Influence of Forest Stand Structure and Competing Understory Vegetation on Ash Regeneration—Potential Effects of Ash Dieback


 ,
, 
Abstract
1. Introduction
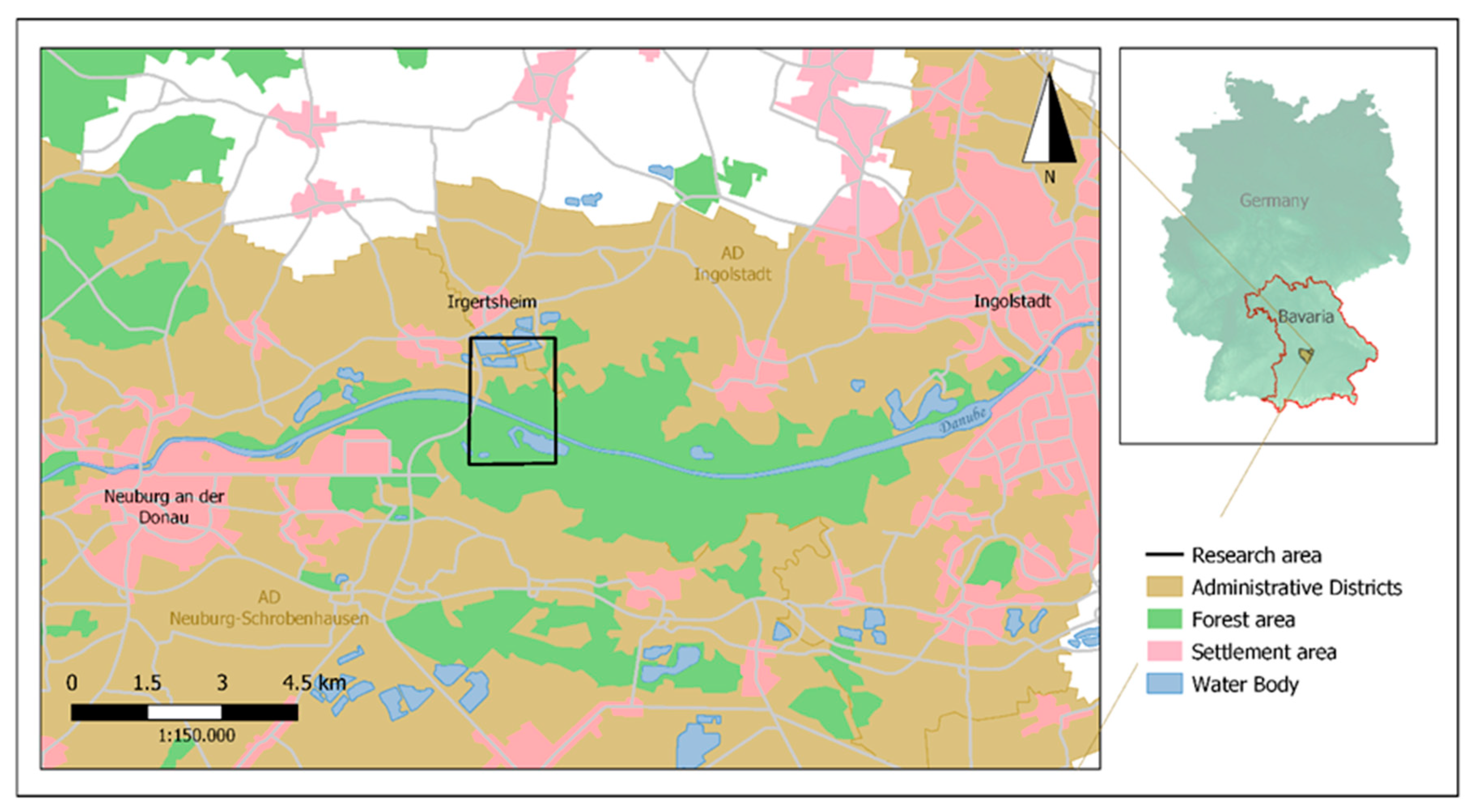
2. Study Area
3. Materials and Methods
4. Results
4.1. Ash Metrics and Abiotic and Biotic Variables
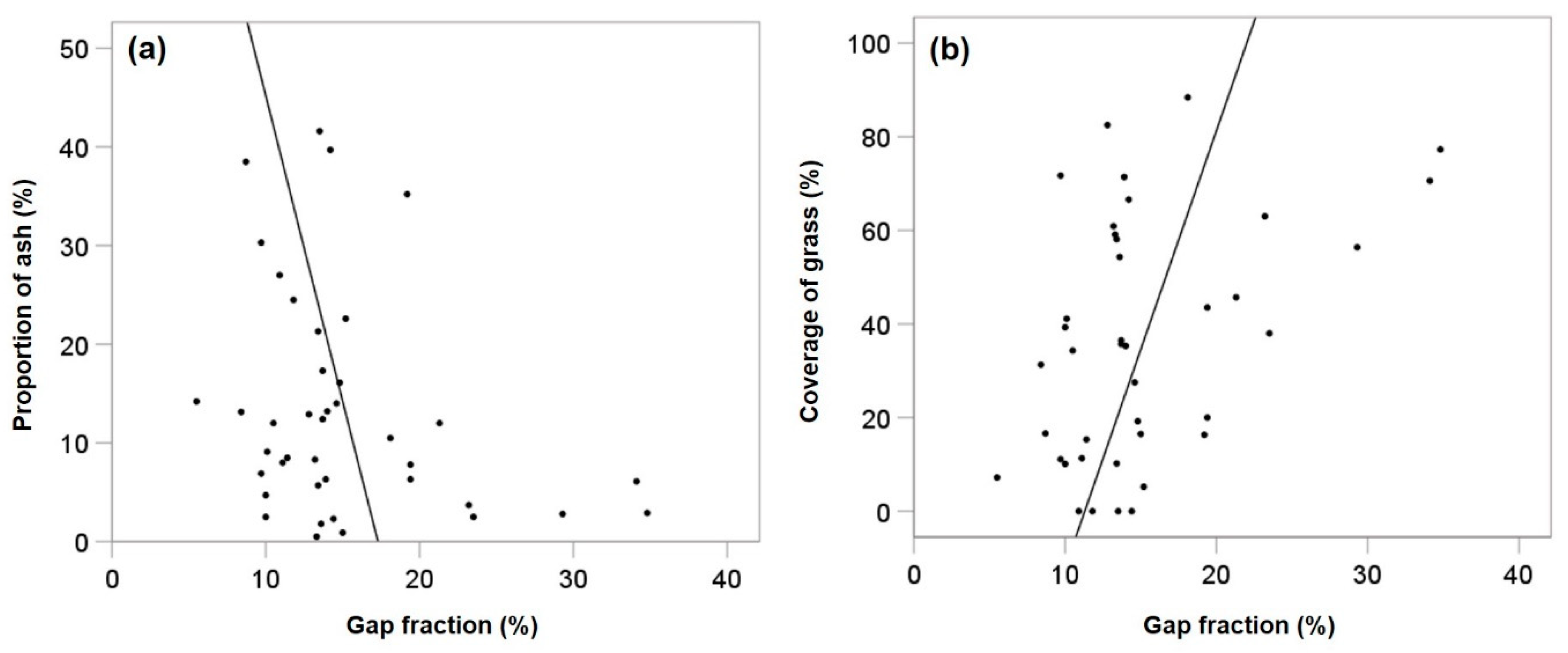
4.2. Relationship between Ash Regeneration, Grass Cover and Abiotic and Biotic Variables
5. Discussion
5.1. Relationships between Variables Related to Light Conditions and Ash Regeneration
5.2. Relationships between Other Abiotic Factors and Ash Regeneration
5.3. Seed Availability Inferred by Distance Measures
5.4. Competition in the Herb Layer
5.5. Relationship between Litter and Ash Regeneration
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bayerische Landesanstalt für Wald und Forstwirtschaft. Die Regionale Natürliche Waldzusammensetzung Bayerns; LWF-Wissen 32; Bayerische Landesanstalt für Wald und Forstwirtschaft: Freising, Germany, 2001. [Google Scholar]
- Mitchell, R.J.; Beaton, J.K.; Bellamy, P.E.; Broome, A.; Chetcuti, J.; Eaton, S.; Ellis, C.J.; Gimona, A.; Harmer, R.; Hester, A.J.; et al. Ash dieback in the UK: A review of the ecological and conservation implications and potential management options. Biol. Conserv. 2014, 175, 95–109. [Google Scholar] [CrossRef]
- Dichtl, T.; Stöger, W. Auenwald im Klimawandel: Forstliche Forschung an Donau und Rhein. LWF Aktuell 2020, 126, 28–29. [Google Scholar]
- Enderle, R.; Fussi, B.; Lenz, H.D.; Langer, G.; Nagel, R.; Metzler, B. Ash dieback in Germany: Research on disease development, resistance and management options. In Dieback of European Ash (Fraxinus spp.)—Consequences and Guidelines for Sustainable Management; Vasaitis, R., Enderle, R., Eds.; European Cooperation in Science & Technology (COST): Uppsala, Sewden, 2017; pp. 89–105. ISBN 978-91-576-8696-1. [Google Scholar]
- Tabari, K.M.; Lust, N.; Neirynk, J. Effect of light and humus on survival and height growth of ash (Fraxinus excelsior L.) seedlings. Silva Gandav. 1998, 63, 36–49. [Google Scholar] [CrossRef]
