
Thermotherapy and Storage Temperature Manipulations Limit the Production of Reactive Oxygen Species in Stored Pedunculate Oak Acorns



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.2. Temperature Manipulations before and during Storage
2.2.1. Thermotherapy
2.2.2. Acclimation to Lower Temperature
2.3. Determination of ROS Release
2.3.1. Release of Hydrogen Peroxide (H2O2)
2.3.2. Release of Superoxide Anion Radical (O2•−)
2.3.3. Release of Hydroxyl Radical (•OH)
2.4. Seed Viability Determination
2.4.1. Germination Test (Hypocotyl Emergence)
2.4.2. Seedling Emergence (Epicotyl Emergence)
2.4.3. Electrolyte Leakage
2.5. Statistical Analyses
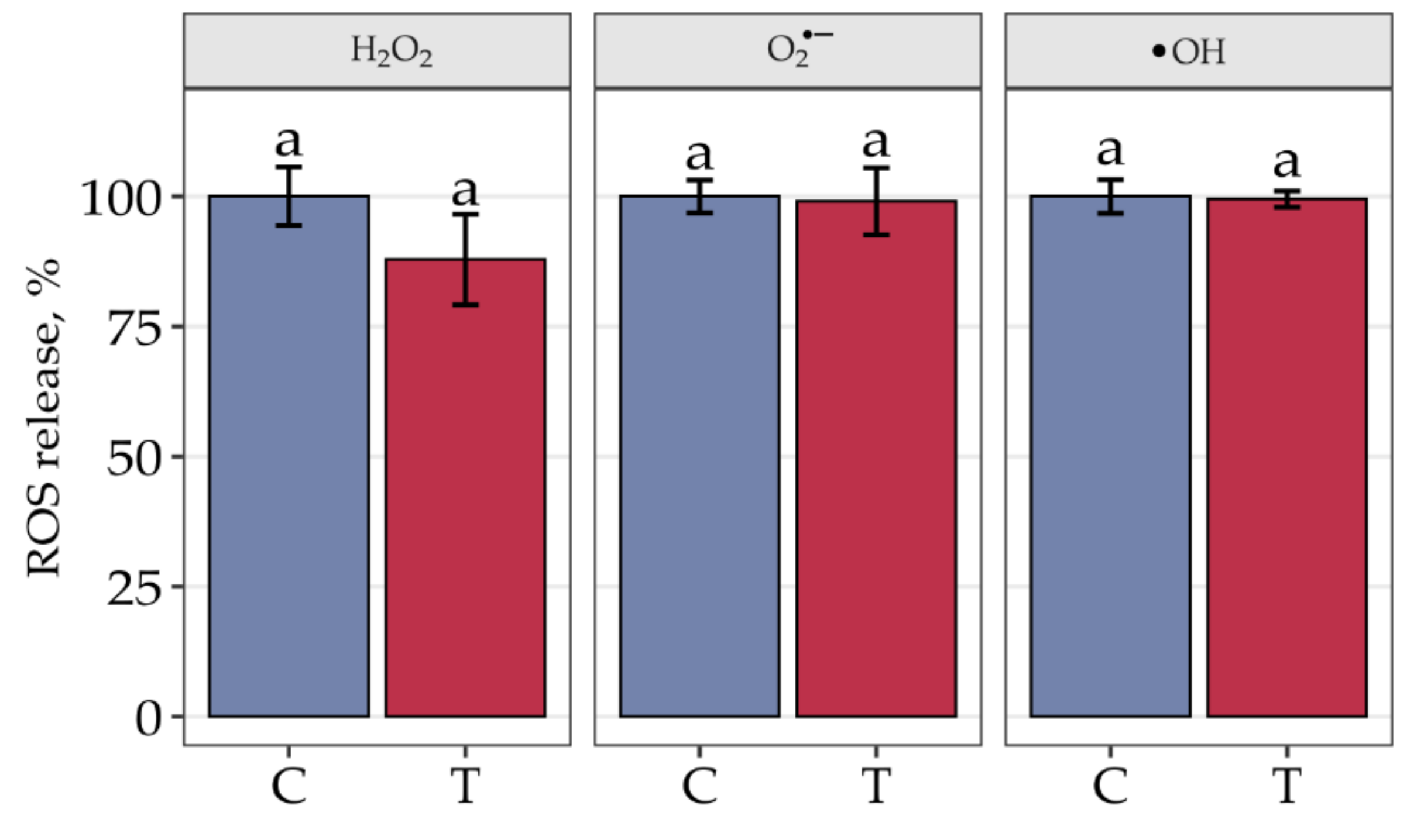
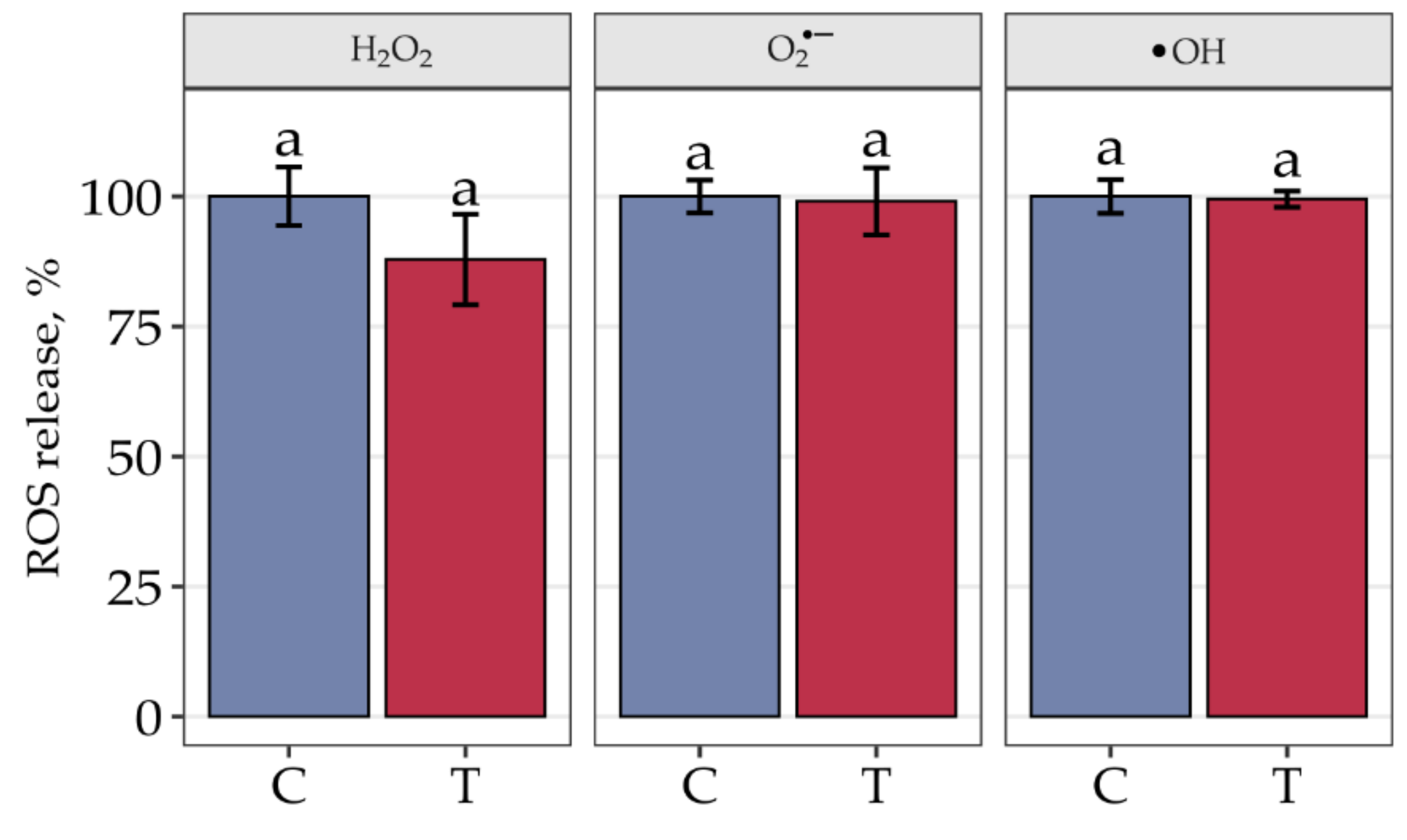
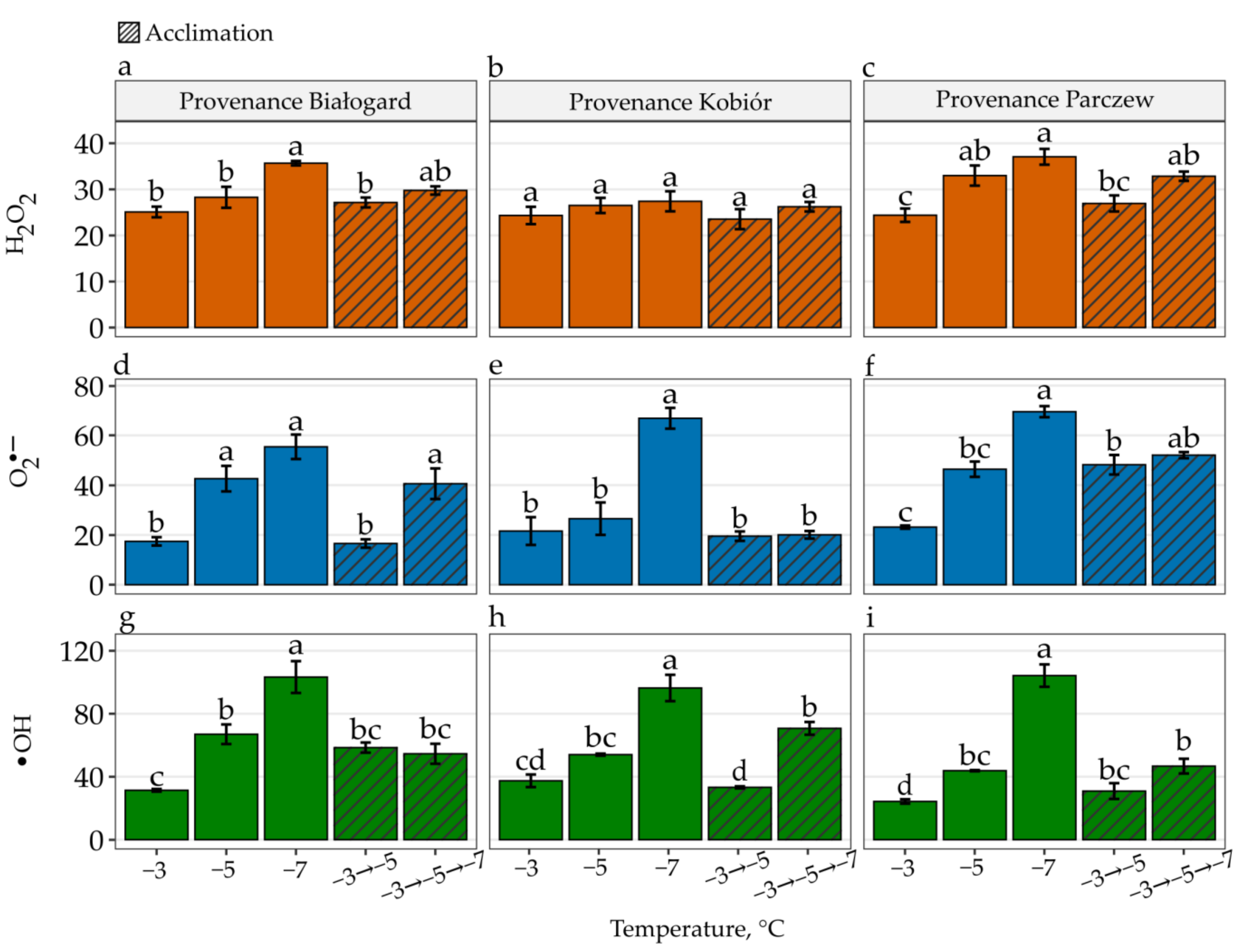
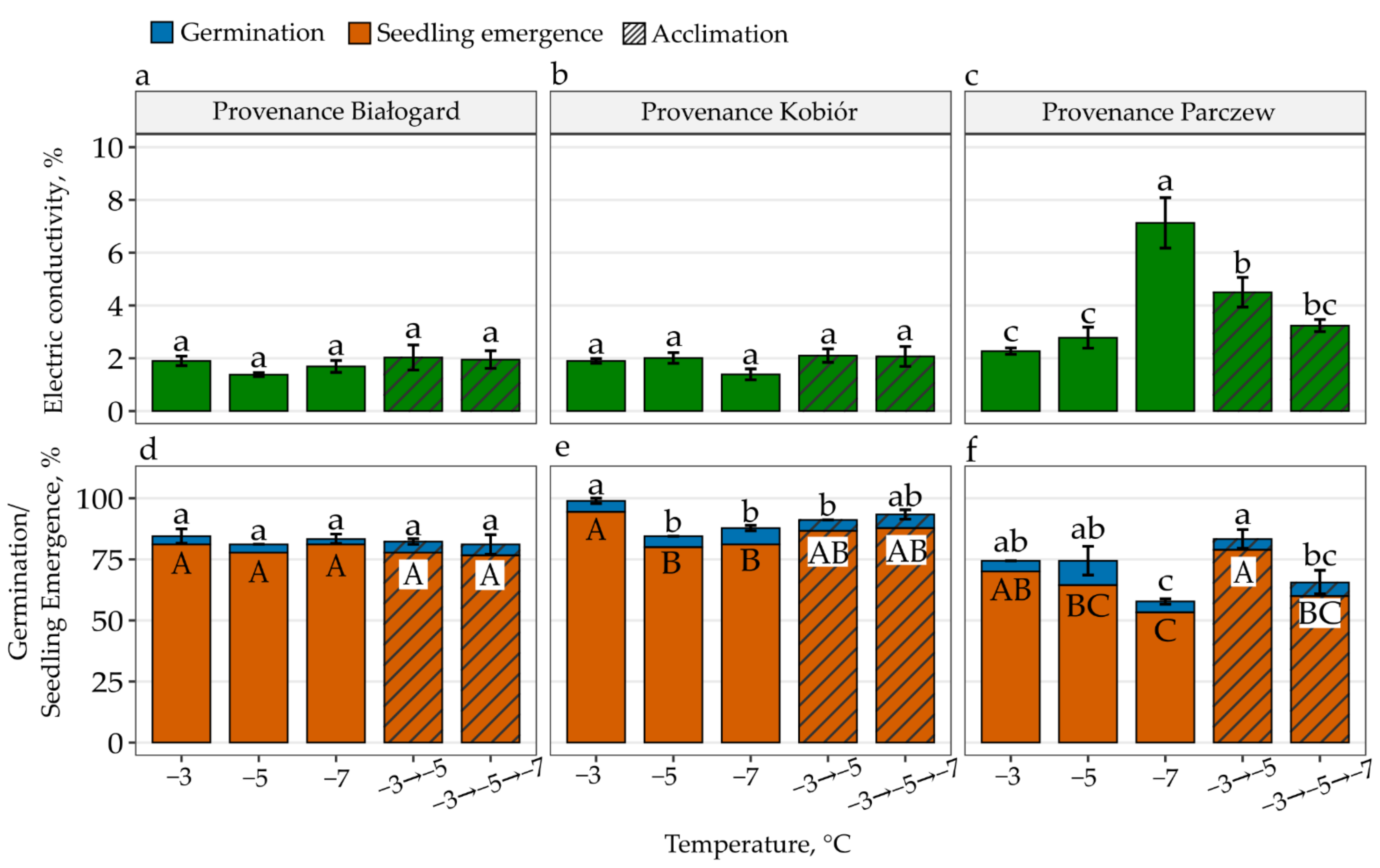
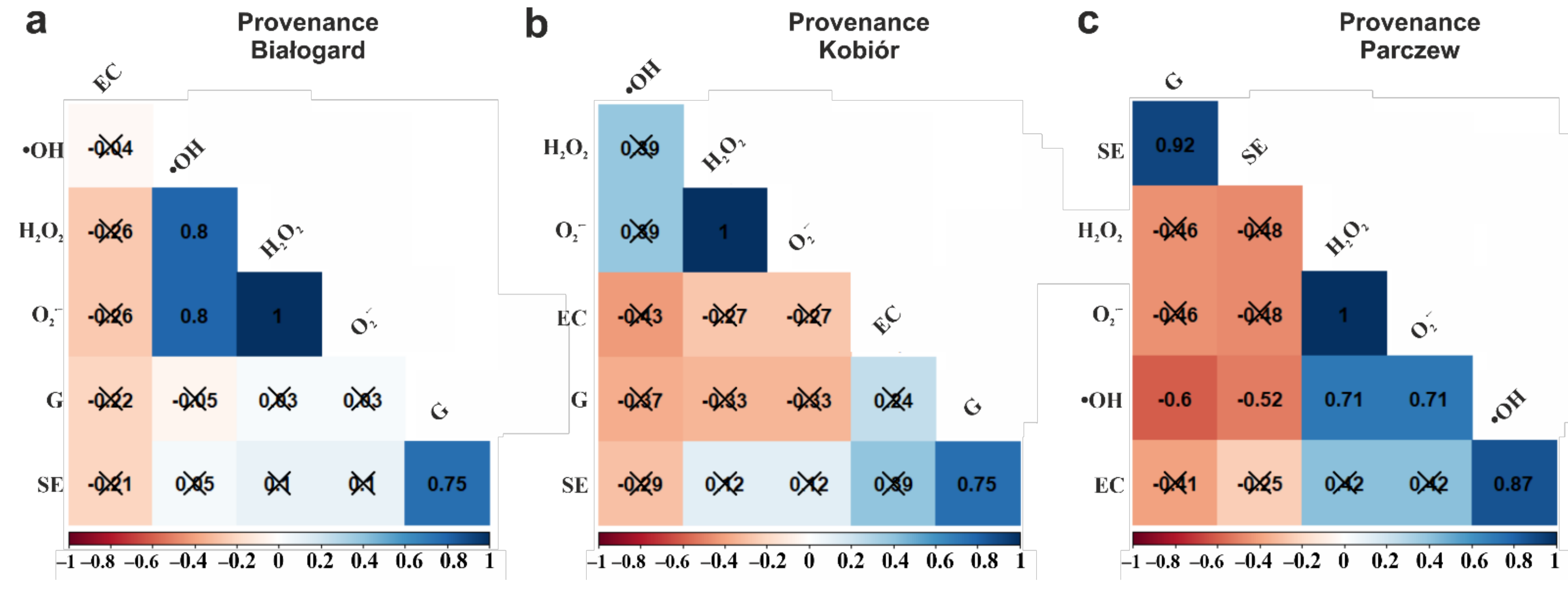
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buckley, L.B.; Huey, R.B. How Extreme Temperatures Impact Organisms and the Evolution of Their Thermal Tolerance. Integr. Comp. Biol. 2016, 56, 98–109. [Google Scholar] [CrossRef] [Green Version]
- Kaspari, M.; Clay, N.A.; Lucas, J.; Yanoviak, S.P.; Kay, A. Thermal Adaptation Generates a Diversity of Thermal Limits in a Rainforest Ant Community. Glob. Chang. Biol. 2015, 21, 1092–1102. [Google Scholar] [CrossRef]
- Gilchrist, G. Chapter 5 The Evolution of Thermal Sensitivity in Changing Environments. Cell Mol. Response Stress 2000, 1, 55–70. [Google Scholar] [CrossRef]
- Mercer, J. Glossary of Terms for Thermal Physiology, Third Edition. Jpn. J. Physiol. 2001, 51, 245–280. [Google Scholar]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat Tolerance in Plants: An Overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Kidokoro, S.; Yoneda, K.; Takasaki, H.; Takahashi, F.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Different Cold-Signaling Pathways Function in the Responses to Rapid and Gradual Decreases in Temperature. Plant Cell 2017, 29, 760–774. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Shi, Y.; Yang, S. Advances and Challenges in Uncovering Cold Tolerance Regulatory Mechanisms in Plants. New Phytol. 2019, 222, 1690–1704. [Google Scholar] [CrossRef] [Green Version]
