Climate and Management Factors Underlying Changes in Beech Forest Herbaceous Layer Plant Communities in the Polish Eastern Carpathians


Abstract
:1. Introduction
2. Materials and Methods
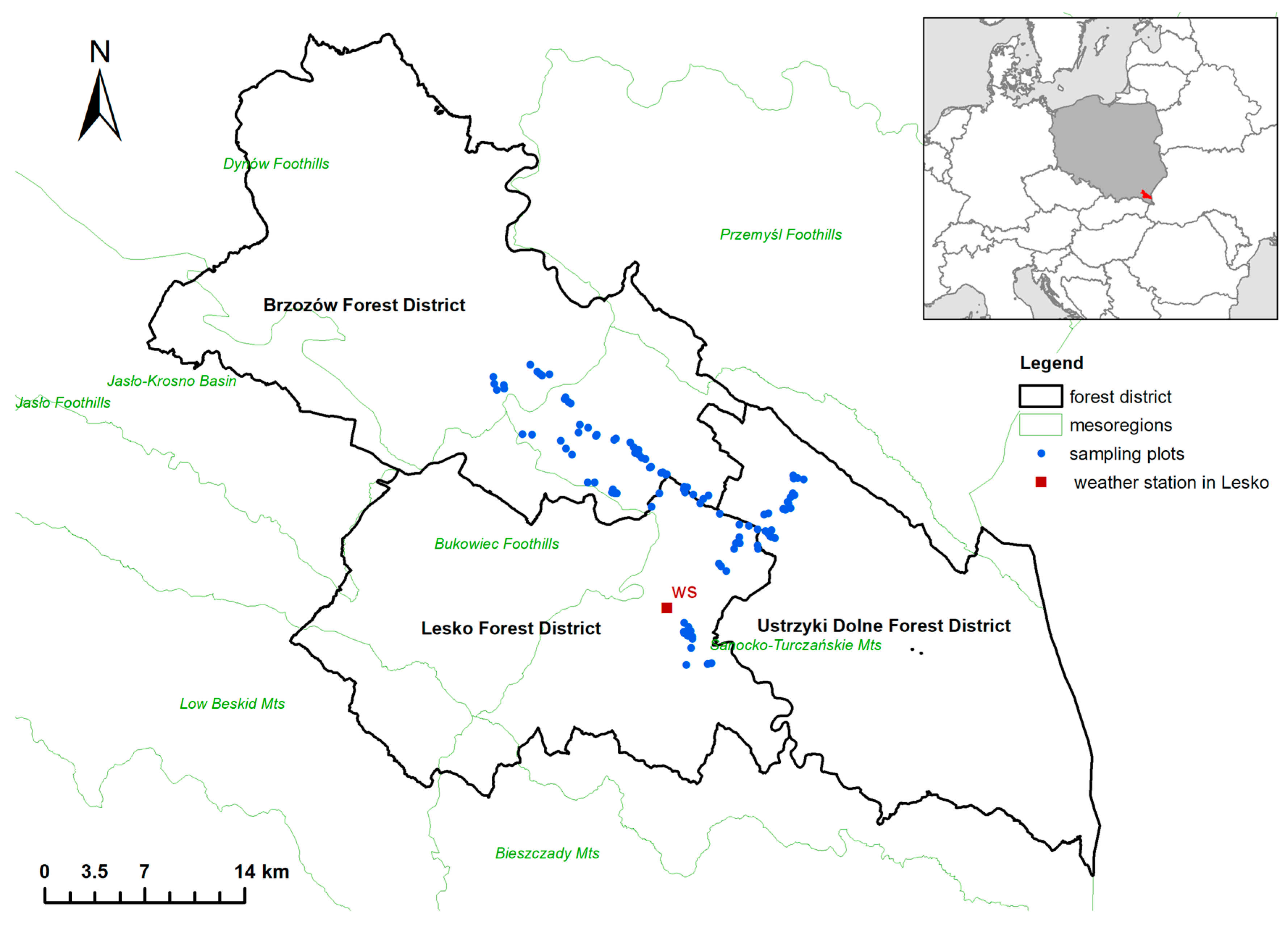
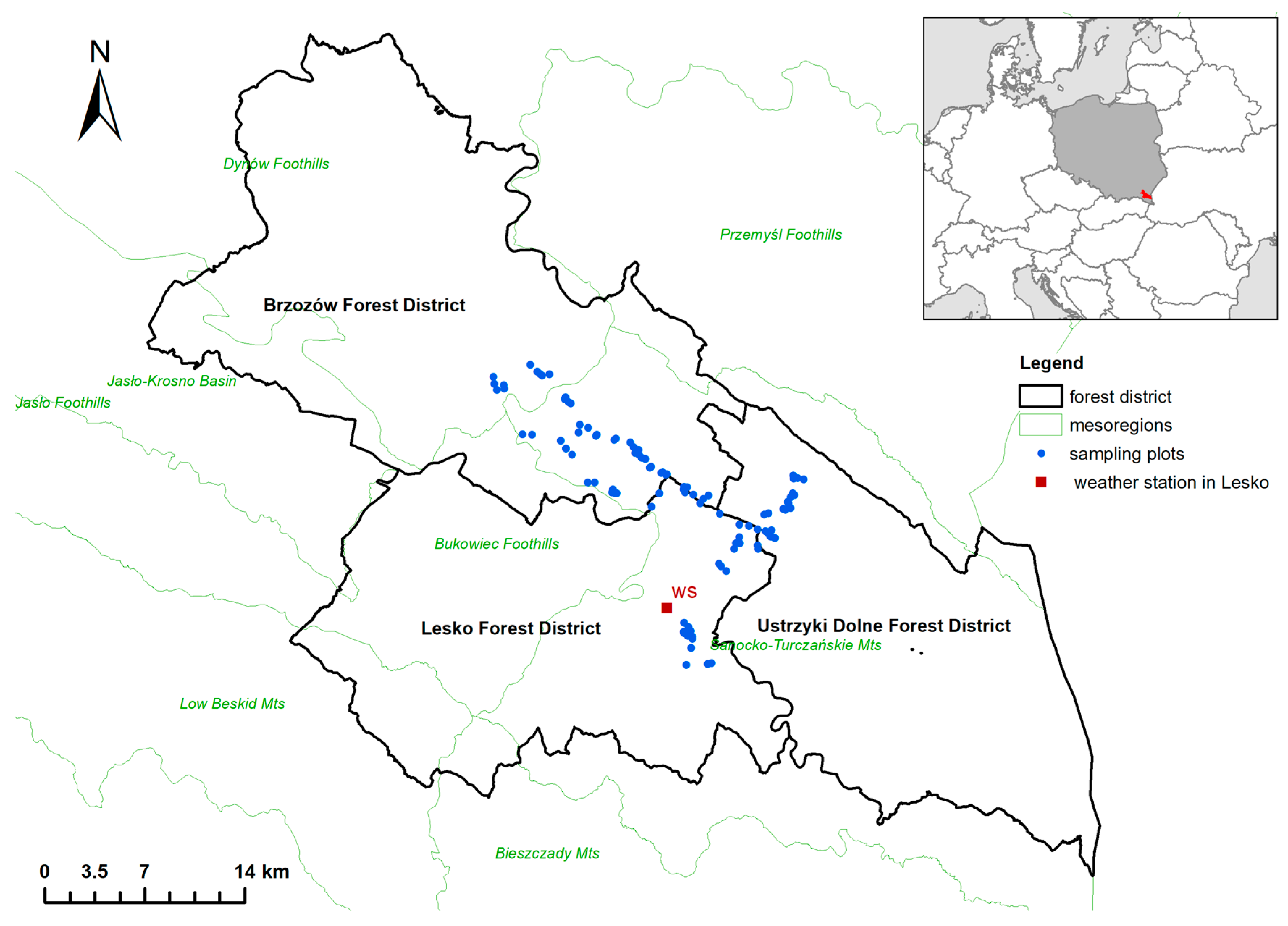
2.1. Study Area
2.2. Data Collection
2.3. Analysis of Forest Vegetation
3. Results