- Nüsslein, S. Waldbauliche Behandlung der Esche. In Beiträge zur Esche Fachtagung zum Baum des Jahres 2001; LWF-Wissen 34; Bayerische Landesanstalt für Wald und Forstwirtschaft: Freising, Germany, 2002; pp. 41–43. [Google Scholar]
- Gulder, H.-J. Standortansprüche und Wurzelwerk der Esche. In Beiträge zur Esche Fachtagung zum Baum des Jahres 2001; LWF-Wissen 34; Bayerische Landesanstalt für Wald und Forstwirtschaft: Freising, Germany, 2002; pp. 50–52. [Google Scholar]
- Schütz, J.P. Modellierung des Höhenwuchses der Esche in der Verjüngungsphase in Abhängigkeit von der Beschattung. In Proceedings of the Jahrestagung 2004 des Deutschen Verbandes forstlicher Forschungsanstalten, Stift Schlägl, Austria, 24–26 May 2004; Deutscher Verband Forstlicher Forschungsanstalten: Göttingen, Germany, 2004; p. 109. [Google Scholar]
- Enderle, R.; Nakou, A.; Thomas, K.; Metzler, B. Susceptibility of autochthonous German Fraxinus excelsior clones to Hymenoscyphus pseudoalbidus is genetically determined. Ann. For. Sci. 2015, 72, 183–193. [Google Scholar] [CrossRef]
- Zhao, Y.-J.; Hosoya, T.; Baral, H.-O.; Hosaka, K.; Kakishima, M. Hymenoscyphus pseudoalbidus, the correct name for Lambertella albida reported from Japan. Mycotaxon 2013, 122, 25–41. [Google Scholar] [CrossRef]
- Gross, A.; Holdenrieder, O.; Pautasso, M.; Queloz, V.; Sieber, T.N. Hymenoscyphus pseudoalbidus, the causal agent of European ash dieback. Mol. Plant Pathol. 2014, 15, 5–21. [Google Scholar] [CrossRef]
- Przybyl, K. Fungi associated with necrotic apical parts of Fraxinus excelsior shoots. For. Pathol. 2002, 32, 387–394. [Google Scholar] [CrossRef]
- Kowalski, T. Chalara fraxinea sp. nov. associated with dieback of ash (Fraxinus excelsior ) in Poland. For. Pathol. 2006, 36, 264–270. [Google Scholar] [CrossRef]
- Pautasso, M.; Aas, G.; Queloz, V.; Holdenrieder, O. European ash (Fraxinus excelsior) dieback—A conservation biology challenge. Biol. Conserv. 2013, 158, 37–49. [Google Scholar] [CrossRef]
