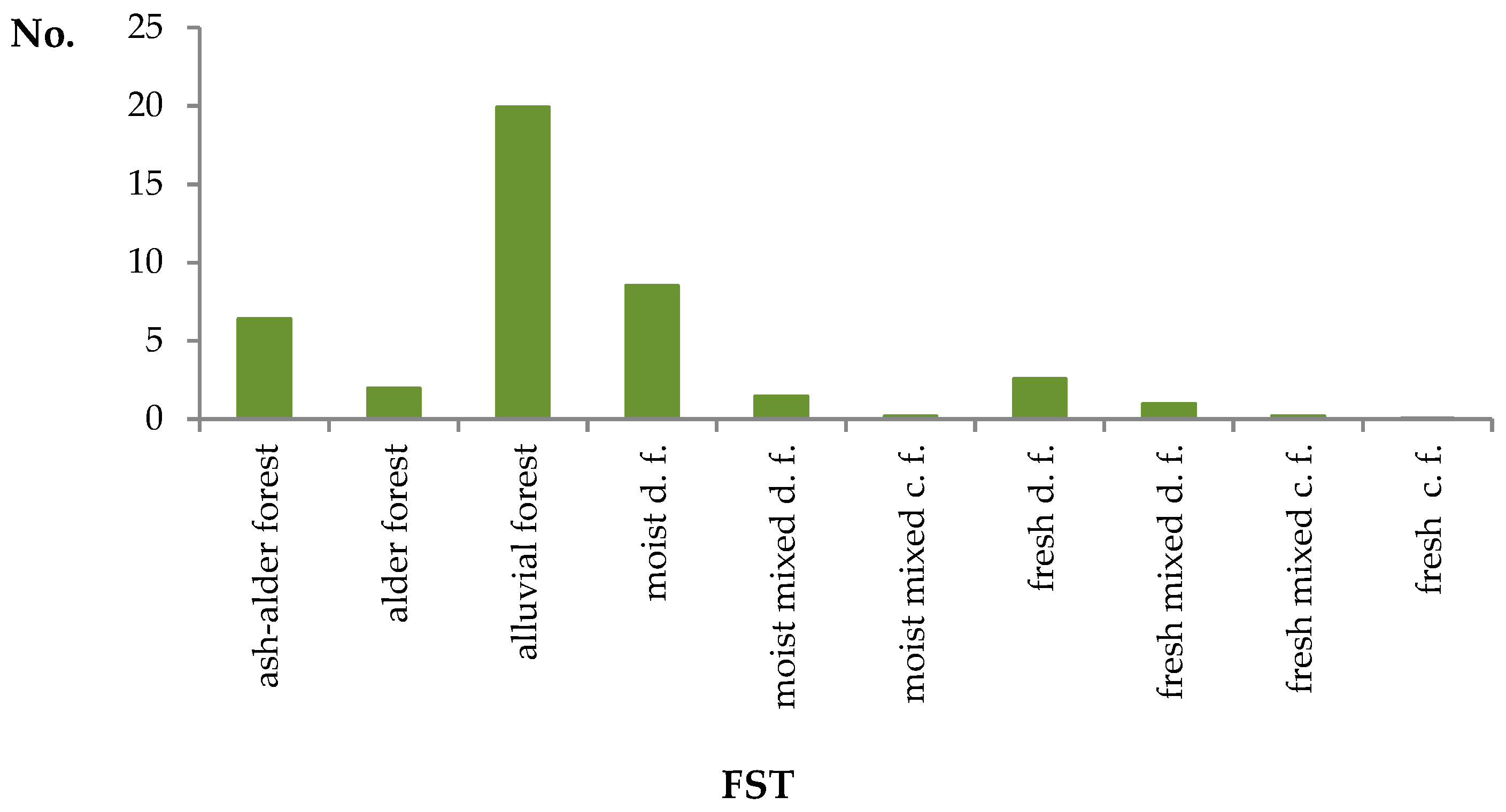
3.1.3. Site Preferences
3.2. Results Based on Field Research on (Randomly) Selected Elm Localities (with Division into Species)
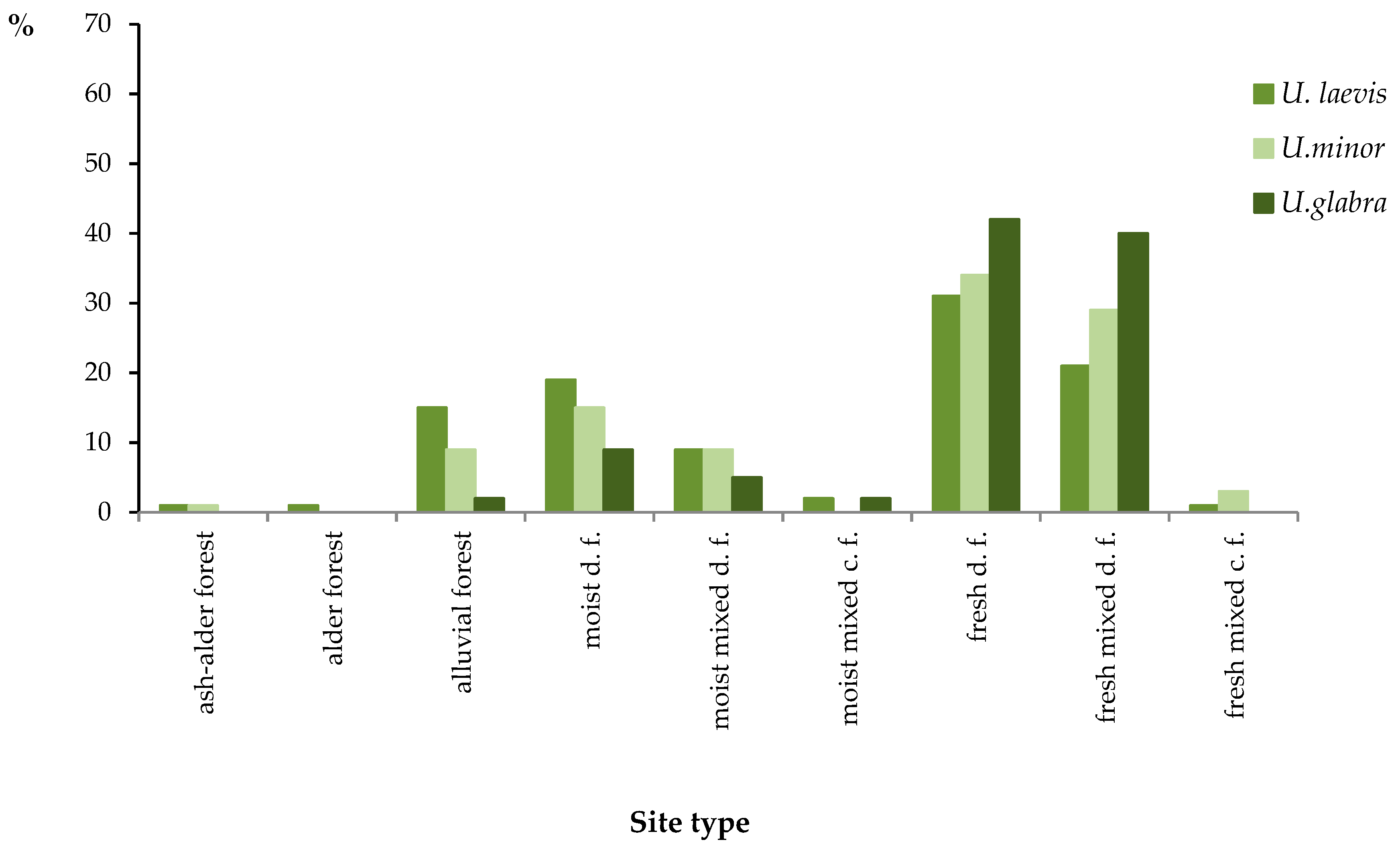
3.2.1. Site Preferences of Individual Elm Species
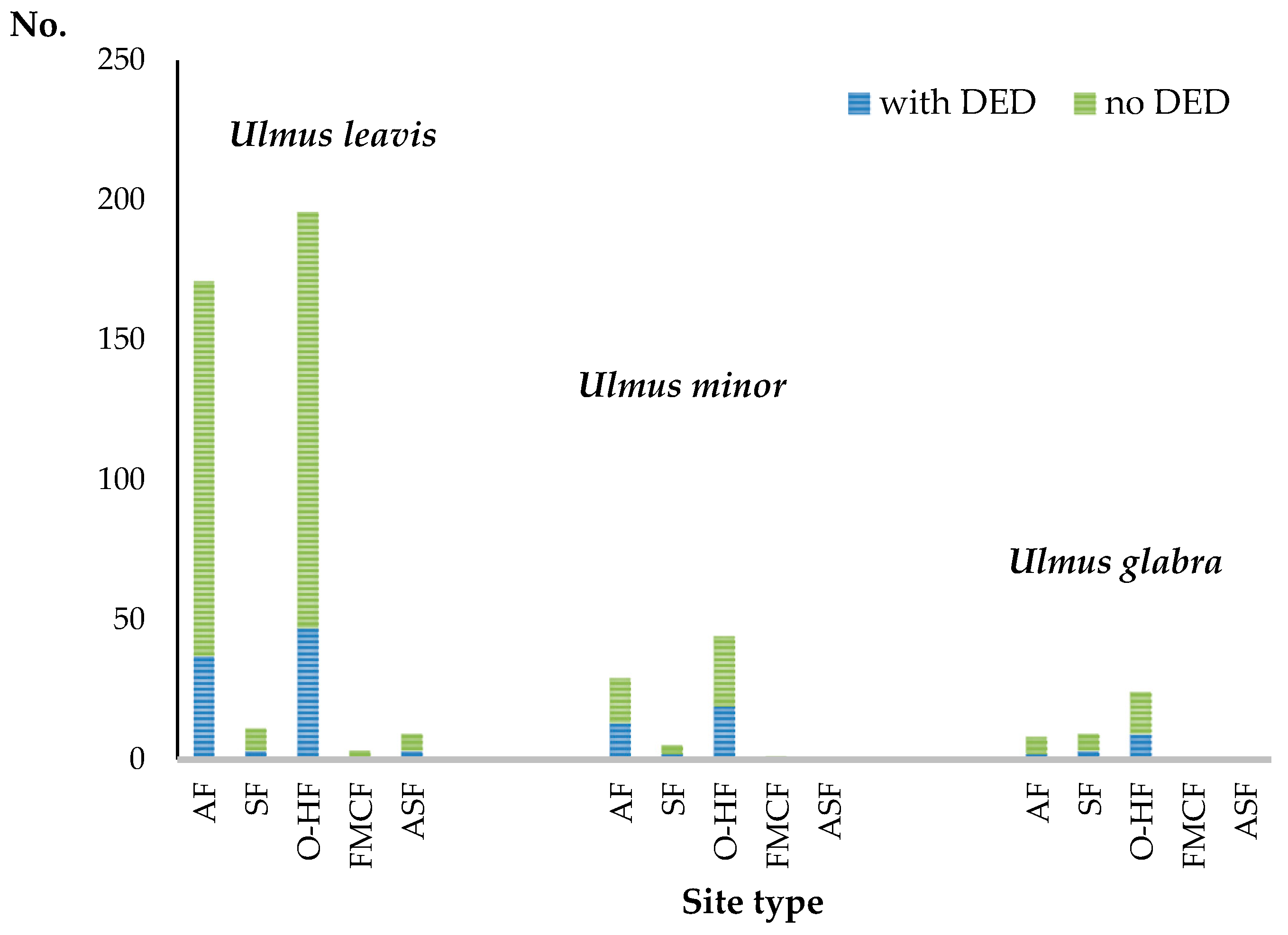
3.2.2. The Occurrence of DED Depends on the Species of Elm and the Habitat
4. Discussion
4.1. Age Distribution
4.2. Number and Area of Forest Plots with Elms
4.3. The Elms Habitat Preferences
4.4. Individual Elm Species Importance in Today’s Forest Communities
4.5. Factors, That May Be Responsible for the Current State of Elms Populations
4.6. Ability to Overestimate the Number of U. minor in Floristic Researches
4.7. Relationship between the Type of Habitat and DED Symptoms
4.8. Suggestions for Silviculture
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boratyńska, K.; Sękiewicz, M.; Boratyński, A. Morfologia, systematyka, zmienność i rozmieszczenie geograficzne. [Morphology, systematics, variability and geographical distribution]. In Wiązy [Elms]; Bugała, W., Boratyński, A., Iszkuło, G., Eds.; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2015; pp. 24–52. [Google Scholar]
- Filipiak, M.; Napierała-Filipiak, A. Zarys ekologii [Outline of ecology]. In Wiązy [Elms]; Bugała, W., Boratyński, A., Iszkuło, G., Eds.; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2015; pp. 133–181. [Google Scholar]
- Brasier, C.M. Ophiostoma novo-ulmi sp. nov., causative agent of the current Dutch elm disease pandemics. Mycopathologia 1991, 115, 151–161. [Google Scholar] [CrossRef]
- Collin, E.; Rondouin, M.; Joyeau, C.; Matz, S.; Raimbault, P.; Harvengt, L.; Bilger, I.; Guibert, M. Conservation and use of elm genetic resources in France: Results and perspectives. iForest—Biogeosci. For. 2020, 13, 41–47. [Google Scholar] [CrossRef]
- Łakomy, P.; Kwaśna, H.; Kuźmiński, R.; Napierała-Filipiak, A.; Filipiak, M.; Behnke, K.; Behnke-Borowczyk, J. Investigation of Ophiostoma population infected elms in Poland. Dendrobiology 2016, 76, 137–144. [Google Scholar] [CrossRef]
- Martín, J.A.; Sobrino-Plata, J.; Rodríguez-Calcerrada, J.; Collada, C.; Gil, L. Breeding and scientific advances in the fight against Dutch elm disease: Will they allow the use of elms in forest restoration? New For. 2019, 50, 183–215. [Google Scholar] [CrossRef]
- Heybroek, H.M. The elm, tree of milk and wine. iForest—Biogeosci. For. 2015, 8, 181–186. [Google Scholar] [CrossRef]