- Fallik, E.; Ilić, Z. The Influence of Physical Treatments on Phytochemical Changes in Fresh Produce after Storage and Marketing. Agronomy 2021, 11, 788. [Google Scholar] [CrossRef]
- Ellis, J.B.; Martin, G.B. Macrosporium solani E&M. Am. Nat. 1882, 16, 1003. [Google Scholar]
- Schröder, T.; Kehr, R.; Procházková, Z.; Sutherland, J.R. Practical Methods for Estimating the Infection Rate of Quercus robur Acorn Seedlots by Ciboria batschiana. For. Pathol. 2004, 34, 187–196. [Google Scholar] [CrossRef]
- Merouani, H.; Trubat, R.; Lourenco, M.; Sampaio, T.; Santos, M.; Cortina, J.; Pereira, J.; Almeida, M.H. Fungus Development, a Factor Limiting the Long-Term Storage of Cork Oak Acorns. IOBC WPRS Bull. 2005, 28, 129. [Google Scholar]
- Delatour, C.; Muller, C.; Bonnet-Masimbert, M. Progress in Acorns Treatment in a Long Term Storage Prospect. In Proceedings of the International Symposium on Forest Tree Seed Storage, Chalk River, ON, Canada, 23–27 September 1980. [Google Scholar]
- Suszka, B.; Muller, C.; Bonnet-Masimbert, M.; Gordon, A. Seeds of Forest Broadleaves: From Harvest to Sowing; Institute National de la Recherche Agronomique: Paris, France, 1996; ISBN 978-2-7380-0659-2. [Google Scholar]
- Finch-Savage, W.; Clay, H.; Budge, S.; Dent, K.; Clarkson, J.; Whipps, J. Biological Control of Sclerotinia Pseudotuberosa and Other Fungi During Moist Storage of Quercus robur Seeds. Eur. J. Plant Pathol. 2003, 109, 615–624. [Google Scholar] [CrossRef]
- Knudsen, I.; Thomsen, K.; Jensen, B.; Poulsen, K. Effects of Hot Water Treatment, Biocontrol Agents, Disinfectants and a Fungicide on Storability of English Oak Acorns and Control of the Pathogen, Ciboria batschiana. For. Pathol. 2004, 34, 47–64. [Google Scholar] [CrossRef]
