
Thinning Response and Potential Basal Area—A Case Study in a Mixed Sub-Humid Low-Elevation Oak-Hornbeam Forest



Abstract
:1. Introduction
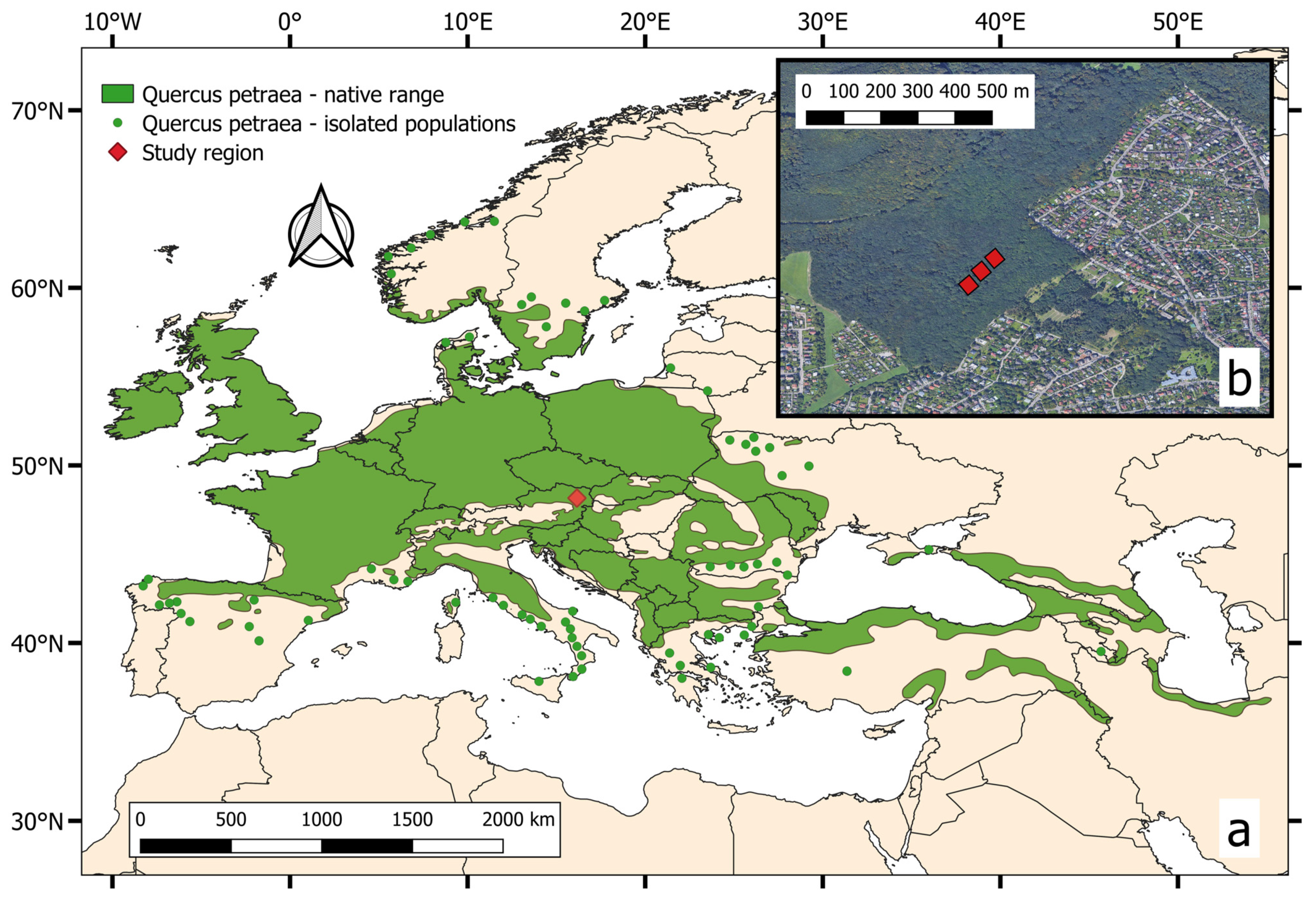
2. Materials and Methods
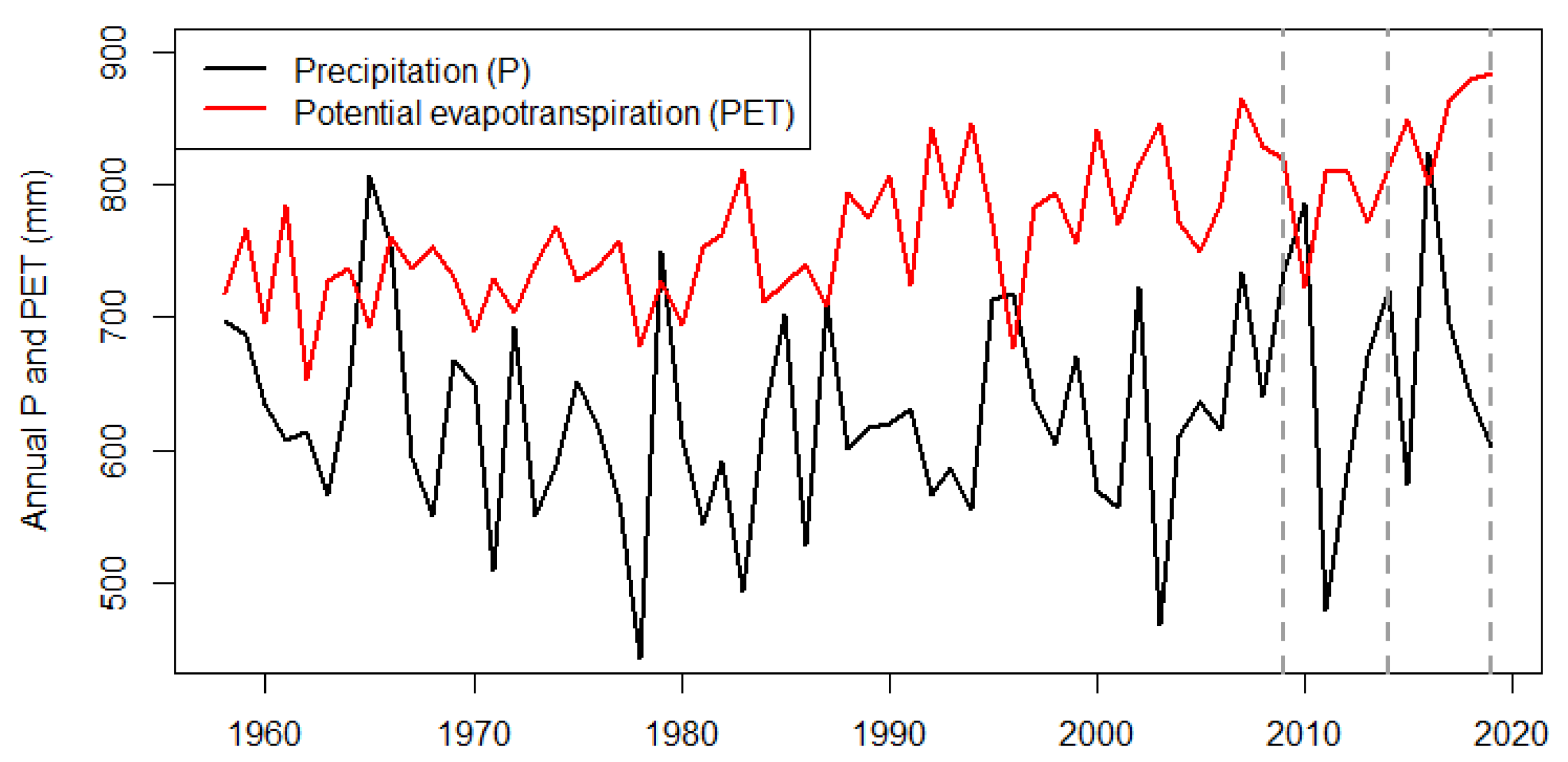
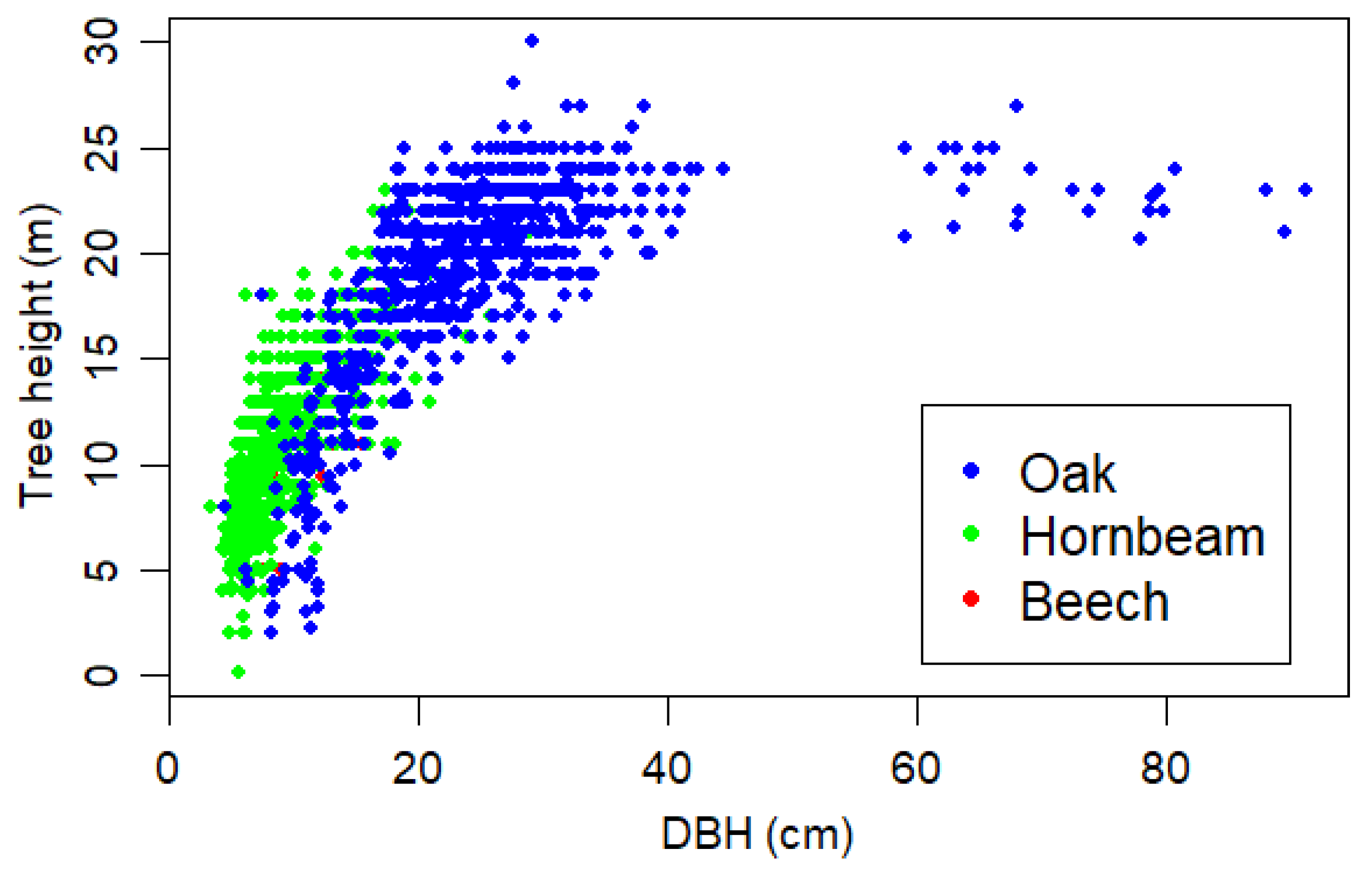
2.1. Data
2.2. Analysis
