
Climate Signals for Growth Variations of F. sylvatica, P. abies, and P. sylvestris in Southeast Germany over the Past 50 Years



Abstract
:1. Introduction
2. Materials and Methods
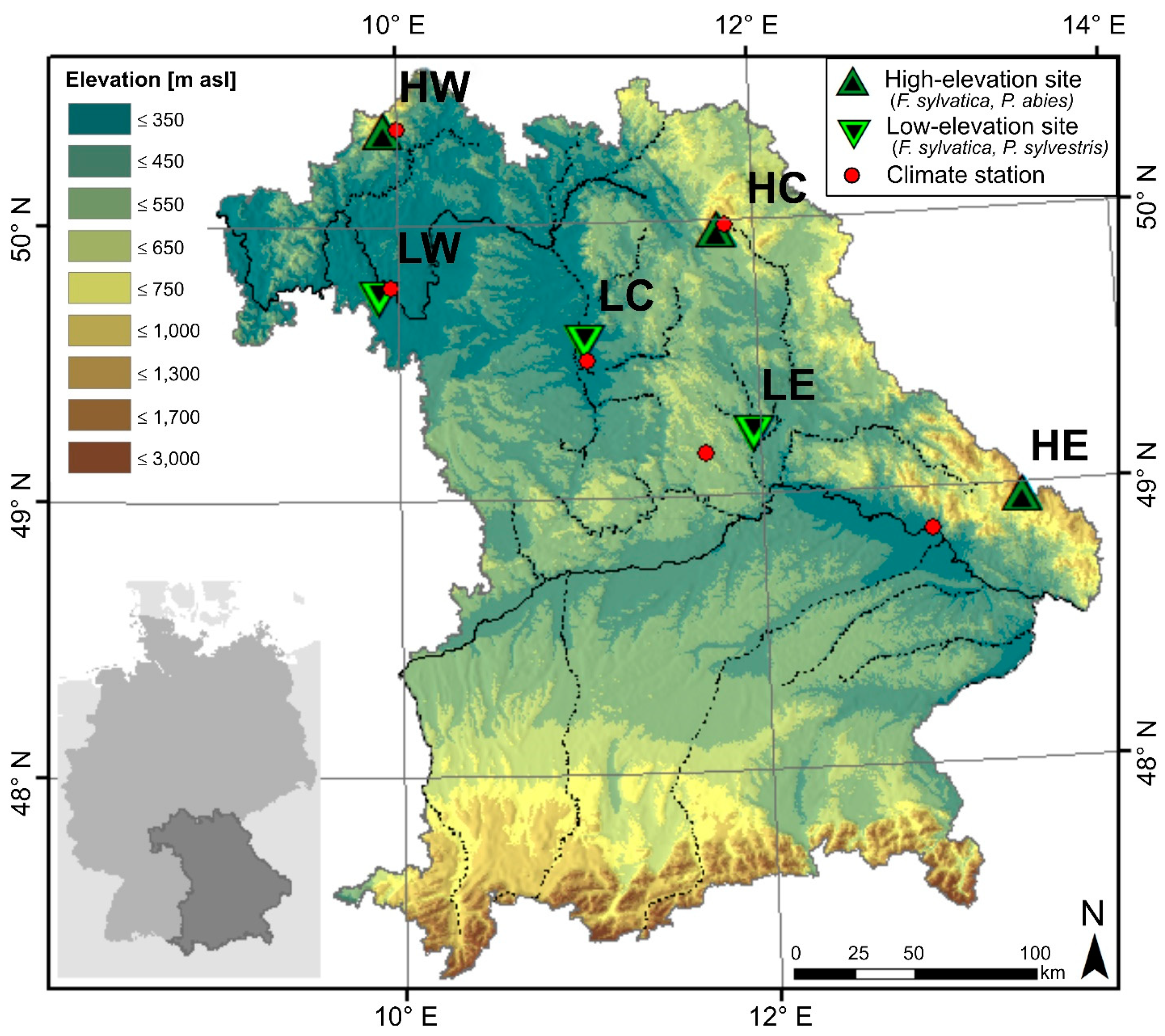
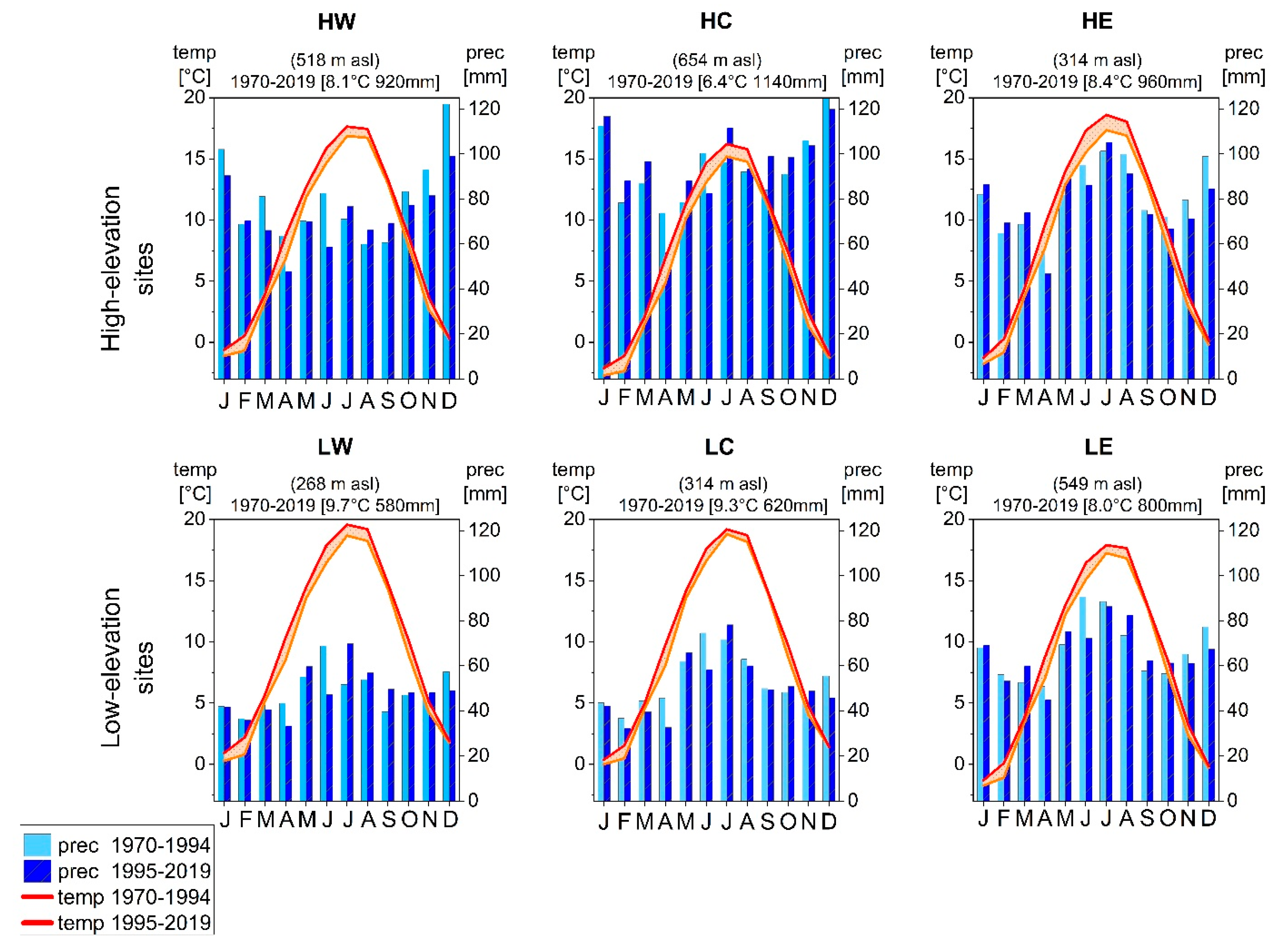
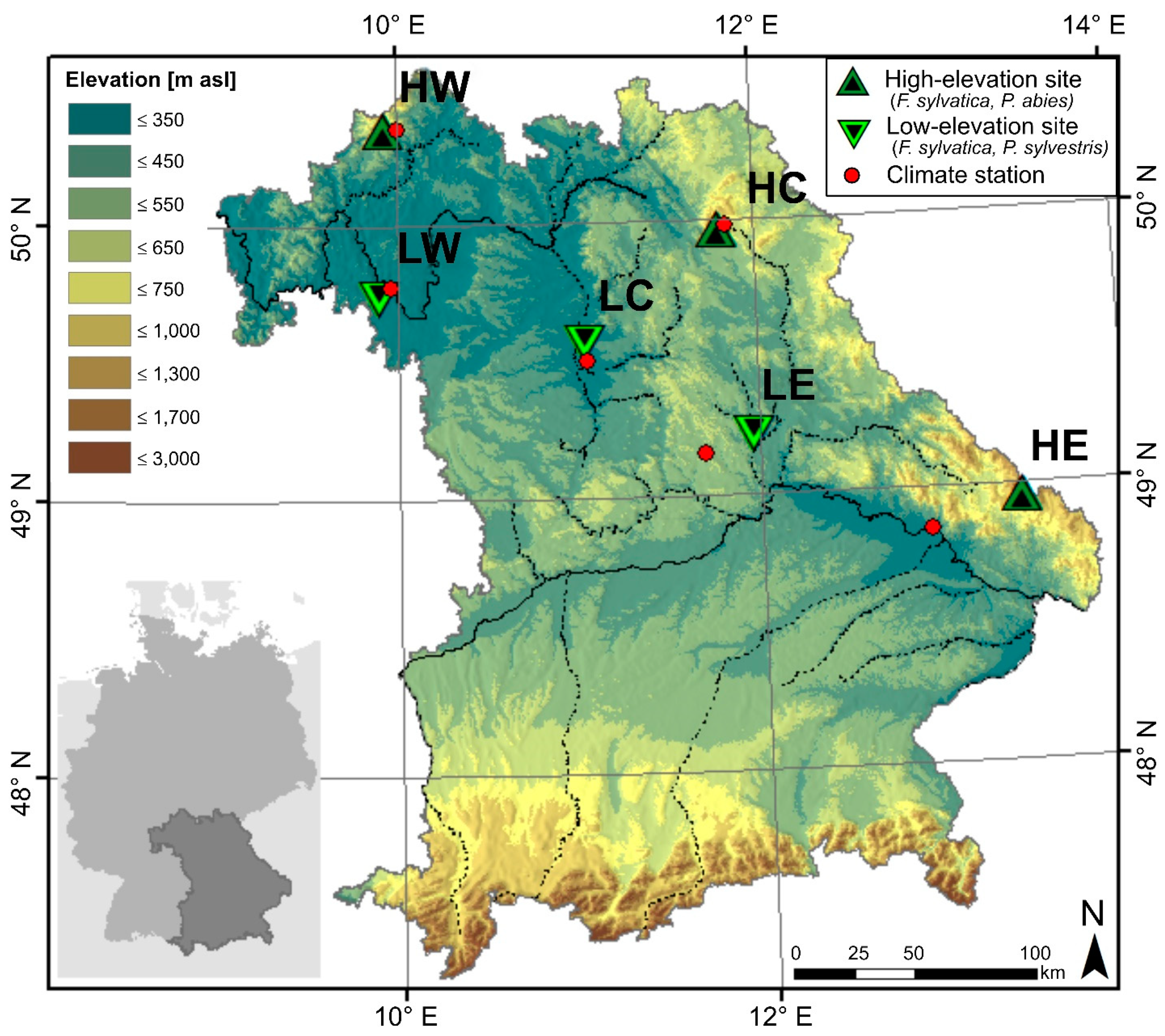
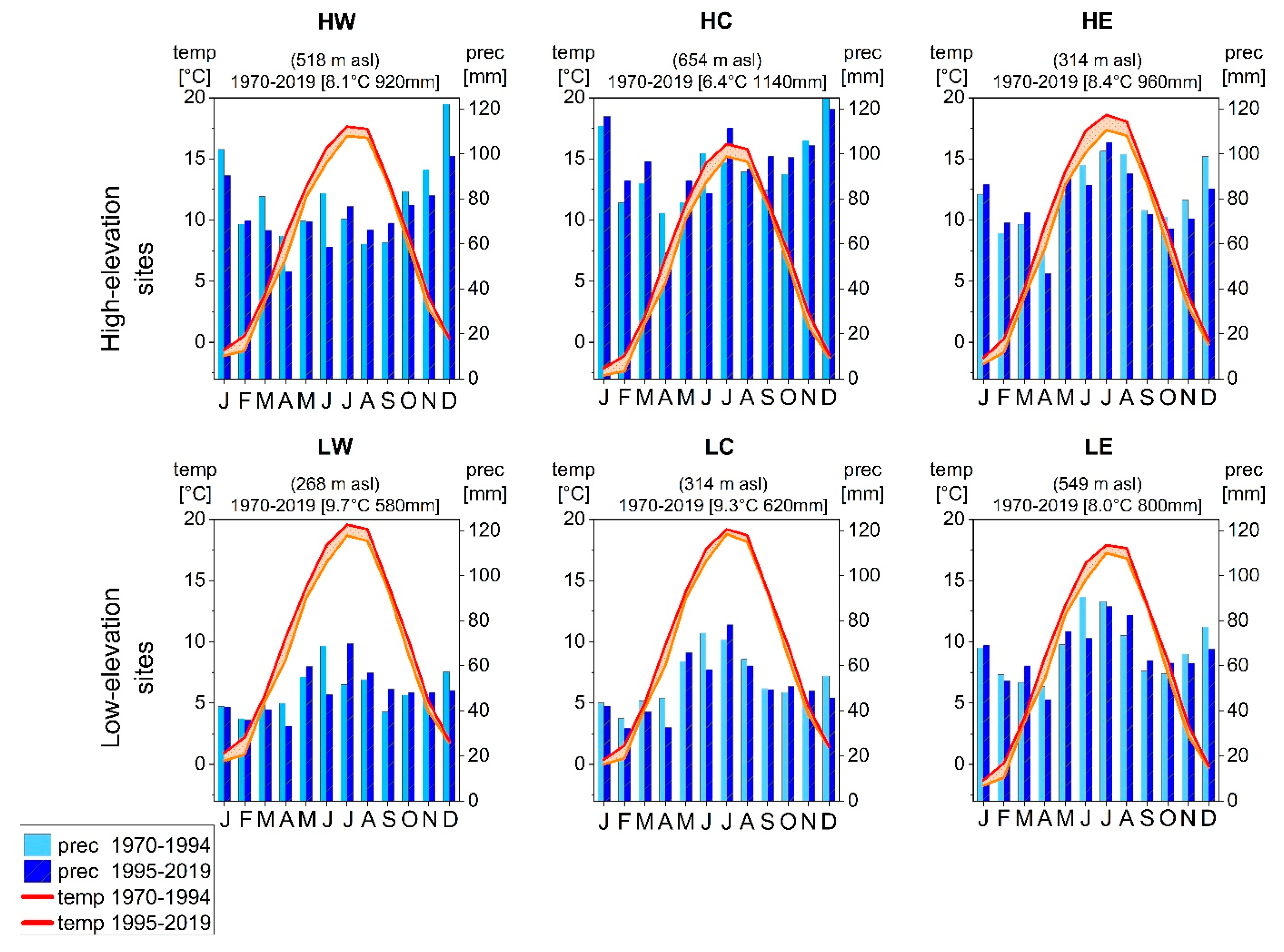
2.1. Study Area and Sample Collection
2.2. Statistical Analysis
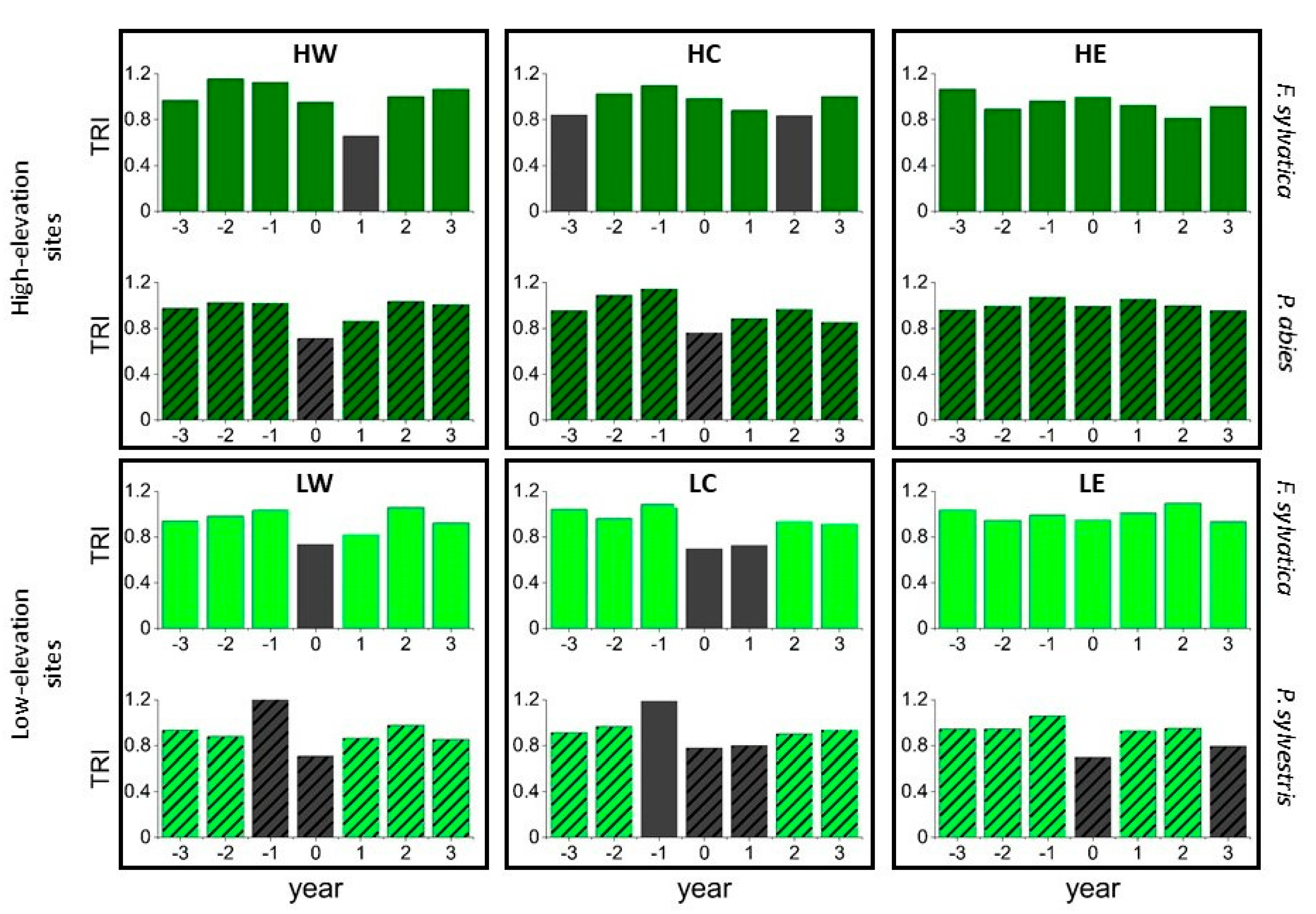
2.3. Pointer Year Analysis and Superposed Epoch Analysis
3. Results
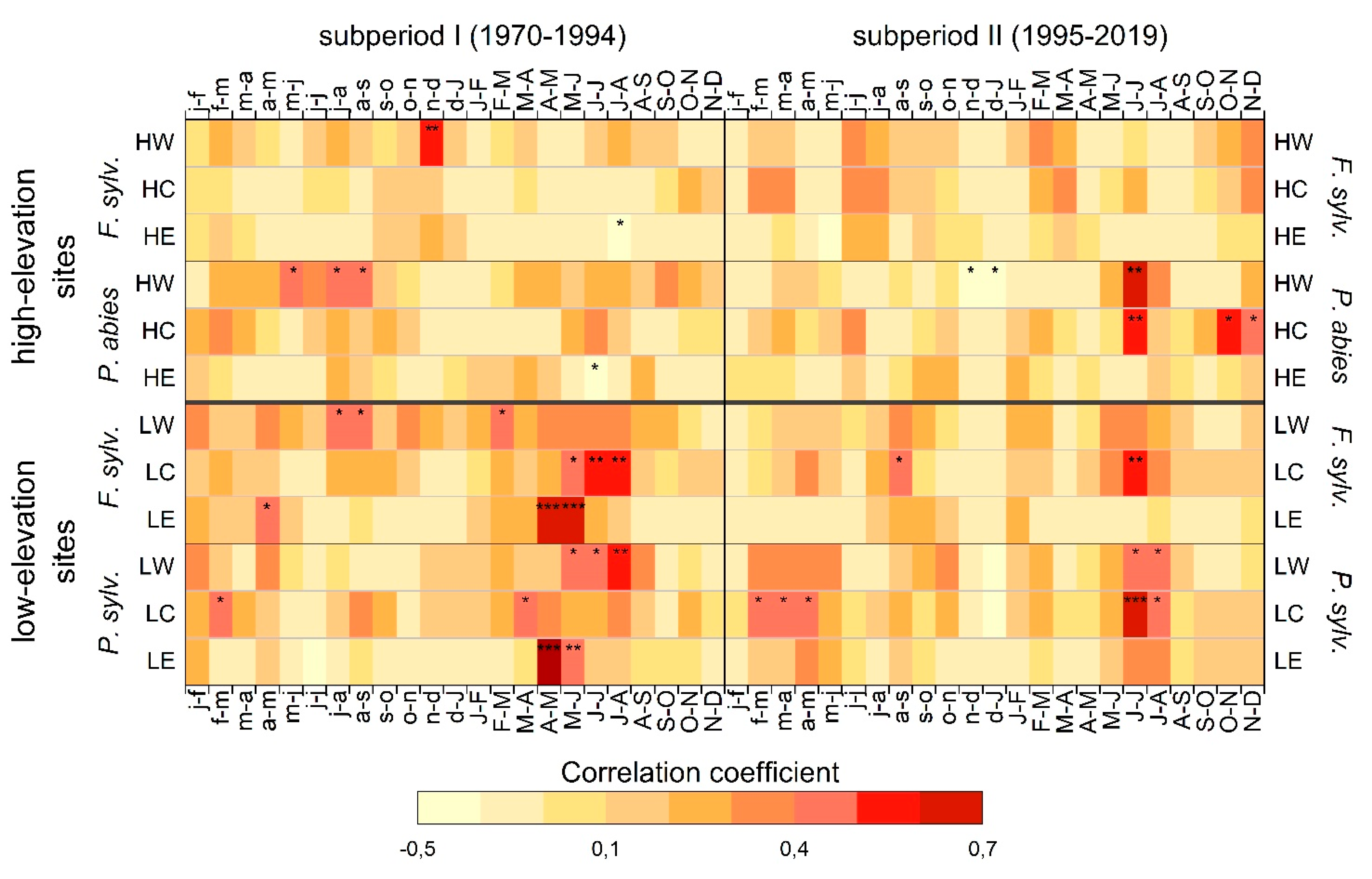
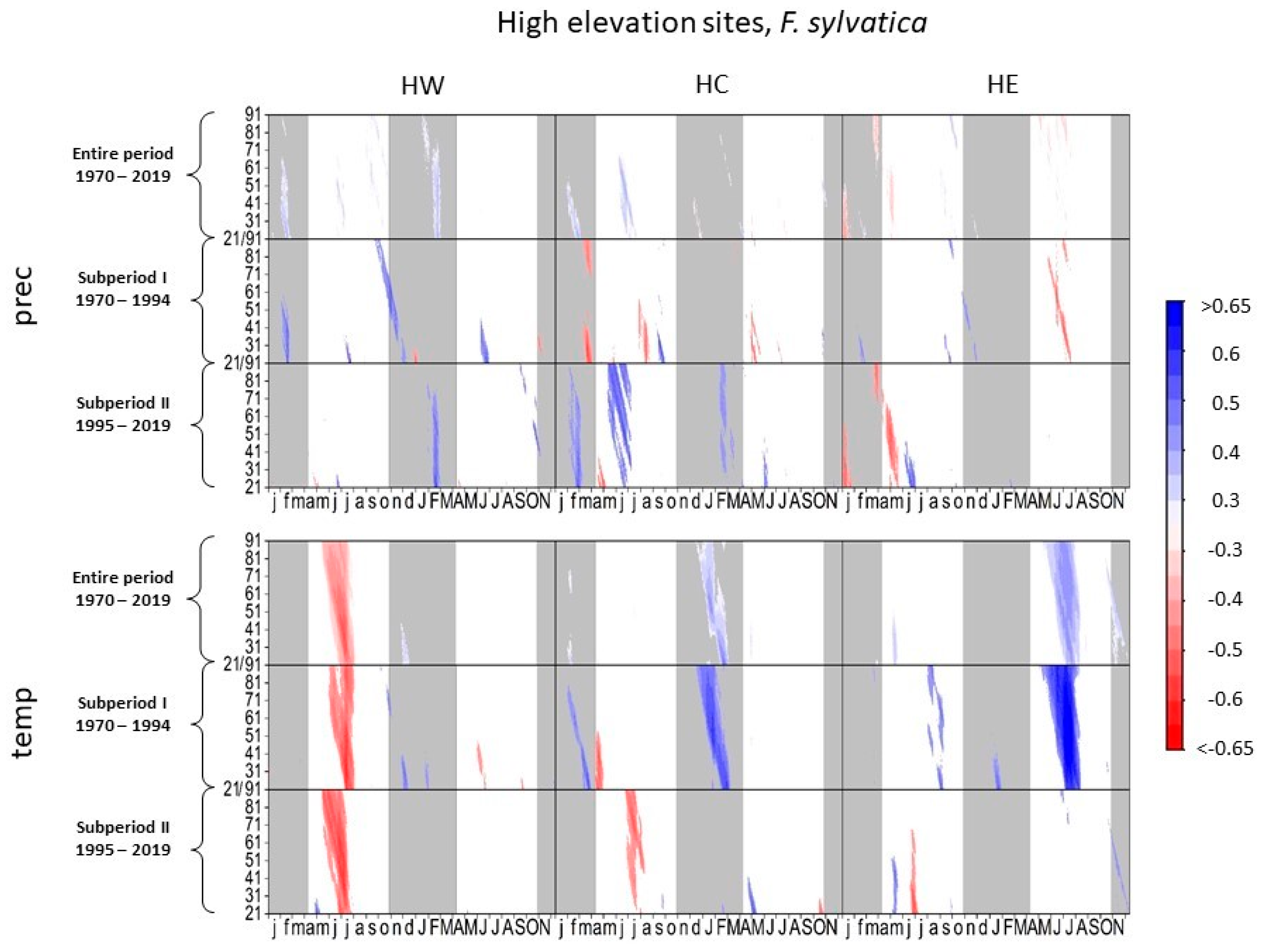
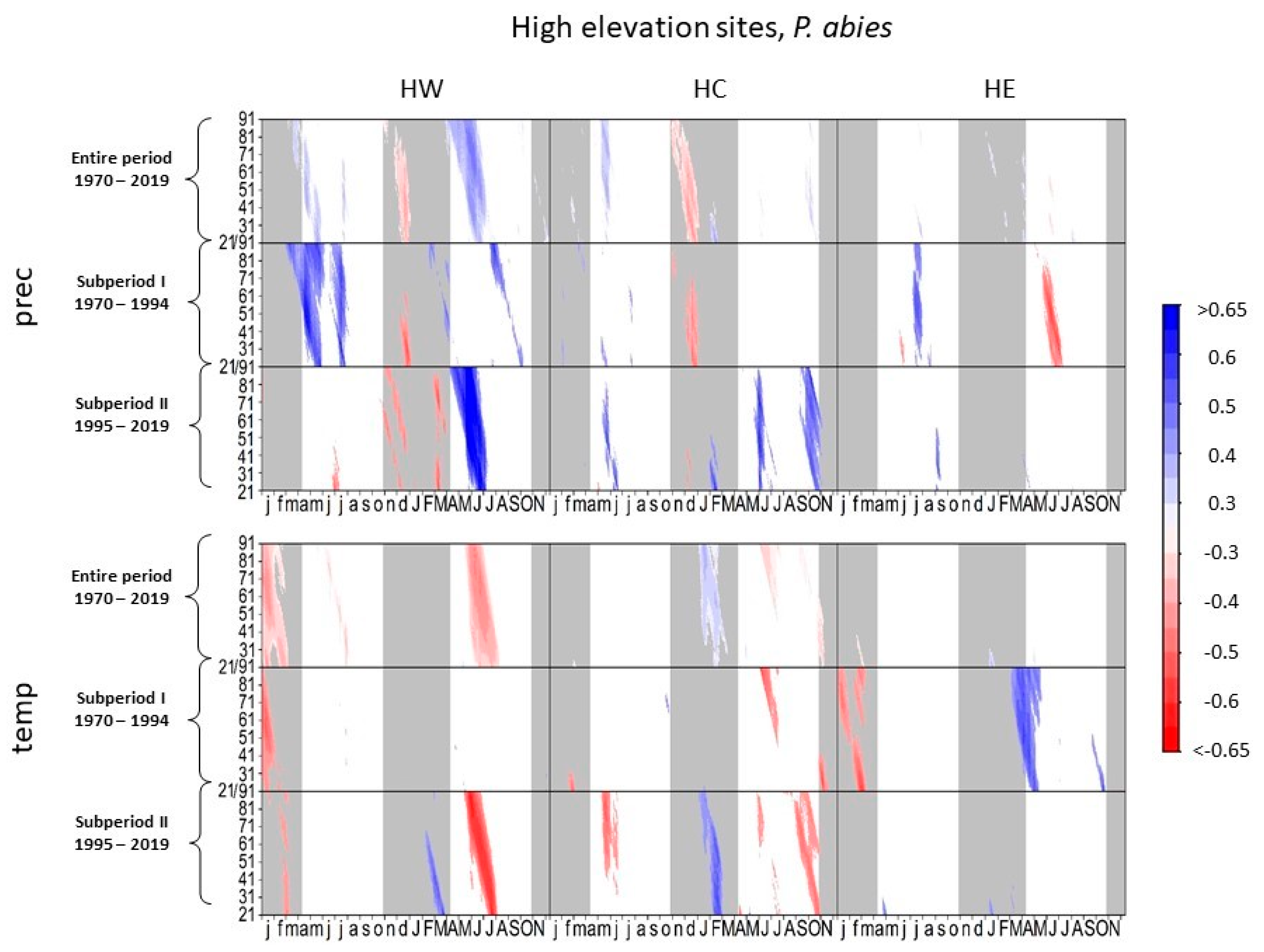
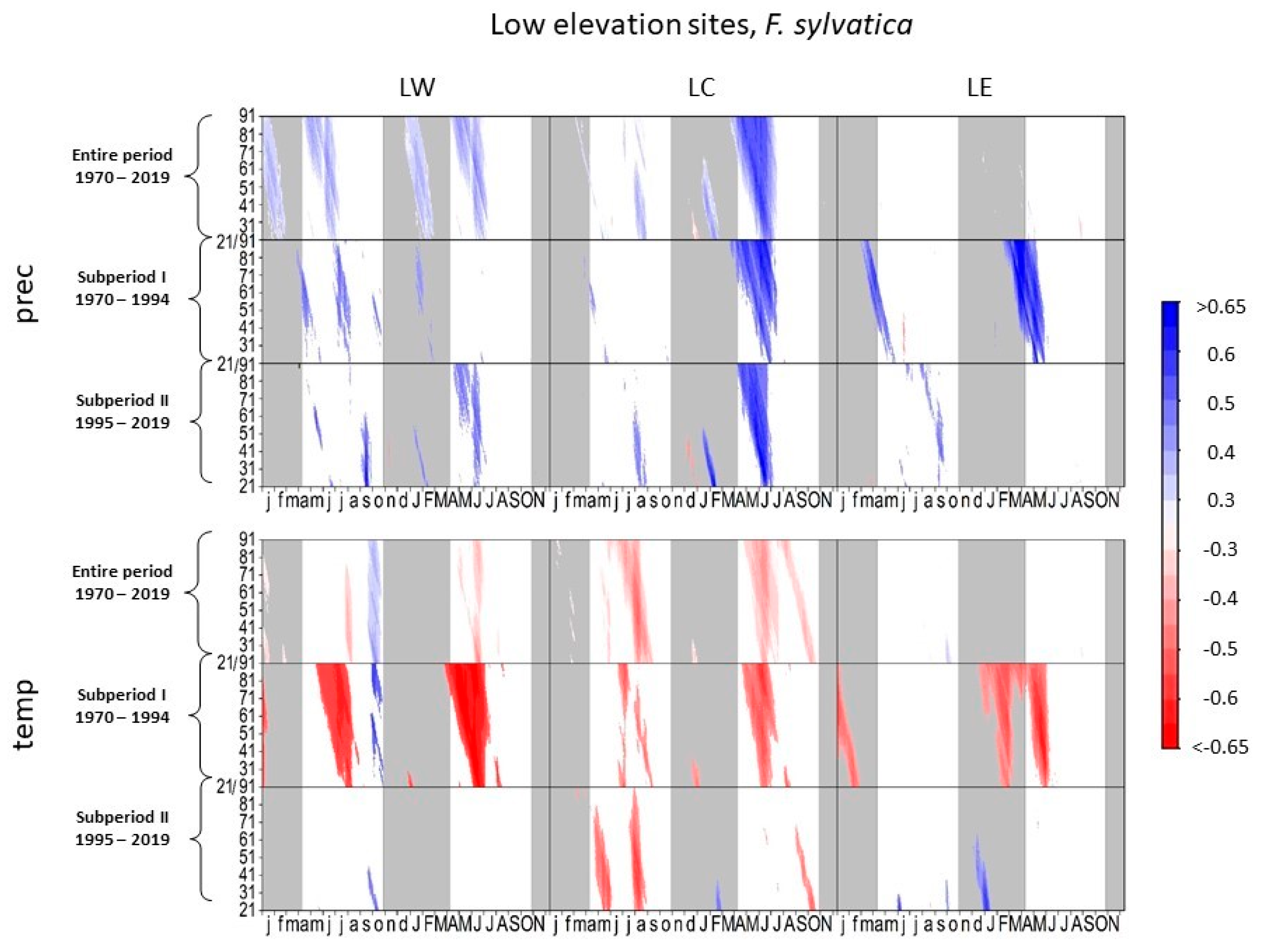
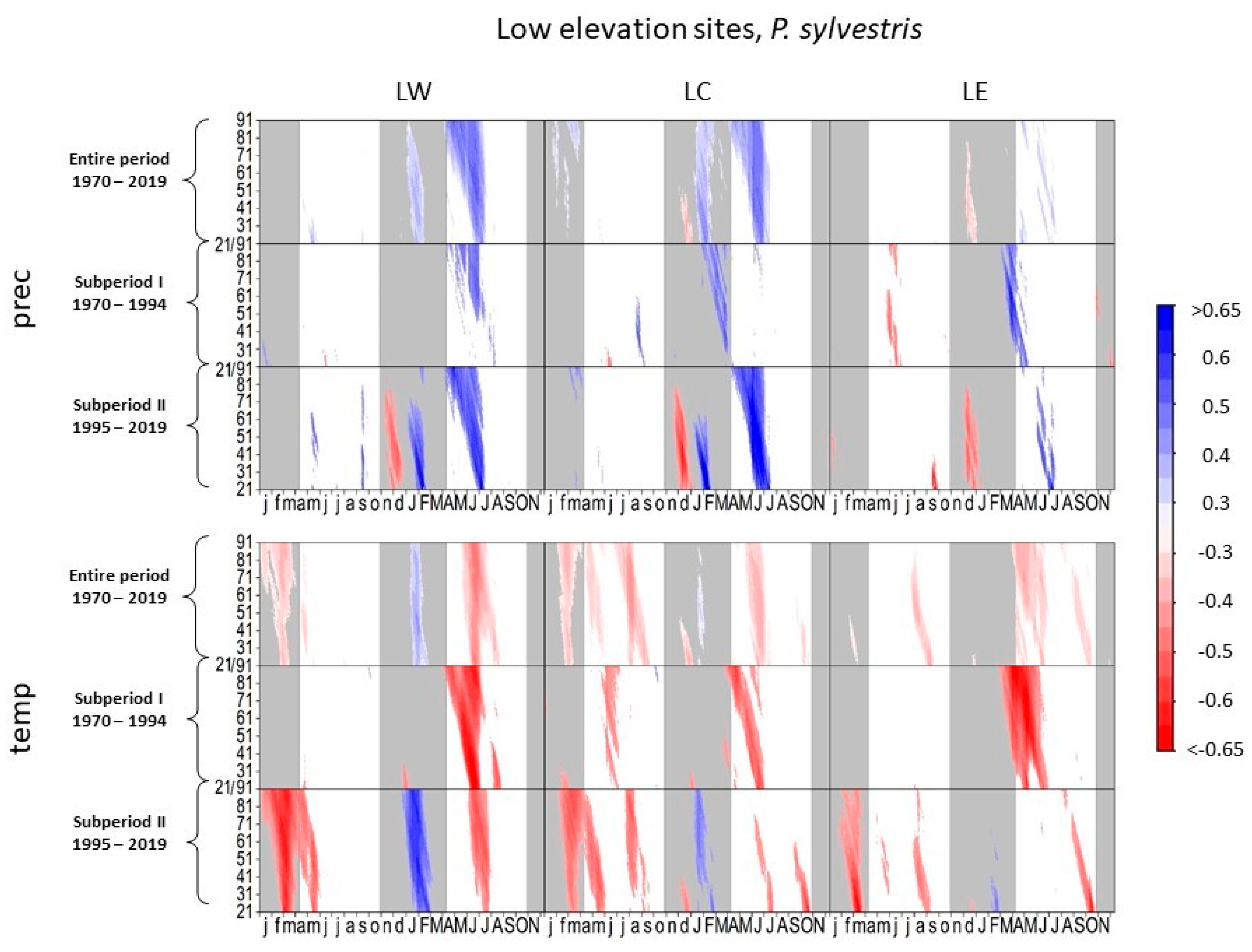
3.1. Climate–Growth Relationships
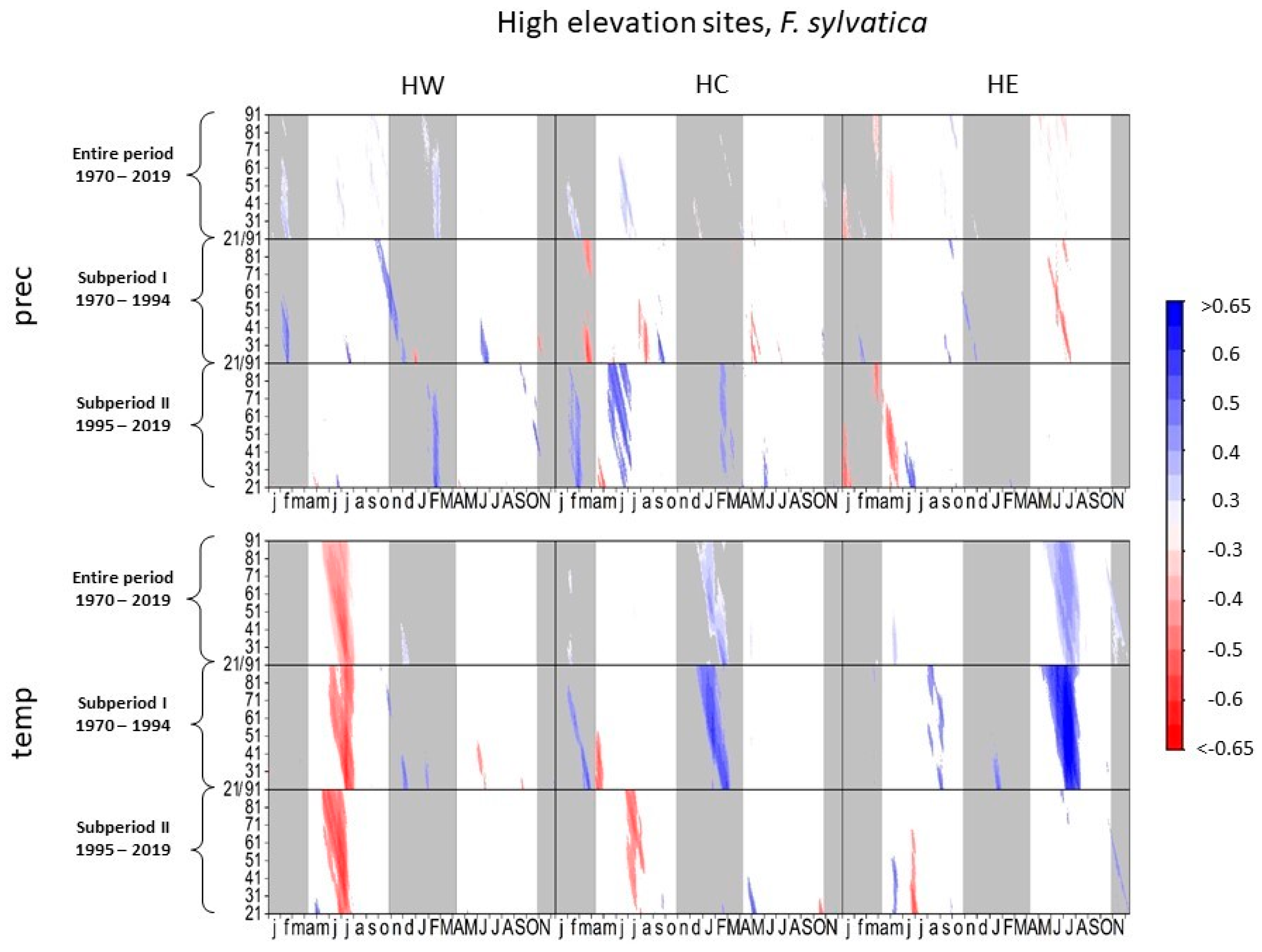
3.1.1. European Beech at High-Elevation Sites
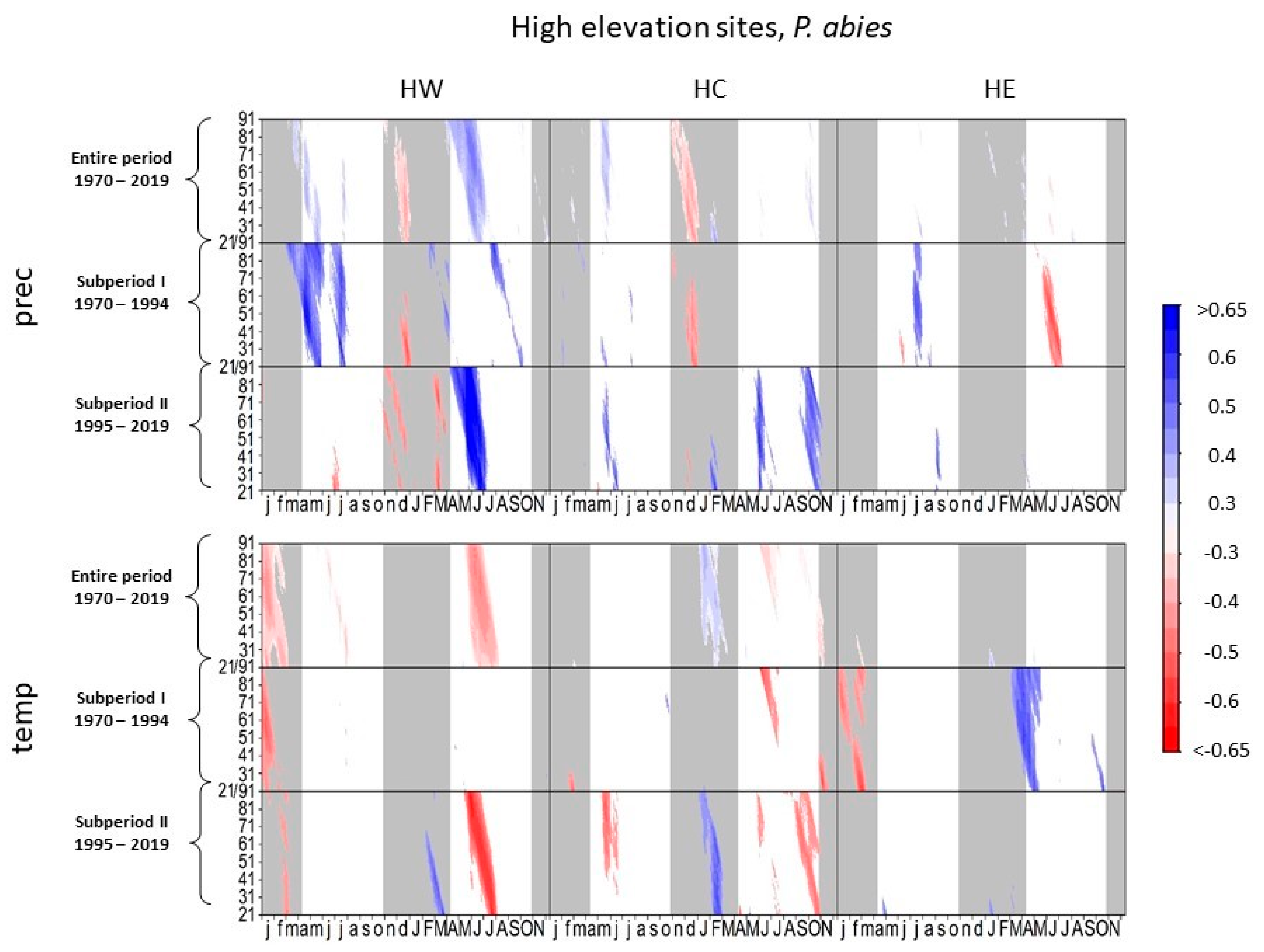
3.1.2. Norway Spruce at High-Elevation Sites
3.1.3. European Beech at Low-Elevation Sites
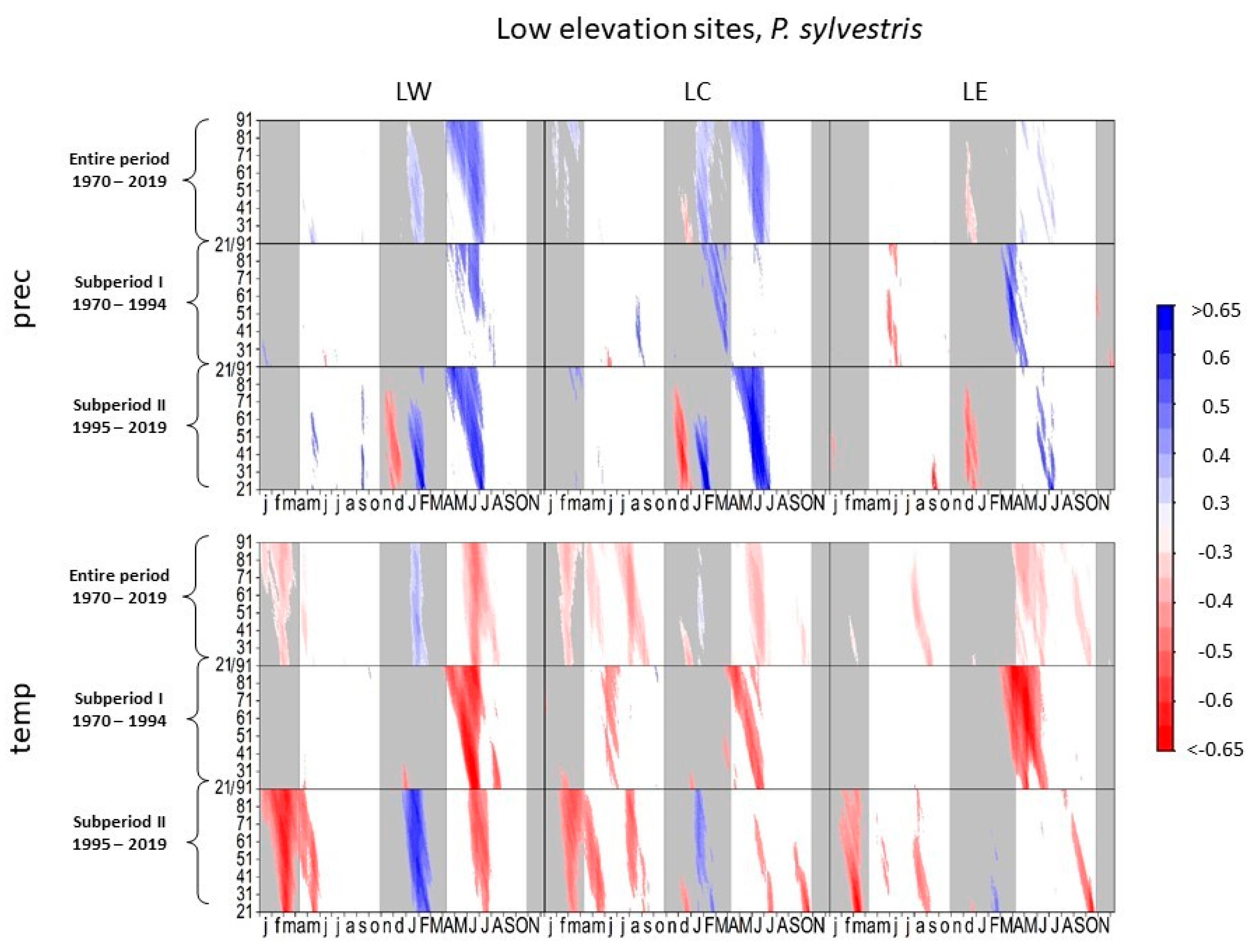
3.1.4. Scots Pine at Low-Elevation Sites
3.2. Tree Responses to 2-Month SPEI
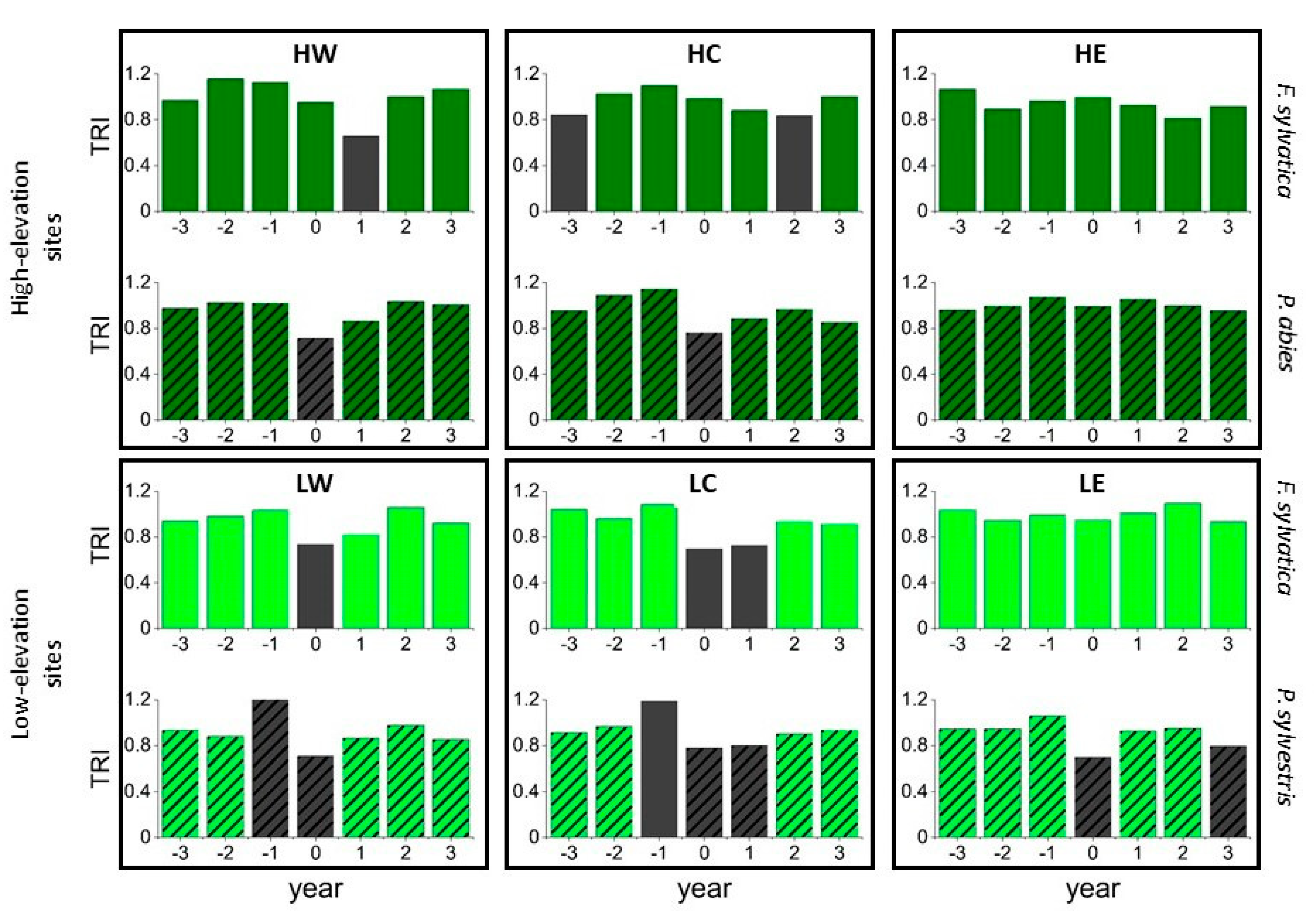
3.3. Pointer Year Analysis and Superposed Epoch Analysis (SEA)