- Giongo, S.; Oliveira Longa, C.M.; Dal Maso, E.; Montecchio, L.; Maresi, G. Evaluating the impact of Hymenoscyphus fraxineus in Trentino (Alps, Northern Italy): First investigations. iForest 2017, 10, 871–878. [Google Scholar] [CrossRef]
- Kowalski, T.; Holdenrieder, O. Pathogenicity of Chalara fraxinea. For. Pathol. 2009, 39, 1–7. [Google Scholar] [CrossRef]
- Skovsgaard, J.P.; Thomsen, I.M.; Skovgaard, I.M.; Martinussen, T. Associations among symptoms of dieback in even-aged stands of ash (Fraxinus excelsior L.). For. Pathol. 2010, 40, 7–18. [Google Scholar] [CrossRef]
- McKinney, L.V.; Nielsen, L.R.; Hansen, J.K.; Kjær, E.D. Presence of natural genetic resistance in Fraxinus excelsior (Oleraceae) to Chalara fraxinea (Ascomycota): An emerging infectious disease. Heredity 2011, 106, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Rysavy, T.; Roloff, A. Ursachen der Vereschung in Mischbeständen und Vorschläge zu ihrer Vermeidung. Forst Und Holz 1994, 49, 392–395. [Google Scholar]
- Metzler, B.; Enderle, R.; Karopka, M.; Toepfner, K.; Aldinger, E. Development of ash dieback in a provenance trial on different sites in southern Germany. In Allgmeine Forst und Jagdzeitung; JD Sauerlaender’s Verlag: Bad Orb, Germany, 2012; Volume 183, pp. 168–180. [Google Scholar]
- McKinney, L.V.; Nielsen, L.R.; Collinge, D.B.; Thomsen, I.M.; Hansen, J.K.; Kjaer, E.D. The ash dieback crisis: Genetic variation in resistance can prove a long-term solution. Plant Pathol. 2014, 63, 485–499. [Google Scholar] [CrossRef]
- Pliūra, A.; Lygis, V.; Suchockas, V. Performance of twenty four European Fraxinus excelsior populations in three Lithuanian progeny trials with a special emphasis on resistance to Chalara fraxinea. Balt. For. 2011, 17, 17–34. [Google Scholar]
- Tabari, K.M.; Lust, N. Monitoring of natural regeneration in a mixed deciduous forest. Silva Gandav. 1999, 64. [Google Scholar] [CrossRef]