- Jaworski, A. Hodowla Lasu. Charakterystyka Hodowlana Drzew i Krzewów Leśnych [ Silviculture, Characteristics of Forest Trees and Shrubs; PWRiL: Warszawa, Poland, 2011. [Google Scholar]
- Mańka, K.; Sobiczewski, P.; Mańka, M.; Fiedorow, Z. Fitopatologia Leśna [Forest Pathology]; PWRiL: Warszawa, Poland, 2005. [Google Scholar]
- Santini, A.; Faccoli, M. Dutch elm disease and elm bark beetles: A century of association. iForest—Biogeosci. For. 2015, 8, 126–134. [Google Scholar] [CrossRef]
- Mackenthun, G.L. Elm Losses and their Causes over a 20 Year Period—A long-term Study of Ulmus in Saxony, Germany. In Proceedings of the Third International Elm Conference, Florence, Italy, 9–11 October 2013; pp. 9–11. [Google Scholar]
- Napierała-Filipiak, A.; Filipiak, M.; Jaworek, J. Rozmieszczenie zasobów drzew z rodzaju wiąz (Ulmus spp.) w lasach Polski w świetle dokumentacji leśnej [Distribution of elms (Ulmus spp.) in Polish forests according to the forestry inventory]. Sylwan 2014, 158, 811–820. [Google Scholar]
- Napierała-Filipiak, A.; Filipiak, M.; Łakomy, P.; Kuźmiński, R.; Gubański, J. Changes in elm (Ulmus) populations of mid-western Poland during the past 35 years. Dendrobiology 2016, 76, 145–156. [Google Scholar] [CrossRef]
- Napierała-Filipiak, A.; Filipiak, M.; Łakomy, P. Changes in the Species Composition of Elms (Ulmus spp.) in Poland. Forests 2019, 10, 1008. [Google Scholar] [CrossRef]
- Chudzińska, M.; Pałucka, M.; Pasławska, A.; Litkowiec, M.; Lewandowski, A.; Kozioł, C. Wyniki wstępnych badań nad zmiennością genetyczną oraz zróżnicowaniem genetycznym między populacjami wiązu górskiego (Ulmus glabra Huds.) w Polsce [Results of preliminary research on genetic variation and genetic differentiation between Wych elm populations]. (Ulmus glabra Huds.) in Poland]. Sylwan 2018, 162, 727–736. [Google Scholar] [CrossRef]
- Nielsen, L.R.; Kjær, E.D. Fine-scale gene flow and genetic structure in a relic Ulmus laevis population at its northern range. Tree Genet Genomes 2010, 6, 643–649. [Google Scholar] [CrossRef]
- Puerto, M.M.; García, F.M.; Mohanty, A.; Martín, J.P. Genetic diversity in relict and fragmented populations of Ulmus glabra Hudson in the Central System of the Iberian Peninsula. Forests 2017, 8, 143. [Google Scholar] [CrossRef]
- Venturas, M.; Fuentes-Utrilla, P.; López, R.; Perea, R.; Fernández, V.; Gascó, A.; Guzmán, P.; Li, M.; Rodríguez-Calcerrada, J.; Miranda, E.; et al. Ulmus laevis in the Iberian Peninsula: A review of its ecology and conservation. iForest 2015, 8, 135–142. [Google Scholar] [CrossRef]