3.1. Changes in Forest Cover
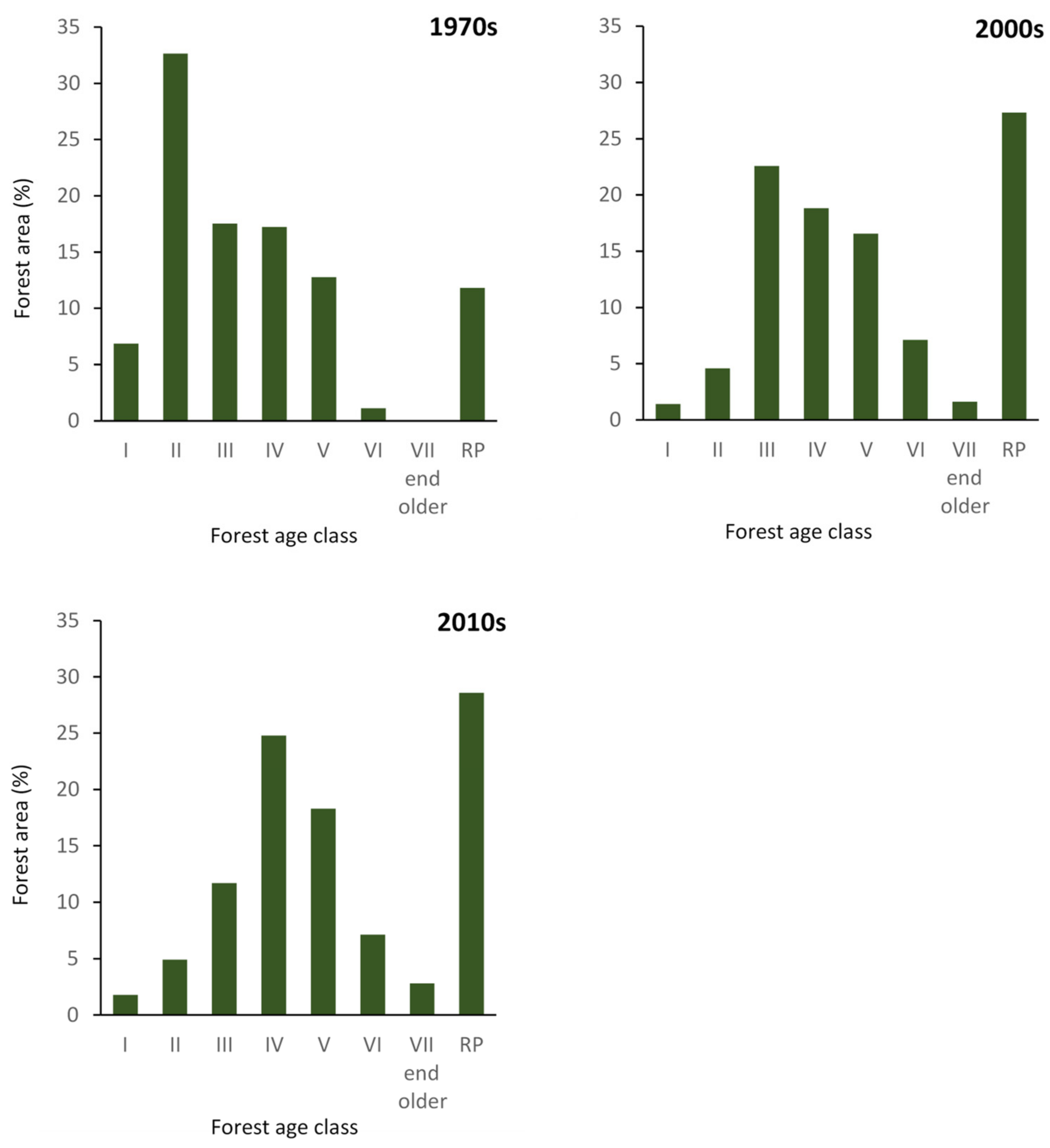
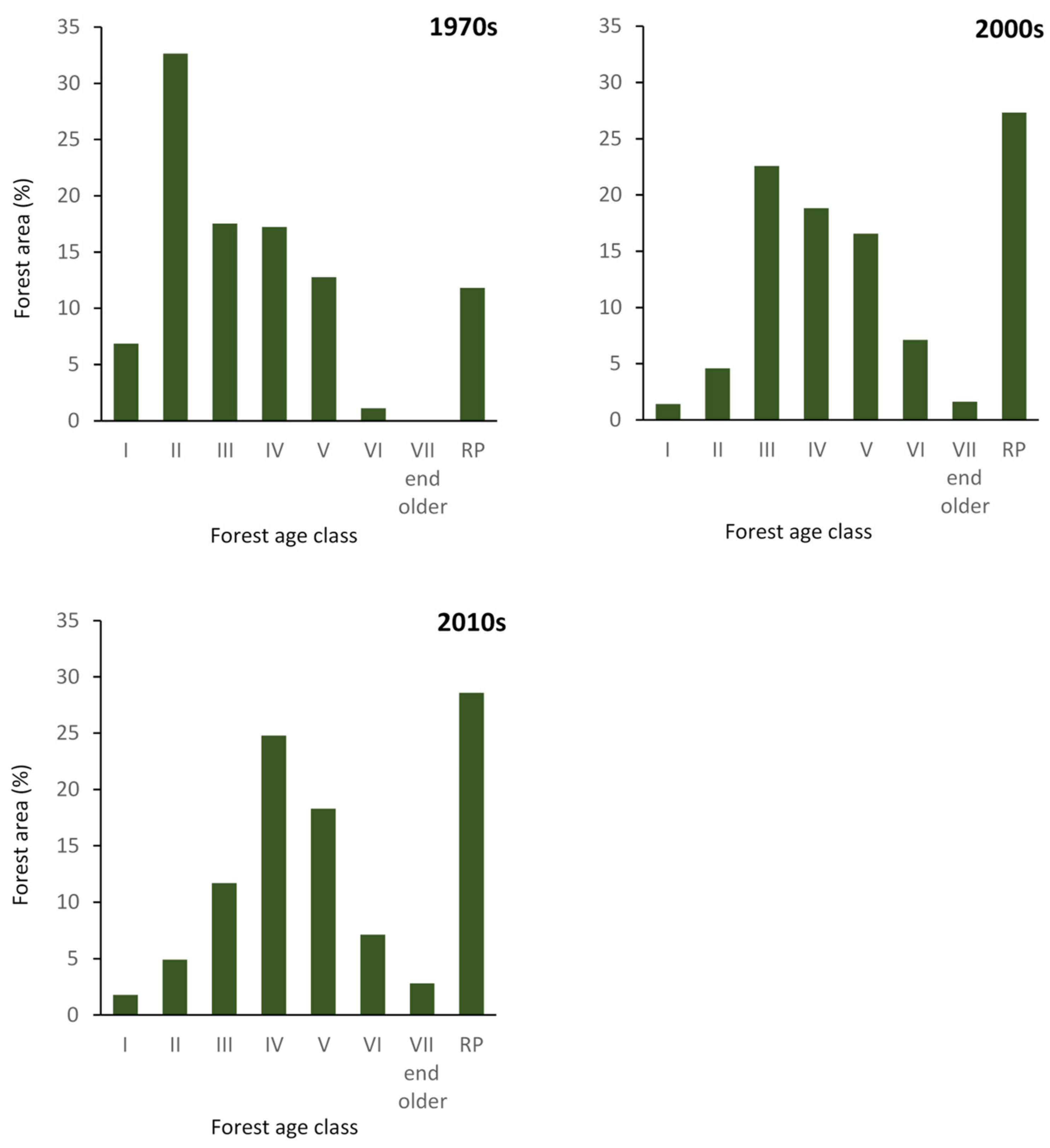
3.2. Changes in Stand Structure and Forest Management
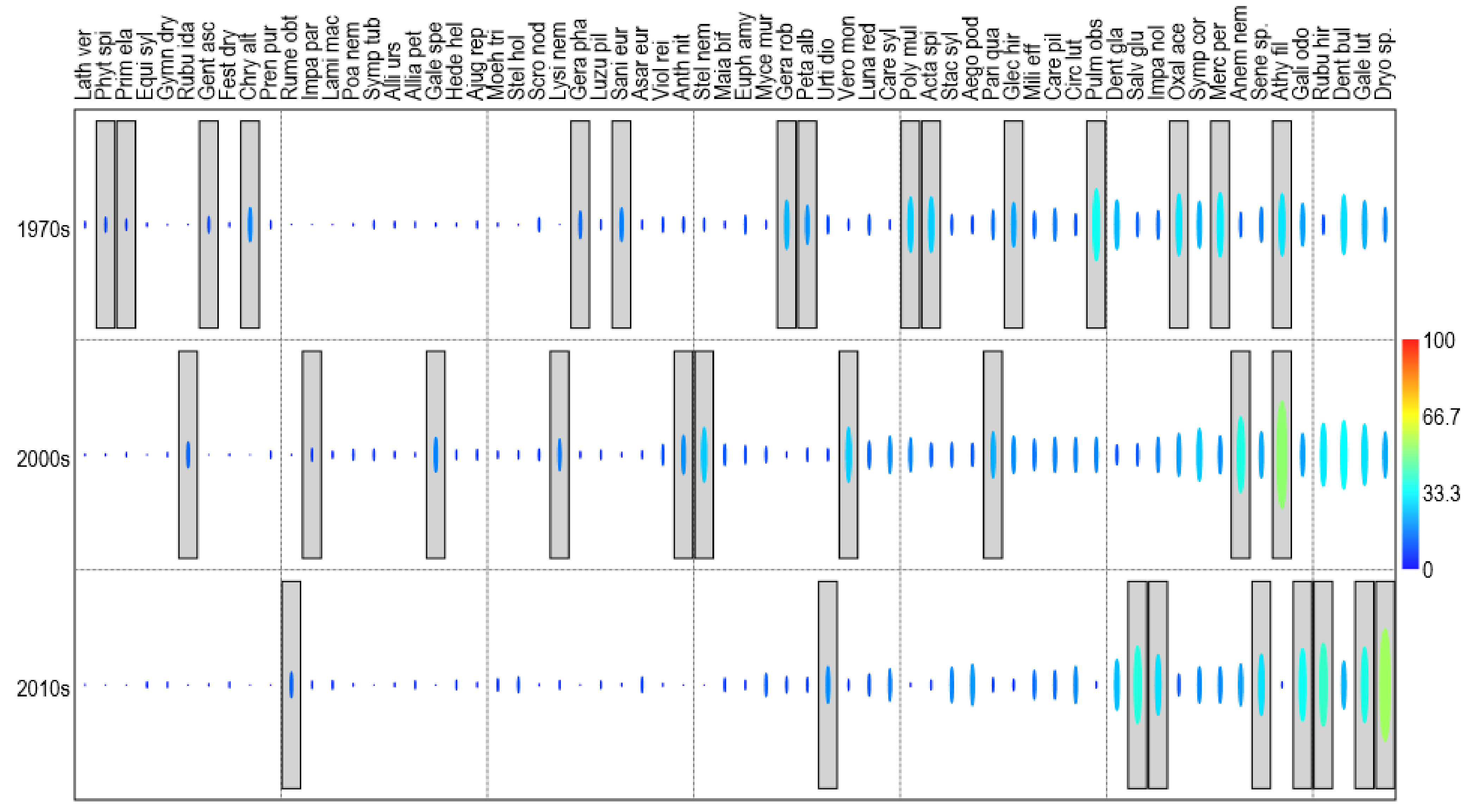
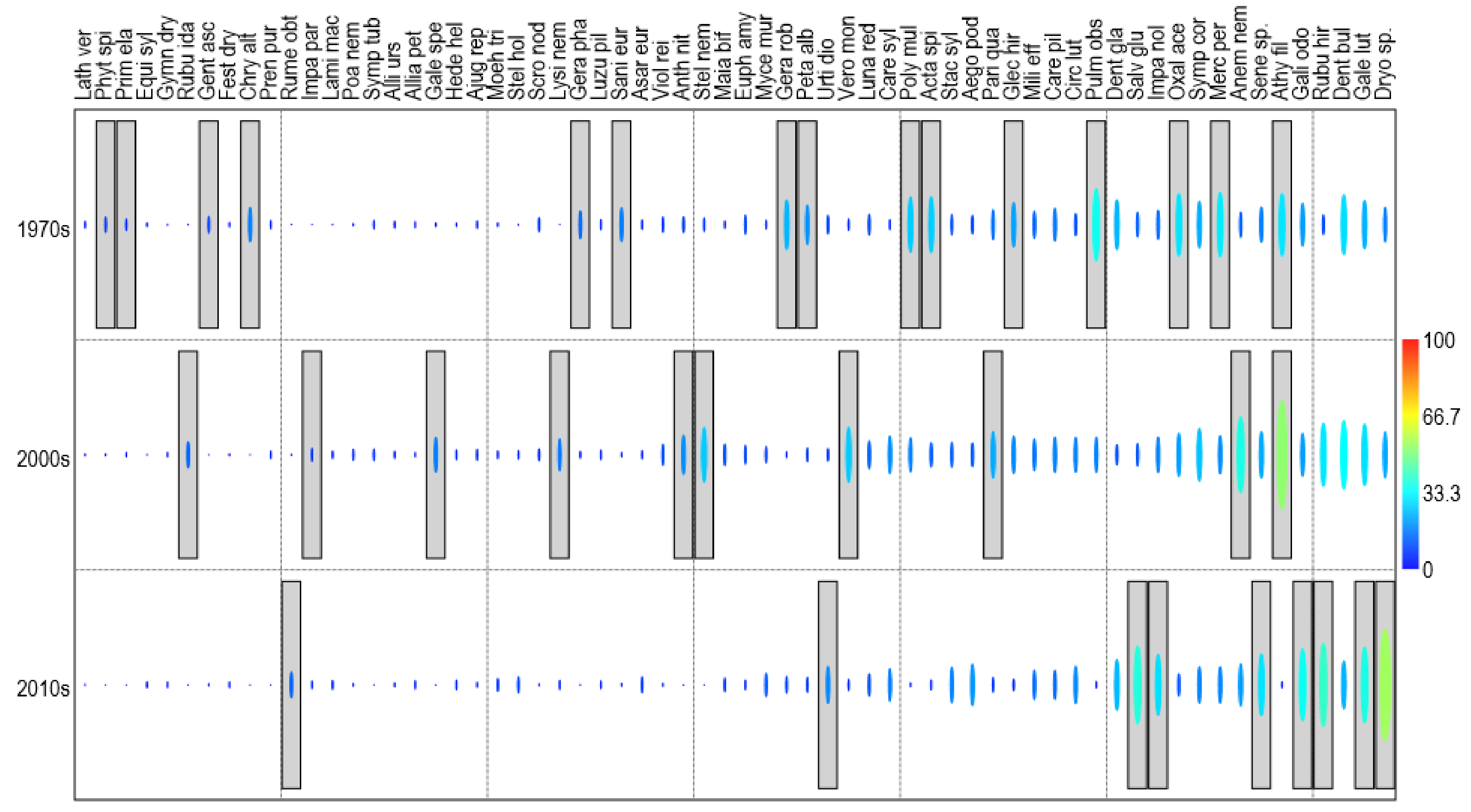
3.3. Changes in Beech Forest Communities
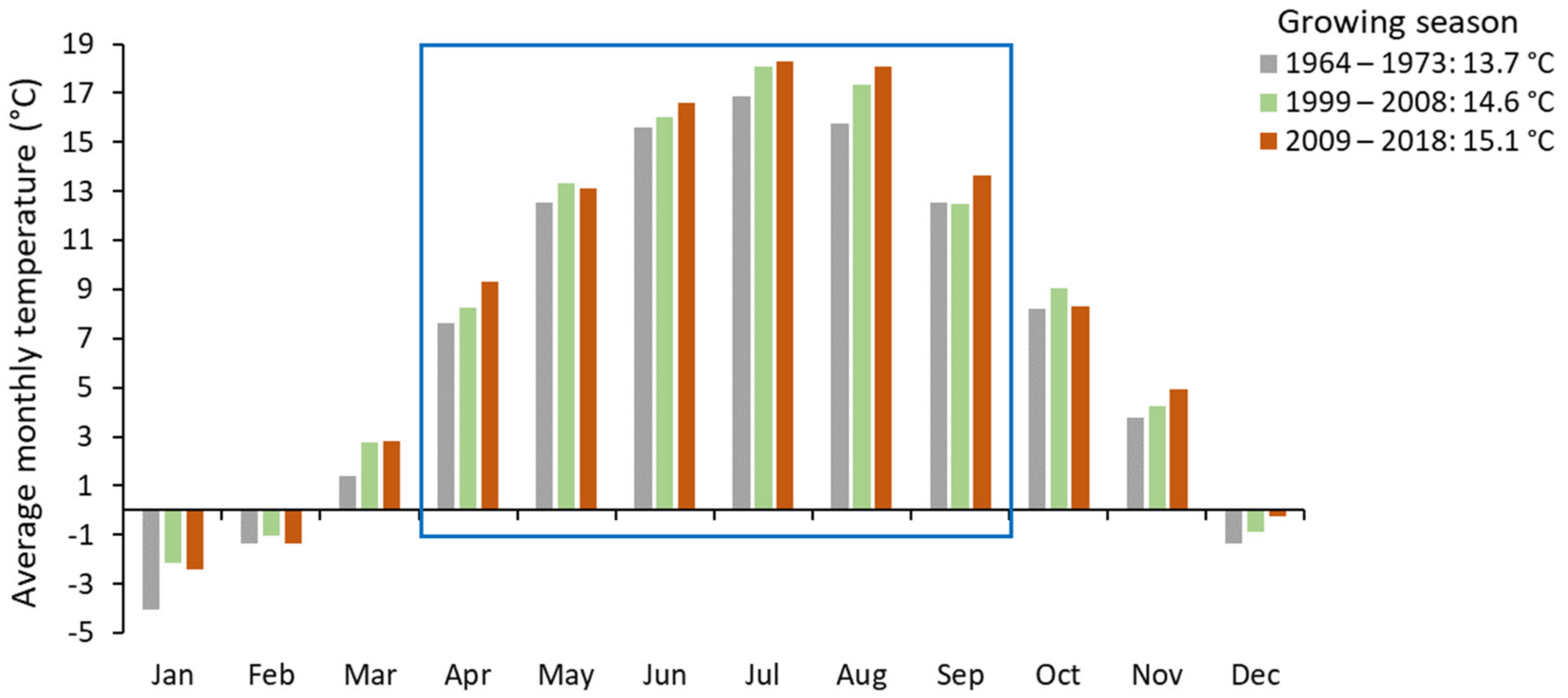
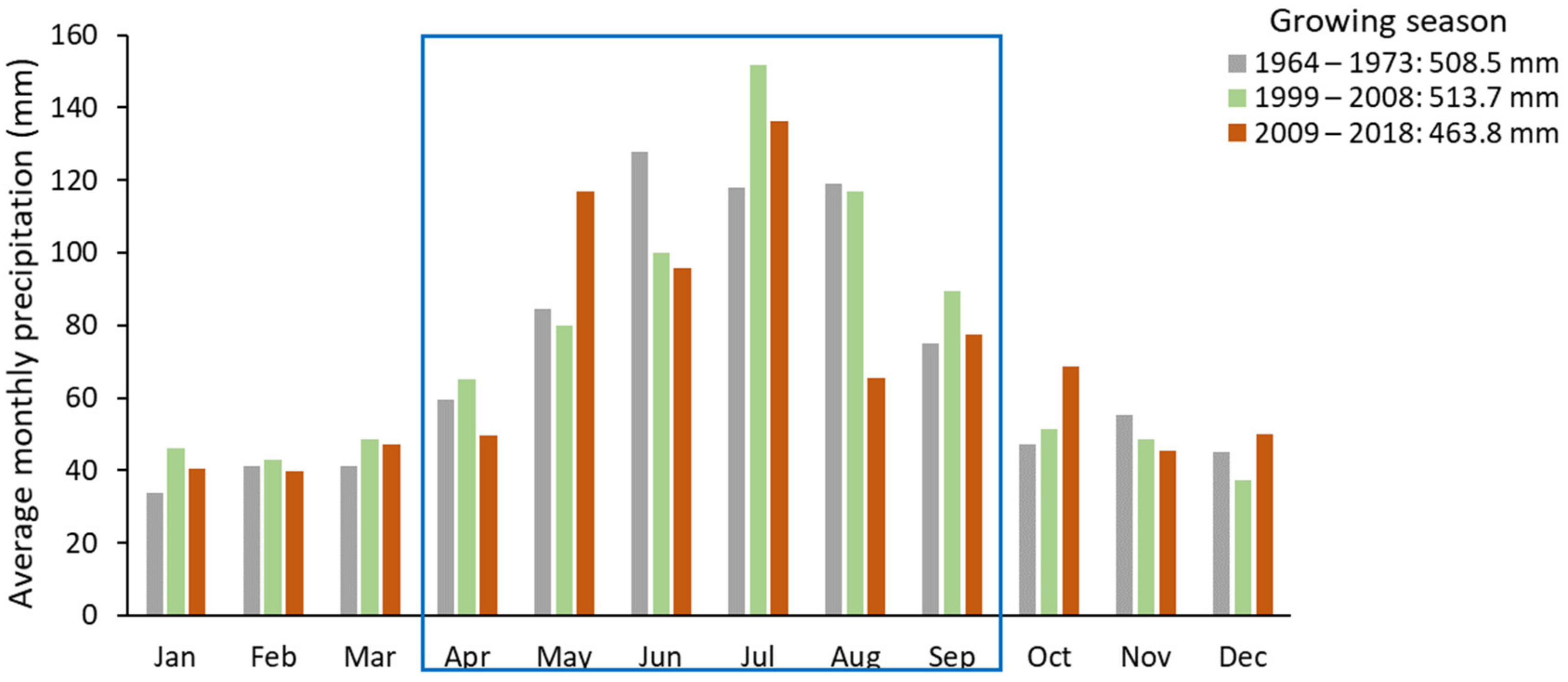
