
Fire and Mechanical Forest Management Treatments Support Different Portions of the Bird Community in Fire-Suppressed Forests


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
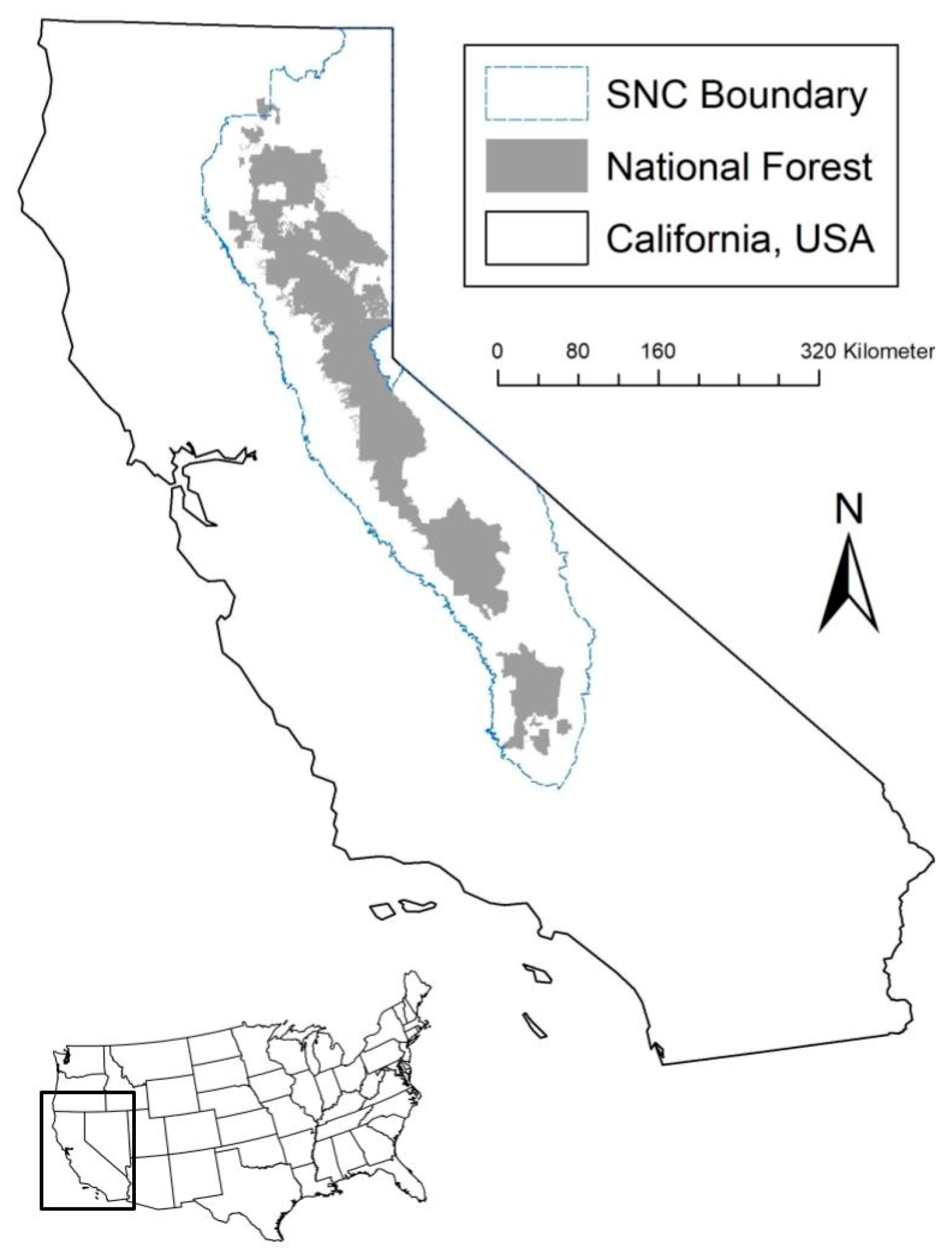
2.1. Study Location
2.2. Site Selection
2.3. Field Surveys
2.4. Bird Abundance Analyses
3. Results
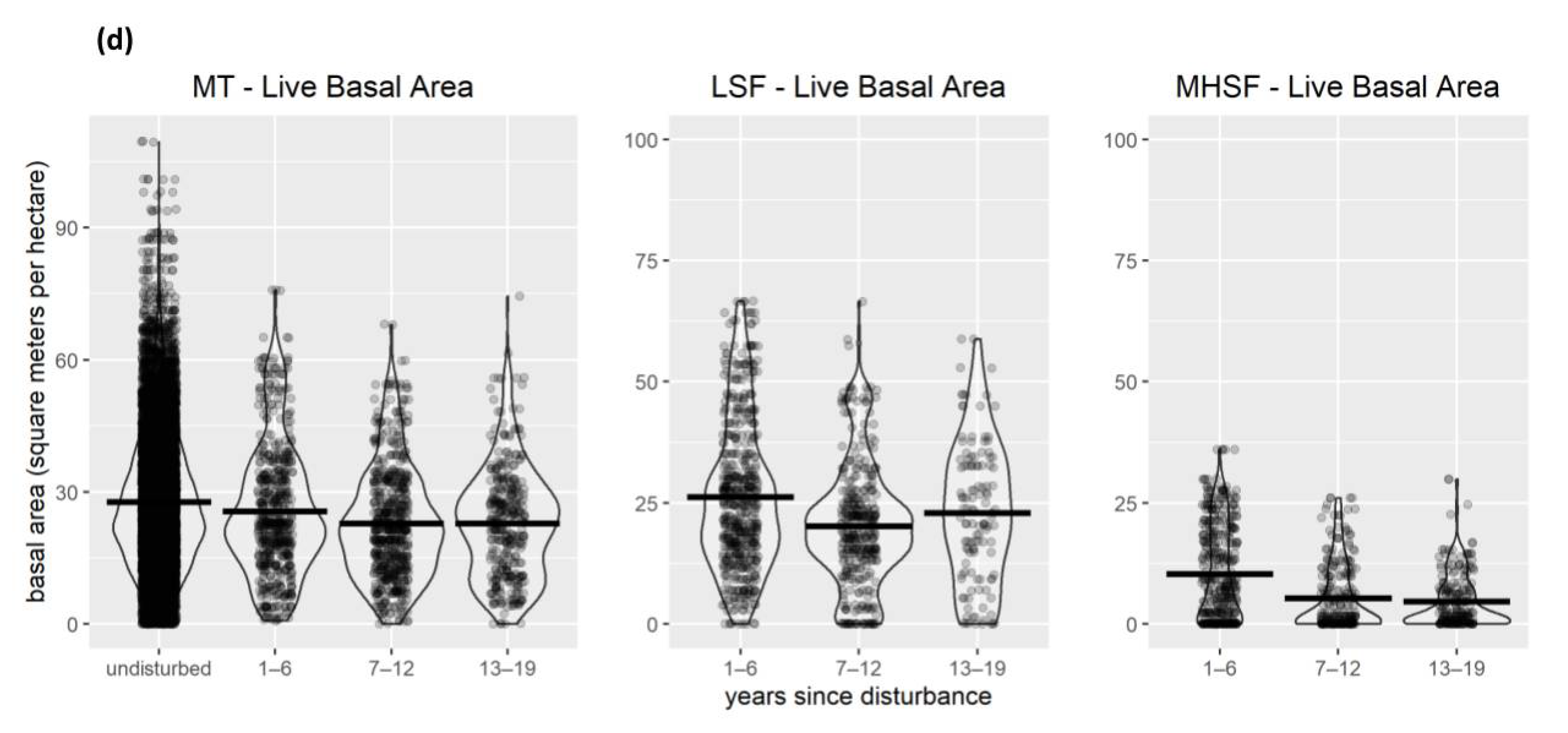
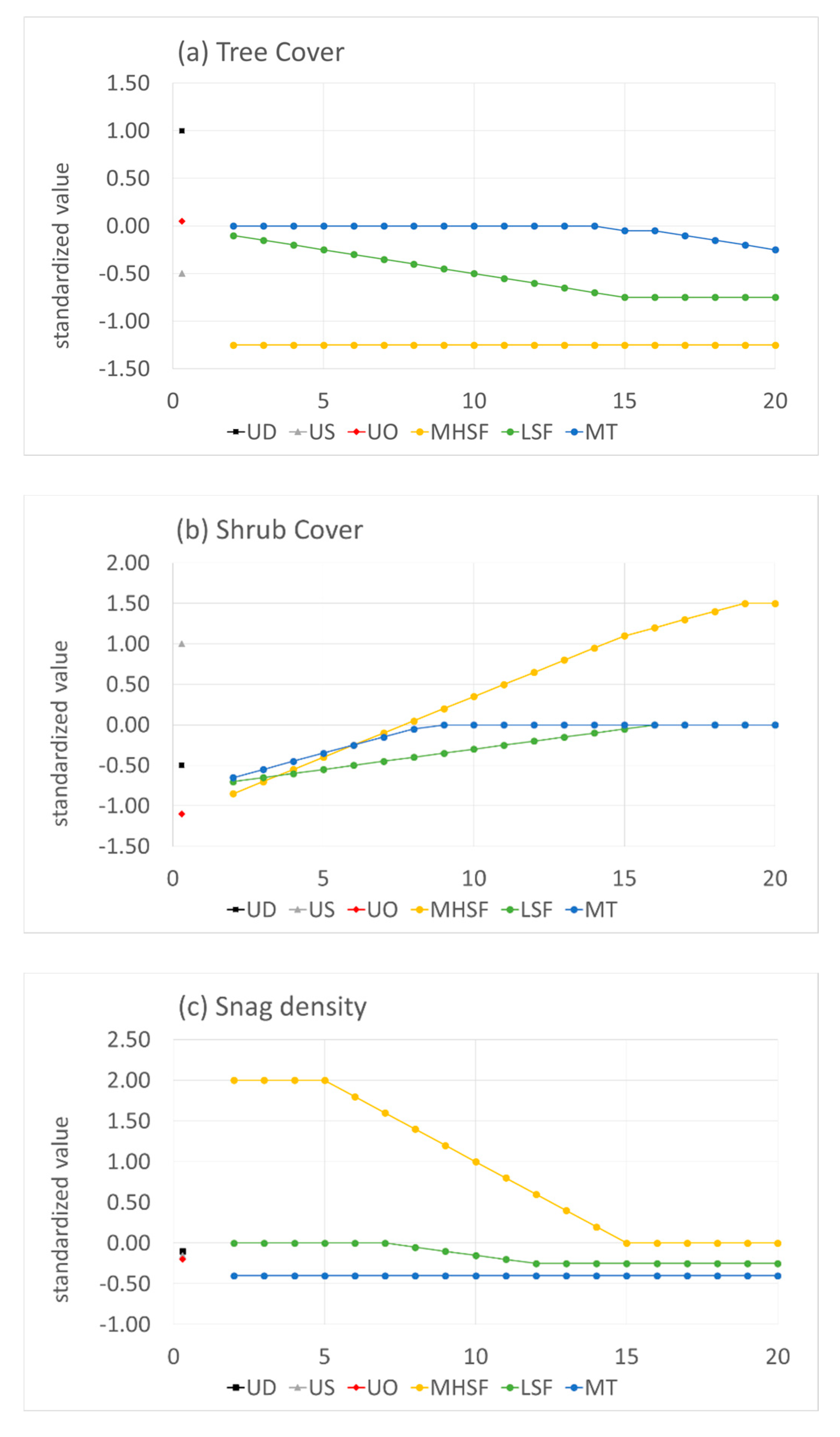
3.1. Vegetation Changes Following Disturbance
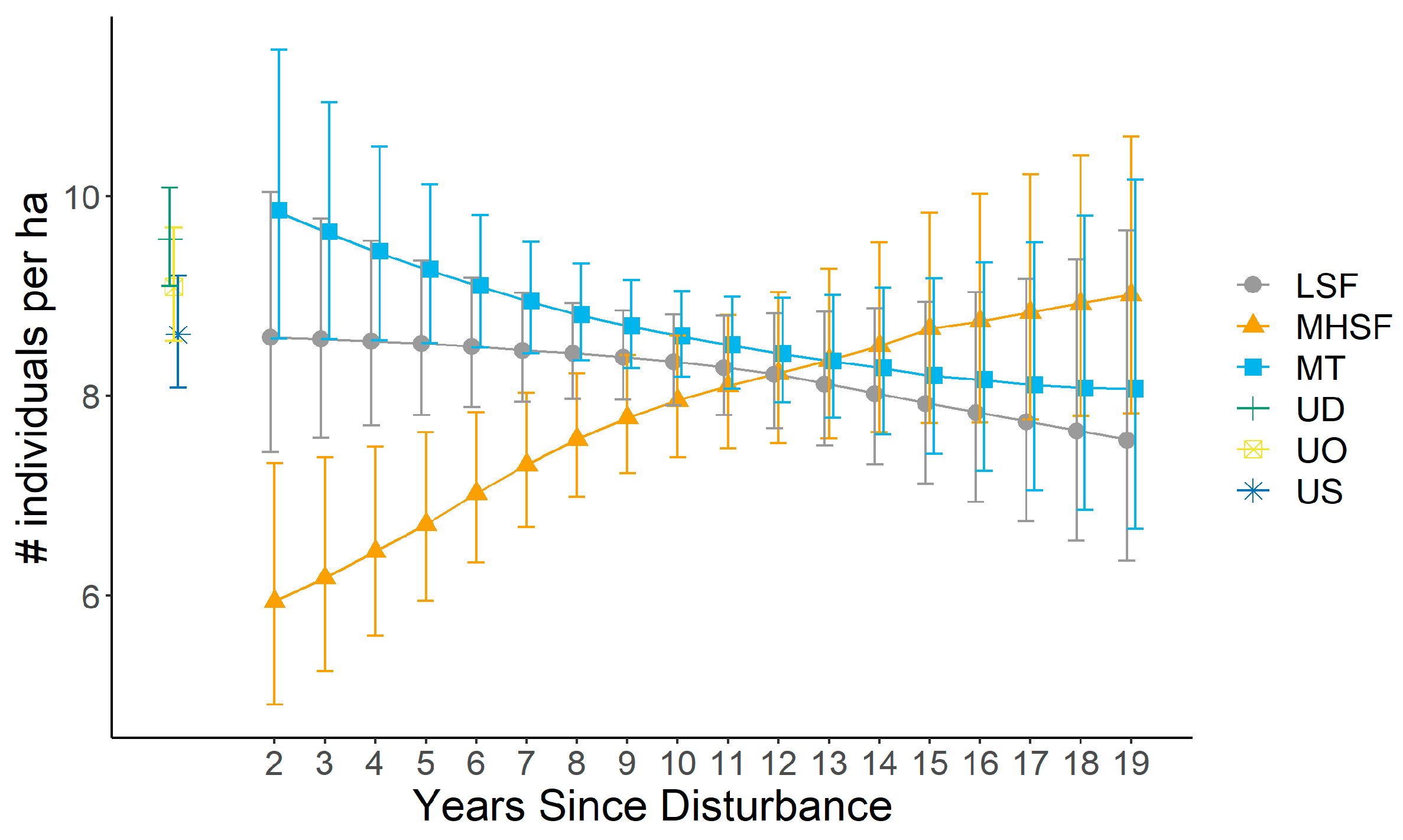
3.2. Bird Abundance and Response to Disturbance
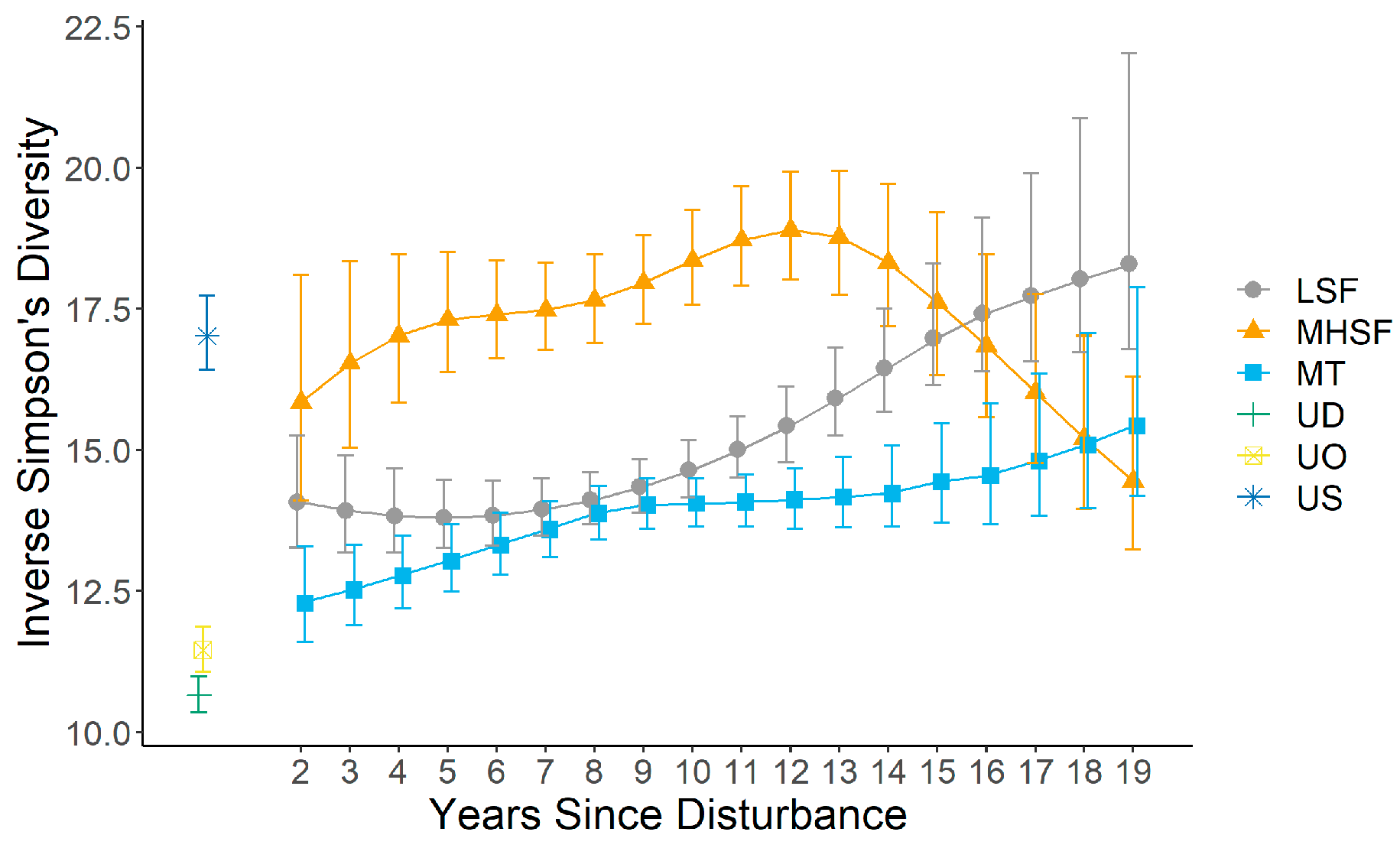
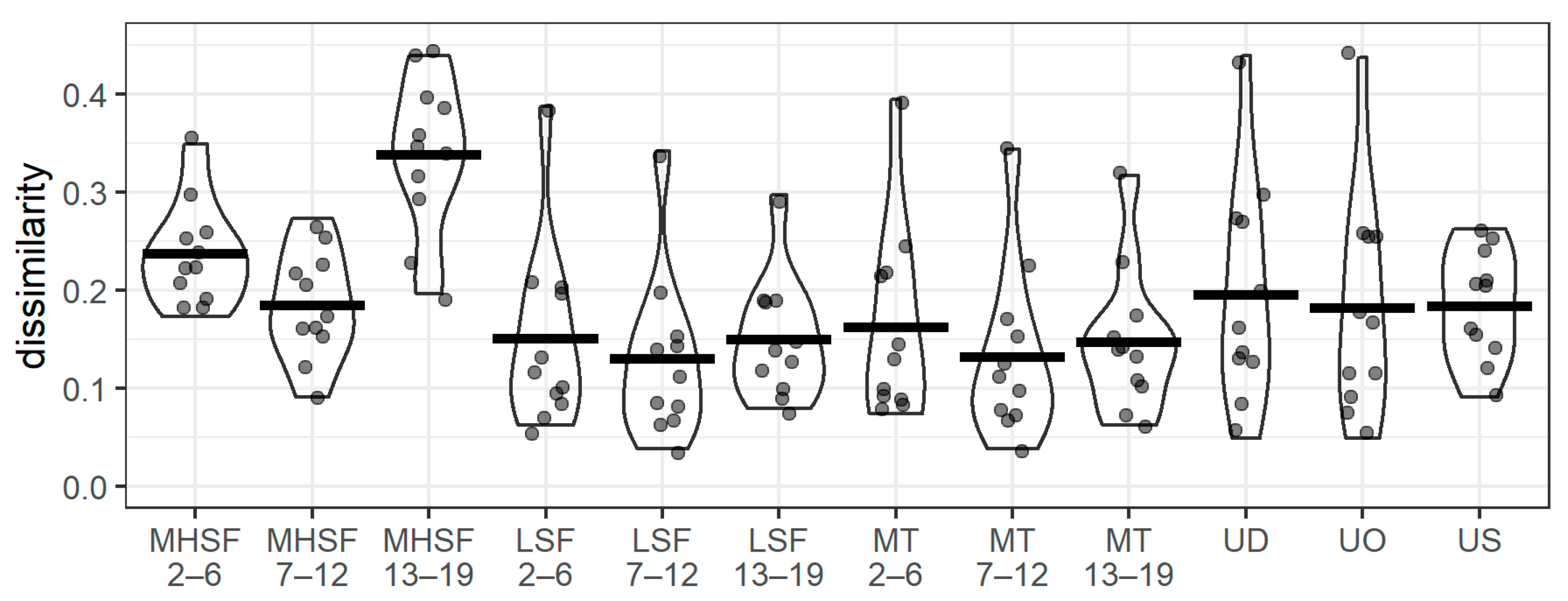
3.3. Community Similarity
4. Discussion
4.1. The Importance of Fire
4.2. Managing Disturbances over Time
4.3. Disturbance in a Disturbance-Starved System
4.4. Analysis Assumptions and Unmodeled Influences on Avian Abundance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Lindenmayer, D.B.; Laurance, W.F. A history of hubris—Cautionary lessons in ecologically sustainable forest management. Biol. Cons. 2012, 151, 11–16. [Google Scholar] [CrossRef]
- O’Hara, K. What is close-to-nature silviculture in a changing world? Forestry 2016, 89, 1–6. [Google Scholar] [CrossRef]
- Randhir, T.O.; Erol, A. Emerging Threats to Forests: Resilience and Strategies at System Scale. Am. J. Plant. Sci. 2013, 4, 739–748. [Google Scholar] [CrossRef]
- FAO and UNEP. The State of the World’s Forests 2020. Forests, Biodiversity and People. Rome. 2020. Available online: https://doi.org/10.4060/ca8642en (accessed on 21 December 2020).
- Turner, M.G.; Donato, D.C.; Romme, W.H. Consequences of spatial heterogeneity for ecosystem services in changing forest landscapes: Priorities for future research. Landsc. Ecol. 2013, 28, 1081–1097. [Google Scholar] [CrossRef]
- Churchill, D.J.; Larson, A.J.; Dahlgreen, M.C.; Franklin, J.F.; Hessburg, P.F.; Lutz, J.A. Restoring forest resilience: From reference spatial patterns to silvicultural prescriptions and monitoring. For. Ecol. Manag. 2013, 291, 442–457. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Allen, C.D.; Franklin, J.F.; Frelich, L.E.; Harvey, B.J.; Higuera, P.E.; Mack, M.C.; Meentemeyer, R.K.; Metz, M.R.; Perry, G.L.; et al. Changing disturbance regimes, ecological memory, and forest resilience. Front. Ecol. Environ. 2016, 14, 369–378. [Google Scholar] [CrossRef]