- Dobrowolska, D.; Hein, S.; Oosterbaan, A.; Wagner, S.; Clark, J.; Skovsgaard, J.P. A review of European ash (Fraxinus excelsior L.): Implications for silviculture. Forestry 2011, 84, 133–148. [Google Scholar] [CrossRef]
- Petritan, A.M.; von Lupke, B.; Petritan, I.C. Effects of shade on growth and mortality of maple (Acer pseudoplatanus), ash (Fraxinus excelsior) and beech (Fagus sylvatica) saplings. Forestry 2007, 80, 397–412. [Google Scholar] [CrossRef]
- Coomes, D.A.; Grubb, P.J. Impacts of root competition in forests and woodlands: A theoretical framework and review of experiments. Ecol. Monogr. 2000, 70, 171–207. [Google Scholar] [CrossRef]
- Flory, S.L.; Clay, K. Non-native grass invasion suppresses forest succession. Oecologia 2010, 164, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Gilliam, F.S. The Ecological Significance of the Herbaceous Layer in Temperate Forest Ecosystems. BioScience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- ForstBW. Jahresbericht 2012 des Landesbetriebes ForstBW; ForstBW: Baden-Württemberg, Germany, 2012; p. 58. [Google Scholar]
- Langer, G.; Bressem, U. Eschentriebsterben—Praxisinformation Nr. 4; Bayerische Landesanstalt für Wald und Forstwirtschaft: Göttingen, Germany, 2016; Available online: https://www.lwf.bayern.de/mam/cms04/waldschutz/dateien/bonitur_von_alteschen.pdf (accessed on 4 December 2020).
- Lygis, V.; Vasiliauskas, R.; Larsson, K.-H.; Stenlid, J. Wood-inhabiting fungi in stems of Fraxinus excelsior in declining ash stands of northern Lithuania, with particular reference to Armillaria cepistipes. Scand. J. For. Res. 2005, 20, 337–346. [Google Scholar] [CrossRef]
- Geiger, R. Überarbeitete Neuausgabe von Geiger, P. Köppen-Geiger / Klima der Erde; Klett-Perthes: Gotha, Germany, 1961. [Google Scholar]
- Bayerische Landesanstalt für Landwirtschaft. Agrarmeteorologie by: Weather Data. (Open Document). Available online: https://dynamax.com/images/uploads/papers/HemiView_Manual.pdf (accessed on 4 December 2020).
