
Winter Roosting by Eastern Red Bats in Ozark Mountain Forests of Missouri


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Capture and Radio-Tracking
2.3. Roost-Site Characterization
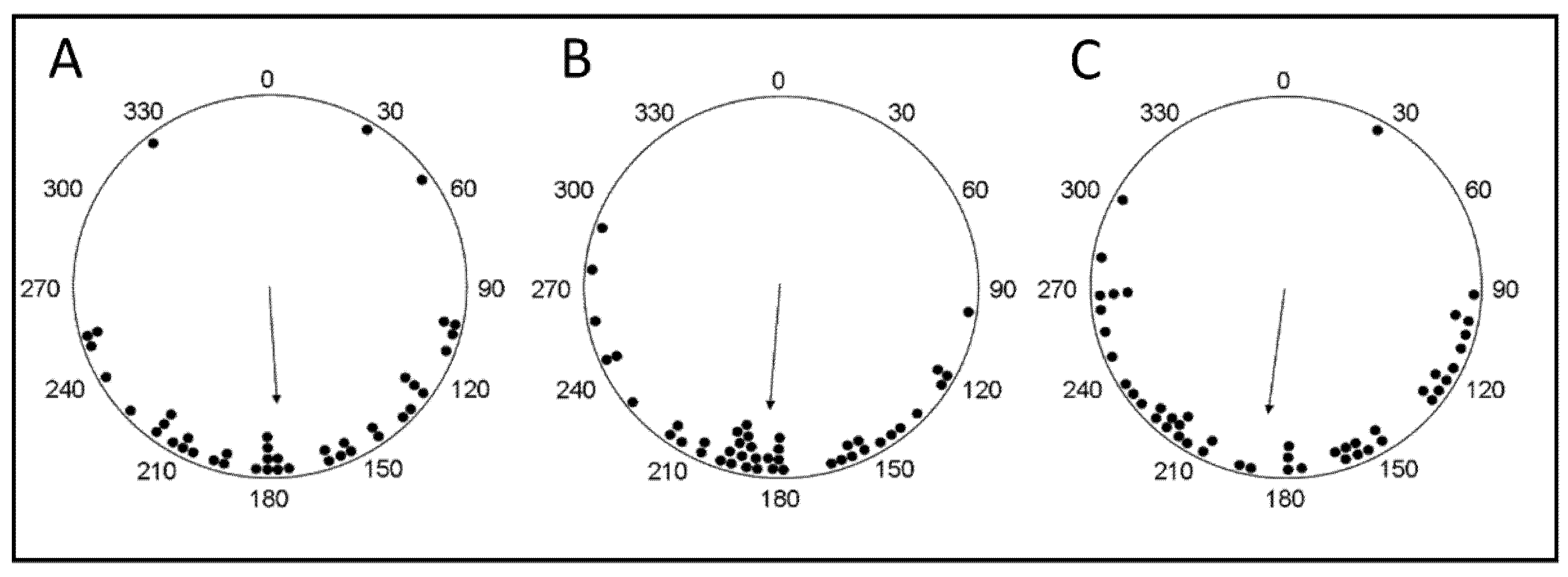
2.4. Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shump, K.A., Jr.; Shump, A.U. Lasiurus borealis. Mamm. Species 1982, 183, 1–6. [Google Scholar] [CrossRef]
- Cryan, P.M. Seasonal distribution of migratory tree bats (Lasiurus and Lasionycteris) in North America. J. Mammal. 2003, 84, 579–593. [Google Scholar] [CrossRef] [Green Version]
- Boyles, J.G.; Timpone, J.C.; Robbins, L.W. Late-winter observations of red bats, Lasiurus borealis, and evening bats, Nycticeius humeralis, in Missouri. Bat Res. News 2003, 44, 59–61. [Google Scholar]
- Mormann, B.M.; Robbins, L.W. Winter roosting ecology of eastern red bats in southwest Missouri. J. Wildl. Manag. 2007, 71, 213–217. [Google Scholar] [CrossRef]
- Davis, W.H.; Lidicker, W.Z., Jr. Winter range of the red bat, Lasiurus borealis. J. Mammal. 1956, 37, 280–291. [Google Scholar] [CrossRef]
- Koontz, T.; Davis, W. Winter roosting of the red bat, Lasiurus borealis. Bat Res. News 1991, 32, 3–4. [Google Scholar]
- Rodrigue, J.L.; Schuler, T.M.; Menzel, M.A. Observations of bat activity during prescribed burning in West Virginia. Bat Res. News 2001, 42, 48–49. [Google Scholar]
- Whitaker, J.O., Jr.; Rose, R.K.; Padgett, T.M. Food of the red bat Lasiurus borealis in winter in the Great Dismal Swamp, North Carolina and Virginia. Am. Midl. Nat. 1997, 137, 408–412. [Google Scholar]
- Winhold, L.; Kurta, A.; Foster, R. Long-term change in an assemblage of North American bats: Are eastern red bats declining? Acta Chiropterol. 2008, 10, 359–366. [Google Scholar] [CrossRef]
- Kunz, T.H.; Arnett, E.B.; Erickson, W.P.; Hoar, A.R.; Johnson, G.D.; Larkin, R.P.; Strickland, M.D.; Thresher, R.W.; Tuttle, M.D. Ecological impacts of wind energy development on bats: Questions, research needs, and hypotheses. Front. Ecol. Environ. 2007, 5, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Arnett, E.B.; Brown, W.K.; Erickson, W.P.; Fiedler, J.K.; Hamilton, B.L.; Henry, T.H.; Jain, A.; Johnson, G.D.; Kerns, J.; Koford, R.R.; et al. Patterns of bat fatalities at wind energy facilities in North America. J. Wildl. Manag. 2008, 72, 61–78. [Google Scholar] [CrossRef]
- Hein, C.D.; Castleberry, S.B.; Miller, K.V. Male Seminole bat winter roost-site selection in a managed forest. J. Wildl. Manag. 2008, 72, 1756–1764. [Google Scholar] [CrossRef]
- Perry, R.W. A review of fire effects on bats and bat habitat in the eastern oaks region. In Proceedings of the 4th Fire in Eastern Oak Forests Conference, Springfield, MO, USA, 17–19 May 2012; USDA Forest Service General Technical Report NRS-P-102. pp. 170–191. [Google Scholar]