- Stephens, S.L.; Collins, B.M.; Fettig, C.J.; Finney, M.A.; Hoffman, C.M.; Knapp, E.E.; North, M.P.; Safford, H.; Wayman, R.B. Drought, Tree Mortality, and Wildfire in Forests Adapted to Frequent Fire. BioScience 2018, 68, 77–88. [Google Scholar] [CrossRef]
- Agee, J.K. The landscape ecology of western forest fire regimes. Northwest. Sci. 1998, 72, 24–34. [Google Scholar]
- Kennedy, P.L.; Fontaine, J.B. Synthesis of Knowledge on the Effects of Fire and Fire Surrogates on Wildlife in US Dry Forests. Oregon State University Agricultural Experiment Station Special Report 1096. 2009. Available online: http://ir.library.oregonstate.edu/jspui/bitstream/1957/12625/1/SR1096.pdf (accessed on 20 December 2020).
- Skinner, C.N.; Chang, C. Fire regimes, past and present. In Sierra Nevada Ecosystem Project: Final Report to Congress, Vol. II, Assessments and Scientific Basis for Management Options; Water Resources Center Report 37; Centers for Water and Wildland Resources, University of California: Davis, CA, USA, 1996; pp. 1041–1069. [Google Scholar]
- Steel, Z.L.; Safford, H.D.; Viers, J.H. The fire frequency-severity relationship and the legacy of fire suppression in California forests. Ecosphere 2015, 6, 1–23. [Google Scholar] [CrossRef]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western US forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef]
- Miller, J.D.; Safford, H.D. Trends in wildfire severity: 1984 to 2010 in the Sierra Nevada, Modoc Plateau, and southern Cascades, California, USA. Fire Ecol. 2012, 8, 41–57. [Google Scholar] [CrossRef]
- Taylor, A.H. Fire regimes and forest changes in mid and upper montane forests of the southern cascades, Lassen Volcanic National Park, California, USA. J. Biogeogr. 2000, 27, 87–104. [Google Scholar] [CrossRef]
