1. Introduction
Originating in the Caucasus and Asia Minor, where it was first domesticated and spread throughout southern Europe, sweet (or European) chestnut,
Castanea sativa Mill., is the only native species of the
Castanea genus in Europe [
1]. During the Middle Ages, sweet chestnut was considered the “mountain cereal”, and today it occupies three climatic sub-regions, growing from sea level up to 1800 m over a wide range of climatic conditions [
2,
3].
Historically, chestnut has been used as an important ingredient in many nutrient products due to its richness in various nutriments [
4]. Moreover, chestnut trees have been distributed widely throughout mainly southern Europe, forming high-density forests in France and Italy, indicating the importance of this tree economically and environmentally [
5]. Like many European forest species, chestnut has notable potential in reducing pollution and climate change effects [
6]. The potential multi-use value of chestnut in the past is still valid today and new uses have proven to be economically important.
From a sustainable arboricultural point of view, it is crucial to combine increased productivity and competitiveness with the maintenance of biodiversity. Landraces and local varieties are saved simply because they fill ecological, cultural and local socioeconomic positions not occupied by modern varieties [
2]. Thus, it is very important to assess the genetic diversity of traditional local varieties. However, assessing their diversity is very difficult, due to the absence of standard references, confusion of “variety” names (homonymy and synonymy) and the existence of multiclonal varieties as a result of the richness of chestnut genetic heritage [
7,
8].
Traditional chestnut varieties are characterised according to the geographical origin, morphology, ripening period and type of use [
8]. In Crete, four traditional varietal names have been reported (Katharokastania, Koutsakera, Rogdiani and Strovliani); nevertheless, their classification is unclear due to the absence of pertinent studies.