- Saugey, D.A.; Heath, D.R.; Heidt, G.A. The bats of the Ouachita Mountains. Proc. Arkansas Acad. Sci. 1989, 43, 71–77. [Google Scholar]
- Moorman, C.E.; Russell, K.R.; Menzel, M.A.; Lohr, S.M.; Ellenberger, J.E.; Van Lear, D.H. Bats roosting in deciduous leaf litter. Bat Res. News 1999, 40, 74–75. [Google Scholar]
- Layne, J.T.; Green, D.; Scesny, A.; Robbins, L.W. Eastern Red Bat Responses to Fire during Winter Torpor. Forests 2021, 12, 1347. [Google Scholar] [CrossRef]
- Van Lear, D.H.; Harlow, R.F. Fire in the eastern United States: Influence on wildlife habitat. In The Role of Fire in Nongame Wildlife Management and Community Restoration: Traditional Uses and New Directions; USDA Forest Service General Technical Report NE-288; USDA: Newton Square, PA, USA, 2002; pp. 2–10. [Google Scholar]
- Carter, T.C.; Ford, W.M.; Menzel, M.A. Fire and bats in the southeast and mid-Atlantic: More questions than answers. In The Role of Fire in Nongame Wildlife Management and Community Restoration: Traditional Uses and New Directions; USDA Forest Service General Technical Report NE-288; USDA: Newton Square, PA, USA, 2002; pp. 139–143. [Google Scholar]
- Keyser, P.D.; Ford, W.M. Influence of fire on mammals in eastern oak forests. In Fire in Eastern Oak Forests: Delivering Science to Land Managers, Proceedings of a Conference; USDA Forest Service General Technical Report NRS-P-1; USDA: Newton Square, PA, USA, 2006; pp. 180–190. [Google Scholar]
- Dickinson, M.B.; Lacki, M.J.; Cox, D.R. Fire and the endangered Indiana bat. In Proceedings of the 3rd Fire in Eastern Oak Forests Conference, Carbondale, IL, USA, 20–22 May 2008; USDA Forest Service General Technical Report NRS-P-46. USDA: Newton Square, PA, USA, 2009; pp. 51–75. [Google Scholar]
- Loeb, S.C. Qualitative synthesis of temperate bat responses to silvicultural treatments—where do we go from here? J. Mammal. 2020, 101, 1513–1525. [Google Scholar] [CrossRef]
- Jorge, M.H.; Sweeten, S.E.; True, M.C.; Freeze, S.R.; Cherry, M.J.; Garrison, E.P.; Taylor, H.; Gorman, M.K.; Ford, W.M. Fire, land cover, and temperature drivers of bat activity in winter. Fire Ecol. 2021, 17, 19. [Google Scholar] [CrossRef]
- Flinn, J.R. Winter Roosting Behavior of Red Bats (Lasiurus borealis): Habitat Use, Microclimate, and Effects of Ambient Temperature on Roost Choice. Master’s Thesis, Missouri State University, Springfield, MO, USA, 2009. [Google Scholar]
- Anthony, E.L.P. Age determination in bats. In Ecological and Behavioral Methods for the Study of Bats; Kunz, T.H., Ed.; Smithsonian Institution Press: Washington, DC, USA, 1988; pp. 47–58. [Google Scholar]
- Vonhof, M.J.; Barclay, R.M.R. Roost-site selection and roosting ecology of forest-dwelling bats in southern British Columbia. Can. J. Zool. 1996, 74, 1797–1805. [Google Scholar] [CrossRef]
- Beyer, H.L. Hawth’s Analysis Tools for ArcGIS. Available online: http://www.spatialecology.com/htools (accessed on 1 December 2008).