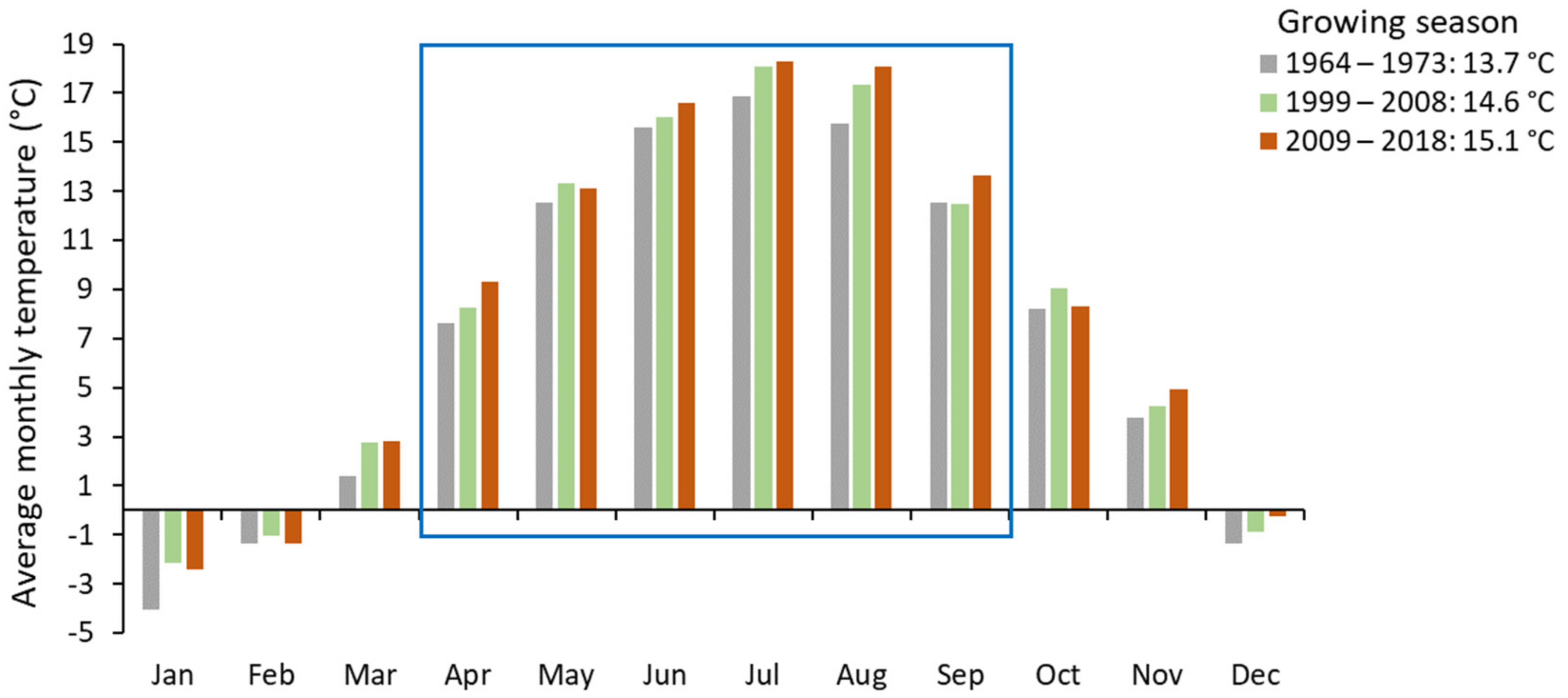
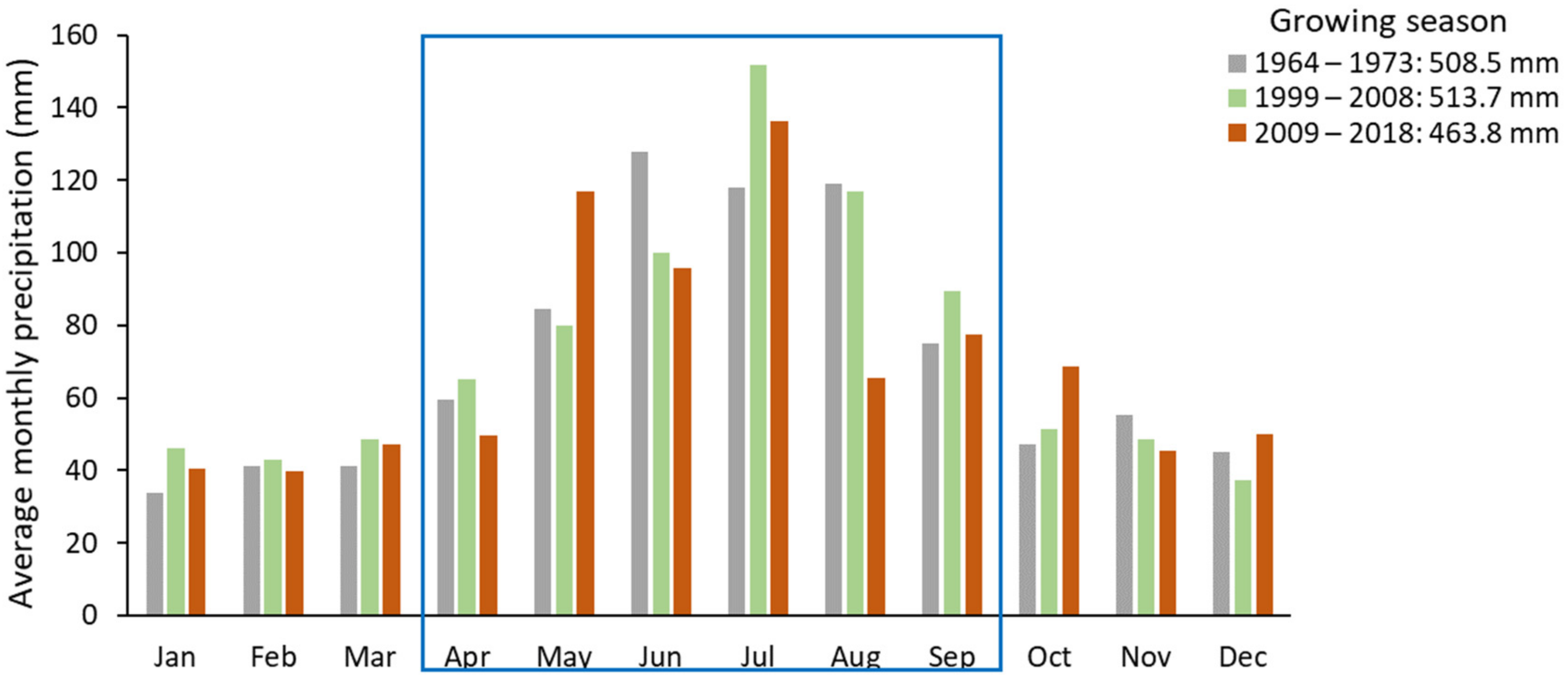
3.4. Climatic Conditions
4. Discussion
4.1. Impact of Changes in Forest Area
4.2. Impact of Changes in Stand Structure and Forest Management
4.3. Impact of Climate Change
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Führer, E. Forest functions, ecosystem stability and management. For. Ecol. Manag. 2000, 132, 29–38. [Google Scholar] [CrossRef]
- Święcicki, Z. (Ed.) Instrukcja Urządzania Lasu. Cz. 1. Instrukcja Sporządzania Planu Urządzenia Lasu dla Nadleśnictwa; CILP: Warsaw, Poland, 2012. (In Polish) [Google Scholar]
- Vedeld, P.; Angelsen, A.; Bojö, J.; Sjaastad, E. Forest environmental incomes and the rural poor. For. Policy Econ. 2007, 9, 869–879. [Google Scholar] [CrossRef]
- Varma, V.K.; Ferguson, I.S.; Wild, I. Decision support system for the sustainable forest management. For. Ecol. Manag. 2000, 128, 49–55. [Google Scholar] [CrossRef]
- European Forest Ecosystems–State and trends EEA Report; European Environment Agency: Copenhagen, Denmark, 2016; p. 5.