This study reports the use of nSSR and EST-SSR markers for the identification of chestnut cultivars present in Crete. The null hypothesis of single-clone genotype-to-cultivar correspondence was tested. The main objective was to characterise the Cretan chestnut varieties genetically and to detect possible homonymies and synonymies.
3. Results
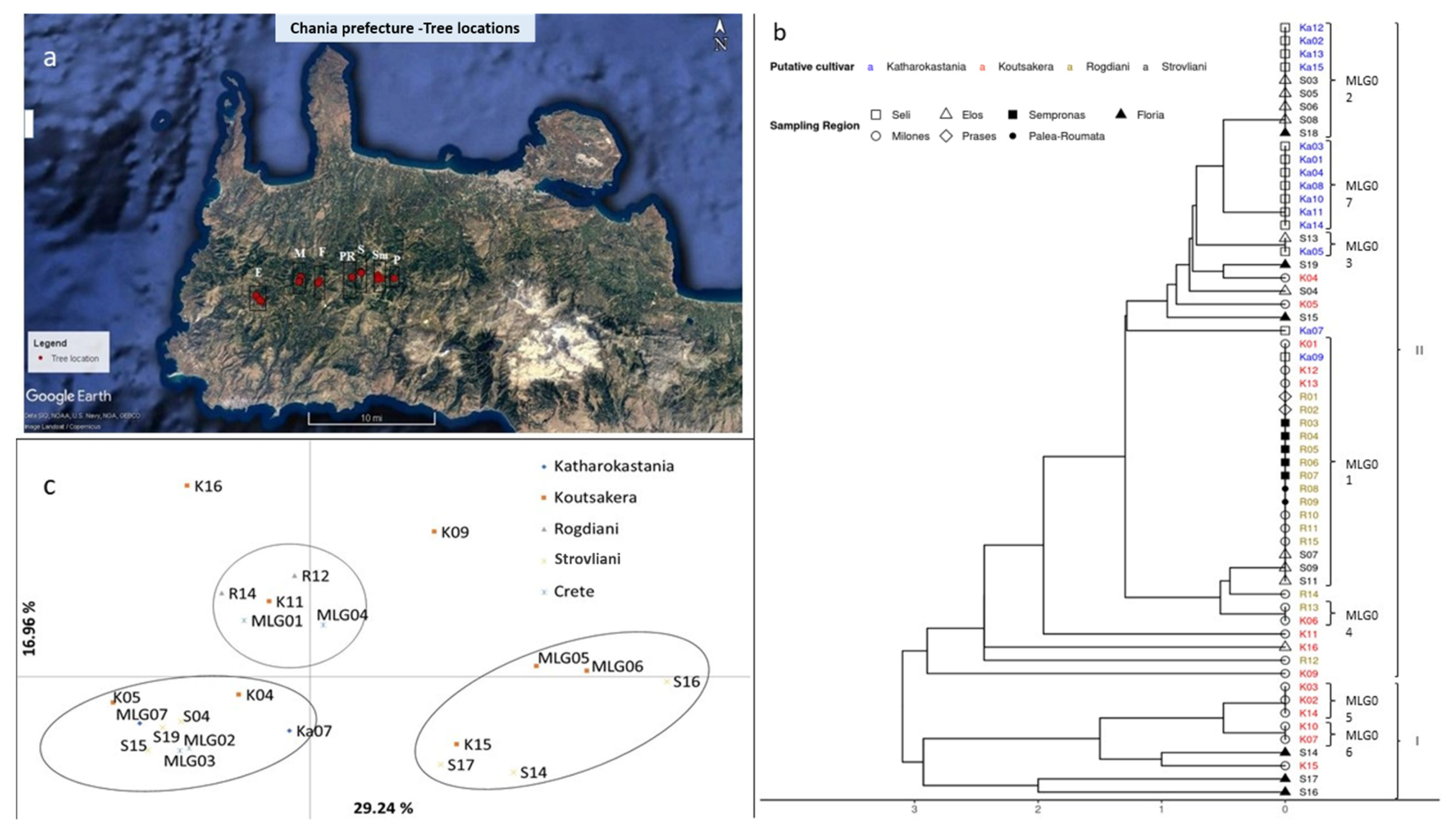
As a result of the multilocus genotype analysis, 22 unique genotypes were identified in the 4 chestnut Cretan cultivars (
Table 2). For the 59 chestnut orchard trees studied, 4 different groups of synonymies (genetically identical cultivars with different names), namely, MLG01 to MLG04, were identified. Regarding homonymies (genetically different cultivars with the same name), two homonymy groups were found in both Katharokastania and Strovliani, six in Rogdiani and eight in Koutsakera (
Table 2).
In total, 26 alleles were detected in the 7 microsatellite loci. The number of alleles per locus varied between two and seven (average of 3.7 alleles per locus). The most polymorphic locus was GOT021 that presented an observed heterozygosity (Ho) of 0.982 followed by FIR110 and CsCAT6, which presented 0.917 (
Table 3). The probability of identity (PI) ranged between 0.615 and 1.000 for WAG004 and CsCAT3, respectively (
Table 3).
The fixation index (Fst = 0.065) shows a relatively low level of differentiation between the cultivars. The average number of migrants ranged between 1.542 for CsCAT3 and 161.8 for GOT021 (
Table 3). According to the AMOVA, 73% of the total diversity resides within cultivars and 27% among cultivars (
Table A2 and
Table A3).
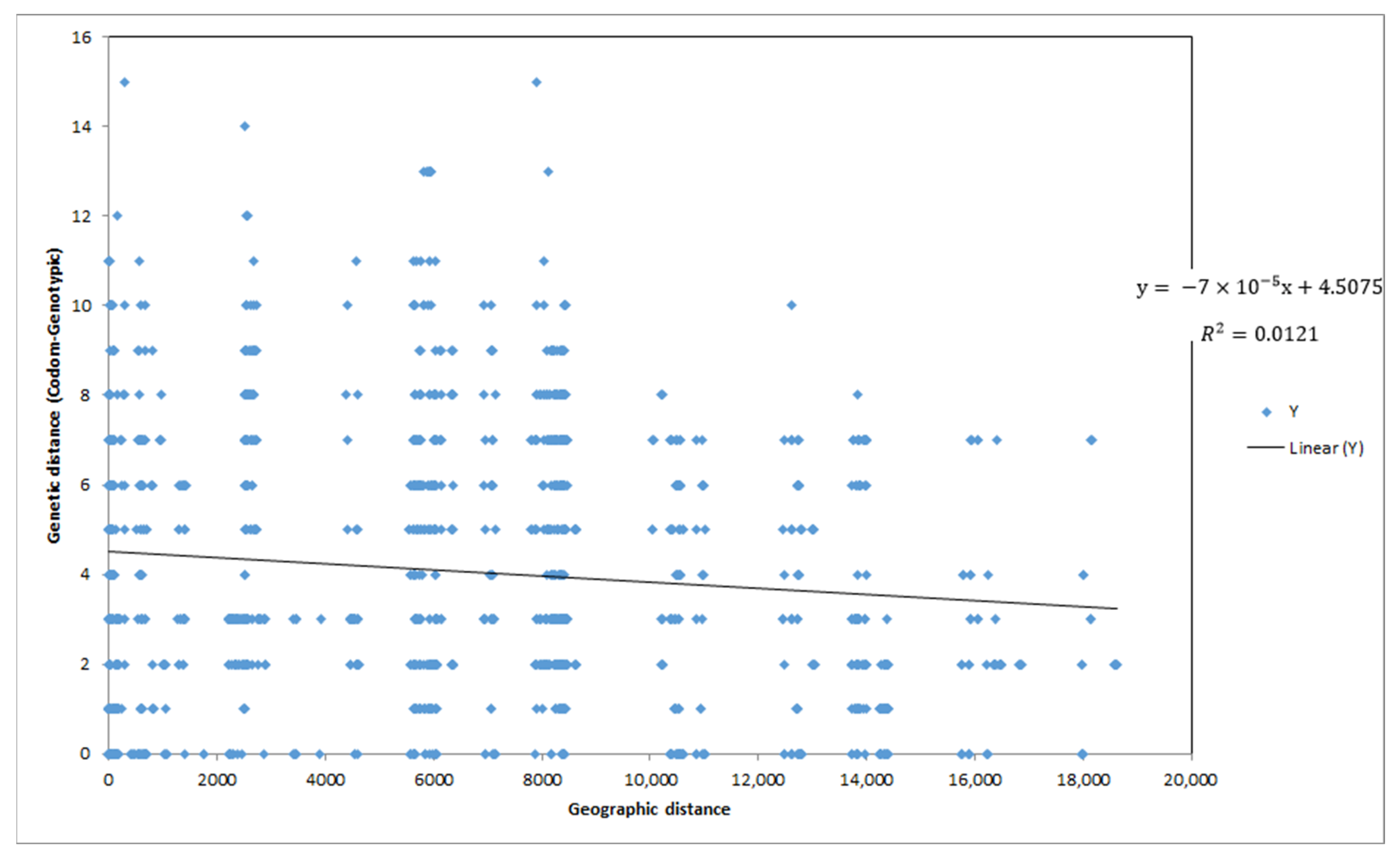
A very weak (Pearson’s
r = 0.11), but statistically significant (
p = 0.04) relationship was found between genetic and geographic distances (
Figure A2).
Moreover, the genetic diversity analysis of the separate nSSR and EST-SSR data sets showed that there are no significant differences in the genetic diversity parameters between them (
Table A4).