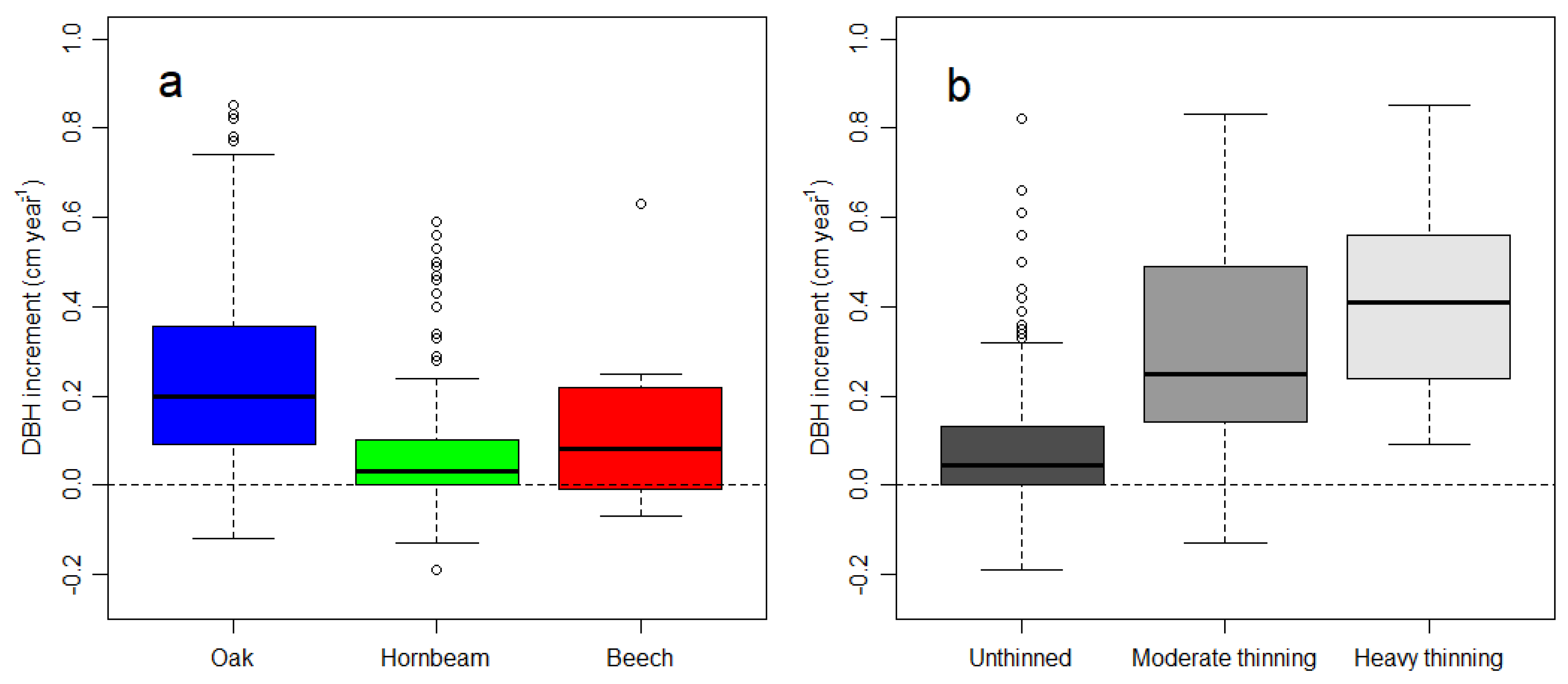
3. Results
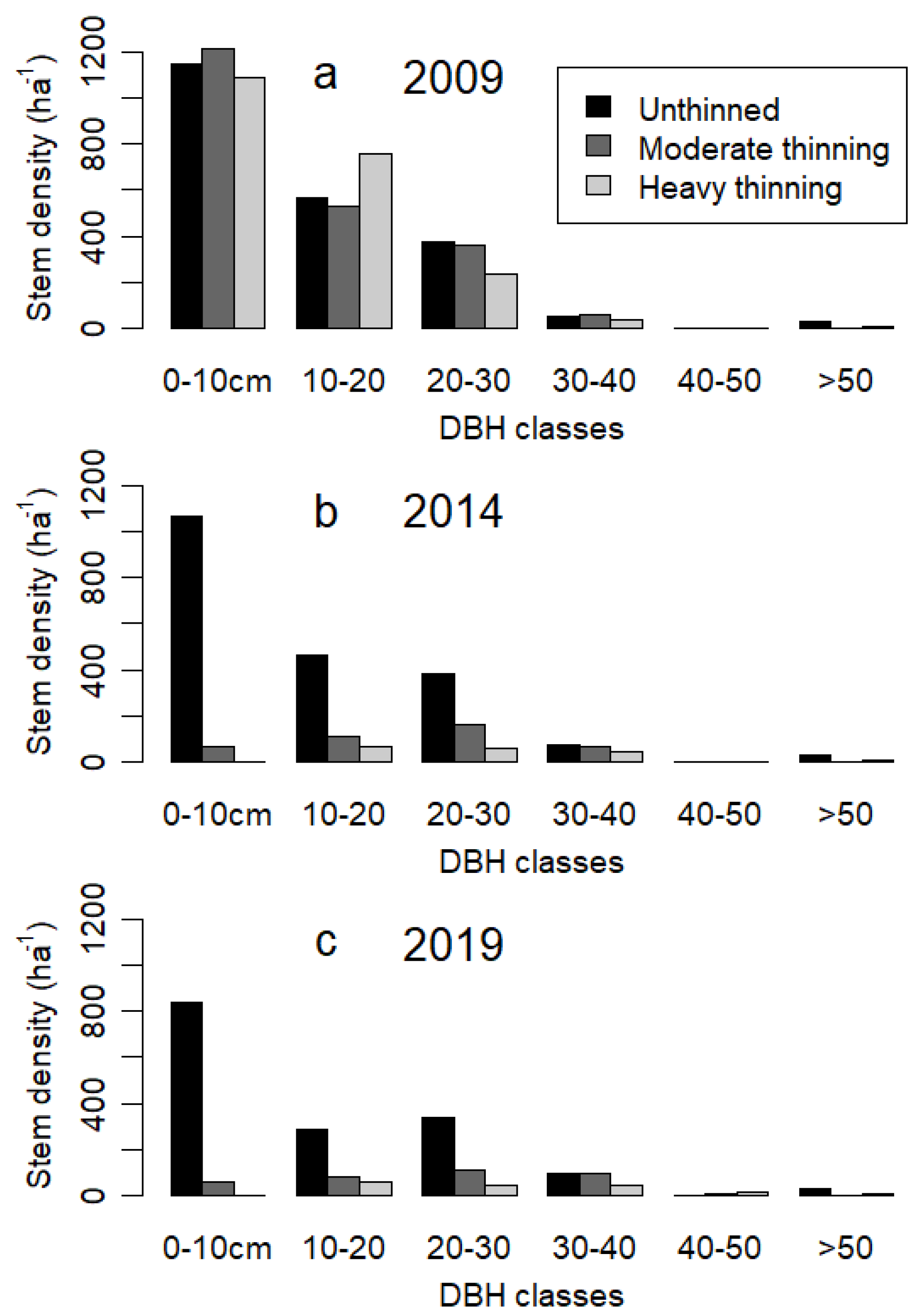
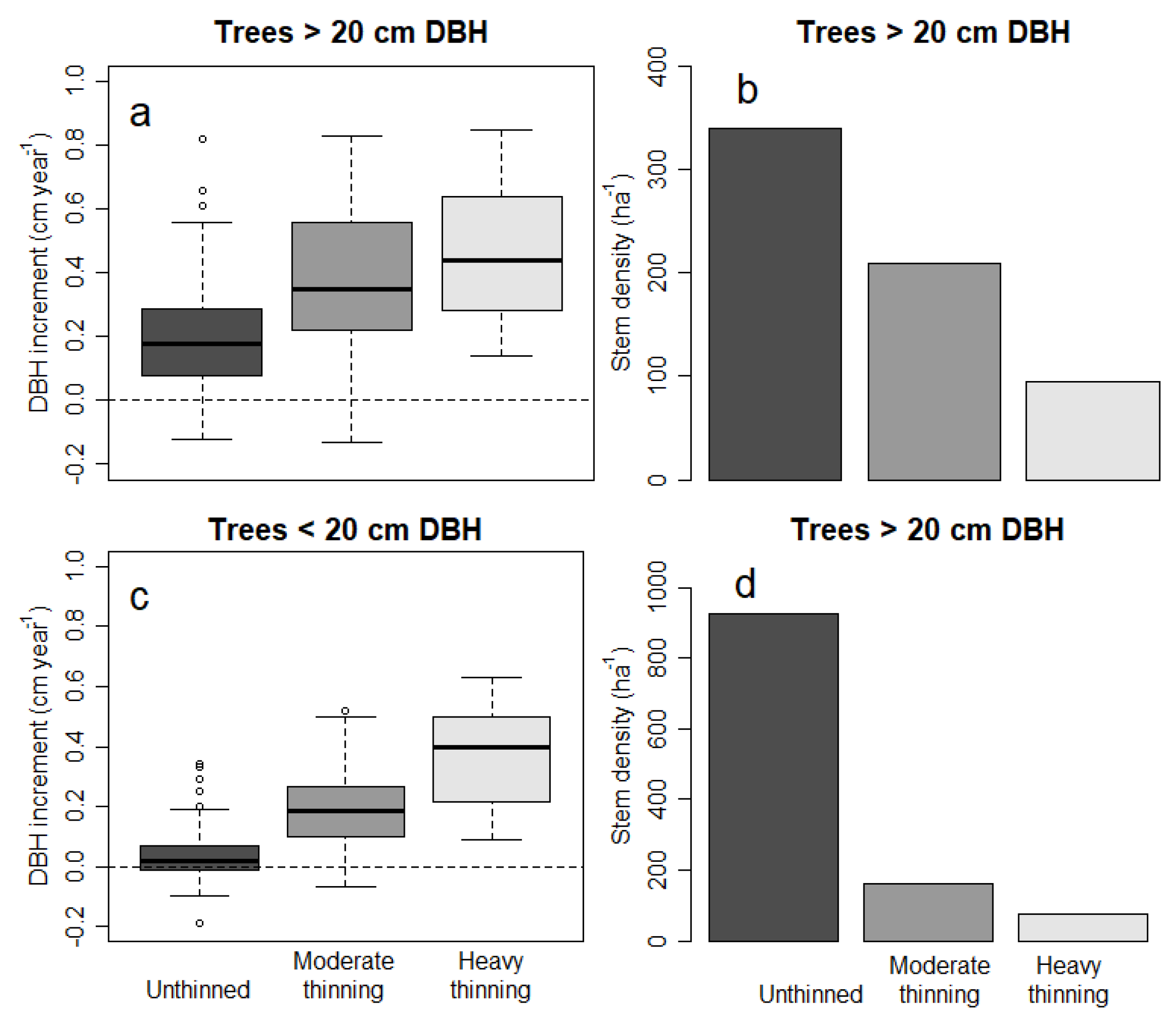
3.1. Thinning and Stand Structure