- Fuentes-Utrilla, P.; Venturas, M.; Hollingsworth, P.M.; Squirrell, J.; Collada, C.; Stone, G.N.; Gil, L. Extending glacial refugia for a European tree: Genetic markers show that Iberian populations of white elm are native relicts and not introductions. Heredity 2014, 112, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Mackenthun, G.L. The role of Ulmus laevis in German floodplain landscapes. For. Syst. 2004, 13, 55–63. [Google Scholar]
- Richens, R.H. Elm; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Heybroek, H.M. The Dutch elm breeding program. In Dutch Elm Disease Research: Cellular and MolecularApproaches; Sticklen, M.B., Sherald, J.L., Eds.; Springer: New York, NY, USA, 1993; pp. 16–25. [Google Scholar]
- Kieliszewska-Rokicka, B. Wymiana gazowa i gospodarka wodna [Gas exchange and water relations]. In Wiązy [Elms]; Bugała, W., Boratyński, A., Iszkuło, G., Eds.; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2015; pp. 82–95. [Google Scholar]
- Babushkina, E.A.; Zhirnova, D.F.; Belokopytova, L.V.; Tychkov, I.I.; Vaganov, E.A.; Krutovsky, K.V. Response of Four Tree Species to Changing Climate in a Moisture-Limited Area of South Siberia. Forests 2019, 10, 999. [Google Scholar] [CrossRef]
- Yuan, Y.; Ren, Y.; Gao, G.; Zhao, Z.; Niu, S. Intra-and interspecific interactions among pioneer trees affect forest-biomass carbon accumulation in a nutrient-deficient reclaimed coal mine spoil. Forests 2020, 11, 819. [Google Scholar] [CrossRef]
- Kassahun, Z.; Yow, J.N.; Renninger, H.J. Diversity or Redundancy in Leaf Physiological and Anatomical Parameters in a Species Diverse, Bottomland Hardwood Forest? Forests 2020, 11, 519. [Google Scholar] [CrossRef]
- Falkengren-Grerup, U.; Brunet, J.; Diekmann, M. Nitrogen mineralisation in deciduous forest soils in south Sweden in gradients of soil acidity and deposition. Environ. Pollut. 1998, 102, 415–420. [Google Scholar] [CrossRef]
- Polyakova, O.; Billor, N. Impact of deciduous tree species on litterfall quality, decomposition rates and nutrient circulation in pine stands. For. Ecol. Manag. 2007, 253, 11–18. [Google Scholar] [CrossRef]
- Li, X.; Dong, W.; Song, Y.; Wang, W.; Zhan, W. Effect of Soil Fauna on Home-Field Advantages of Litter Mass Loss and Nutrient Release in Different Temperate Broad-Leaved Forests. Forests 2019, 10, 1033. [Google Scholar] [CrossRef]
- Pogrebniak, P. Podstawy Typologii Leśnej [Basics of Forest Typology]; PWRiL: Warszawa, Poland, 1961. [Google Scholar]