- Schwab, A.; Stammel, B.; Kiehl, K. Seed dispersal via a new watercourse in a reconnected floodplain: Differences in species groups and seasonality. Restor. Ecol. 2018, 26, S103–S113. [Google Scholar] [CrossRef]
- Margraf, C. Die Vegetationsentwicklung der Donauauen Zwischen Ingolstadt und Neuburg: Vegetationskundlich-ökologische Studie über den Wandel Einer Auenlandschaft 30 Jahre Nach Staustufenbau; Hoppea, Denkschriften der Regensburgischen Botanischen Gesellschaft: Regensburg, Germany, 2004. [Google Scholar]
- Lang, P.; Ewald, J. Predictive modelling and monitoring of Ellenberg moisture value validates restoration success in floodplain forests. Appl. Veg. Sci. 2014, 17, 543–555. [Google Scholar] [CrossRef]
- Doben, K.; Doppler, G.; Freudenberger, W.; Jerz, H.; Meyer, R.K.F.; Mielke, H.; Ott, W.-D.; Rohrmüller, J.; Schmidt-Kaler, H.; Schwerd, K.; et al. Geologische Karte von Bayern; 4. Auflage; Bayerisches Geologisches Landesamt: Munich, Germany, 1996. [Google Scholar]
- Lenz, H.; Straßner, L.; Baumann, M.; Baier, U. Boniturschlüssel zur Einstufung der Vitalität von Alteschen. Afz-Der Wald 2012, 3, 18–129. [Google Scholar]
- Londo, G. The decimal scale for releves of permanent quadrats. Vegetatio 1976, 33, 61–64. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulißen, D. (Eds.) Zeigerwerte von Pflanzen in Mitteleuropa; 2. Auflage; Verlag Erich Goltze GmbH & Co. KG: Göttingen, Germany, 1992. [Google Scholar]
- Monteith, J.L. Principles of Environmental Physics; Reprinted with corr.; Arnold: London, UK, 1973; ISBN 0713123753. [Google Scholar]
- Delta-T Devices. User Manual for HemiView, Version 2.1. 1999. Available online: https://dynamax.com/images/uploads/papers/HemiView_Manual.pdf (accessed on 4 December 2020).
- Welles, J.M.; Norman, J.M. Instrument for Indirect Measurement of Canopy Architecture. Agron. J. 1991, 83, 818–825. [Google Scholar] [CrossRef]
- Marigo, G.; Peltier, J.-P.; Girel, J.; Pautou, G. Success in the demographic expansion of Fraxinus excelsior L. Trees 2000, 15, 1–13. [Google Scholar] [CrossRef]
- Welles, J.M.; Cohen, S. Canopy structure measurement by gap fraction analysis using commercial instrumentation. J. Exp. Bot. 1996, 47, 1335–1342. [Google Scholar] [CrossRef]
- Mölder, A.; Bernhardt-Römermann, M.; Schmidt, W. Herb-layer diversity in deciduous forests: Raised by tree richness or beaten by beech? For. Ecol. Manag. 2008, 256, 272–281. [Google Scholar] [CrossRef]
- Thomas, P.A. Biological Flora of the British Isles: Fraxinus excelsior. J. Ecol. 2016, 104, 1158–1209. [Google Scholar] [CrossRef]
- Kerr, G.; Cahalan, C. A review of site factors affecting the early growth of ash (Fraxinus excelsior L.). For. Ecol. Manag. 2004, 188, 225–234. [Google Scholar] [CrossRef]
- Wagner, S. Ein Modell zur Fruchtausbreitung der Esche (Fraxinus excelsior L.) unter Berücksichtigung von Richtungseffekten. Allg. Forst Jagdztg. 1997, 168, 149–155. [Google Scholar]
- BMEL. The Forests in Germany—Selected Results of the Third National Forest Inventory; BMEL: Bonn, Germany, 2014. [Google Scholar]