4. Discussion
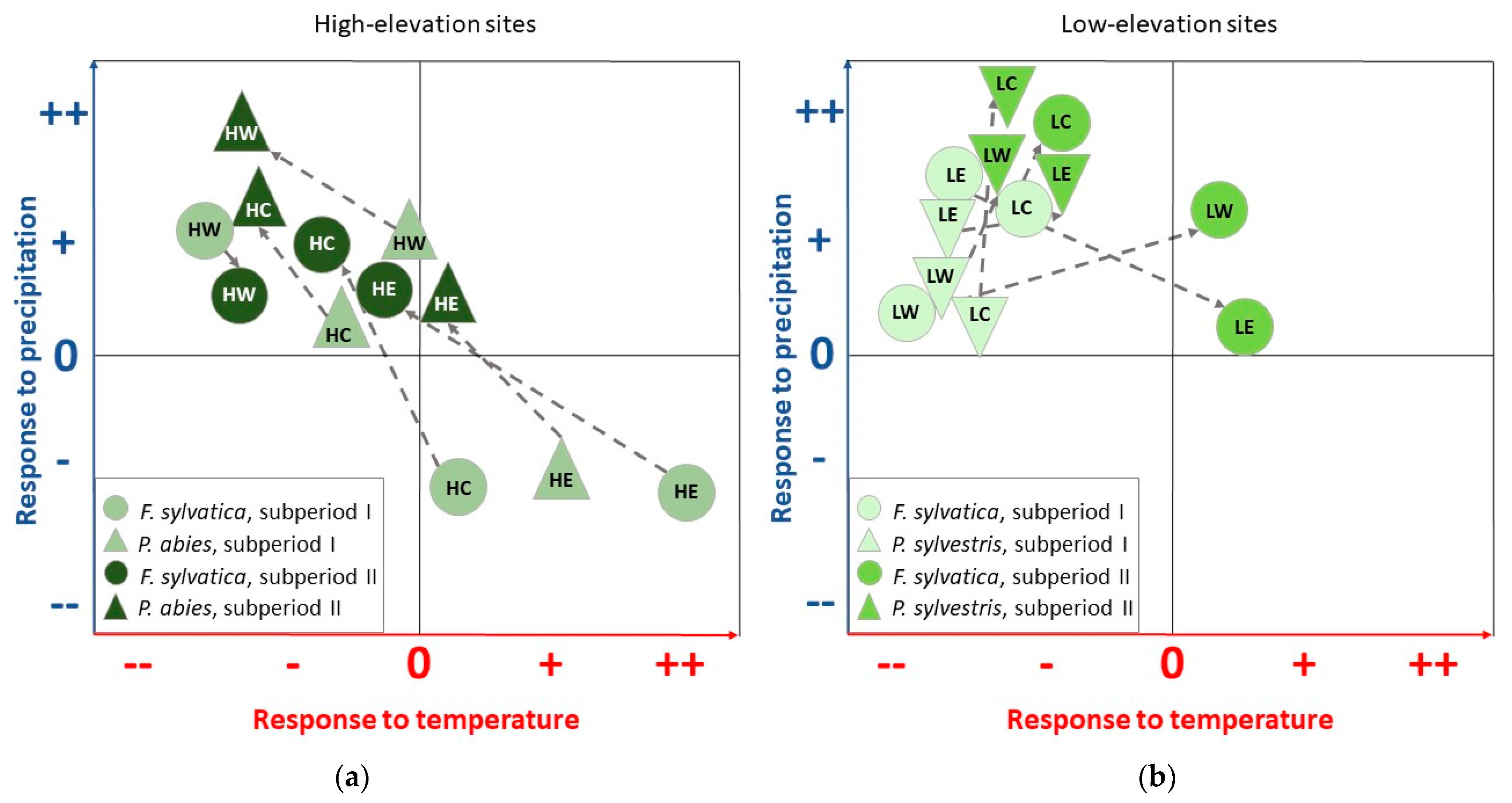
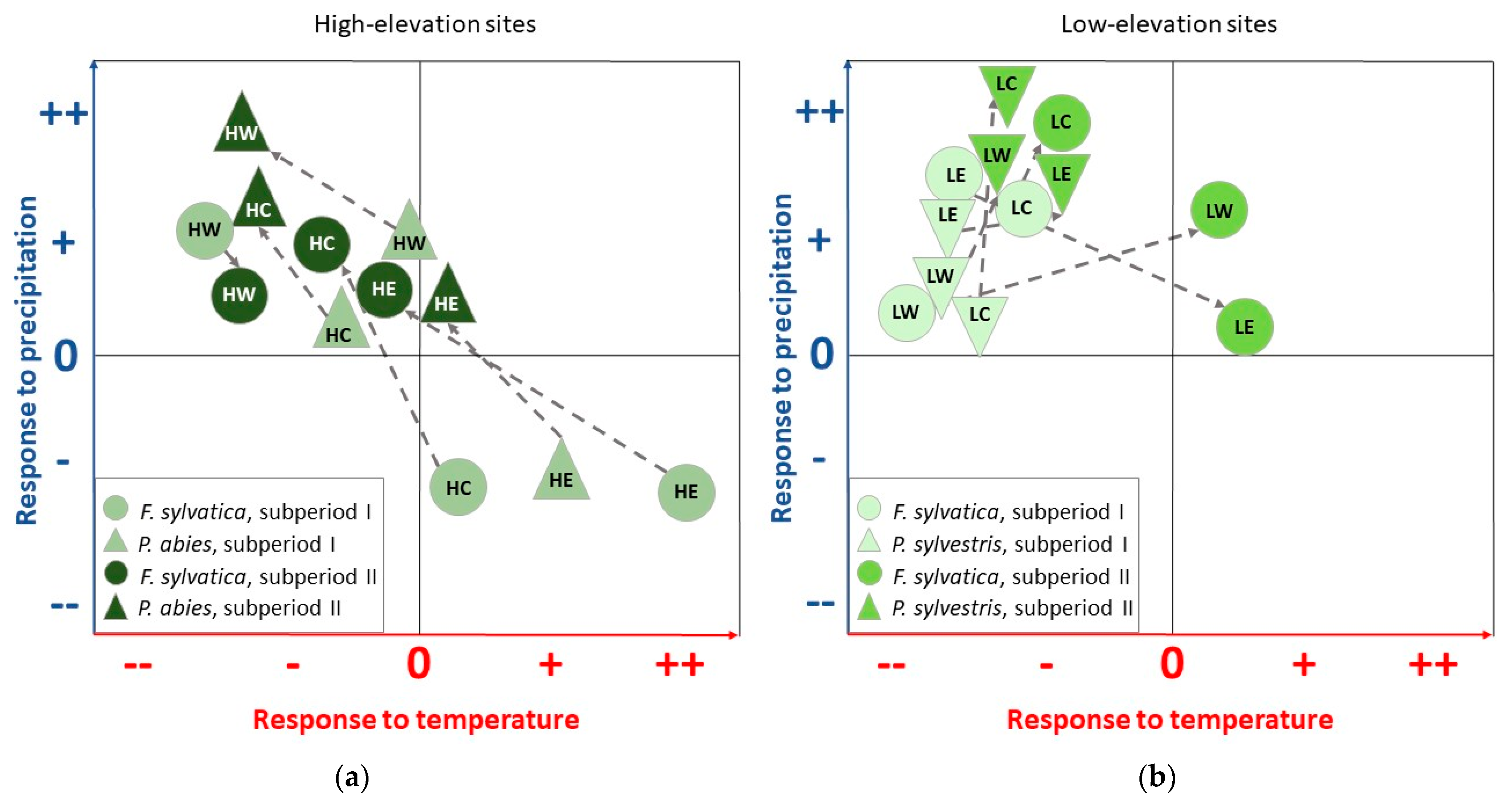
4.1. Climate–Growth Relationships
4.1.1. European Beech and Norway Spruce at High-Elevation Sites
4.1.2. European Beech and Scots Pine at Low-Elevation Sites
4.2. Pointer Year Analysis and Superposed Epoch Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kosanic, A.; Kavcic, I.; van Kleunen, M.; Harrison, S. Climate change and climate change velocity analysis across Germany. Sci. Rep. 2019, 9, 2196. [Google Scholar] [CrossRef]
- Christensen, J.H.; Hewitson, B.; Busuioc, A.; Chen, A.; Gao, X.; Held, I.; Jones, R.; Kolli, R.K.; Kwon, W.-T.; Laprise, R.; et al. Regional Climate Projections. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007. [Google Scholar]
- Buras, A.; Rammig, A.; Zang, C.S. Quantifying impacts of the 2018 drought on European ecosystems in comparison to 2003. Biogeosciences 2020, 17, 1655–1672. [Google Scholar] [CrossRef] [Green Version]
- Schuldt, B.; Buras, A.; Arend, M.; Vitasse, Y.; Beierkuhnlein, C.; Damm, A.; Gharun, M.; Grams, T.E.E.; Hauck, M.; Hajek, P.; et al. A first assessment of the impact of the extreme 2018 summer drought on Central European forests. Basic Appl. Ecol. 2020, 45, 86–103. [Google Scholar] [CrossRef]