- Kranjec Orlović, J.; Drvodelić, D.; Vukelić, M.; Rukavina, M.; Diminić, D.; Oršanić, M. Impact of Thermotherapy and Short-Term Storage on Quercus robur L. Acorn Mycobiota and Germination. Forests 2021, 12, 528. [Google Scholar] [CrossRef]
- Suzuki, N.; Mittler, R. Reactive Oxygen Species and Temperature Stresses: A Delicate Balance between Signaling and Destruction. Physiol. Plant. 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Pandey, P.; Ramegowda, V.; Senthil-Kumar, M. Shared and Unique Responses of Plants to Multiple Individual Stresses and Stress Combinations: Physiological and Molecular Mechanisms. Front. Plant Sci. 2015, 6, 723. [Google Scholar] [CrossRef] [Green Version]
- Loayza, F.E.; Brecht, J.K.; Simonne, A.H.; Plotto, A.; Baldwin, E.A.; Bai, J.; Lon-Kan, E. Enhancement of the Antioxidant Capacity of Ripe Tomatoes by the Application of a Hot Water Treatment at the Mature-Green Stage. Postharvest Biol. Technol. 2020, 161, 111054. [Google Scholar] [CrossRef]
- Rajjou, L.; Debeaujon, I. Seed Longevity: Survival and Maintenance of High Germination Ability of Dry Seeds. C. R. Biol. 2008, 331, 796–805. [Google Scholar] [CrossRef]
- Renard, J.; Niñoles, R.; Martínez-Almonacid, I.; Gayubas, B.; Mateos-Fernández, R.; Bissoli, G.; Bueso, E.; Serrano, R.; Gadea, J. Identification of Novel Seed Longevity Genes Related to Oxidative Stress and Seed Coat by Genome-Wide Association Studies and Reverse Genetics. Plant Cell Environ. 2020, 43, 2523–2539. [Google Scholar] [CrossRef]
- Harman, D. Aging: A Theory Based on Free Radical and Radiation Chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [Green Version]
- Bailly, C. The Signalling Role of ROS in the Regulation of Seed Germination and Dormancy. Biochem. J. 2019, 476, 3019–3032. [Google Scholar] [CrossRef]
- Jeevan Kumar, S.P.; Rajendra Prasad, S.; Banerjee, R.; Thammineni, C. Seed Birth to Death: Dual Functions of Reactive Oxygen Species in Seed Physiology. Ann. Bot. 2015, 116, 663–668. [Google Scholar] [CrossRef] [Green Version]