- Stephens, S.L.; Martin, R.E.; Clinton, N.E. Prehistoric fire area and emissions from California forests, woodlands, shrublands, and grasslands. For. Ecol. Manag. 2007, 251, 205–216. [Google Scholar] [CrossRef]
- Williams, M.A.; Baker, W.L. Spatially extensive reconstructions show variable-severity fire and heterogeneous structure in historical western United States dry forests. Glob. Ecol. Biogeog. 2012, 21, 1042–1052. [Google Scholar] [CrossRef]
- Mallek, C.; Safford, H.; Viers, J.; Miller, J. Modern departures in fire severity and area vary by forest type, Sierra Nevada and southern Cascades, California, USA. Ecosphere 2013, 4, 1–28. [Google Scholar] [CrossRef]
- Parsons, D.J.; DeBenedetti, S.H. Impact of fire suppression on a mixed-conifer forest. For. Ecol. Manag. 1979, 2, 21–33. [Google Scholar] [CrossRef]
- Minnich, R.A.; Barbour, M.G.; Burk, J.H.; Fernau, R.F. Sixty years of change in California conifer forests of the San Bernardino mountains. Cons. Biol. 1995, 9, 902–914. [Google Scholar] [CrossRef]
- Barbour, M.; Kelley, E.; Maloney, P.; Rizzo, D.; Royce, E.; Fites-Kaufman, J. Present and past old growth forests of the Lake Tahoe Basin, Sierra Nevada, USA. J. Veg. Sci. 2002, 13, 461–472. [Google Scholar] [CrossRef]
- Vankat, J.L.; Major, J. Vegetation changes in Sequoia National Park, California. J. Biogeogr. 1978, 5, 377–402. [Google Scholar] [CrossRef]
- Nagel, T.A.; Taylor, A.H. Fire and persistence of montane chaparral in mixed conifer forest landscapes in the northern Sierra Nevada, Lake Tahoe Basin, California, USA. J. Torrey Bot. Soc. 2005, 132, 442–457. [Google Scholar] [CrossRef]
- Rogers, P.C.; Shepperd, W.D.; Bartos, D.L. Aspen in the Sierra Nevada: Regional conservation of a continental species. Nat. Areas J. 2007, 27, 183–193. [Google Scholar] [CrossRef]
- Beaty, R.M.; Taylor, A.H. Fire history and the structure and dynamics of a mixed conifer forest landscape in the northern Sierra Nevada, Lake Tahoe Basin, California, USA. For. Ecol. Manag. 2005, 255, 707–719. [Google Scholar] [CrossRef]
- Collins, B.M.; Everett, R.G.; Stephens, S.L. Impacts of fire exclusion and recent managed fire on forest structure in old growth Sierra Nevada mixed-conifer forests. Ecosphere 2011, 2, 1–14. [Google Scholar] [CrossRef]