- Gilliam, F.S. Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition. J. Ecol. 2006, 94, 1176–1191. [Google Scholar] [CrossRef]
- Plue, J.; van Gils, B.; de Schrijver, A.; Peppler-Lisbach, C.; Verheyen, K.; Hermy, M. Forest herb layer response to long-term light deficit along a forest developmental series. Acta Oecol. 2013, 53, 63–72. [Google Scholar] [CrossRef]
- Lyon, J.; Sharpe, W.E. Impacts of Hay-Scented Fern on Nutrition of Northern Red Oak Seedlings. J. Plant Nutr. 2003, 26, 487–502. [Google Scholar] [CrossRef]
- Oijen, D.; Feijen, M.; Hommel, P.; Ouden, J.; Waal, R. Effects of tree species composition on within-forest distribution of understorey species. Appl. Veg. Sci. 2005, 8, 155–166. [Google Scholar] [CrossRef]
- Langenbruch, C.; Helfrich, M.; Flessa, H. Effects of beech (Fagus sylvatica), ash (Fraxinus excelsior) and lime (Tilia spec.) on soil chemical properties in a mixed deciduous forest. Plant Soil 2012, 352, 389–403. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variable Category | Statisitics | Ash Seedling Density [m−2] | Proportion of Ash | Ash % among Seedlings | |||
|---|---|---|---|---|---|---|---|
| Ash regeneration metrics | Mean | 2.2 | 13.2 | 38.3 | |||
| Standard deviation | 2.0 | 11.3 | 23.4 | ||||
| Minimum | 0.1 | 0.5 | 0 | ||||
| Maximum | 8.5 | 41.6 | 100 | ||||
| LAI | G | M | R | N | |||
| Variables related to light conditions and abiotic factors | Mean | 2.1 | 15.2 | 5.4 | 7.1 | 6.7 | |
| Standard deviation | 0.5 | 6.4 | 0.6 | 0.2 | 0.8 | ||
| Minimum | 1.0 | 5.5 | 3.7 | 6.5 | 4.4 | ||
| Maximum | 3.4 | 34.8 | 6.0 | 7.7 | 8.0 | ||
| Dist. Ash | N Species | Herb Layer % | Litter % | Soil % | Grass % | ||
| Variables related to vegetation and biotic factors | Mean | 13.5 | 10.3 | 73.1 | 16.7 | 3.4 | 36.2 |
| Standard deviation | 17.6 | 2.5 | 20.8 | 19.8 | 6.5 | 26.0 | |
| Minimum | 0.8 | 4.0 | 18.0 | 0.0 | 0.0 | 0.0 | |
| Maximum | 50.0 | 17.0 | 98.0 | 75.0 | 40.0 | 88.4 |
| Species | Coverage % | Species | Coverage % |
|---|---|---|---|
| Aegopodium podagraria (h) | 11.9 | Juniperus communis (s) | 0.2 |
| Acer pseudoplatanus (t) | 6.5 | Vincetoxicum hirundinaria (h) | 0.1 |
| Fraxinus excelsior (t) | 6.4 | Aconitum napellus (h) | 0.1 |
| Acer campestre (t) | 2.1 | Anthriscus sylvestris (h) | 0.1 |
| Rubus silvaticus (s) | 1.3 | Corylus avellana (s) | 0.1 |
| Carpinus betulus (t) | 1.1 | Galeopsis tetrahit (h) | 0.1 |
| Viola odorata (h) | 1.1 | Galium mollugo (h) | 0.1 |
| Cornus mas (s) | 1.0 | Glechoma hederacea (h) | 0.1 |
| Geum urbanum (h) | 0.9 | Hypericum perforatum (h) | 0.1 |
| Asarum europaeum (h) | 0.8 | Viburnum lantana (s) | 0.1 |
| Viola reichenbachiana (h) | 0.7 | Centaurea jacea (h) | 0.1 |
| Lonicera xylosteum (s) | 0.6 | Hieracium murorum (h) | 0.1 |
| Crataegus monogyna (s) | 0.6 | Hypericum hirsutum (h) | 0.1 |
| Galium aparine (h) | 0.5 | Lotus corniculatus (h) | 0.1 |
| Clematis vitalba (l) | 0.4 | Pimpinella saxifrage (h) | 0.1 |
| Cornus sanguinea (s) | 0.4 | Prunus padus (t) | 0.1 |
| Euphorbia cyparissias (h) | 0.4 | Rosa canina (s) | 0.1 |
| Quercus robur (t) | 0.4 | Veronica arvensis (h) | <0.1 |