- Bayerisches Staatsministerium für Ernährung; Landwirtschaft und Forsten. Ergebnisse der Waldzustandserhebung 2019; STMELF: München, Germany, 2019; Volume 2019. [Google Scholar]
- Friedrichs, D.A.; Trouet, V.; Büntgen, U.; Frank, D.C.; Esper, J.; Neuwirth, B.; Löffler, J. Species-specific climate sensitivity of tree growth in Central-West Germany. Trees 2009, 23, 729–739. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early warnings of tree dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef] [Green Version]
- Buras, A.; Schunk, C.; Zeiträg, C.; Herrmann, C.; Kaiser, L.; Lemme, H.; Straub, C.; Taeger, S.; Gößwein, S.; Klemmt, H.-J.; et al. Are Scots pine forest edges particularly prone to drought-induced mortality? Environ. Res. Lett. 2018, 13, 25001. [Google Scholar] [CrossRef]
- Bhuyan, U.; Zang, C.; Menzel, A. Different responses of multispecies tree ring growth to various drought indices across Europe. Dendrochronologia 2017, 44, 1–8. [Google Scholar] [CrossRef]
- Baumbach, L.; Niamir, A.; Hickler, T.; Yousefpour, R. Regional adaptation of European beech (Fagus sylvatica) to drought in Central European conditions considering environmental suitability and economic implications. Reg. Environ. Chang. 2019, 19, 1159–1174. [Google Scholar] [CrossRef]
- Zang, C.; Rothe, A.; Weis, W.; Pretzsch, H. Tree Suitability under climate change conditions: Susceptibility of major forest tree species from tree-ring widths. Allg. Forst Jagdztg. 2011, 182, 98–112. [Google Scholar]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.-J.; Lasch, P.; Eggers, J.; van der Maaten-Theunissen, M.; et al. Climate change and European forests: What do we know, what are the uncertainties, and what are the implications for forest management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartl-Meier, C.; Zang, C.; Büntgen, U.; Esper, J.; Rothe, A.; Göttlein, A.; Dirnböck, T.; Treydte, K. Uniform climate sensitivity in tree-ring stable isotopes across species and sites in a mid-latitude temperate forest. Tree Physiol. 2015, 35, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Kraus, C.; Zang, C.; Menzel, A. Elevational response in leaf and xylem phenology reveals different prolongation of growing period of common beech and Norway spruce under warming conditions in the Bavarian Alps. Eur. J. Forest Res. 2016, 135, 1011–1023. [Google Scholar] [CrossRef]