- Taillie, P.J.; Burnett, R.D.; Roberts, L.J.; Campos, B.R.; Peterson, M.N.; Moorman, C.E. Interacting and non-linear avian responses to mixed-severity wildfire and time since fire. Ecosphere 2018, 9, e02291. [Google Scholar] [CrossRef]
- Steel, Z.L.; Campos, B.R.; Frick, W.F.; Burnett, R.D.; Safford, H.D. The effects of wildfire severity and pyrodiversity on bat occupancy and diversity in fire-suppressed forests. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Brawn, J.D.; Robinson, S.K.; Thompson III, F.R. The role of disturbance in the ecology and conservation of birds. Ann. Rev. Ecol. Syst. 2001, 32, 251–276. [Google Scholar] [CrossRef]
- Smucker, K.M.; Hutto, R.L.; Steele, B.M. Changes in bird abundance after wildfire: Importance of fire severity and time since fire. Ecol. Appl. 2005, 15, 1535–1549. [Google Scholar] [CrossRef]
- Tingley, M.W.; Ruiz-Gutiérrez, V.; Wilkerson, R.L.; Howell, C.A.; Siegel, R.B. Pyrodiversity promotes avian diversity over the decade following forest fire. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161703. [Google Scholar] [CrossRef]
- Hutto, R.L. The Ecological Importance of Severe Wildfires: Some like It Hot. Ecol. Appl. 2008, 18, 1827–1834. [Google Scholar] [CrossRef]
- Miller, J.D.; Knapp, E.E.; Key, C.H.; Skinner, C.N.; Isbell, C.J.; Creasy, R.M.; Sherlock, J.W. Calibration and validation of the relative differenced Normalized Burn Ratio (RdNBR) to three measures of fire severity in the Sierra Nevada and Klamath Mountains, California, USA. Remote Sens. Environ. 2009, 113, 645–656. [Google Scholar] [CrossRef]
- Agee, J.K.; Skinner, C.N. Basic principles of forest fuel reduction treatments. For. Ecol. Manag. 2005, 211, 83–96. [Google Scholar] [CrossRef]
- Driscoll, D.A.; Lindenmayer, D.B.; Bennett, A.F.; Bode, M.; Bradstock, R.A.; Cary, G.J.; Clarke, M.F.; Dexter, N.; Fensham, R.; Friend, G.; et al. Fire management for biodiversity conservation: Key research questions and our capacity to answer them. Biol. Cons. 2010, 143, 1928–1939. [Google Scholar] [CrossRef]