- Walters, C. Orthodoxy, Recalcitrance and in-between: Describing Variation in Seed Storage Characteristics Using Threshold Responses to Water Loss. Planta 2015, 242, 397–406. [Google Scholar] [CrossRef]
- Berjak, P.; Pammenter, N.W. Implications of the Lack of Desiccation Tolerance in Recalcitrant Seeds. Front. Plant Sci. 2013, 4, 478. [Google Scholar] [CrossRef] [Green Version]
- Wawrzyniak, M.K.; Kalemba, E.M.; Ratajczak, E.; Chmielarz, P. Oxidation Processes Related to Seed Storage and Seedling Growth of Malus sylvestris, Prunus avium and Prunus padus. PLoS ONE 2020, 15, e0234510. [Google Scholar] [CrossRef]
- Kalemba, E.M.; Suszka, J.; Ratajczak, E. The Role of Oxidative Stress in Determining the Level of Viability of Black Poplar (Populus nigra) Seeds Stored at Different Temperatures. Funct. Plant Biol. FPB 2015, 42, 630–642. [Google Scholar] [CrossRef] [Green Version]
- Doody, C.N.; O’Reilly, C. Drying and Soaking Pretreatments Affect Germination in Pedunculate Oak. Ann. For. Sci. 2008, 65, 509. [Google Scholar] [CrossRef] [Green Version]
- GOSLING, P.G. The Effect of Drying Quercus robur Acorns to Different Moisture Contents, Followed by Storage, Either with or without Imbibition. For. Int. J. For. Res. 1989, 62, 41–50. [Google Scholar] [CrossRef]
- Ozbingol, N. Increasing Acorn Moisture Content Followed by Freezing-Storage Enhances Germination in Pedunculate Oak. Forestry 2005, 78, 73–81. [Google Scholar] [CrossRef]
- Suszka, B.; Tylkowski, T. Storage of Acorns of the English Oak (Quercus robur L.) over 1–5 Winters. Arbor. Kornickie 1980, 25, 199–229. [Google Scholar]
- Tylkowski, T. Height Increment of 1-Year Shoots of the English Oak (Quercus robur L.) and the Northern Red Oak (Quercus borealis Michx = Q. rubra L.) from 4-Year-Old Roots of Seedlings Raised from Acorns Stored over 1–5 Winters. Arbor. Kórn. 1982, 27, 357–365. [Google Scholar]
- Chmielarz, P.; Michalak, M.; Pałucka, M.; Wasileńczyk, U. Successful Cryopreservation of Quercus robur Plumules. Plant Cell Rep. 2011, 30, 1405–1414. [Google Scholar] [CrossRef] [PubMed]