- Mittempergher, L.; Santini, A. The history of elm breeding. For. Syst. 2004, 13, 161–177. [Google Scholar]
- Mioduszewski, S.; Korczyk, A.F. Growth of Ulmus glabra Huds. grafts in the clone archive in Bielsk Forest District. Leśne Prace Badawcze [For. Res. Pap.] 2013, 74, 149–159. [Google Scholar] [CrossRef][Green Version]
- Piou, D.; Benest, F.; Collin, E. Est-il possible de tirer des enseignements des introductions anciennes? L’exemple de la graphiose de l’orme [Are there any lessons to be learnt from long-standing introductions of pathogens? The case of Dutch Elm Disease]. Revue Forestière Française 2018, 70, 621–637. [Google Scholar] [CrossRef]
- Martín, J.A.; Solla, A.; Esteban, L.G.; De Palacios, P.; Gil, L. Bordered pit and ray morphology involvement in elm resistance to Ophiostoma novo-ulmi. Can. J. Bot. 2009, 39, 420–429. [Google Scholar] [CrossRef]
- Solheim, H.; Eriksen, R.; Hietala, A.M. Dutch elm disease has currently a low incidence on wych elm in Norway. For. Path. 2011, 41, 182–188. [Google Scholar] [CrossRef]
- Urban, J.; Dvořák, M. Sap flow-based quantitative indication of progression of Dutch elm disease after inoculation with Ophiostoma novo-ulmi. Trees 2014, 28, 1599–1605. [Google Scholar] [CrossRef]
- Menkis, A.; Östbrant, I.L.; Wågström, K.; Vasaitis, R. Dutch elm disease on the island of Gotland: Monitoring disease vector and combat measures. Scand. J. For. Res. 2016, 31, 237–241. [Google Scholar] [CrossRef]
- Flynn, L.E.; Mitchell, F.J.G. Comparison of a recent elm decline with the mid-Holocene Elm Decline. Veg. His. Archaeob. 2018, 1–8. [Google Scholar] [CrossRef]
- Martín, J.A.; Solla, A.; Ruiz-Villar, M.; Gil, L. Vessel length and conductivity of Ulmus branches: Ontogenetic changes and relation to resistance to Dutch elm disease. Trees 2013, 27, 1239–1248. [Google Scholar] [CrossRef]
- Gong, J.; Zhang, Z.; Zhang, C.; Zhang, J.; Ran, A. Ecophysiological responses of three tree species to a high-altitude environment in the Southeastern Tibetan Plateau. Forests 2018, 9, 48. [Google Scholar] [CrossRef]
- Sepúlveda, P.; Johnstone, D.M. A novel way of assessing plant vitality in urban trees. Forests 2019, 10, 2. [Google Scholar] [CrossRef]
- Bohn, U.; Gollub, G.; Hettwer, C.; Neuhäuslová, Z.; Raus, T.; Schlüter, H.; Weber, H. Karte der natürlichen Vegetation Europas Map of the Natural Vegetation of Europe Maßstab/Scale 1: 2 500 000; Bundesamt für Naturschutz (BfN)/Federal Agency for Nature Conservation: Bonn, Germany, 2004. [Google Scholar]