- Vitali, V.; Büntgen, U.; Bauhus, J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob. Chang. Biol. 2017, 23, 5108–5119. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Kremer, A.; Nagy, L.; Újvári-Jármay, É.; Ducousso, A.; Kóczán-Horváth, A.; Hansen, J.K.; Mátyás, C. Common garden comparisons confirm inherited differences in sensitivity to climate change between forest tree species. PeerJ 2019, 7, e6213. [Google Scholar] [CrossRef] [PubMed]
- Spathelf, P.; van der Maaten, E.; van der Maaten-Theunissen, M.; Campioli, M.; Dobrowolska, D. Climate change impacts in European forests: The expert views of local observers. Ann. For. Sci. 2014, 71, 131–137. [Google Scholar] [CrossRef]
- Neuwirth, B.; Schweingruber, F.H.; Winiger, M. Spatial patterns of central European pointer years from 1901 to 1971. Dendrochronologia 2007, 24, 79–89. [Google Scholar] [CrossRef]
- McCarty, J. Ecological Consequences of Recent Climate Change. Conserv. Biol. 2001, 15, 320–331. [Google Scholar] [CrossRef]
- Walentowski, H.; Gulder, H.-J.; Kölling, C.; Ewald, J.; Türk, W. Die regionale natürliche Waldzusammensetzung Bayerns: Berichte aus der bayerischen Landesanstalt für Wald und Forstwirtschaft. LWF-Bericht 2001, 32, 10–14. [Google Scholar]
- Knutzen, F.; Dulamsuren, C.; Meier, I.C.; Leuschner, C. Recent Climate Warming-Related Growth Decline Impairs European Beech in the Center of Its Distribution Range. Ecosystems 2017, 20, 1494–1511. [Google Scholar] [CrossRef]
- Dulamsuren, C.; Hauck, M.; Kopp, G.; Ruff, M.; Leuschner, C. European beech responds to climate change with growth decline at lower, and growth increase at higher elevations in the center of its distribution range (SW Germany). Trees 2017, 31, 673–686. [Google Scholar] [CrossRef]
- Bauwe, A.; Koch, M.; Kallweit, R.; Konopatzky, A.; Strohbach, B.; Lennartz, B. Tree-ring growth response of Scots pine (Pinus sylvestris L.) to climate and soil water availability in the lowlands of North-Eastern Germany. Balt. For. 2013, 212–225. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Jevšenak, J.; Levanič, T. dendroTools: R package for studying linear and nonlinear responses between tree-rings and daily environmental data. Dendrochronologia 2018, 48, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Beck, W.; Heinzig, P. A new tool to discovering realistic climate-growth relationships. FREIJ 2018, 2, 49–52. [Google Scholar] [CrossRef]
- DWD Climate Data Center. Station Observations for Germany; Version v19.3; Deutscher Wetterdienst: Offenbach, Germany, 2020. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press Inc. (London) LTD.: London, UK, 1976. [Google Scholar]
- Eckstein, D.; Bauch, J. Beitrag zur Rationalisierung eines dendrochronologischen Verfahrens und zur Analyse seiner Aussagesicherheit. Forstwiss. Cent. 1969, 88, 230–250. [Google Scholar] [CrossRef]
- Rinn, F. TSAP-Win -User Reference: Time Series Analysis and Presentation for Dendrochronology and Related Applications; Version 4.64 for Microsoft Windows; Rinntech-Metriwerk GmbH & Co. KG: Heidelberg, Germany, 2011. [Google Scholar]
- Grissino-Mayer, H.D.; Fritts, H.C. The International Tree-Ring Data Bank: An enhanced global database serving the global scientific community. Holocene 1997, 235–238. [Google Scholar] [CrossRef]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 1 July 2021).