- Chmielarz, P.; Grenier-de March, G.; Boucaud, M.-T. Cryopreservation of Quercus robur L. Embryogenic Calli. Cryo Lett. 2005, 26, 349–356. [Google Scholar] [PubMed]
- Chmielarz, P. Frost resistance of Quercus robur L. acorns. In Treatment and Storage of Oak Seeds. Present Situation and Research; Mitt.Biol.Bundesanst.Land-Forstwirtsch: Berlin-Dahlem, Germany, 1997; pp. 76–81. [Google Scholar]
- Ballesteros, D.; Hill, L.M.; Lynch, R.T.; Pritchard, H.W.; Walters, C. Longevity of Preserved Germplasm: The Temperature Dependency of Aging Reactions in Glassy Matrices of Dried Fern Spores. Plant Cell Physiol. 2019, 60, 376–392. [Google Scholar] [CrossRef]
- Taylor, A.G. Seed Storage, Germination, Quality, and Enhancements. In Physiology of Vegetable Crops, 2nd ed.; Wien, H.C., Stuetzel, H., Eds.; CAB International: Wallingford, UK, 2020; p. 496. ISBN 978-1786393777. [Google Scholar]
- Petrovic, S.; Sobajic, S.; Rakic, S.; Tomic, A.; Kukic, J. Investigation of Kernel Oils of Quercus robur and Quercus cerris. Chem. Nat. Compd. 2004, 40, 420–422. [Google Scholar] [CrossRef]
- Ballesteros, D.; Pritchard, H.W.; Walters, C. Dry Architecture: Towards the Understanding of the Variation of Longevity in Desiccation-Tolerant Germplasm. Seed Sci. Res. 2020, 30, 142–155. [Google Scholar] [CrossRef]
- Xia, K.; Hill, L.M.; Li, D.-Z.; Walters, C. Factors Affecting Stress Tolerance in Recalcitrant Embryonic Axes from Seeds of Four Quercus (Fagaceae) Species Native to the USA or China. Ann. Bot. 2014, 114, 1747–1759. [Google Scholar] [CrossRef]
- Hendry, G.A.F. Oxygen, Free Radical Processes and Seed Longevity. Seed Sci. Res. 1993, 3, 141–153. [Google Scholar] [CrossRef]
- Varghese, B.; Sershen, N.; Berjak, P.; Varghese, D.; Pammenter, N.W. Differential Drying Rates of Recalcitrant Trichilia dregeana Embryonic Axes: A Study of Survival and Oxidative Stress Metabolism. Physiol. Plant. 2011, 142, 326–338. [Google Scholar] [CrossRef]