The UPGMA dendrogram distributes the 59 Cretan chestnut individuals across two main clusters at the six allele difference point (
Figure 1b). In general, cluster analysis shows a certain Rogdiani group. Moreover, most Katharokastania trees are also clustered together in two MLGs, which differ by only one allele. On the other hand, no clear classification could be made for Koutsakera and especially Strovliani.
In particular, cluster I, represented by a small number of individuals, gathers six putative Koutsakera trees, out of which five are distributed across MLG05 and MLG06 and differ by one allele and one more sample that bears a three allele difference to both MLGs. Cluster I also contains three putative Strovliani trees. Cluster II, which contains the rest of the 59 individuals studied, is represented as follows: (a) 12 Rogdiani individuals, sampled from four different areas, are present as one genotype (MLG01); (b) two other Rogdiani samples separated from the previous by only one allele; (c) an outlier Rogdiani individual “R12”, which differed from the main Rogdiani MLG by a five allele difference is also present; (d) Katharokastania individuals, sampled from the Seli region, grouped together in two MLGs (MLG02, MLG07) (MLG02 contains four Katharokastania individuals with only one allele difference from MLG07, which has seven Katharokastania trees); (e) three separated outlier Katharokastania trees, Ka05, Ka07 and Ka09, with a two to three allele difference from the two other Katharokastania MLGs; (f) Strovliani individuals, which show no clear grouping, present a first group of five individuals being genotypically identical to Katharokastania MLG02, a second group of three Strovliani individuals (S07, S09 and S11) present in the putative Rogdiani cluster group (MLG01), while four remaining Strovliani samples (S13, S19, S04 and S15) are scattered with differences of up to three alleles from MLG01 and MLG02. On the other hand, the remaining Koutsakera individuals are dispersed without an obvious pattern across various subgroups of cluster II (
Figure 1b).