- Kelly, L.T.; Brotons, L. Using fire to promote biodiversity. Science 2017, 355, 1264–1265. [Google Scholar] [CrossRef] [PubMed]
- DellaSala, D.A.; Bond, M.L.; Hanson, C.T.; Hutto, R.L.; Odion, D.C. Complex early seral forests of the Sierra Nevada: What are they and how can they be managed for ecological integrity? Nat. Areas J. 2014, 34, 310–324. [Google Scholar] [CrossRef]
- Hutto, R.L.; Patterson, D.A. Positive effects of fire on birds may appear only under narrow combinations of fire severity and time-since-fire. Int. J. Wildland Fire 2016, 25, 1074–1085. [Google Scholar] [CrossRef]
- Hutto, R.L.; Hutto, R.R.; Hutto, P.L. Patterns of bird species occurrence in relation to anthropogenic and wildfire disturbance: Management implications. For. Ecol. Manag. 2020, 461, 117942. [Google Scholar] [CrossRef]
- Noss, R.F.; Franklin, J.F.; Baker, W.L.; Schoennagel, T.; Moyle, P.B. Managing fire-prone forests in the western United States. Front. Ecol. Environ. 2006, 4, 481–487. [Google Scholar] [CrossRef]
- Patry, C.; Kneeshaw, D.; Wyatt, S.; Grenon, F.; Messier, C. Forest ecosystem management in North America: From theory to practice. For. Chron. 2013, 89, 525–537. [Google Scholar] [CrossRef]
- Arno, S.F.; Fiedler, C.E. Mimicking Nature’s Fire: Restoring Fire-Prone Forests in the West; Island Press: Washington, DC, USA, 2005; p. 256. [Google Scholar]
- Long, J.N. Emulating natural disturbance regimes as a basis for forest management: A North American view. For. Ecol. Manag. 2009, 257, 1868–1873. [Google Scholar] [CrossRef]
- North, M.P.; Keeton, W.S. Emulating natural disturbance regimes: An emerging approach for sustainable forest management. In Patterns and Processes in Forest Landscapes; Springer: Berlin/Heidelberg, Germany, 2008; pp. 341–372. [Google Scholar]
- North, M.P.; Stine, P.; O’Hara, K.; Zielinski, W.; Stephens, S. An Ecosystem Management Strategy for Sierran Mixed-Conifer Forests; General Technical Report PSW-GTR-220; US Department of Agriculture, Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2009.