- Schopfer, P.; Plachy, C.; Frahry, G. Release of Reactive Oxygen Intermediates (Superoxide Radicals, Hydrogen Peroxide, and Hydroxyl Radicals) and Peroxidase in Germinating Radish Seeds Controlled by Light, Gibberellin, and Abscisic Acid. Plant Physiol. 2001, 125, 1591–1602. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.S.; Kim, J.W.; Cha, Y.-N.; Kim, C. A Quantitative Nitroblue Tetrazolium Assay for Determining Intracellular Superoxide Anion Production in Phagocytic Cells. J. Immunoassay Immunochem. 2006, 27, 31–44. [Google Scholar] [CrossRef]
- Giertych, M.J.; Suszka, J. Consequences of Cutting off Distal Ends of Cotyledons of Quercus robur Acorns before Sowing. Ann. For. Sci. 2011, 68, 433–442. [Google Scholar] [CrossRef] [Green Version]
- R: A Language and Environment for Statistical Computing. Available online: https://www.gbif.org/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed on 30 August 2021).
- Corrplot: Visualization of a Correlation Matrix Version 0.90 from CRAN. Available online: https://rdrr.io/cran/corrplot/ (accessed on 30 August 2021).
- Jagadish, S.V.K.; Way, D.A.; Sharkey, T.D. Plant Heat Stress: Concepts Directing Future Research. Plant Cell Environ. 2021, 44, 1992–2005. [Google Scholar] [CrossRef]
- Fan, Y.; Ma, C.; Huang, Z.; Abid, M.; Jiang, S.; Dai, T.; Zhang, W.; Ma, S.; Jiang, D.; Han, X. Heat Priming During Early Reproductive Stages Enhances Thermo-Tolerance to Post-Anthesis Heat Stress via Improving Photosynthesis and Plant Productivity in Winter Wheat (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 805. [Google Scholar] [CrossRef] [PubMed]
- Vierling, E. The Roles of Heat Shock Proteins in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 579–620. [Google Scholar] [CrossRef]
- Lin, M.; Chai, K.; Ko, S.; Kuang, L.; Lur, H.; Charng, Y. A Positive Feedback Loop between HEAT SHOCK PROTEIN101 and HEAT STRESS-ASSOCIATED 32-KD PROTEIN Modulates Long-Term Acquired Thermotolerance Illustrating Diverse Heat Stress Responses in Rice Varieties. Plant Physiol. 2014, 164, 2045–2053. [Google Scholar] [CrossRef] [PubMed]