A PCoA was carried out on the 22 clone-corrected data set. The first two components account for 59.73% of the total variance (
Figure 1c). Three different groups could be differentiated. Group I is represented by Rogdiani samples, with two Rogdiani individuals (R12 and R14), in addition to two MLG groups, MLG04 (K06 and R13) and MLG01 (mostly Rogdiani samples) (
Table 2;
Figure 1c). Group II is dominated by Katharokastania trees, containing Ka07, MLG07 (Katharokastania individuals), MLG02 (three Katharokastania individuals and five Strovliani individuals), MLG3 (Ka05 and S13), K05 and three Strovliani (S04, S15, S19) (
Table 2;
Figure 1c). Group III, consisting of Koutsakera and Strovliani cultivars, where Koutsakera dominates with two MLGs (MLG05 and MLG06) and one individual (K15), while Strovliani is represented by three individuals (S14, S16 and S17) (
Table 2;
Figure 1c).
As far as the sampling regions are concerned, diversity statistics are displayed in
Table 4. Allelic richness is reported (results for four gene copies) instead of the number of alleles, in order to account for the unbalanced sampling. The highest allelic richness value occurred in Floria (AR = 2.10) and the lowest in Sempronas (AR = 1.82). An interesting finding was the detection of five private alleles in the Milones region. An AMOVA partitioned the genetic diversity as 22% between regions and 78% within regions (
Table 5).
4. Discussion
In Crete, chestnut has always been an important component of the natural landscape and traditional agroforestry systems, but it has rarely been assessed genetically before. The present work offers a first genetic assessment of the main Cretan chestnut cultivar germplasm that consists of four widespread cultivars, by using a set of SSR primers. The multilocus genotype analysis showed that in these four Cretan chestnut cultivars there are twenty-two unique genotypes identified. Therefore, each cultivar appears to be a multiclonal variety. This result has also been found regarding other varieties and cultivars, for instance, in Greece [
7] and Spain [
2].
The results obtained with both nSSR and EST-SSR markers showed high levels of diversity for the Cretan chestnut cultivars, confirming the results obtained in previous studies conducted in European chestnut populations [
9,
12]. AMOVA revealed that the cultivars are genetically different (presenting some unique allele combinations) but share a common gene pool. These results are in agreement with the relevant literature on chestnut, where most of the variation (~70%) accounted for intracultivar differences [
24]. Moreover, the study of Poljak et al. [
25] on sweet chestnut wild germplasm and cultivated varieties sampled in central Europe and the western part of the Balkan Peninsula showed that most of the genetic diversity was attributed to the differences between individuals within populations (84.1%), which also supports our findings regarding the partitioning of variation within chestnut cultivars.
The Cretan cultivars exhibit a high level of gene diversity with an observed heterozygosity (Ho = 0.7). These findings are in accordance with the results of Martín et al. [
26], who used nine EST-SSR markers for evaluating Spanish and Italian cultivars (Ho = 0.5 and Ho = 0.6, respectively). On a cultivar basis, both allelic number and observed heterozygosity differ, a finding reflecting the presence of different numbers of genotypes per cultivar. The inbreeding coefficient (F) presents a negative value for all cultivars indicative of a high heterozygosity, and perhaps in relation to cultivar selection for fitness-related traits (such as growth and nut production). Heterozygosity has on numerous occasions been associated with growth and fructification, a phenomenon documented for several years now [
27]. In this case, it could be a result attributable to the repeated artificial selection process for higher growth and nut production.
Despite analysing 59 genotypes from a rather restricted geographic area (~2380 km
2), the polymorphism level was notable. The number of alleles per locus was lower compared to the results of other sweet chestnut studies, which nevertheless refer to wider areas. Martín et al. [
2] genotyped 100 chestnut trees grown in Andalusia (Huelva and Malaga) with seven microsatellites and detected an average of 5.4 and 7.4 alleles per locus for Huelva and Malaga, respectively. Torello Marinoni et al. [
9] genotyped 68 chestnut trees collected in different valleys in northwestern Italy with 10 SSRs and identified 80 alleles with an average of 8 alleles per locus. Moreover, Martín et al. [
28] genotyped 239 chestnuts trees in the north, centre and south (Andalusia) of Spain with seven microsatellites and found 13.14 alleles per locus.