| Ligustrum vulgare (s) | 0.4 | Clinopodium vulgare (h) | <0.1 |
| Paris quadrifolia (h) | 0.4 | Euphorbia amygdaloides (h) | <0.1 |
| Ranunculus nemorosus (h) | 0.3 | Lamium album (h) | <0.1 |
| Lamium maculatum (h) | 0.3 | Plantago lanceolata(h) | <0.1 |
| Mercurialis perennis (h) | 0.3 | Primula elatior (h) | <0.1 |
| Tilia cordata (t) | 0.3 | Prunella vulgaris (h) | <0.1 |
| Ulmus minor (t) | 0.2 | Salvia pratensis (h) | <0.1 |
| Circaea lutetiana (h) | 0.2 | Tilia platyphyllos (t) | <0.1 |
| Daphne mezereum (s) | 0.2 | Viburnum lantana (h) | <0.1 |
| Origanum vulgare (h) | 0.2 | Viola melissifolia (h) | <0.1 |
| Viola riviniana (h) | <0.1 |
| Variables | LAI | G | M | R | N | ||
|---|---|---|---|---|---|---|---|
| ash seedling density | rs | 0.400 | −0.385 | 0.036 | −0.330 | 0.101 | |
| p | 0.011 | 0.014 | 0.842 | 0.038 | 0.533 | ||
| proportion of ash | rs | 0.330 | −0.272 | −0.190 | −0.355 | −0.197 | |
| p | 0.037 | 0.089 | 0.241 | 0.025 | 0.222 | ||
| ash % among seedlings | rs | 0.245 | −0.172 | −0.200 | −0.204 | 0.014 | |
| p | 0.128 | 0.288 | 0.215 | 0.207 | 0.931 | ||
| grass % | rs | −0.346 | 0.319 | −0.354 | 0.157 | −0.519 | |
| p | 0.029 | 0.045 | 0.025 | 0.333 | 0.001 | ||
| herb % | rs | −0.164 | 0.203 | −0.023 | 0.277 | 0.012 | |
| p | 0.311 | 0.210 | 0.887 | 0.084 | 0.939 | ||
| dist. ash | N species | herb % | grass % | litter % | soil % | ||
| ash seedling density | rs | 0.196 | −0.042 | −0.523 | −0.546 | 0.619 | 0.112 |
| p | 0.273 | 0.795 | 0.001 | 0.000 | 0.000 | 0.496 | |
| proportion of ash | rs | 0.110 | −0.088 | −0.517 | −0.323 | 0.674 | 0.159 |
| p | 0.544 | 0.591 | 0.001 | 0.042 | 0.000 | 0.335 | |
| ash % among seedlings | rs | 0.092 | −0.005 | −0.318 | −0.360 | 0.444 | −0.046 |
| p | 0.609 | 0.977 | 0.046 | 0.023 | 0.005 | 0.782 | |
| grass % | rs | −0.228 | 0.059 | 0.571 | 1 | 0.457 | −0.347 |
| p | 0.202 | 0.719 | 0.000 | 0.003 | 0.030 | ||
| herb % | rs | −0.134 | 0.214 | 1 | 0.571 | 0.809 | −0.335 |
| p | 0.457 | 0.186 | 0.000 | 0.000 | 0.037 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jochner-Oette, S.; Rohrer, T.; Eisen, A.-K.; Tönnes, S.; Stammel, B. Influence of Forest Stand Structure and Competing Understory Vegetation on Ash Regeneration—Potential Effects of Ash Dieback. Forests 2021, 12, 128. https://doi.org/10.3390/f12020128
Jochner-Oette S, Rohrer T, Eisen A-K, Tönnes S, Stammel B. Influence of Forest Stand Structure and Competing Understory Vegetation on Ash Regeneration—Potential Effects of Ash Dieback. Forests. 2021; 12(2):128. https://doi.org/10.3390/f12020128
Chicago/Turabian StyleJochner-Oette, Susanne, Tanja Rohrer, Anna-Katharina Eisen, Selina Tönnes, and Barbara Stammel. 2021. "Influence of Forest Stand Structure and Competing Understory Vegetation on Ash Regeneration—Potential Effects of Ash Dieback" Forests 12, no. 2: 128. https://doi.org/10.3390/f12020128
APA StyleJochner-Oette, S., Rohrer, T., Eisen, A.-K., Tönnes, S., & Stammel, B. (2021). Influence of Forest Stand Structure and Competing Understory Vegetation on Ash Regeneration—Potential Effects of Ash Dieback. Forests, 12(2), 128. https://doi.org/10.3390/f12020128