- Hurteau, S.R.; Sisk, T.D.; Block, W.M.; Dickson, B.G. Fuel-reduction treatment effects on avian community structure and diversity. J. Wildl. Manag. 2008, 72, 1168–1174. [Google Scholar] [CrossRef]
- Verschuyl, J.; Riffell, S.; Miller, D.; Wigley, T.B. Biodiversity response to intensive biomass production from forest thinning in North American forests–a meta-analysis. For. Ecol. Manag. 2011, 261, 221–232. [Google Scholar] [CrossRef]
- White, A.M.; Zipkin, E.F.; Manley, P.N.; Schlesinger, M.D. Simulating avian species and foraging group responses to fuel reduction treatments in coniferous forests. For. Ecol. Manag. 2013, 304, 261–274. [Google Scholar] [CrossRef]
- Stephens, S.L.; Bigelow, S.W.; Burnett, R.D.; Collins, B.M.; Gallagher, C.V.; Keane, J.; Kelt, D.A.; North, M.P.; Roberts, L.J.; Stine, P.A.; et al. California spotted owl, songbird, and small mammal responses to landscape fuel treatments. BioScience 2014, 64, 893–906. [Google Scholar] [CrossRef]
- Hutto, R.L.; Keane, R.E.; Sherriff, R.L.; Rota, C.T.; Eby, L.A.; Saab, V.A. 2016. Toward a more ecologically informed view of severe forest fires. Ecosphere 2016, 7, 1–13. [Google Scholar] [CrossRef]
- Stephens, S.L.; Moghaddas, J.J. Silvicultural and reserve impacts on potential fire behavior and forest conservation: Twenty-five years of experience from Sierra Nevada mixed conifer forests. Biol. Cons. 2005, 125, 369–379. [Google Scholar] [CrossRef]
- Stephens, S.L.; Moghaddas, J.J. Experimental fuel treatment impacts on forest structure, potential fire behavior, and predicted tree mortality in a California mixed conifer forest. For. Ecol. Manag. 2005, 215, 21–36. [Google Scholar] [CrossRef]
- Stephens, S.L.; Moghaddas, J.J.; Edminster, C.; Fiedler, C.E.; Haase, S.; Harrington, M.; Keeley, J.E.; Knapp, E.E.; McIver, J.D.; Metlen, K.; et al. Fire treatment effects on vegetation structure, fuels, and potential fire severity in western U.S. forests. Ecol. Appl. 2009, 19, 305–320. [Google Scholar] [CrossRef]
- Collins, B.M.; Moghaddas, J.J.; Stephens, S.L. Initial changes in forest structure and understory plant communities following fuel reduction activities in a Sierra Nevada mixed conifer forest. For. Ecol. Manag. 2007, 239, 102–111. [Google Scholar] [CrossRef]
- Collins, B.M.; Stephens, S.L.; Moghaddas, J.J.; Battles, J. Challenges and approaches in planning fuel treatments across fire-excluded forested landscapes. J. For. 2010, 108, 24–31. [Google Scholar]
- Askins, R.A. Restoring forest disturbances to sustain populations of shrubland birds. Restor. Manag. Notes 1998, 16, 166–173. [Google Scholar] [CrossRef]
- Swanson, M.E.; Franklin, J.F.; Beschta, R.L.; Crisafulli, C.M.; DellaSala, D.A.; Hutto, R.L.; Lindenmayer, D.B.; Swanson, F.J. The forgotten stage of forest succession: Early-successional ecosystems on forest sites. Front. Ecol. Environ. 2011, 9, 117–125. [Google Scholar] [CrossRef]
- North, M.P.; Stephens, S.L.; Collins, B.M.; Agee, J.K.; Aplet, G.; Franklin, J.F.; Fulé, P.Z. Reform forest fire management. Science 2015, 349, 1280–1281. [Google Scholar] [CrossRef] [PubMed]
- North, M.P.; Brough, A.; Long, J.; Collins, B.M.; Bowden, P.; Yasuda, D.; Miller, J.; Sugihara, N. Constraints on mechanized treatment significantly limit mechanical fuels reduction extent in the Sierra Nevada. J. For. 2015, 113, 40–48. [Google Scholar] [CrossRef]
- Moreira, F.; Ascoli, D.; Safford, H.; Adams, M.A.; Morena, J.M.; Pereira, J.M.C.; Catry, F.X.; Armesto, J.; Bond, W.; Gonzalez, M.E.; et al. Wildfire Management in Mediterranean-type regions: Paradigm change needed. Environ. Res. Lett. 2020, 15, 011001. [Google Scholar] [CrossRef]
- McRae, D.J.; Duchesne, L.C.; Freedman, B.; Lynham, T.J.; Woodley, S. Comparisons between wildfire and forest harvesting and their implications in forest management. Environ. Rev. 2001, 9, 223–260. [Google Scholar] [CrossRef]
- Reich, P.B.; Bakken, P.; Carlson, D.; Frelich, L.E.; Friedman, S.K.; Grigal, D.F. Influence of logging, fire, and forest type on biodiversity and productivity in southern boreal forests. Ecology 2001, 82, 2731–2748. [Google Scholar] [CrossRef]
- Fontaine, J.B.; Kennedy, P.L. Meta-analysis of avian and small-mammal response to fire severity and fire surrogate treatments in US fire-prone forests. Ecol. Appl. 2012, 22, 1547–1561. [Google Scholar] [CrossRef]
- Fontaine, J.B.; Donato, D.C.; Robinson, W.D.; Law, B.E.; Kauffman, J.B. Bird communities following high-severity fire: Response to single and repeat fires in a mixed-evergreen forest, Oregon, USA. For. Ecol. Manag. 2009, 257, 1496–1504. [Google Scholar] [CrossRef]
- Roberts, L.J.; Burnett, R.D.; Fogg, A.M.; Geupel, G.R. PRBO MIS Final Study Plan and Sampling Protocols for Mountain Quail, Hairy Woodpecker, Fox Sparrow, and Yellow Warbler; PRBO Contribution 1714; PRBO Conservation Science: Petaluma, CA, USA, 2011. [Google Scholar]
- Sierra Nevada Avian Monitoring Information Network. Available online: https://data.prbo.org/apps/snamin/ (accessed on 3 November 2020).
- Stevens, D.L.; Olsen, A.R. Spatially balanced sampling of natural resources. J. Am. Stat. Assoc. 2004, 99, 262–278. [Google Scholar] [CrossRef]
- Theobald, D.M.; Stevens, D.L.; White, D.; Urquhart, N.S.; Olsen, A.R.; Norman, J.B. Using GIS to generate spatially balanced random survey designs for natural resource applications. Environ. Manag. 2007, 40, 134–146. [Google Scholar] [CrossRef]
- US Department of Agriculture, Forest Service. Pacific Southwest Region Past 20 Years Accomplishments—(FACTS). Vector Digital Data. 2017. Available online: https://www.fs.usda.gov/detail/r5/landmanagement/gis/?cid=stelprd3811519 (accessed on 9 November 2019).
- USDA Forest Service. Pacific Southwest Region Fire, Fuels and Aviation Management. VegBurnSeverity 18_1 Vector Digital Data. 2018. Available online: https://www.fs.usda.gov/detail/r5/landmanagement/gis/?cid=stelprd3805100 (accessed on 11 February 2020).
- North, M.P.; Schwartz, M.W.; Collins, B.M.; Keane, J.J. Current and Projected Condition of Mid-Elevation Sierra Nevada Forests; General Technical Report PSW-GTR-254; US Department of Agriculture, Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2017; pp. 109–157.