- Aggestam, F.; Konczal, A.; Sotirov, M.; Wallin, I.; Paillet, Y.; Spinelli, R.; Lindner, M.; Derks, J.; Hanewinkel, M.; Winkel, G. Can nature conservation and wood production be reconciled in managed forests? A review of driving factors for integrated forest management in Europe. J. Environ. Manage. 2000, 268, 110670. [Google Scholar] [CrossRef]
- Blicharska, M.; Angelstam, P.; Giessen, L.; Hilszczański, J.; Hermanowicz, E.; Holeksa, J.; Jacobsen, J.B.; Jaroszewicz, B.; Konczal, A.; Konieczny, A.; et al. Between biodiversity conservation and sustainable forest management—A multidisciplinary assessment of the emblematic Białowieża Forest case. Biol. Conserv. 2020, 248, 108614. [Google Scholar] [CrossRef]
- Maier, C.; Winkel, G. Implementing nature conservation through integrated forest management: A street-level bureaucracy perspective on the German public forest sector. For. Policy Econ. 2017, 82, 14–29. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human Domination of Earth’s Ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson, J.; Nilsson, S.G.; Menozzi, P. Biodiversity, disturbances, ecosystem function and management of European forests. For. Ecol. Manag. 2000, 132, 39–50. [Google Scholar] [CrossRef]
- Kindstrand, C.; Norman, J.; Boman, M.; Mattsson, L. Attitudes towards various forest functions: A comparison between private forest owners and forest officers. Scand. J. For. Res. 2008, 23, 133–136. [Google Scholar] [CrossRef]
- Kindler, E. A comparison of the concepts: Ecosystem services and forest functions to improve interdisciplinary exchange. For. Policy Econ. 2016, 67, 52–59. [Google Scholar] [CrossRef]
- Stephen, R.; Shifley, R.; Moser, K.; Nowak, D.J.; Miles, P.D.; Butler, B.J.; Aguilar, F.X.; DeSantis, R.D.; Greenfield, E.J. Five anthropogenic factors that will radically alter forest conditions and management need in the Northern United States. For. Sci. 2014, 60, 914–925. [Google Scholar] [CrossRef]
- Ortiz, J.C.; Espinosa, C.I.; Dahik, C.Q.; Mendoza, Z.A.; Ortiz, E.C.; Gusmán, E.; Weber, M.; Hildebrandt, P. Influence of Anthropogenical Factors on the Diversity and Structure of a Dry Forest in the Central Part of the Tumbesian Region (Ecuador-Perú). Forests 2019, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- Gautam, K.H.; Devoe, N.N. Ecological and anthropogenic niches of sal (Shorea robusta Gaertn. f.) forest and prospects for multiple-product forest management—A review. Forestry 2006, 79, 81–101. [Google Scholar] [CrossRef]
- Puettmann, K.J.; Wilson, S.M.; Baker, S.C.; Donoso, P.J.; Drössler, L.; Amente, G.; Harvey, B.D.; Knoke, T.; Lu, Y.; Nocentini, S.; et al. Silvicultural alternatives to conventional even-aged forest management—What limits global adoption? For. Ecosyst. 2015, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Tylianakis, J.M.; Didham, R.K.; Bascompte, J.; Wardle, D.A. Global change and species interactions in terrestrial ecosystems. Ecol. Lett. 2008, 11, 1351–1363. [Google Scholar] [CrossRef]
- Kuemmeerle, T.; Hostert, P.; Radeloff, V.C.; Perzanowski, K.; Kruhlov, I. Post-socialist forest disturbance in the Carpathian Border region of poland, Slovakia, and Ukraine. Ecol. Appl. 2007, 17, 1279–1295. [Google Scholar] [CrossRef]
- Kuemmeerle, T.; Hostert, P.; Radeloff, V.C.; Linden, S.; Perzanowski, K.; Kruhlov, I. Cross-border Comparison of Post-socialist Farmland Abandonment in the Carpathians. Ecosystems 2008, 11, 614–628. [Google Scholar] [CrossRef]
- Kubijowicz, W. Życie Pasterskie w Beskidach Wschodnich; Prace Instytutu Geografii UJ: Kraków, Poland, 1926. (In Polish) [Google Scholar]
- Skała, M.; Wolski, J. Krajobraz (nie)pamięci–teraźniejszość nadpisująca przeszłość. Osadnictwo i ludowość. In Bojkowszczyzna Zachodnia-Wczoraj, Dziś i Jutro, Tom 2; Instytut Geografii i Przestrzennego Zagospodarowania in Stanisława Leszczyckiego Monografie: Warsaw, Poland, 2016; Volume 37, pp. 347–377. (In Polish) [Google Scholar]
- Durak, T.; Żywiec, M.; Kapusta, P.; Holeksa, J. Impact of land use and climate changes on expansion of woody species on subalpine meadows in the Eastern Carpathians. For. Ecol. Manag. 2015, 339, 127–135. [Google Scholar] [CrossRef]
- Kozak, J. Forest Cover Change in the Western Carpathians in the Past 180 Years. A Case Study in the Orawa Region in Poland. Mt. Res. Dev. 2003, 23, 369–375. [Google Scholar] [CrossRef] [Green Version]
- Kozak, J.; Kaim, D. Forecom; Instytut Geografii i Gospodarki Przestrzennej UJ: Krakow, Poland, 2016. (In Polish) [Google Scholar]
- Kozak, J.; Ziółkowska, E.; Vogt, P.; Dobosz, M.; Kaim, D.; Kolecka, N.; Ostafin, K. Forest-Cover Increase Does Not Trigger Forest-Fragmenntation Decrease: Case Study from the Polish Carpathians. Sustainability 2018, 10, 1472. [Google Scholar] [CrossRef] [Green Version]
- Matlack, G.R.; Monde, J. Consequences of low mobility in spatially and temporally heterogeneous ecosystems. J. Ecol. 2004, 92, 1025–1035. [Google Scholar] [CrossRef]
- Matlack, G.R. Slow plants in a fast forest: Local dispersal as a predictor of species frequencies in a dynamic landscape. J. Ecol. 2005, 93, 50–59. [Google Scholar] [CrossRef]
- Marszałek, E. The forest management in the Carpathian part of Regional Directorate of the State Forest in Krosno and its influence on the protection of nature. Rocz. Bieszcz. 2011, 19, 59–75. (In Polish) [Google Scholar]
- Varga, A.; Molnár, Z.; Biró, M.; Demeter, L.; Gellény, K.; Miókovics, E.; Molnár, Á.; Molnár, K.; Ujházy, N.; Ulicsni, V.; et al. Changing year-round habitat use of extensively grazing cattle, sheep and pigs in East-Central Europe between 1940 and 2014: Consequences for conservation and policy. Agric. Ecosyst. Environ. 2016, 234, 142–153. [Google Scholar] [CrossRef] [Green Version]
- Ustawa z dnia 28 Września 1991 r. o Lasach. Dz. U. 1991 Nr 101 poz. 444 (t.j. Dz. U. z 2020 r. poz. 1463). Available online: https://isap.sejm.gov.pl/isap.nsf/DocDetails.xsp?id=WDU19911010444 (accessed on 16 March 2021). (In Polish)
- Helmy, N.; Essl, F.; Mirtl, M.; Dirnoböck, T. Multiple environmental changes drive forest floor cegetation in a temperate mountain forest. Ecol. Evol. 2017, 7, 2155–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, M.; Weland, N.; Platner, C.; Schaefer, M.; Leuschner, C.; Thomas, F.M. Nutrient release from decomposing leaf litter of temperate deciduous forest trees along a gradient of increasing tree species diversity. Soil Biol. Biochem. 2009, 41, 2122. [Google Scholar] [CrossRef]