- Matuszkiewicz, J.M. Rola wiązów w zespołach roślinnych Polski [The role of the elms in plant associations in Poland]. In Wiązy [Elms]; Bugała, W., Boratyński, A., Iszkuło, G., Eds.; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2015; pp. 181–235. [Google Scholar]
- Forest Data Bank. Available online: https://www.bdl.lasy.gov.pl/portal/ (accessed on 5 October 2020).
- Hartmann, G.; Nienhaus, F.; Butin, H. Farbatlas Waldschäden: Diagnose von Baumkrankheiten; Eugen Ulmer KG: Stuttgart, Germany, 2007. [Google Scholar]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 6th ed.; The Iowa State University Press: Ames, IA, USA, 1976; pp. 327–329. [Google Scholar]
- Milewski, W. Lasy w Polsce 2017; Centrum Informacyjne Lasów Państwowych: Warszawa, Poland, 2017. Available online: http://www.lasy.gov.pl/pl/informacje/publikacje/do-poczytania/lasy-w-polsce-1/lasy-w-polsce-2017-pl-internet.pdf (accessed on 30 December 2017).
- Martín-Benito, D.; García-Vallejo, M.C.; Pajares, J.A.; López, D. Triterpenes in elms in Spain. Can. J. For. Res. 2005, 35, 199–205. [Google Scholar] [CrossRef]
- Filipiak, M.; Napierała-Filipiak, A.; Banacki, J. Wiąz górski (Ulmus glabra Huds.), wiąz szypułkowy (U. laevis Pall.) i wiąz polny (U. minor Mill.) na terenie Bałtyckiej Krainy Przyrodniczo-Leśnej [Wych elm (Ulmus glabra Huds.), European white elm (U. laevis Pall.) and field elm (U. minor Mill.) in the Bltic Natural Forest Region]. Acta Sci. Pol. Silv. Colendar. Ratio Ind. Lignar. 2018, 17, 333–344. [Google Scholar]
- Głaz, J. Wiąz w Lasach Państwowych. [Elm in the State Forests]. Sylwan 1986, 130, 23–33. [Google Scholar]
- Danielewicz, W. Ekologiczne Uwarunkowania Zasięgów drzew i Krzewów na Aluwialnych Obszarach doliny Odry. [Ecological Determinants of the Range of Trees and Shrubs in the Alluvial Areas of the Oder Valley]; Wydawnictwo Uniwersytetu Przyrodniczego: Poznań, Poland, 2008. [Google Scholar]
- Kosiński, P. Rozmieszczenie Oraz Warunki Występowania Drzew i Krzewów w Polskiej części Sudetów Wschodnich [Distribution and Conditions of Occurrence of Trees and Shrubs in the Polish Part of the Eastern Sudety Mts]; Acta Botanica Silesiaca, Monographiae 1, Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2007. [Google Scholar]
- Martínez-Arias, C.; Sobrino-Plata, J.; Macaya-Sanz, D.; Aguirre, N.M.; Collada, C.; Gil, L.; Martín, J.A.; Rodríguez-Calcerrada, J. Changes in plant function and root mycobiome caused by flood and drought in a riparian tree. Tree Phys. 2020, 40, 886–903. [Google Scholar] [CrossRef]