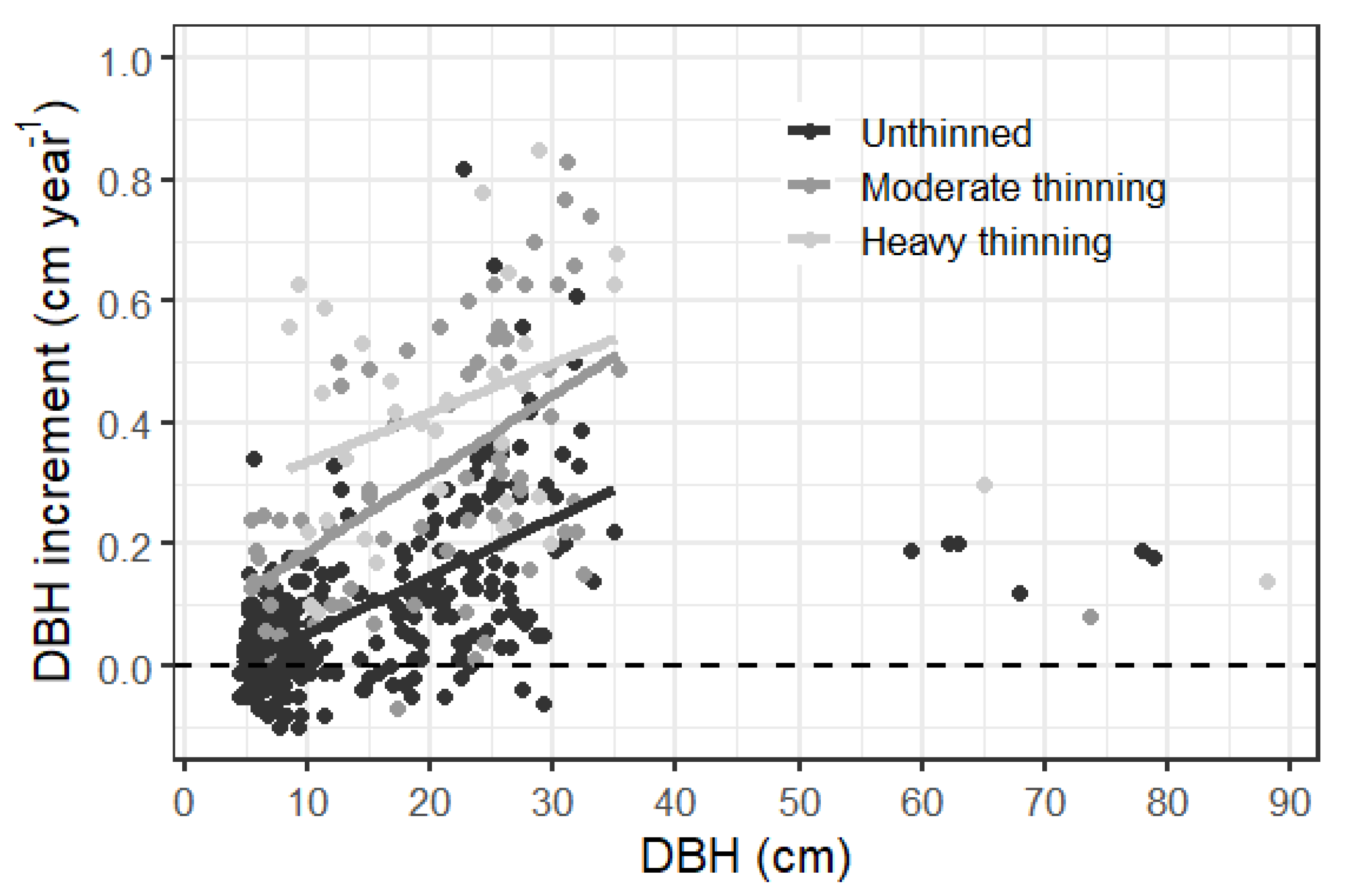
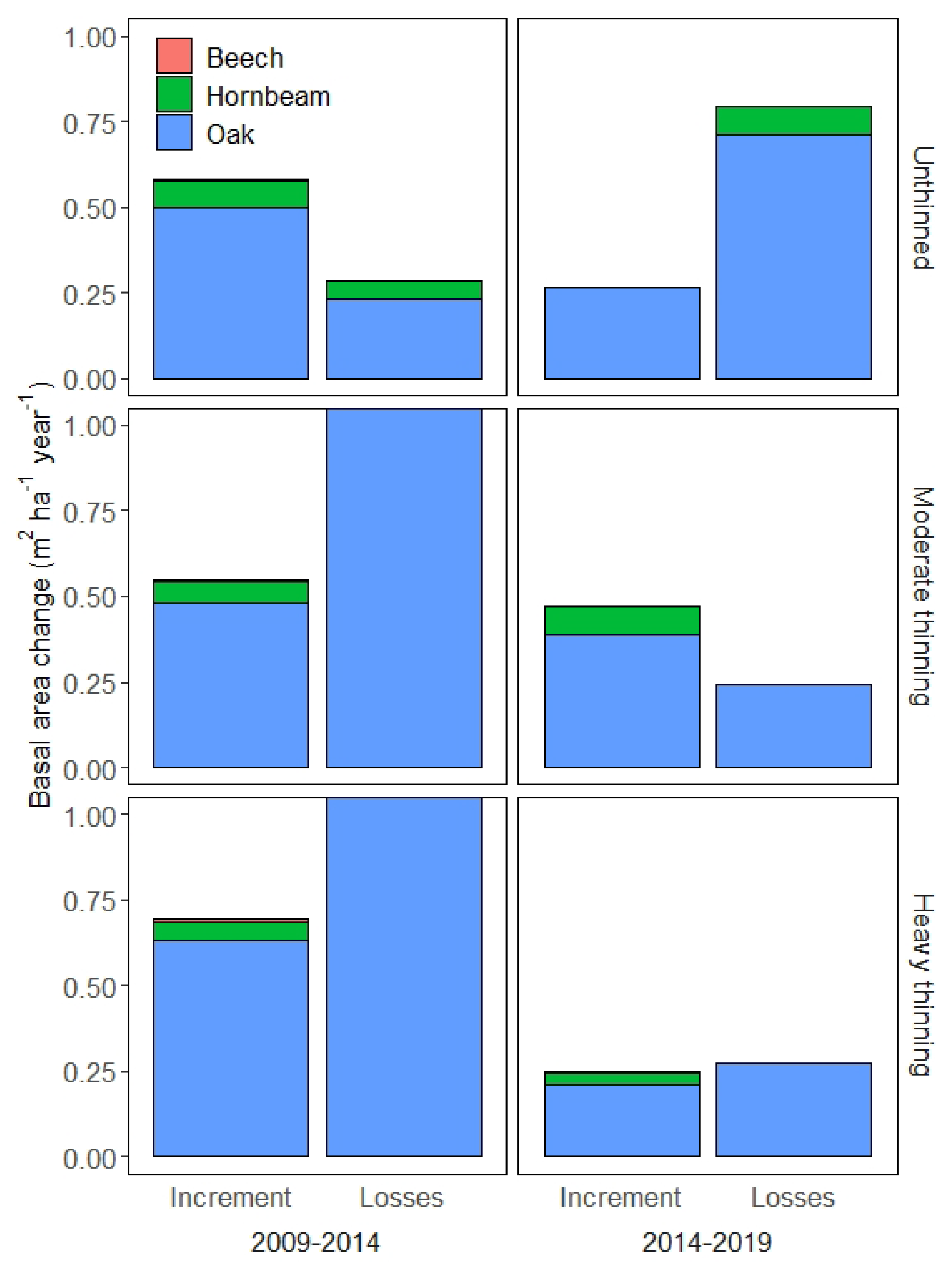
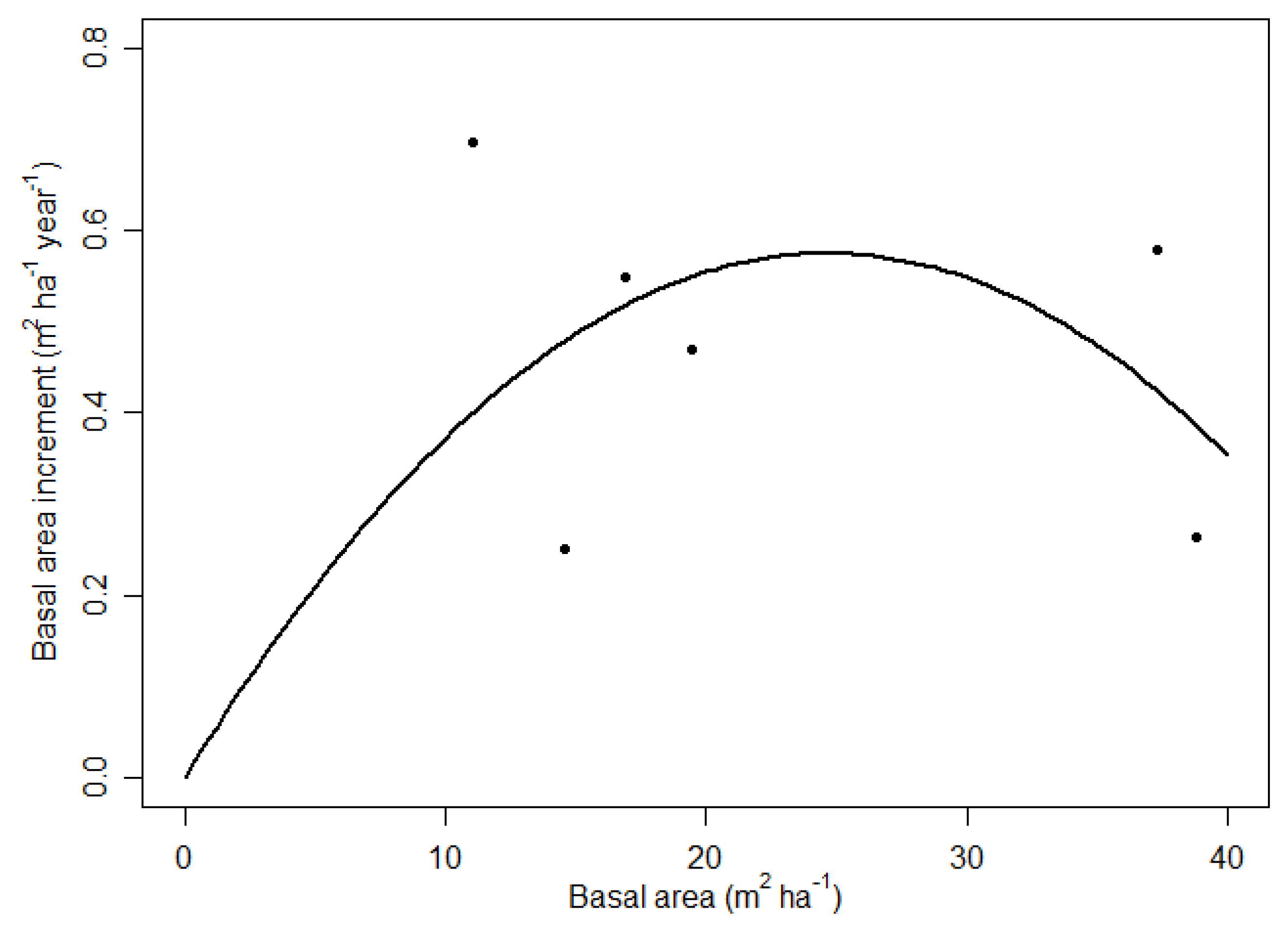
3.2. Increment and Losses
4. Discussion
4.1. Thinning Response
4.2. Maximum Basal Area
4.3. Harvesting versus Natural Mortality
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Br. 2017, 12, 662–666. [Google Scholar] [CrossRef]
- BFW. Österreichische Waldinventur. 2017. Available online: http://bfw.ac.at/rz/wi.home (accessed on 4 March 2021).