- Charng, Y.-Y.; Liu, H.-C.; Liu, N.-Y.; Chi, W.-T.; Wang, C.-N.; Chang, S.-H.; Wang, T.-T. A Heat-Inducible Transcription Factor, HsfA2, Is Required for Extension of Acquired Thermotolerance in Arabidopsis. Plant Physiol. 2007, 143, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-H.; Alam, I.; Kim, Y.-G.; Sharmin, S.A.; Lee, K.-W.; Lee, S.-H.; Lee, B.-H. Overexpression of a Chloroplast-Localized Small Heat Shock Protein OsHSP26 Confers Enhanced Tolerance against Oxidative and Heat Stresses in Tall Fescue. Biotechnol. Lett. 2012, 34, 371–377. [Google Scholar] [CrossRef]
- Suzuki, N.; Katano, K. Coordination Between ROS Regulatory Systems and Other Pathways Under Heat Stress and Pathogen Attack. Front. Plant Sci. 2018, 9, 490. [Google Scholar] [CrossRef]
- Wang, L.; Ma, K.-B.; Lu, Z.-G.; Ren, S.-X.; Jiang, H.-R.; Cui, J.-W.; Chen, G.; Teng, N.-J.; Lam, H.-M.; Jin, B. Differential Physiological, Transcriptomic and Metabolomic Responses of Arabidopsis Leaves under Prolonged Warming and Heat Shock. BMC Plant Biol. 2020, 20, 86. [Google Scholar] [CrossRef]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Robert, U.; Etuk, S.; Ubong, A.; Umoren, G.; Agbasi, O.; Abdulrazzaq, Z. Thermal and Mechanical Properties of Fabricated Plaster of Paris Filled with Groundnut Seed Coat and Waste Newspaper Materials for Structural Application. Epitoanyag-J. Silic. Based Compos. Mater. 2020, 72, 72–78. [Google Scholar] [CrossRef]
- Yi, X.; Wang, Z.; Liu, C.; Liu, G.; Zhang, M. Acorn Cotyledons Are Larger than Their Seedlings’ Need: Evidence from Artificial Cutting Experiments. Sci. Rep. 2015, 5, 8112. [Google Scholar] [CrossRef]
- Finch-Savage, W.E. Embryo Water Status and Survival in the Recalcitrant Species Quercus robur L.: Evidence for a Critical Moisture Content. J. Exp. Bot. 1992, 43, 663–669. [Google Scholar] [CrossRef]