- Wawro, E. Winnice w Polsce [Vineyards in Poland]; Multico: Warszawa, Poland, 2015. [Google Scholar]
- Peterken, G.F.; Mountford, E.P. Long-term change in an unmanaged population of elm subjected to Dutch elm disease. J. Ecol. 1998, 86, 205–218. [Google Scholar] [CrossRef]
- Oheimb, G.; Brunet, J. Long-term vegetation changes in a south Swedish deciduous forest. Acta Oecol. 2007, 31, 229–242. [Google Scholar] [CrossRef]
- Modrý, M.; Hubený, D.; Rejšek, K. Differential response of naturally regenerated European shade tolerant tree species to soil type and light availability. For. Ecol. Manag. 2004, 188, 185–195. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
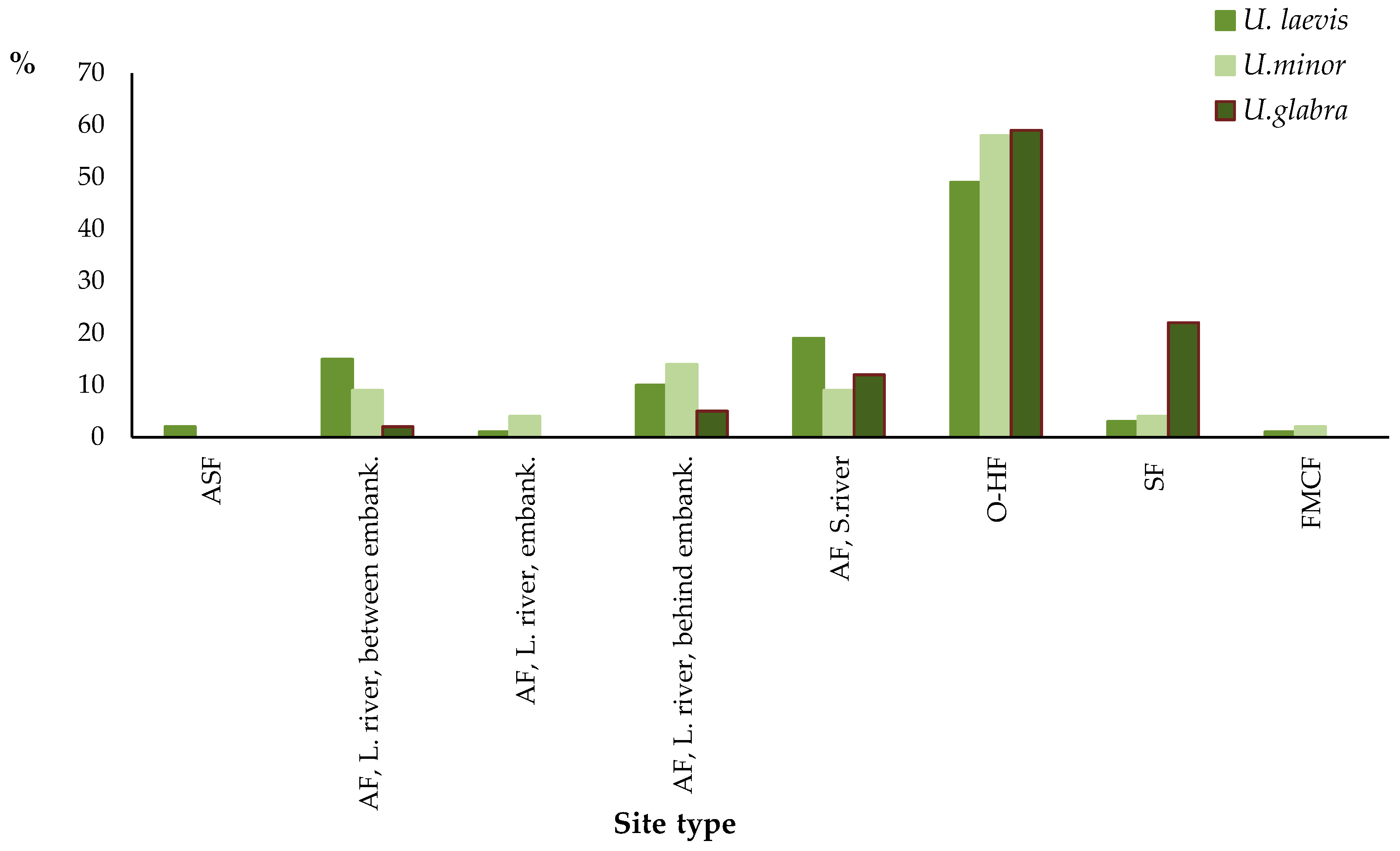{kind=link}
{kind=link}
{kind=link}
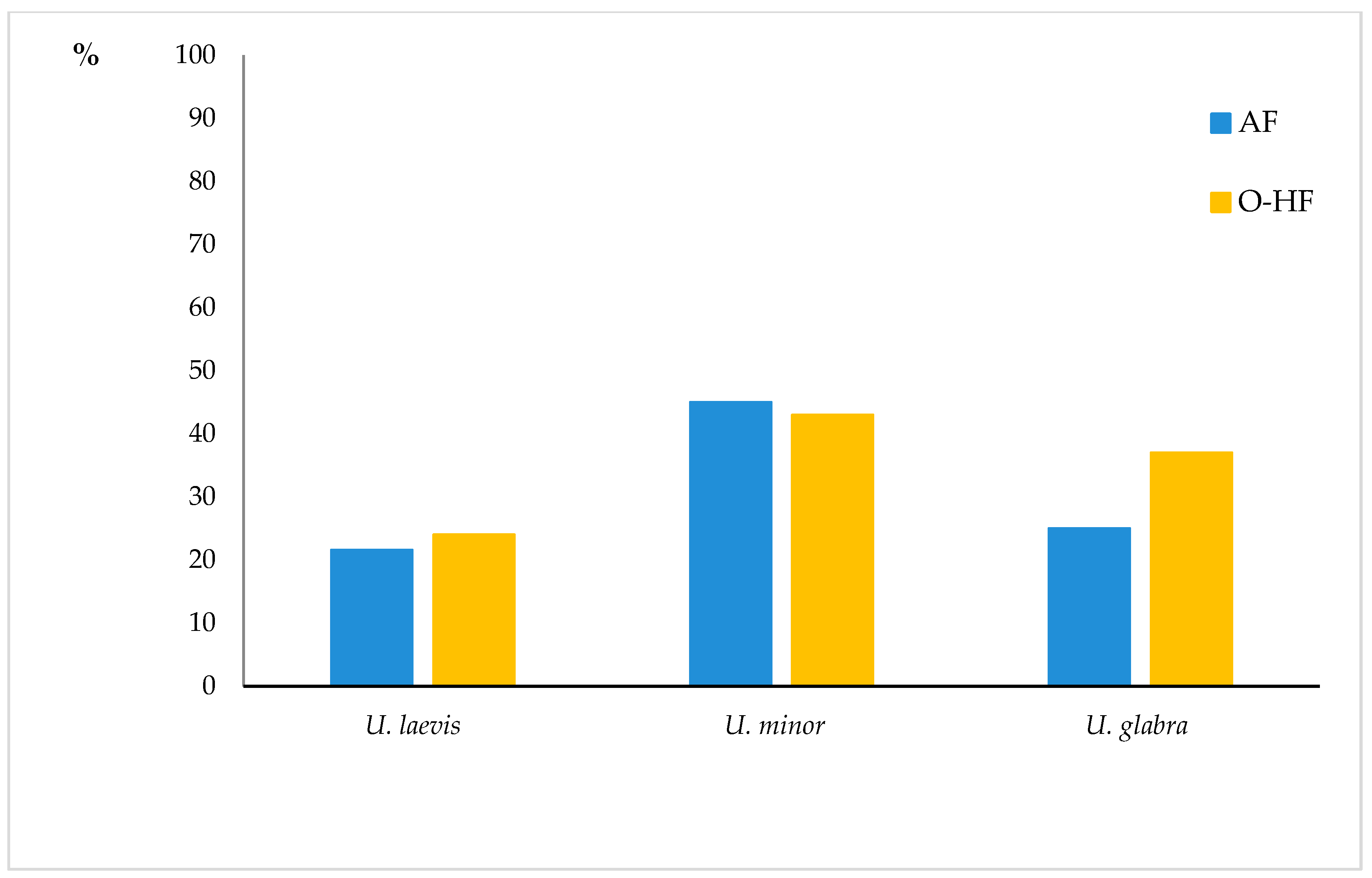{kind=link}
| Area (km2) | Forest Area (km2) | Forest Area (%) | Average. Annual Temperature (°C) | Vegetation Period (Days) | Average Annual Rainfall (mm) | Dominant Forest Site Types | Elevation m.a.s.l |
|---|---|---|---|---|---|---|---|
| 220,406 | 59,968 | 27 | 8.0 | 200–220 | 500–650 | FCF—43% FMCF—21% DCF—8.4% FMDF—11.9% FDF—8.9% MMCF—6% MMDF—5% MDF—2% AF—1% AAF—1%, ASF—1% * | 40–718 |
| Share of Elms in Forest Stands | No | Area (ha) | Reduced Area (ha) |
|---|---|---|---|
| <5% | 5523 | 21,863 | 546 |
| 5–10% | 23,412 | 95,258 | 7144 |
| 10% | 3552 | 10,830 | 1083 |
| 20% | 1092 | 3198 | 639 |
| 30% | 389 | 957 | 287 |
| 40% | 174 | 425 | 170 |
| 50% | 102 | 200 | 100 |
| 60% | 70 | 229 | 137 |
| 70% | 53 | 126 | 88 |
| 80% | 40 | 135 | 108 |
| 90% | 14 | 27 | 24 |
| 100% | 162 | 378 | 378 |
| FST | No. Ulmus | Wooded Area |
|---|---|---|
| ash-alder forest | 5.2 | 0.8 |
| alder forest | 3.0 | 1.5 |
| alluvial forest | 12.4 | 0.6 |
| moist d.f. | 18.0 | 2.0 |
| moist mixed d.f. | 7.4 | 4.9 |
| moist mixed c.f. | 1.1 | 6.3 |
| fresh d.f. | 27.5 | 10.1 |
| fresh mixed d.f. | 18.4 | 18.1 |
| fresh mixed c.f. | 5.8 | 28.3 |
| fresh c.f. | 1.1 | 27.5 |
| Chi test | ||
| chi-square | 786.69 | |
| degrees of freedom | 9 | |
| p-value | 0.000 | |
| Yats’ chi-square | 775.3 | |
| Yats’ p-value | 0.000 |
| Site Type | U. laevis | U. minor | U. glabra |
|---|---|---|---|
| ASF | 9 | 0 | 0 |
| AF, L. river, between embank. | 58 | 8 | 1 |
| AF, L. river, embank. | 5 | 3 | 0 |
| AF, L. river, behind embank. | 40 | 11 | 2 |
| AF, S. river | 73 | 7 | 5 |
| O-HF | 191 | 46 | 24 |
| SF | 11 | 3 | 9 |
| FMCF | 3 | 1 | 0 |