The fixation index (Fst) of GOT021 is extremely low compared to the other EST-SSRs (Fst = 0.002), while the number of migrants shows a high value compared to the other EST-SSRs (Nm = 161.8), which indicates that this EST-SSR marker is an outlier marker. Moreover, as a functional marker, it is associated with loci involved in response to drought stress or trait of particular interest [
13,
29]. This result is in accordance with the findings of numerous relevant studies on
Castanea sativa and several
Quercus species, where GOT021 was one of the EST-SSR markers associated with abiotic stress. It was shown, for instance, to be under divergent selection in
Quercus species [
13,
30]. Moreover, [
13,
28,
29] did not find strong evidence that this marker is under selection, but they detected private alleles for both tolerant and susceptible
Castanea sativa trees under drought stress. Additional sampling on a larger area and further analysis (such as a water stress experiment using dedicated genotypes) could provide better insight on GOT021 into adaptation of Cretan chestnut populations.
Cluster analysis revealed a clear connection between some cultivar names as provided by the farmers and the molecular result, while, for some others, no clear correspondence was found. In fact, the Rogdiani cultivar individuals with the same genetic profile sampled from four different regions showed a high degree of confidence regarding cultivar identity. However, it should be pointed out that putative samples from the other three cultivars were also shown to have the same genotype (MLG01). The Katharokastania cultivar samples were almost exclusively grouped into two closely related MLGs (MLG02, MLG07); however, sampling Katharokastania individuals from only one region (Seli), in addition to the presence of some outlier Katharokastania individuals within other cultivar groups, blurs the assertion for cultivar identity. Regarding the Koutsakera and Strovliani cultivars, no clear molecular identification was possible, as individuals from both cultivars are dispersed across various subgroups. Nevertheless, it is worth noting that a subsample of putative Koutsakera samples was grouped into a unique cluster identified in the dendrogram (cluster I) as well as in the PCoA.
The PCoA results are in accordance with the cluster analysis results, revealing the Rogdiani and Katharokastania cultivar identity (the latter with less confidence, as some individuals from other cultivars were grouped within the Katharokastania group). On the other hand, no clear differentiation for the Koutsakera and Strovliani cultivars was seen. As the chestnut cultivars studied are classified as widespread traditional varieties in Crete, it is not uncommon to have cases where misnaming of some trees can occur between farmers, especially if those cultivars share some fruit characteristics.
There were 18 homonym and 4 synonym groups distinguished in this study. As the original cultivar identification by name was given during sampling by the respective owners of the orchards where these traditional varieties are present, there appears to be some confusion in practice as to cultivar identity. The same was observed in Spain; [
2] reported two homonym and four synonym groups found among the Spanish cultivars studied. Given the amount of genetic variation found and the cases of homonyms and synonyms, the sample size per nominal cultivar to assess intracultivar variation should be increased in future studies. The four cultivars could be distinguished in two groups. Katharokastania and Rogdiani are relatively low-number multilocus genotype cultivars (five and four MLGs, respectively), while Koutsakera and Strovliani can be considered as high-number multilocus genotype cultivars (10 and 9 MLGs, respectively). In terms of chestnut crop uniformity, the nuts produced from Katharokastania and Rogdiani are expected to be more uniform given their low MLG number. On the other hand, Koutsakera and Strovliani present low uniformity which reduces the market value of their chestnut crop. In fact, Koutsakera, a low uniformity variety, has the highest standard deviation in nut area, while Katharokastania, a high uniformity variety, has the least (El Chami et al., unpublished results). Nevertheless, under strong environmental pressure (for example, under climatic change), more MLGs within a cultivar should offer better long-term orchard stability.
No relationship was found between genetic and geographic distances, similar to the results reported for Spanish cultivars [
2,
24]. Due to isolation by distance, in typical natural populations, genetic and spatial distances are usually positively correlated. On the other hand, an orchard where the placement of cultivars on the ground is an anthropogenic exercise explains the absence of a relation between genetic and geographic distance. This finding has also been observed in a similar study in Greece [
10]. This result also indicates that the chestnut orchards in the Chania region are either planted or grafted onto wild rootstock; the presence of any remnants of old-growth natural chestnut forest in the studied region is not supported by our results. Groups of trees from all regions exhibited similar levels of genetic diversity. Surprisingly, five out of the six private alleles detected in this study originated in Milones, which might reflect a greater significance of this area as a centre of traditional chestnut cultivar diversity. However, some caution should be exercised in this interpretation, due to the unbalanced sampling design. The fact that AMOVA partitioned most of the genetic variation within regions (78% within regions, 22% between regions) points towards the employment of similar cultivation and/or domestication practices in the past centuries throughout the investigated area.


 ,
, 



{kind=link}
{kind=link}
{kind=link}