- Verheyn, K.; Baeten, L.; De Frenne, P.; Bernhardt-Römermann, M.; Brunet, J.; Cornelis, J.; Decocq, G.; Dierschke, H.; Eriksson, O.; Hédl, R.; et al. Driving factors behind the eutrophication signal in understorey plant communities of deciduous temperate forests. J. Ecol. 2012, 100, 352–365. [Google Scholar] [CrossRef]
- Majchrowska, A. The History of State Forest and Forestry in Poland. Manag. North. Eur. For. 2018, 12, 318–358. [Google Scholar] [CrossRef]
- Jaworski, A.; Kołodziej, Z. Beech (Fagus sulvatica L.) forest of a selection structure in the Bieszczady Mountains (Southeastern Poland). J. For. Sci. 2004, 50, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Jaworski, A. Hodowla Lasu. Tom, I. Sposoby Zagospodarowania, Odnawianie Lasu, Przebudowa i Przemiana Drzewostanów; Powszechne Wydawnictwo Rolnicze i Leśne: Warsaw, Poland, 2018. (In Polish) [Google Scholar]
- Ellenberg, H.; Leuscher, C. Vegetation Mitteleuropas mit den Alpen in Ökologischer, Dynamischer und Historischer Sicht; Ulmer: Stuttgart, Germany, 2010. (In German) [Google Scholar]
- Mölder, A.; Streit, M.; Schmidt, W. When beech strikes back: How strict nature conservation reduces herb-layer diversity and productivity in Central European deciduous forests. For. Ecol. Manag. 2014, 319, 51–61. [Google Scholar] [CrossRef]
- Augusto, L.; Dupouey, J.; Ranger, J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann. For. Sci. 2003, 60, 823–831. [Google Scholar] [CrossRef]
- Durak, T.; Holeksa, J. Biotic homogenisation and differentiation along a habitat gradient resulting from the ageing of manager beech stands. For. Ecol. Manag. 2015, 351, 47–56. [Google Scholar] [CrossRef]
- Vacek, S.; Černý, T.; Vacek, Z.; Podrázský, V.; Mikeska, M.; Králíček, I. Long-term changes in vegetation annd site conditions in beech and spruce forest of lower mountains ranges of Central Europe. For. Ecol. Manag. 2017, 398, 75–90. [Google Scholar] [CrossRef]
- Vacek, S.; Prokůpková, A.; Vacek, Z.; Bulušek, D.; Šimůnek, V.; Králíček, I.; Prausová, R.; Hájek, V. Growth response of mixed beech forests to climate change, various management and game pressure in Central Europe. J. For. Sci. 2019, 65, 331–345. [Google Scholar] [CrossRef]
- Bosela, M.; Štefančík, I.; Petráš, R.; Vacek, S. The effects of climate Warming on the growth of European beech forest depend critically on thinning strategy and site productivity. Agric. For. Meteorol. 2016, 222, 21–31. [Google Scholar] [CrossRef]
- Krupková, L.; Havránková, K.; Krejza, J.; Sedlák, P.; Marek, M.V. Impact of water scarcity on spruce and beech forests. J. For. Res. 2019, 30, 899–909. [Google Scholar] [CrossRef]
- Parpan, V.I.; Stojko, S.M.; Parpan, T.V. Phytocoenotical and ecological characterization of beech forests (Fagus sylvaticae L.) of Ukraine and possibility to expand their area due to global Warming. Infrastruktura i Ekologia Terenów Wiejskich 2013, 2, 29–44. [Google Scholar]
- Melo, M.; Lapin, M.; Kapolková, H.; Pecho, J.; Kružicová, A. Climate Trends in the Slovak Part of the Carpathians. In The Carpathians: Integrating Nature and Society Towards Sustainability. Environmental Science and Engineering; Kozak, J., Ostapowicz, K., Bytnerowicz, A., Wyżga, B., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 131–150. [Google Scholar] [CrossRef]
- Henrichs, S.; Winterhoff, W.; Schmidt, W. Vegetation dynamics of beech forests on limestone in central Germany over half a century–Effects of climate change, forest management, eutrophication or game browsing? Biodivers. Ecol. 2012, 4, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Zellweger, F.; De Frenne, P.; Lenoir, J.; Van Gansbeke, P.; Verheyen, K.; Bernhardt-Römermann, M.; Baeten, L.; Hédl, R.; Berki, I.; Brunet, J.; et al. Forest microclimate dynamics drive plant responses to Warming. Science 2020, 368, 772–775. [Google Scholar] [CrossRef]
- Felsmann, K.; Baudis, M.; Kayler, Z.E.; Puhlmann, H.; Ulrich, A.; Gessler, A. Responses of the structure and function of the understory plant communities to preciptation reduction across forest ecosystems in Germany. Ann. For. Sci. 2018, 75, 3. [Google Scholar] [CrossRef] [Green Version]
- Archaux, F.; Wolters, V. Impact of summer drought on forest biodiversity: Why do we know? Ann. For. Sci. 2006, 63, 645–652. [Google Scholar] [CrossRef]
- Dzwonko, Z. Forest communities of the Gory Słonne Range (Polish Eastern Carpathians). Fragm. Flor. Geobot. 1977, 23, 161–200. [Google Scholar]
- Skiba, S.; Drewnik, M. Soil map of the polish carpathian mountains. Rocz. Bieszcz. 2003, 11, 15–20. [Google Scholar]
- Meteorological Data. Averages and Monthly Totals. Lesko. Available online: https://meteomodel.pl (accessed on 16 March 2021).
- Trampler, T.; Mąkosa, K.; Girżda, A.; Bąkowski, J.; Dmyterko, E. Siedliskowe Podstawy Hodowli Lasu; PWRiL: Warsaw, Poland, 1990. (In Polish) [Google Scholar]
- CORINE land Cover-CLC. Available online: https://clc.gios.gov.pl (accessed on 16 March 2021).
- Braun−Blanquet, J. Pflanzensoziologie, Grundzüge der Vegetationskunde; Springer: Vienna, Austria, 1964. [Google Scholar]
- Thom, D.; Sommerfeld, A.; Sebald, J.; Hagge, J.; Müller, J.; Seidl, R. Effect of disturbance patterns and deadwood on the microclimate in European beech forests. Agric. For. Meteorol. 2020, 291, 108066. [Google Scholar] [CrossRef]
- Ewald, J.; Jehl, H.; Braun, L.; Lohberger, E. Die Vagetation des Nationalparks Bayerischer Wald als Ausdruck von Standort und Walddynarnik. Tuexenia 2011, 31, 9–38. [Google Scholar]
- Hermy, M.; Honnay, O.; Firbank, L.; Grashof−Bokdam, C.; Lawesson, J.E. An ecological comparison between ancient and other forest plant species of Europe, and the implications for forest conservation. Biol. Conserv. 1999, 91, 9–22. [Google Scholar] [CrossRef]
- Dzwonko, Z.; Loster, S. Wskaźnikowe gatunki roślin starych lasów i ich znaczenie dla ochrony przyrody i kartografii roślinności. IGiPZ PAN. Pr. Geogr. 2001, 178, 120–132. (In Polish) [Google Scholar]
- Matuszkiewicz, W. Przewodnik do Oznaczania Zbiorowisk Roślinnych Polski. [A Guide for the Identification of Polish Plant Communities]; Wydawnictwo Naukowe PWN: Warsaw, Poland, 2001. (In Polish) [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzenin Mitteleuropa. Scripta Geobot. 1992, 18, 1–248. [Google Scholar]
- Diekmann, M. Species indicator values as an important tool in applied plant ecology a review. Basic Appl. Ecol. 2003, 4, 493–506. [Google Scholar] [CrossRef]
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Rozkrut, D. Roczniki Statystyczne Leśnictwa 2020; GUS: Warsaw, Poland, 2020. (In Polish) [Google Scholar]
- MacDonald, D.; Crabtree, J.R.; Wiesinger, G.; Dax, T.; Stamou, N.; Fleury, P.; Lazpita, J.G.; Gibon, A. Agricultural abandonment in mountain areas of Europe: Environmental consequences and policy response. J. Environ. Manage. 2000, 59, 47–69. [Google Scholar] [CrossRef]