- Haneca, K.; Van acker, J.; Beeckman, H. Growth trends reveal the forest structure during Roman and Medieval times in Western Europe: A comparison between archaeological and actual oak ring series (Quercus robur and Quercus petraea). Ann. For. Sci. 2005, 62, 797–805. [Google Scholar] [CrossRef] [Green Version]
- Pötzelsberger, E.; Wolfslehner, B.; Hasenauer, H. Climate change impacts on key forest functions of the Vienna Woods. Eur. J. For. Res. 2015, 134, 481–496. [Google Scholar] [CrossRef]
- Oettel, J.; Lapin, K.; Kindermann, G.; Steiner, H.; Schweinzer, K.; Frank, G.; Essl, F. Patterns and drivers of deadwood volume and composition in different forest types of the Austrian natural forest reserves. For. Ecol. Manag. 2020, 463, 118016. [Google Scholar] [CrossRef]
- Mölder, A.; Meyer, P.; Nagel, R.V. Integrative management to sustain biodiversity and ecological continuity in Central European temperate oak (Quercus robur, Q. petraea) forests: An overview. For. Ecol. Manag. 2019, 437, 324–339. [Google Scholar] [CrossRef]
- Ortmann-Ajkai, A.; Csicsek, G.; Hollos, R.; Kevey, B.; Borhidi, A. Comparison of spontaneous regeneration in unmanaged oak (Quercus robur L.) and beech (Fagus sylvatica L.) forests: Implications for close-to-nature silviculture. Austrian J. For. Sci. 2016, 133, 223–250. [Google Scholar]
- Bruckman, V.J.; Terada, T.; Fukuda, K.; Yamamoto, H.; Hochbichler, E. Overmature periurban Quercus–Carpinus coppice forests in Austria and Japan: A comparison of carbon stocks, stand characteristics and conversion to high forest. Eur. J. For. Res. 2016, 135, 1–13. [Google Scholar] [CrossRef]
- Stimm, K.; Heym, M.; Uhl, E.; Tretter, S.; Pretzsch, H. Height growth-related competitiveness of oak (Quercus petraea (Matt.) Liebl. and Quercus robur L.) under climate change in Central Europe. Is silvicultural assistance still required in mixed-species stands? For. Ecol. Manag. 2021, 482, 118780. [Google Scholar] [CrossRef]
- Zeller, L.; Pretzsch, H. Effect of forest structure on stand productivity in Central European forests depends on developmental stage and tree species diversity. For. Ecol. Manag. 2019, 434, 193–204. [Google Scholar] [CrossRef]
- Assmann, E. The Principles of Forest Yield Study; Pergamon Press: New York, NY, USA, 1970. [Google Scholar]
- Gavinet, J.; Ourcival, J.M.; Gauzere, J.; García de Jalón, L.; Limousin, J.M. Drought mitigation by thinning: Benefits from the stem to the stand along 15 years of experimental rainfall exclusion in a holm oak coppice. For. Ecol. Manag. 2020, 473, 118266. [Google Scholar] [CrossRef]
- Hochbichler, E.; Bellos, P.; Lick, E. Biomass functions for estimating needle and branch biomass of spruce (Picea abies) and Scots pine (Pinus sylvestris) and branch biomass of beech (Fagus sylvatica) and oak (Quercus robur and petrea). Austrian J. For. Sci. 2006, 123, 35–46. [Google Scholar]
- Suchomel, C.; Pyttel, P.; Becker, G.; Bauhus, J. Biomass equations for sessile oak (Quercus petraea (Matt.) Liebl.) and hornbeam (Carpinus betulus L.) in aged coppiced forests in southwest Germany. Biomass Bioenergy 2012, 1–9. [Google Scholar] [CrossRef]
- Del Río, M.; Calama, R.; Cañellas, I.; Roig, S.; Montero, G. Thinning intensity and growth response in SW-European Scots pine stands. Ann. For. Sci. 2008, 65, 308. [Google Scholar] [CrossRef] [Green Version]
- Jensen, F.S.; Skovsgaard, J.P. Precommercial thinning of pedunculate oak: Recreational preferences of the population of Denmark for different thinning practices in young stands. Scand. J. For. Res. 2009, 24, 28–36. [Google Scholar] [CrossRef]
- Tyrväinen, L.; Silvennoinen, H.; Kolehmainen, O. Ecological and aesthetic values in urban forest management. Urban For. Urban Green. 2003, 1, 135–149. [Google Scholar] [CrossRef]
- Fedorová, B.; Kadavý, J.; Adamec, Z.; Kneifl, M.; Knott, R. Response of diameter and height increment to thinning in oak-hornbeam coppice in the southeastern part of the Czech Republic. J. For. Sci. 2016, 62, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, A.; Trouvé, R.; Seynave, I.; Lebourgeois, F. Decreasing stand density favors resistance, resilience, and recovery of Quercus petraea trees to a severe drought, particularly on dry sites. Ann. For. Sci. 2020, 77. [Google Scholar] [CrossRef]
- Breda, N.; Granier, A.; Aussenac, G. Effects of thinning on soil and tree water relations, transpiration and growth in an oak forest (Quercus petraea (Matt.) Liebl.). Tree Physiol. 1995, 15, 295–306. [Google Scholar] [CrossRef]
- Tinya, F.; Kovács, B.; Aszalós, R.; Tóth, B.; Csépányi, P.; Németh, C.; Ódor, P. Initial regeneration success of tree species after different forestry treatments in a sessile oak-hornbeam forest. For. Ecol. Manag. 2020, 459, 117810. [Google Scholar] [CrossRef]
- Führer, E.; Horváth, L.; Jagodics, A.; Machon, A.; Szabados, I. Application of a new aridity index in Hungarian forestry practice. Idojaras 2011, 115, 205–216. [Google Scholar]