| Chi test | |||
| Chi-square | 49.99 | ||
| Degrees offreedom | 14 | ||
| p-value | 0.00 | ||
| Yets’ chi-square | 38.72 | ||
| Yets’ p-value | 0.00 |
| A | B | C | D | E | F | G | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Site Type | U. laevis | U. glabra | Site Type | U. laevis | U. minor | Site Type | U. laevis | U. minor | Habitat Type | U. laevis | U. minor | TSL | U. laevis | U.minor | U. glabra | TSL | U. laevis | U. glabra | TSL | U.minor | U. glabra |
| ASF | 9 | 0 | ASF | 9 | 0 | AF, L. river, between | 58 | 8 | Moist sites | 180 | 26 | slope forest | 9 | 3 | 11 | slope forest | 9 | 11 | slope forest | 3 | 11 |
| AF, L. river, between embank. | 58 | 1 | AF, L. river, between embank. | 58 | 8 | embank. | Fresh sites | 210 | 53 | other site types | 379 | 76 | 32 | other site types | 379 | 32 | other site types | 76 | 32 | ||
| AF, L. river, embank. | 5 | 0 | AF, L. river, embank. | 5 | 3 | AF, L. river, embank. | 5 | 3 | |||||||||||||
| AF, L. river, behind embank. | 40 | 2 | AF, L. river, behind embank. | 40 | 11 | AF, L. river, behind embank. | 40 | 11 | |||||||||||||
| AF, S. river | 73 | 5 | AF, S. river | 73 | 7 | AF, S. river | 73 | 7 | |||||||||||||
| O-HF | 191 | 24 | |||||||||||||||||||
| SF | 11 | 9 | |||||||||||||||||||
| FMCF | 3 | 0 | |||||||||||||||||||
| Chi Test | Chi Test | Chi Test | Chi Test | Chi Test | Chi Test | Chi Test | |||||||||||||||
| Chi-square | 37.89 | Chi-square | 9.82 | Chi-square | 8,05 | Chi-square | 4.67 | chi-square | 48.75 | chi-square | 47.34 | chi-square | 13.01 | ||||||||
| Degrees of freedom | 7 | Degrees of freedom | 4 | Degrees of freedom | 3 | Degrees of freedom | 1 | degrees of freedom | 2 | degrees of freedom | 1 | degrees of freedom | 1 | ||||||||
| p-value | 0 | p-value | 0.04 | p-value | 0.04 | p-value | 0.03 | p-value | 0 | p-value | 0 | p-value | 0 | ||||||||
| Yats’ chi-square | 30.62 | Yats’ chi-square | 6 | Yats’ chi-square | 5.25 | Yats’ chi-square | 4.155 | Yats’ chi-square | 43.41 | Yats’ chi-square | 42.23 | Yats’ chi-square | 10.95 | ||||||||
| Yats’ p-value | 0 | Yats’ p-value | 0.2 | Yats’ p-value | 0.15 | Yats’ p-value | 0.04 | Yats’ p-value | 0 | Yats’ p-value | 0 | Yats’ p-value | 3 | 0.001 | |||||||
| TSL | U. laevis | U.minor | U. glabra |
|---|---|---|---|
| ash-alder forest | 5 | 1 | 0 |
| alder forest | 4 | 0 | 0 |
| alluvial forest | 59 | 7 | 1 |
| moist d.f. | 74 | 12 | 4 |
| moist mixed d.f. | 35 | 7 | 2 |
| moist mixed c.f. | 8 | 0 | 1 |
| fresh mixed d.f. | 82 | 23 | 16 |