- Efron, B.; Tibshirani, R. Bootstrap Methods for Standard Errors, Confidence Intervals, and Other Measures of Statistical Accuracy. Stat. Sci. 1986, 1, 54–77. [Google Scholar] [CrossRef]
- van der Maaten-Theunissen, M.; van der Maaten, E.; Bouriaud, O. pointRes: An R package to analyze pointer years and components of resilience. Dendrochronologia 2015, 35, 34–38. [Google Scholar] [CrossRef]
- Schweingruber, F.H.; Eckstein, D.; Serre-Bachet, F.; Bräker, O.U. Identification, presentation and interpretation of event years and pointer years in dendrochronology. Dendrochronologia 1990, 1990, 9–38. [Google Scholar]
- Bunn, A.G. Statistical and visual crossdating in R using the dplR library. Dendrochronologia 2010, 28, 251–258. [Google Scholar] [CrossRef]
- Dittmar, C.; Zech, W.; Elling, W. Growth variations of Common beech (Fagus sylvatica L.) under different climatic and environmental conditions in Europe—A dendroecological study. For. Ecol. Manag. 2003, 173, 63–78. [Google Scholar] [CrossRef]
- Graham, E.A.; Mulkey, S.S.; Kitajima, K.; Phillips, N.G.; Wright, S.J. Cloud cover limits net CO2 uptake and growth of a rainforest tree during tropical rainy seasons. Proc. Natl. Acad. Sci. USA 2003, 100, 572–576. [Google Scholar] [CrossRef] [Green Version]
- Di Filippo, A.; Biondi, F.; Čufar, K.; Luis, M.d.; Grabner, M.; Maugeri, M.; Presutti Saba, E.; Schirone, B.; Piovesan, G. Bioclimatology of beech (Fagus sylvatica L.) in the Eastern Alps: Spatial and altitudinal climatic signals identified through a tree-ring network. J. Biogeogr. 2007, 34, 1873–1892. [Google Scholar] [CrossRef]
- Rozas, V.; Camarero, J.J.; Sangüesa-Barreda, G.; Souto, M.; García-González, I. Summer drought and ENSO-related cloudiness distinctly drive Fagus sylvatica growth near the species rear-edge in northern Spain. Agric. For. Meteorol. 2015, 201, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Dulamsuren, C.; Klinge, M.; Degener, J.; Khishigjargal, M.; Chenlemuge, T.; Bat-Enerel, B.; Yeruult, Y.; Saindovdon, D.; Ganbaatar, K.; Tsogtbaatar, J.; et al. Carbon pool densities and a first estimate of the total carbon pool in the Mongolian forest-steppe. Glob. Chang. Biol. 2016, 22, 830–844. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Jevšenak, J.; Tychkov, I.; Gričar, J.; Levanič, T.; Tumajer, J.; Prislan, P.; Arnič, D.; Popkova, M.; Shishov, V.V. Growth-limiting factors and climate response variability in Norway spruce (Picea abies L.) along an elevation and precipitation gradients in Slovenia. Int. J. Biometeorol. 2021, 65, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Scharnweber, T.; Manthey, M.; Criegee, C.; Bauwe, A.; Schröder, C.; Wilmking, M. Drought matters—Declining precipitation influences growth of Fagus sylvatica L. and Quercus robur L. in north-eastern Germany. For. Ecol. Manag. 2011, 262, 947–961. [Google Scholar] [CrossRef]
- Dittmar, C.; Elling, W. Jahrringbreite von Fichte und Buche in Abhängigkeit von Witterung und Höhenlage: Radial growth of Norway spruce and Euripean beech in relation to weather and altitude. Forstw. Cbl. 1999, 118, 251–270. [Google Scholar] [CrossRef]
- Lebourgeois, F.; Bréda, N.; Ulrich, E.; Granier, A. Climate-tree-growth relationships of European beech (Fagus sylvatica L.) in the French Permanent Plot Network (RENECOFOR). Trees 2005, 19, 385–401. [Google Scholar] [CrossRef]
- Weber, P.; Bugmann, H.; Rigling, A. Radial growth responses to drought of Pinus sylvestris and Quercus pubescens in an inner-Alpine dry valley. J. Veg. Sci. 2007, 18, 777–792. [Google Scholar] [CrossRef]
- Muffler, L.; Weigel, R.; Hacket-Pain, A.J.; Klisz, M.; Maaten, E.; Wilmking, M.; Kreyling, J.; Maaten-Theunissen, M. Lowest drought sensitivity and decreasing growth synchrony towards the dry distribution margin of European beech. J. Biogeogr. 2020, 47, 1910–1921. [Google Scholar] [CrossRef]
- Cavin, L.; Jump, A.S. Highest drought sensitivity and lowest resistance to growth suppression are found in the range core of the tree Fagus sylvatica L. not the equatorial range edge. Glob. Chang. Biol. 2017, 23, 362–379. [Google Scholar] [CrossRef] [Green Version]
- Walentowski, H.; Falk, W.; Mette, T.; Kunz, J.; Bräuning, A.; Meinardus, C.; Zang, C.; Sutcliffe, L.M.E.; Leuschner, C. Assessing future suitability of tree species under climate change by multiple methods: A case study in southern Germany. Ann. For. Res. 2014, 60, 101–126. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, J.; Hauck, M.; Dulamsuren, C.; Leuschner, C. Climate Warming-Related Growth Decline Affects Fagus sylvatica, But Not Other Broad-Leaved Tree Species in Central European Mixed Forests. Ecosystems 2015, 18, 560–572. [Google Scholar] [CrossRef]
- Sykes, M.T.; Prentice, I.C. Climate change, tree species distributions and forest dynamics: A case study in the mixed conifer/northern hardwoods zone of northern Europe. Clim. Chang. 1996, 34, 161–177. [Google Scholar] [CrossRef]
- Oberhuber, W. Soil water availability and evaporative demand affect seasonal growth dynamics and use of stored water in co-occurring saplings and mature conifers under drought. Trees 2017, 31, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity—Evidence since the middle of the 20th century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef] [Green Version]
- Weemstra, M.; Eilmann, B.; Sass-Klaassen, U.G.W.; Sterck, F.J. Summer droughts limit tree growth across 10 temperate species on a productive forest site. For. Ecol. Manag. 2013, 306, 142–149. [Google Scholar] [CrossRef]
- Butt, N.; Bebber, D.P.; Riutta, T.; Crockatt, M.; Morecroft, M.D.; Malhi, Y. Relationships between tree growth and weather extremes: Spatial and interspecific comparisons in a temperate broadleaf forest. For. Ecol. Manag. 2014, 334, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Gazol, A.; Camarero, J.J.; Anderegg, W.R.L.; Vicente-Serrano, S.M. Impacts of droughts on the growth resilience of Northern Hemisphere forests. Glob. Ecol. Biogeogr. 2017, 26, 166–176. [Google Scholar] [CrossRef]
- Oberhuber, W.; Kofler, W. Topographic influences on radial growth of Scots pine (Pinus sylvestris L.) at small spatial scales. Plant Ecol. 2000, 146, 231–240. [Google Scholar] [CrossRef]
- Menzel, A.; Helm, R.; Zang, C. Patterns of late spring frost leaf damage and recovery in a European beech (Fagus sylvatica L.) stand in south-eastern Germany based on repeated digital photographs. Front. Plant Sci. 2015, 6, 110. [Google Scholar] [CrossRef] [Green Version]
- Príncipe, A.; van der Maaten, E.; van der Maaten-Theunissen, M.; Struwe, T.; Wilmking, M.; Kreyling, J. Low resistance but high resilience in growth of a major deciduous forest tree (Fagus sylvatica L.) in response to late spring frost in southern Germany. Trees 2017, 31, 743–751. [Google Scholar] [CrossRef]
- Hilton, G.M.; Packham, J.R. Variation in the masting of common beech (Fagus sylvatica L.) in northern Europe over two centuries (1800–2001). Forestry 2003, 76, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Overgaard, R.; Gemmel, P.; Karlsson, M. Effects of weather conditions on mast year frequency in beech (Fagus sylvatica L.) in Sweden. Forestry 2007, 80, 555–565. [Google Scholar] [CrossRef]
- Meinardus, C.; Bräuning, A. Zur Trockenstresstoleranz von Eichen und Buchen. LWF Aktuell 2011, 85, 9–11. [Google Scholar]
- Buras, A.; Rammig, A.; Zang, C.S. A novel approach for the identification of pointer years. Dendrochronologia 2020, 63, 125746. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name and Acronym | Species | lon[°E] | lat[°N] | Elev. [m asl] | Climate Station MAP/MAT | Geology | Soils |
|---|---|---|---|---|---|---|---|
| Bad Brückenau (HW) | F. sylvatica | 9.93 | 50.35 | 810 | 10.0° E 50.35° N, 518 m | Basalt | Cambisols |
| P. abies | 920 mm/8.1 °C | ||||||
| Wunsiedel (HC) | F. sylvatica | 11.80 | 49.97 | 795 | 11.84° E 49.98° N, 654 m | Sandstone | Cambisols (podsolic) |
| P. abies | 1140 mm/6.4 °C | ||||||
| Grafenau (HE) | F. sylvatica | 13.43 | 48.96 | 975 | 12.92° E 48.85° N, 314 m | Granite | Cambisols |
| P. abies | 13.53 | 48.91 | 990 | 960 mm/8.4 °C | Gleysols | ||
| Würzburg (LW) | F. sylvatica | 9.88 | 49.73 | 335 | 9.96° E 49.77° N, 268 m | Limestone | Cambisols/Luvisols |
| P. sylvestris | 580 mm/9.7 °C | ||||||
| Tennenlohe (LC) | F. sylvatica | 11.03 | 49.55 | 300 | 11.05° E 49.5° N, 314 m | Sand | Cambisols/Podsols |
| P. sylvestris | 620 mm/9.3 °C | ||||||
| Burglengenfeld (LE) | F. sylvatica | 11.95 | 49.21 | 415 | 11.69° E 49.15° N, 549 m | Limestone | Cambisols |
| P. sylvestris | 12.07 | 49.20 | 395 | 800 mm/8.0 °C |
| Year | Study Site | Species | Reason |
|---|---|---|---|
| 1976 | HW | F. sylvatica/P. abies | Drought year+ Year after a dry year |
| HC | P. abies | ||
| LW | F. sylvatica/P. sylvestris | ||
| LC | F. sylvatica/P. sylvestris | ||
| LE | F. sylvatica/P. sylvestris | ||
| 2011 | HW | F. sylvatica | Late frost event in spring |
| HC | F. sylvatica/P. abies | ||
| HE | F. sylvatica/P. abies | ||
| LW | F. sylvatica | ||
| 2018 | HW | P. abies | Drought year |
| HC | P. abies | ||
| LC | F. sylvatica/P. sylvestris | ||
| LE | P. sylvestris | ||
| 2019 | HW | P. abies | Year after a drought year+ drought year/dry summer |
| HC | P. abies | ||
| HE | P. abies | ||
| LW | P. sylvestris | ||
| LC | P. sylvestris | ||
| 2003 | LW | F. sylvatica/P. sylvestris | Drought year |
| LC | F. sylvatica | ||
| 1998 | LC | F. sylvatica/P. sylvestris | Year after a dry year + dry spring/summer |
| LW | F. sylvatica | ||
| 1991 | HW | P. abies | Drought year |
| HE | P. abies | ||
| 2012 | LW | P. sylvestris | Dry spring/summer |
| LE | P. sylvestris | ||
| 2009 | LW | F. sylvatica | Dry summer/autumn |
| LE | F. sylvatica | ||
| 2000 | HC | F. sylvatica | Dry and warm spring |
| LE | F. sylvatica | ||
| 2013 | LC | P. sylvestris | Dry spring+ |
| LE | P. sylvestris | Year after a dry year | |
| 2004 | HW | F. sylvatica | Year after a drought year |
| HC | F. sylvatica | ||
| 2010 | HW | F. sylvatica | Year after a warm summer/autumn |
| LE | F. sylvatica | ||
| 1987 | HC | F. sylvatica | Wet and cold summer |
| HE | F. sylvatica | ||
| 1996 | HE | F. sylvatica/P. abies | Cold summer |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Debel, A.; Meier, W.J.-H.; Bräuning, A. Climate Signals for Growth Variations of F. sylvatica, P. abies, and P. sylvestris in Southeast Germany over the Past 50 Years. Forests 2021, 12, 1433. https://doi.org/10.3390/f12111433
Debel A, Meier WJ-H, Bräuning A. Climate Signals for Growth Variations of F. sylvatica, P. abies, and P. sylvestris in Southeast Germany over the Past 50 Years. Forests. 2021; 12(11):1433. https://doi.org/10.3390/f12111433
Chicago/Turabian StyleDebel, Annette, Wolfgang Jens-Henrik Meier, and Achim Bräuning. 2021. "Climate Signals for Growth Variations of F. sylvatica, P. abies, and P. sylvestris in Southeast Germany over the Past 50 Years" Forests 12, no. 11: 1433. https://doi.org/10.3390/f12111433
APA StyleDebel, A., Meier, W. J.-H., & Bräuning, A. (2021). Climate Signals for Growth Variations of F. sylvatica, P. abies, and P. sylvestris in Southeast Germany over the Past 50 Years. Forests, 12(11), 1433. https://doi.org/10.3390/f12111433