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice-Hall: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Perry, R.W.; Saugey, D.A.; Crump, B.G. Winter roosting ecology of silver-haired bats in an Arkansas Forest. Southeast. Nat. 2010, 9, 563–572. [Google Scholar] [CrossRef]
- Dunbar, M.B.; Tomasi, T.E. Arousal patterns, metabolic rate, and an energy budget of eastern red bats (Lasiurus borealis) in winter. J. Mammal. 2006, 87, 1096–1102. [Google Scholar] [CrossRef] [Green Version]
- Schaffalitzky De Muckadell, M. Environmental factors in development stages of trees. In Tree Growth; Kozlowski, T.T., Ed.; Ronald Press: New York, NY, USA, 1962; pp. 289–297. [Google Scholar]
- Kalcounis-Rüppell, M.C.; Psyllakis, J.M.; Brigham, R.M. Tree roost selection by bats: An empirical synthesis using meta-analysis. Wildl. Soc. Bull. 2005, 33, 175–190. [Google Scholar] [CrossRef]
- Elmore, L.W.; Miller, D.A.; Vilella, F.J. Selection of diurnal roosts by red bats (Lasiurus borealis) in an intensively managed pine forest in Mississippi. For. Ecol. Manag. 2004, 199, 11–20. [Google Scholar] [CrossRef]
- Mager, K.J.; Nelson, T.A. Roost-site selection by Eastern red bats (Lasiurus borealis). Am. Mildl. Nat. 2001, 145, 120–126. [Google Scholar] [CrossRef]
- Menzel, M.A.; Carter, T.C.; Ford, W.M.; Chapman, B.R.; Ozier, J. Summer roost tree selection by eastern red, Seminole, and evening bats in the upper Coastal Plain of South Carolina. Proc. Annu. Conf. Southeast. Assoc. Fish Wildl. Agencies 2000, 54, 304–313. [Google Scholar]
- Perry, R.W.; Thill, R.E.; Carter, S.A. Sex-specific roost selection by adult red bats in a diverse forested landscape. For. Ecol. Manag. 2007, 253, 48–55. [Google Scholar] [CrossRef]
- Perry, R.W. A review of factors affecting cave climates for hibernating bats in temperate North America. Environ. Rev. 2013, 21, 28–39. [Google Scholar] [CrossRef]
- Perry, R.W. Potential energy expenditure by litter-roosting bats associated with temperature profiles under leaf litter during winter. J. Therm. Biol. 2013, 38, 467–473. [Google Scholar] [CrossRef]
- Fenton, M.B. Population Studies of Myotis lucifugus (Chiroptera: Vespertilionidae) in Ontario. Life Sci. Contr. R. Ont. Mus. 1970, 77, 34. [Google Scholar]
- Haarsma, A.; Kaal, R. Predation of wood mice (Apodemus sylvaticus) on hibernating bats. Popul. Ecol. 2016, 58, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Avery, M.I. Winter Activity of Pipistrelle Bats. J. Anim. Ecol. 1985, 54, 721–738. [Google Scholar] [CrossRef]
- Heyerdahl, E.; Brubaker, L.; Agee, J. Spatial controls of historical fire regimes: A multiscale example from the interior west, USA. Ecology 2001, 82, 660–678. [Google Scholar] [CrossRef]
- Dillon, G.K.; Holden, Z.A.; Morgan, P.; Crimmins, M.A.; Heyerdahl, E.K.; Luce, C.H. Both topography and climate affected forest and woodland burn severity in two regions of the western US, 1984 to 2006. Ecosphere 2011, 2, 1–33. [Google Scholar] [CrossRef]
- Perry, R.W.; McDaniel, V.L. Temperatures below leaf litter during winter prescribed burns: Implications for litter-roosting bats. Int. J. Wildland Fire 2015, 24, 544–549. [Google Scholar] [CrossRef]