- Ralph, C.J.; Geupel, G.R.; Pyle, P.; Martin, T.E.; DeSante, D.F. Field Methods for Monitoring Landbirds; General Technical Report PSW-GTR 144; US Department of Agriculture, Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 1993.
- Ralph, C.J.; Droege, S.; Sauer, J.R. Managing and monitoring birds using point counts: Standards and applications. In Monitoring Bird Populations by Point Counts; Ralph, C.J., Sauer, J.R., Droege, S., Eds.; General Technical Report PSW-GTR 149; US Department of Agriculture, Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 1995; pp. 161–169. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Fiske, I.; Chandler, R. Unmarked: An R Package for Fitting Hierarchical Models of Wildlife Occurrence and Abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 28 May 2020).
- Burnett, R.D.; Roberts, L.J. A Quantitative Evaluation of the Conservation Umbrella of Spotted Owl Management Areas in the Sierra Nevada. PLoS ONE 2015, 10, e0123778. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.J.; Burnett, R.; Tietz, J.; Veloz, S. Recent drought and tree mortality effects on the avian community in southern Sierra Nevada: A glimpse of the future? Ecol. Appl. 2019, 29, e01848. [Google Scholar] [CrossRef] [PubMed]
- Sierra Nevada Ecosystem Project. Sierra Nevada Region Digital Elevation Model. 1999. Available online: www.ice.ucdavis.edu/snep/dataset.asp?dataset=318 (accessed on 15 March 2013).
- Flint, L.E.; Flint, A.L.; Thorne, J.H.; Boynton, R. Fine-scale hydrologic modeling for regional landscape applications: The California Basin Characterization Model development and performance. Ecol. Process. 2013, 2, 25. [Google Scholar] [CrossRef]
- Flint, L.E.; Flint, A.L. California Basin Characterization Model: A Dataset of Historical and Future Hydrologic Response to Climate Change. U.S. Geological Survey Data Release. 2014. Available online: https://doi.org/10.5066/f76t0jpb (accessed on 16 September 2019).
- Flint, L.E.; Flint, A.L.; Thorne, J.H.; Boynton, R. California BCM (Basin Characterization Model) Downscaled Climate and Hydrology—30-year Summaries. California Climate Commons. 2014. Available online: http://cida.usgs.gov/thredds/catalog/CA-BCM-2014/catalog.html (accessed on 16 September 2019).
- Van Gunst, K.J.; Weisberg, P.J.; Yang, J.; Fan, Y. Do denser forests have greater risk of tree mortality: A remote sensing analysis of density-dependent forest mortality. For. Ecol. Manag. 2016, 359, 19–32. [Google Scholar] [CrossRef]
- Gao, B.C. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Heiberger, R.H. HH: Statistical Analysis and Data Display: Heiberger and Holland. R Package Version 3.1-34. 2017. Available online: https://CRAN.R-project.org/package=HH (accessed on 12 December 2019).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-1. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 28 May 2020).
- Morris, E.K.; Caruso, T.; Buscot, F.; Fischer, M.; Hancock, C.; Maier, T.S.; Meiners, T.; Müller, C.; Obermaier, E.; Prati, D.; et al. Choosing and using diversity indices: Insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef]
- McKelvey, K.S.; Johnston, J.D. Historical perspectives on forests of the Sierra Nevada and the transverse ranges of southern California; forest conditions at the turn of the century. In The California Spotted Owl: A Technical Assessment of Its Current Status; General Technical Report PSW-GTR-133; USDA Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 1996; pp. 225–246. [Google Scholar]
- Franklin, J.F.; Johnson, K.N. A restoration framework for federal forests in the Pacific Northwest. J. For. 2012, 110, 429–439. [Google Scholar] [CrossRef]
- Fontaine, J.B. Influences of High Severity Fire and Postfire Salvage Logging on Avian and Small Mammal Communities of the Siskiyou Mountains, Oregon, USA. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 2007. [Google Scholar]
- North, M.P.; Stevens, J.T.; Greene, D.F.; Coppoletta, M.; Knapp, E.E.; Latimer, A.M.; Restaino, C.M.; Tompkins, R.E.; Welch, K.R.; York, R.A.; et al. Tamm Review: Reforestation for resilience in dry western US forests. For. Ecol. Manag. 2019, 432, 209–224. [Google Scholar] [CrossRef]
- Betts, M.G.; Hagar, J.C.; Rivers, J.W.; Alexander, J.D.; McGarigal, K.; McComb, B.C. Thresholds in forest bird occurrence as a function of the amount of early-seral broadleaf forest at landscape scales. Ecol. Appl. 2010, 20, 2116–2130. [Google Scholar] [CrossRef]
- Raphael, M.G.; White, M. Use of snags by cavity-nesting birds in the Sierra Nevada. Wildl. Monog. 1984, 86, 3–66. [Google Scholar]
- Latif, Q.S.; Saab, V.A.; Dudley, J.G.; Markus, A.; Mellen-McLean, K. Development and evaluation of habitat suitability models for nesting white-headed woodpecker (Dryobates albolarvatus) in burned forest. PLoS ONE 2020, 15, e0233043. [Google Scholar] [CrossRef] [PubMed]
- Stillman, A.N.; Siegel, R.B.; Wilkerson, R.L.; Johnson, M.; Tingley, M.W. Age-dependent habitat relationships of a burned forest specialist emphasise the role of pyrodiversity in fire management. J. Appl. Ecol. 2019, 56, 880–890. [Google Scholar] [CrossRef]
- Steel, Z.L.; Fogg, A.M.; Roberts, L.J.; Burnett, R.D.; Safford, H.D. When bigger isn’t better—Implications of large high-severity wildfire patches on avian diversity and community composition. Divers. Dist. 2021. [Google Scholar] [CrossRef]
- Shea, P.J.; Laudenslayer, W.F.; Ferrell, G.; Borys, R. Girdled versus bark beetle-created ponderosa pine snags: Utilization by cavity-dependent species and differences in decay rate and insect diversity. In Proceedings of the Symposium on the Ecology and Management of Dead Wood in Western Forests, Reno, NV, USA, 2–4 November 1999; General Technical Report PSW-GTR-181. USDA Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2002; pp. 145–153. [Google Scholar]
- Rota, C.T.; Rumble, M.A.; Lehman, C.P.; Kesler, D.C.; Millspaugh, J.J. Apparent foraging success reflects habitat quality in an irruptive species, the Black-backed Woodpecker. Condor Ornith. Appl. 2015, 117, 178–191. [Google Scholar] [CrossRef]