- Nabuurs, G.-J.; Lindner, M.; Verkerk, P.J.; Gunia, K.; Deda, P.; Michalak, R.; Grassi, G. First signs of carbon sink saturation in European forest biomass. Nat. Clim. Chang. 2013, 3, 792–796. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, T.; Kimberley, M.O.; Beets, P.N. Natural forests in New Zealand – a large terrestrial carbon pool in a national state of equilibrium. For. Ecosyst. 2021, 8. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958-2015. Sci. Data 2018, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity - evidence since the middle of the 20th century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Creed, I.F.; Spargo, A.T.; Jones, J.A.; Buttle, J.M.; Adams, M.B.; Beall, F.D.; Booth, E.G.; Campbell, J.L.; Clow, D.; Elder, K.; et al. Changing forest water yields in response to climate warming: Results from long-term experimental watershed sites across North America. Glob. Chang. Biol. 2014, 20, 3191–3208. [Google Scholar] [CrossRef] [Green Version]
- San-Miguel-Ayanz, J.; de Rigo, D.; Caudullo, G.; Houston Durrant, T.; Mauri, A. European Atlas of Forest Tree Species; Publication Office of the European Union: Luxembourg, Belgium, 2016; ISBN 978-92-79-52833-0. [Google Scholar]
- Hasenauer, H.; Eastaugh, C.S. Assessing Forest Production Using Terrestrial Monitoring Data. Int. J. For. Res. 2012, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Pollanschütz, J. Formzahlfunktionen der Hauptbaumarten Österreichs. Informationsd. Forstl. Bundesversuchsanstalt Wien 1974, 153, 341–343. [Google Scholar]
- Marschall, J. Hilfstafeln für die Forsteinrichtung, 5th ed.; Österreichischer Agrarverlag: Wien, Austria, 1992. [Google Scholar]
- Burrows, W.H.; Henry, B.K.; Back, P.V.; Hoffmann, M.B.; Tait, L.J.; Anderson, E.R.; Menke, N.; Danaher, T.; Carter, J.O.; McKeon, G.M. Growth and carbon stock change in eucalypt woodlands in northeast Australia: Ecological and greenhouse sink implications. Glob. Chang. Biol. 2002, 8, 769–784. [Google Scholar] [CrossRef]
- Bitterlich, W. Die Winkelzaehlprobe. Allg. Forst-und Holzwirtschaftliche Zeitung 1948, 59, 4–5. [Google Scholar]
- Neumann, M.; Moreno, A.; Mues, V.; Härkönen, S.; Mura, M.; Bouriaud, O.; Lang, M.; Achten, W.M.J.; Thivolle-Cazat, A.; Bronisz, K.; et al. Comparison of carbon estimation methods for European forests. For. Ecol. Manag. 2016, 361, 397–420. [Google Scholar] [CrossRef]
- Harmon, M.E.; Fasth, B.G.; Yatskov, M.; Kastendick, D.; Rock, J.; Woodall, C.W. Release of coarse woody detritus-related carbon: A synthesis across forest biomes. Carbon Balance Manag. 2020, 15, 1–21. [Google Scholar] [CrossRef]
- Pukkala, T. Does management improve the carbon balance of forestry? Forestry 2017, 90, 125–135. [Google Scholar] [CrossRef]
- Thom, D.; Seidl, R. Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biol. Rev. 2016, 91, 760–781. [Google Scholar] [CrossRef] [PubMed]
- Dean, C.; Kirkpatrick, J.B.; Osborn, J.; Doyle, R.B.; Fitzgerald, N.B.; Roxburgh, S.H. Novel 3D geometry and models of the lower regions of large trees for use in carbon accounting of primary forests. AoB Plants 2018, 10, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Roxburgh, S.H.; Wood, S.W.; Mackey, B.G.; Woldendorp, G.; Gibbons, P. Assessing the carbon sequestration potential of managed forests: A case study from temperate Australia. J. Appl. Ecol. 2006, 43, 1149–1159. [Google Scholar] [CrossRef]
- Grotti, M.; Chianucci, F.; Puletti, N.; Fardusi, M.J.; Castaldi, C.; Corona, P. Spatio-temporal variability in structure and diversity in a semi-natural mixed oak-hornbeam floodplain forest. Ecol. Indic. 2019, 104, 576–587. [Google Scholar] [CrossRef]
- Petritan, A.M.; Biris, I.A.; Merce, O.; Turcu, D.O.; Petritan, I.C. Structure and diversity of a natural temperate sessile oak (Quercus petraea L.) - European Beech (Fagus sylvatica L.) forest. For. Ecol. Manag. 2012, 280, 140–149. [Google Scholar] [CrossRef]
- Woldendorp, G.; Keenan, R.J.; Barry, S.; Spencer, R.D. Analysis of sampling methods for coarse woody debris. For. Ecol. Manag. 2004, 198, 133–148. [Google Scholar] [CrossRef]
- Stereńczak, K.; Mielcarek, M.; Wertz, B.; Bronisz, K.; Zajączkowski, G.; Jagodziński, A.M.; Ochał, W.; Skorupski, M. Factors influencing the accuracy of ground-based tree-height measurements for major European tree species. J. Environ. Manag. 2019, 231, 1284–1292. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unthinned | Moderate Thinning | Heavy Thinning | |||
|---|---|---|---|---|---|
| Measurement date | 2009 | 2009 − | 2009 + | 2009 − | 2009 + |
| BA (m2 ha−1) | 37.3 | 37.0 | 16.9 | 36.4 | 11.1 |
| NHA (ha−1) | 1736 | 2170 | 410 | 2135 | 180 |
| Species mixture (%) | 88, 12, <1 | 69, 30, 1 | 87, 12, 1 | 59, 41, <1 | 91, 8, 1 |
| Mean tree height (m) | 12.2 | 12.6 | 17.1 | 12.9 | 17.9 |
| Volume (m3 ha−1) | 327 | 308 | 157 | 300 | 106 |
| Yield class (m3 ha−1 yr−1) | 4.5 | 4.5 | 4.5 | ||
| Plot size (ha) | 0.25 | 0.2 | 0.2 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neumann, M.; Hasenauer, H. Thinning Response and Potential Basal Area—A Case Study in a Mixed Sub-Humid Low-Elevation Oak-Hornbeam Forest. Forests 2021, 12, 1354. https://doi.org/10.3390/f12101354
Neumann M, Hasenauer H. Thinning Response and Potential Basal Area—A Case Study in a Mixed Sub-Humid Low-Elevation Oak-Hornbeam Forest. Forests. 2021; 12(10):1354. https://doi.org/10.3390/f12101354
Chicago/Turabian StyleNeumann, Mathias, and Hubert Hasenauer. 2021. "Thinning Response and Potential Basal Area—A Case Study in a Mixed Sub-Humid Low-Elevation Oak-Hornbeam Forest" Forests 12, no. 10: 1354. https://doi.org/10.3390/f12101354
APA StyleNeumann, M., & Hasenauer, H. (2021). Thinning Response and Potential Basal Area—A Case Study in a Mixed Sub-Humid Low-Elevation Oak-Hornbeam Forest. Forests, 12(10), 1354. https://doi.org/10.3390/f12101354