- Kozak, J.; Estreguil, C.; Troll, M. Forest cover changes in the northern Carpathians in the 20th century: A slow transition. J. Land Use Sci. 2007, 2, 127–146. [Google Scholar] [CrossRef]
- Kozak, J. Forest Cover Changes and Their Drivers in the Polish Carpathian Mountains since 1800. In Reforesting Landscapes Linking Pattern and Process; Nagendra, H., Southworth, J., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2010; pp. 253–273. [Google Scholar] [CrossRef]
- Kozak, J.; Szwagrzyk, M. Have there been forest transitions? Forest transition theory revisited in the context of the Modifiable Areal Unit Problem. R. Geogr. Soc. IBG 2016, 48, 504–512. [Google Scholar] [CrossRef]
- Bugno-Pogoda, A.; Durak, R.; Durak, T. Impact of Forest Management on the Temporal Dynamics of Herbaceous Plant Diversity in the Carpathian Beech Forests over 40 Years. Biology 2021, 10, 406. [Google Scholar] [CrossRef] [PubMed]
- Ujházy, K.; Hederová, L.; Máliš, F.; Ujházyová, M.; Bosela, M.; Čiliak, M. Overstorey dynamics controls plant diversity in age-class temperate forests. For. Ecol. Manag. 2017, 391, 96–105. [Google Scholar] [CrossRef]
- Schütz, J.P. Silvicultural tools to develop irregular and diverse forest structures. Forestry 2002, 75, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Raymond, P.; Bedard, S.; Roy, V.; Larouche, C.; Tremblay, S. The irregular shelterwood system: Review, classification, and potential application to forests affected by partial disturbances. J. For. 2009, 107, 405–413. [Google Scholar]
- Langenbruch, C.; Helfrich, M.; Flessa, H. Effects of beech (Fagus sylvatica), ash (Fraxinus excelsior) and lime (Tilia spec.) on soil chemical properties in a mixed deciduous forest. Plant Soil. 2012, 352, 389–403. [Google Scholar] [CrossRef] [Green Version]
- Facelli, J.M.; Pickett, S.T.A. Plant litter: Its dynamics and effects on plant community structure. Bot. Rev. 1991, 57, 1–32. [Google Scholar] [CrossRef]
- Depauw, L.; Perring, M.P.; Landuyt, D.; Maes, S.L.; Blondeel, H.; De Lombaerde, E.; Brūmelis, G.; Brunet, J.; Closset-Kopp, D.; Czerepko, J.; et al. Light availability and land-use history drive biodiversity and functional changes in forest herb layer communities. J. Ecol. 2019, 108, 1411–1425. [Google Scholar] [CrossRef]
- Bergstedt, J.; Hagner, M.; Milberg, P. Effects on vegetation composition of a modified forest harvesting and propagation method compared with clear-cutting, scarification and planting. Appl. Veg. Sci. 2008, 11, 159–168. [Google Scholar] [CrossRef]
- Durak, T.; Bugno-Pogoda, A.; Durak, R. Application of forest inventories to assess the forest developmental stages on plots dedicated to long-term vegetation studies. For. Ecol. Manag. 2021, 489, 119041. [Google Scholar] [CrossRef]
- Keenan, R.J. Climate change impacts and adaptation in forest management: A review. Ann. For. Sci. 2015, 72, 145–167. [Google Scholar] [CrossRef] [Green Version]
- Kovács, B.; Tinya, F.; Ódor, P. Stand structural drivers of microclimate in mature temperate mixed forests. Agric. For. Meteorol. 2017, 234–235, 11–21. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Year | ||
|---|---|---|---|
| 1970s | 2000s | 2010s | |
| % | |||
| Pinus sylvestris L. | 21.7 | 22.0 | 20.9 |
| Larix decidua Mill. | 1.0 | 2.5 | 2.6 |
| Picea abies (L.) H. Karst. | 2.4 | 2.9 | 2.7 |
| Abies alba Mill. | 28.2 | 27.0 | 30.9 |
| Fagus sylvatica L. | 27.6 | 31.4 | 34.1 |
| Quercus robur L. | 1.4 | 1.6 | 1.1 |
| Acer pseudoplatanus L. | 1.0 | 4.1 | 3.0 |
| Fraxinus excelsior L. | 0.1 | 0.8 | 0.3 |
| Carpinus betulus L. | 3.2 | 2.0 | 1.6 |
| Betula pendula Roth | 4.3 | 1.7 | 0.8 |
| Alnus glutinosa (L.) Gaertn. | 0.2 | 0.2 | 0.2 |
| Alnus incana (L.) Moench | 8.5 | 3.6 | 1.6 |
| Populus tremula L. | 0.2 | 0.1 | 0.1 |
| Salix sp. | 0.0 | 0.2 | 0.0 |
| Species | Year | ||
|---|---|---|---|
| 1970s | 2000s | 2010s | |
| % | |||
| Fagus sylvatica | 47.43 | 54.04 | 54.85 |
| Abies alba | 10.54 | 10.43 | 14.08 |
| Acer pseudoplatanus | 7.11 | 8.58 | 7.33 |
| Fraxinus excelsior | 1.49 | 2.74 | 0.06 |
| Carpinus betulus | 0.19 | 0.28 | 0.56 |
| Tilia cordata Mill. | 0.01 | 0.00 | 0.22 |
| Acer platanoides L. | 0.00 | 0.28 | 0.72 |
| Pinus sylvestris | 0.03 | 0.00 | 0.00 |
| Larix decidua | 0.01 | 0.00 | 0.00 |
| Populus tremula | 0.02 | 0.00 | 0.00 |
| Quercus robur | 0.02 | 0.01 | 0.00 |
| Ulmus glabra Huds. | 0.28 | 0.65 | 0.00 |
| Variables | Unit | Test Score | Mean (±SE) Values | ||
|---|---|---|---|---|---|
| F, ×Chi2 | 1970s | 2000s | 2010s | ||
| Total_Cov | % | ×0.96 | 88.0 (±0.97) ns | 86.0 (±0.96) ns | 85.5 (±1.74) ns |
| Herb_Cov | % | 3.8 * | 81.9 (±1.86) a | 80.7 (±1.90) a | 75.2 (±2.24) b |
| Number_Herb | - | 36.9 *** | 24.5 (±0.84) a | 26.9 (±0.95) a | 18.4 (±0.71) b |
| Number_Seedl | - | ×59.0 *** | 5.5 (±0.27) a | 6.0 (±0.23) a | 3.0 (±0.17) b |
| Number_Ancient | - | ×52.1 *** | 18.7 (±0.69) a | 20.0 (±0.80) a | 12.3 (±0.49) b |
| S_beech_forest | - | 7.0 *** | 3.9 (0.2) a | 3.7 (0.2) a | 3.1 (0.2) b |
| Fast species (ANE, END, EPI) | - | ×44.5 *** | 7.9 (±0.37) a | 7.7 (±0.36) a | 4.4 (±0.22) b |
| Slow species (MYR, HYD, BAR, AUT) | - | ×46.6 *** | 4.7 (±0.24 ) a | 6.0 (±0.30) b | 3.0 (±0.22) c |
| EIVs L | - | 3.4 * | 3.8 (±0.04) a | 3.8 (±0.03) a | 4.0 (±0.05) b |
| EIVs T | - | 2.3 | 5.0 (±0.01) ns | 5.0 (±0.02) ns | 5.0 (±0.02) ns |
| EIVs F | - | 4.1 * | 5.6 (±0.02) ab | 5.6 (±0.02) a | 5.5 (±0.03) b |
| EIVs R | - | ×10.2 ** | 6.5 (±0.04) a | 6.3 (±0.06) b | 6.4 (±0.07) b |
| EIVs N | - | ×3.4 | 6.2 (±0.04) ns | 6.2 (±0.06) ns | 6.2 (±0.06) ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bugno-Pogoda, A.; Durak, T. Climate and Management Factors Underlying Changes in Beech Forest Herbaceous Layer Plant Communities in the Polish Eastern Carpathians. Forests 2021, 12, 1446. https://doi.org/10.3390/f12111446
Bugno-Pogoda A, Durak T. Climate and Management Factors Underlying Changes in Beech Forest Herbaceous Layer Plant Communities in the Polish Eastern Carpathians. Forests. 2021; 12(11):1446. https://doi.org/10.3390/f12111446
Chicago/Turabian StyleBugno-Pogoda, Anna, and Tomasz Durak. 2021. "Climate and Management Factors Underlying Changes in Beech Forest Herbaceous Layer Plant Communities in the Polish Eastern Carpathians" Forests 12, no. 11: 1446. https://doi.org/10.3390/f12111446
APA StyleBugno-Pogoda, A., & Durak, T. (2021). Climate and Management Factors Underlying Changes in Beech Forest Herbaceous Layer Plant Communities in the Polish Eastern Carpathians. Forests, 12(11), 1446. https://doi.org/10.3390/f12111446