{kind=link}
| Tree Species Group | Available | Roosts | z | p |
|---|---|---|---|---|
| Oak (Quercus spp.) | 34 | 91 | 6.47 | <0.001 |
| Hickory (Carya spp.) | 17 | 2 | −3.11 | 0.002 |
| Shortleaf pine (Pinus echinate Mill.) | 10 | 7 | −1.01 | 0.311 |
| Other species | 19 | 0 | −2.12 | 0.016 |
| Elm (Ulmus spp.) * | 8 | 0 | ||
| Eastern redcedar (Juniperus virginiana L.) * | 2 | 0 | ||
| Flowering dogwood (Cornus florida L.) * | 8 | 0 | ||
| Eastern redbud (Cercis canadensis L.) * | 2 | 0 |
| Variables | AICc | ∆AICc |
|---|---|---|
| Tree roosts | ||
| Slope, Aspect, Elevation, Snags/ha | 57.153 | 0 |
| Slope, Aspect, Elevation, Snags/ha, RoadDis | 58.375 | 1.22 |
| Slope, Aspect, Elevation, Snags/ha, Decid/ha | 58.637 | 1.48 |
| Slope, Aspect, Elevation, Snags/ha, Decid/ha, Conifer/ha, RoadDist | 59.371 | 2.22 |
| Slope, Aspect, Elevation | 62.472 | 5.32 |
| Litter roosts | ||
| Slope, Aspect, Litter Depth, Litter Cover | 91.041 | 0 |
| Slope, Aspect, Litter Depth, Litter Cover, Stem Density | 93.231 | 2.19 |
| Slope, Aspect, Litter Depth, Litter Cover, Stem Density, Elevation | 94.418 | 3.38 |
| Aspect, Litter Cover | 94.976 | 3.94 |
| Aspect, Litter Depth, Litter Cover | 96.625 | 5.58 |
| Variable | Estimate | SE | LCL | UCL | Odds |
|---|---|---|---|---|---|
| Tree roosts | |||||
| Slope | * 0.154 | 0.077 | 0.004 | 0.305 | 1.167 |
| Southerly aspect | * 1.019 | 0.276 | 0.477 | 1.561 | 2.770 |
| Elevation | * 0.118 | 0.033 | 0.053 | 0.182 | 1.125 |
| Snags/ha | * −0.223 | 0.010 | −0.042 | −0.003 | 0.977 |
| Deciduous trees/ha | 0.002 | 0.002 | −0.001 | 0.006 | 1.002 |
| Distance to roads | −0.001 | 0.001 | −0.002 | 0.001 | 0.999 |
| Litter roosts | |||||
| Slope | * 0.140 | 0.054 | 0.035 | 0.245 | 1.156 |
| Southerly aspect | * 1.146 | 0.245 | 0.665 | 1.626 | 3.145 |
| Litter depth | −0.018 | 0.024 | −0.066 | 0.030 | 0.982 |
| Litter cover | * 0.089 | 0.034 | 0.023 | 0.155 | 1.093 |
| Land Cover Class | Available % | Used % | z | p |
|---|---|---|---|---|
| Upland deciduous forest | 65 | 80 | 3.32 | <0.001 |
| Mixed shortleaf pine-oak forest and woodland | 28 | 16 | −2.50 | 0.013 |
| Other LULC classes | 6 | 2 | −2.90 | 0.004 |
| Shortleaf pine forest and woodland * | 3 | 0 | ||
| Deciduous woodland * | 1 | 0 | ||
| Glade complex * | 1 | 2 | ||
| Cool-season grassland * | 1 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flinn, J.R.; Perry, R.W.; Robbins, L.W. Winter Roosting by Eastern Red Bats in Ozark Mountain Forests of Missouri. Forests 2021, 12, 1769. https://doi.org/10.3390/f12121769
Flinn JR, Perry RW, Robbins LW. Winter Roosting by Eastern Red Bats in Ozark Mountain Forests of Missouri. Forests. 2021; 12(12):1769. https://doi.org/10.3390/f12121769
Chicago/Turabian StyleFlinn, Joshua R., Roger W. Perry, and Lynn W. Robbins. 2021. "Winter Roosting by Eastern Red Bats in Ozark Mountain Forests of Missouri" Forests 12, no. 12: 1769. https://doi.org/10.3390/f12121769
APA StyleFlinn, J. R., Perry, R. W., & Robbins, L. W. (2021). Winter Roosting by Eastern Red Bats in Ozark Mountain Forests of Missouri. Forests, 12(12), 1769. https://doi.org/10.3390/f12121769