- Bagne, K.E.; Purcell, K.L.; Rotenberry, J.T. Prescribed fire, snag population dynamics, and avian nest site selection. For. Ecol. Manag. 2008, 255, 99–105. [Google Scholar] [CrossRef]
- Beedy, E.C. Bird communities and forest structure in the Sierra Nevada of California. Condor 1981, 83, 97–105. [Google Scholar] [CrossRef]
- Verner, J.; Larson, T.A. Richness of breeding bird species in mixed-conifer forests of the Sierra Nevada, California. Auk 1989, 106, 447–463. [Google Scholar]
- Willson, M.F.; Comet, T.A. Bird communities of northern forests: Patterns of diversity and abundance. Condor 1996, 98, 337–349. [Google Scholar] [CrossRef]
- Tingley, M.W.; Stillman, A.N.; Wilkerson, R.L.; Sawyer, S.C.; Siegel, R.B. Black-backed woodpecker occupancy in burned and beetle-killed forests: Disturbance agent matters. For. Ecol. Manag. 2020, 455, 117694. [Google Scholar] [CrossRef]
- George, T.L.; Zack, S. Bird occupancy and richness in ponderosa pine forests with contrasting forest structure and fire history. Can. J. For. Res. 2008, 38, 936–942. [Google Scholar] [CrossRef]
- Russell, R.E.; Royle, J.A.; Saab, V.A.; Lehmkuhl, J.F.; Block, W.M.; Sauer, J.R. Modeling the effects of environmental disturbance on wildlife communities: Avian responses to prescribed fire. Ecol. Appl. 2009, 19, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Bagne, K.E.; Purcell, K.L. Short-term responses of birds to prescribed fire in fire-suppressed forests of California. J. Wild. Manag. 2011, 75, 1051–1060. [Google Scholar] [CrossRef]
- Knapp, E.E.; Lydersen, J.M.; North, M.P.; Collins, B.M. Efficacy of variable density thinning and prescribed fire for restoring forest heterogeneity to mixed-conifer forest in the central Sierra Nevada, CA. For. Ecol. Manag. 2017, 406, 228–241. [Google Scholar] [CrossRef]
- Bock, C.E.; Lynch, J.F. Breeding bird populations of burned and unburned conifer forest in the Sierra Nevada. Condor 1970, 72, 182–189. [Google Scholar] [CrossRef]
- Raphael, M.G.; Morrison, M.L.; Yoder-Williams, M.P. Breeding bird populations during twenty-five years of postfire succession in the Sierra Nevada. Condor 1987, 89, 614–626. [Google Scholar] [CrossRef]
- White, A.M.; Manley, P.N.; Tarbill, G.L.; Richardson, T.W.; Russell, R.E.; Safford, H.D.; Dobrowski, S.Z. Avian community responses to post-fire forest structure: Implications for fire management in mixed conifer forests. Anim. Cons. 2016, 19, 256–264. [Google Scholar] [CrossRef]
- Kelly, L.T.; Bennett, A.F.; Clarke, M.F.; McCarthy, M.A. Optimal fire histories for biodiversity conservation. Cons. Biol. 2015, 29, 473–481. [Google Scholar] [CrossRef]
- Safford, H.D.; Stevens, J.T. Natural Range of Variation for Yellow Pine and Mixed-Conifer Forests in the Sierra Nevada, Southern Cascades, and Modoc and Inyo National Forests, California, USA; General Technical Report PSW-GTR-256; US Department of Agriculture, Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2017.
- Stephens, S.L.; McIver, J.D.; Boerner, R.E.J.; Fettig, C.J.; Fontaine, J.B.; Hartsough, B.R.; Kennedy, P.L.; Schwilk, D.W. The effects of forest fuel-reduction treatments in the United States. Bioscience 2012, 62, 549–560. [Google Scholar] [CrossRef]
- Franklin, J.F.; Fites-Kaufmann, J.A. Assessment of late-successional forests of the Sierra Nevada. In Sierra Nevada Ecosystem Project: Final Report to Congress, Vol. II, Assessments and Scientific Basis for Management Options; Centers for Water and Wildland Resources, University of California: Davis, CA, USA, 1996; Volume 37, pp. 627–656. [Google Scholar]
- Thompson, F.R., III; DeGraaf, R.M. Conservation approaches for woody, early succesional communities in the eastern United States. Wildl. Soc. Bull. 2001, 29, 483–494. [Google Scholar]
- Siegel, R.B.; DeSante, D.F. Bird communities in thinned versus unthinned sierran mixed conifer stands. Wilson Bull. 2003, 115, 155–165. [Google Scholar] [CrossRef]
- North, M.P.; Collins, B.M.; Stephens, S.L. Using fire to increase the scale, benefits, and future maintenance of fuels treatments. J. For. 2012, 110, 392–401. [Google Scholar] [CrossRef]
- Westphal, M.I.; Field, S.A.; Possingham, H.P. Optimizing landscape configuration: A case study of woodland birds in the Mount Lofty Ranges, South Australia. Landsc. Urban Plan. 2007, 81, 56–66. [Google Scholar] [CrossRef]
- Stephens, S.L.; Battaglia, M.A.; Churchill, D.J.; Collins, B.M.; Coppoletta, M.; Hoffman, C.M.; Lydersen, J.M.; North, M.P.; Parsons, R.A.; Ritter, S.M.; et al. Forest Restoration and Fuels Reduction: Convergent or Divergent? BioScience 2021, 71, 85–101. [Google Scholar]
- Zollner, P.A.; Roberts, L.J.; Gustafson, E.J.; He, H.S.; Radeloff, V. Influence of forest planning alternatives on landscape pattern and ecosystem processes in northern Wisconsin, USA. For. Ecol. Manag. 2008, 254, 429–444. [Google Scholar] [CrossRef]
- Lucash, M.S.; Scheller, R.M.; Gustafson, E.J.; Sturtevant, B.R. Spatial resilience of forested landscapes under climate change and management. Landsc. Ecol. 2017, 32, 953–969. [Google Scholar] [CrossRef]
- Steel, Z.L.; Koontz, M.J.; Safford, H.D. The changing landscape of wildfire: Burn pattern trends and implications for California’s yellow pine and mixed conifer forests. Landsc. Ecol. 2018, 33, 1159–1176. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roberts, L.J.; Burnett, R.; Fogg, A. Fire and Mechanical Forest Management Treatments Support Different Portions of the Bird Community in Fire-Suppressed Forests. Forests 2021, 12, 150. https://doi.org/10.3390/f12020150
Roberts LJ, Burnett R, Fogg A. Fire and Mechanical Forest Management Treatments Support Different Portions of the Bird Community in Fire-Suppressed Forests. Forests. 2021; 12(2):150. https://doi.org/10.3390/f12020150
Chicago/Turabian StyleRoberts, Lance Jay, Ryan Burnett, and Alissa Fogg. 2021. "Fire and Mechanical Forest Management Treatments Support Different Portions of the Bird Community in Fire-Suppressed Forests" Forests 12, no. 2: 150. https://doi.org/10.3390/f12020150
APA StyleRoberts, L. J., Burnett, R., & Fogg, A. (2021). Fire and Mechanical Forest Management Treatments Support Different Portions of the Bird Community in Fire-Suppressed Forests. Forests, 12(2), 150. https://doi.org/10.3390/f12020150