| fresh d.f. | 120 | 27 | 17 |
| fresh mixed c.f. | 3 | 2 | 0 |
| Chi test | |||
| chi-square | 22.285 | ||
| degrees offreedom | 16 | ||
| p-value | 0.1342 | ||
| Yats’ chi-square | 14.628 | ||
| Yats’ p-value | 0.552 |
| A | B | C | D | E | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TSL | U. laevis | U.minor | U. glabra | TSL | U. laevis | U. glabra | TSL | U. laevis | U.minor | U. glabra | TSL | U. laevis | U. minor | TSL | U. laevis | U. glabra |
| alluvial forest | 59 | 7 | 1 | alluvial forest | 59 | 1 | moist sites | 185 | 27 | 8 | moist sites | 185 | 27 | moist sites | 185 | 8 |
| fresh mixed d.f. and fresh d.f. | 202 | 50 | 33 | fresh mixed d.f. and fresh d.f. | 202 | 33 | fresh sites | 205 | 52 | 33 | fresh sites | 205 | 52 | fresh sites | 205 | 33 |
| Chi test | Chi test | Chi test | Chi test | Chi test | ||||||||||||
| chi-square | 9.651 | chi-square | 7.179 | chi-square | 14.853 | chi-square | 4.662 | chi-square | 11.699 | |||||||
| degrees of freedom | 2 | degrees of freedom | 1 | degrees of freedom | 2 | degrees of freedom | 1 | degrees of freedom | 1 | |||||||
| p-value | 0.0084 | p-value | 0.007 | p-value | 0.0006 | p-value | 0.0308 | p-value | 0.00063 | |||||||
| Yats’ chi-square | 7.929 | Yats’ chi-square | 6.017 | Yats’ chi-square | 13.39 | Yats’ chi-square | 4.142 | Yats’ chi-square | 10.597 | |||||||
| Yats’ p-value | 0.0189 | Yats’ p-value | 0.014 | Yats’ p-value | 0.0124 | Yats’ p-value | 0.0418 | Yats’ p-value | 0.00113 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Napierała-Filipiak, A.; Filipiak, M.; Jaworek-Jakubska, J. The Populations and Habitat Preferences of Three Elm Species in Conditions Prevailing on Plains of Poland. Forests 2021, 12, 162. https://doi.org/10.3390/f12020162
Napierała-Filipiak A, Filipiak M, Jaworek-Jakubska J. The Populations and Habitat Preferences of Three Elm Species in Conditions Prevailing on Plains of Poland. Forests. 2021; 12(2):162. https://doi.org/10.3390/f12020162
Chicago/Turabian StyleNapierała-Filipiak, Anna, Maciej Filipiak, and Justyna Jaworek-Jakubska. 2021. "The Populations and Habitat Preferences of Three Elm Species in Conditions Prevailing on Plains of Poland" Forests 12, no. 2: 162. https://doi.org/10.3390/f12020162
APA StyleNapierała-Filipiak, A., Filipiak, M., & Jaworek-Jakubska, J. (2021). The Populations and Habitat Preferences of Three Elm Species in Conditions Prevailing on Plains of Poland. Forests, 12(2), 162. https://doi.org/10.3390/f12020162