- Finch-Savage, W.E. Seed Development in the Recalcitrant Species Quercus robur L.: Germinability and Desiccation Tolerance. Seed Sci. Res. 1992, 2, 17–22. [Google Scholar] [CrossRef]
- Rajjou, L.; Lovigny, Y.; Groot, S.P.C.; Belghazi, M.; Job, C.; Job, D. Proteome-Wide Characterization of Seed Aging in Arabidopsis: A Comparison between Artificial and Natural Aging Protocols. Plant Physiol. 2008, 148, 620–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendall, E.; Mckersie, B. Free Radical and Freezing Injury to Cell Membranes of Winter Wheat. Physiol. Plant. 1989, 76, 86–94. [Google Scholar] [CrossRef]
- Devireddy, A.R.; Tschaplinski, T.J.; Tuskan, G.A.; Muchero, W.; Chen, J.-G. Role of Reactive Oxygen Species and Hormones in Plant Responses to Temperature Changes. Int. J. Mol. Sci. 2021, 22, 8843. [Google Scholar] [CrossRef]
- Richards, S.L.; Wilkins, K.A.; Swarbreck, S.M.; Anderson, A.A.; Habib, N.; Smith, A.G.; McAinsh, M.; Davies, J.M. The Hydroxyl Radical in Plants: From Seed to Seed. J. Exp. Bot. 2015, 66, 37–46. [Google Scholar] [CrossRef]
- Laohavisit, A.; Richards, S.L.; Shabala, L.; Chen, C.; Colaço, R.D.D.R.; Swarbreck, S.M.; Shaw, E.; Dark, A.; Shabala, S.; Shang, Z.; et al. Salinity-Induced Calcium Signaling and Root Adaptation in Arabidopsis Require the Calcium Regulatory Protein Annexin1. Plant Physiol. 2013, 163, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Okuda, T.; Matsuda, Y.; Sagisaka, S. Formation of Hydroxyl Radicals in the Stems of Winter Wheat Treated with Freezing Temperature. Biosci. Biotechnol. Biochem. 1994, 58, 1189–1190. [Google Scholar] [CrossRef]
- Chmielarz, P.; Suszka, J. Wawrzyniak MK Desiccation Sensitivity and Frost Resistance of Oak (Quercus robur L.) Seeds. Ann. For. Sci. 2021. (under review). [Google Scholar]
- Baek, K.-H. Production of Reactive Oxygen Species by Freezing Stress and the Protective Roles of Antioxidant Enzymes in Plants. J. Agric. Chem. Environ. 2012, 01, 34–40. [Google Scholar] [CrossRef]
- Berwal, M.K.; Ram, C. Superoxide Dismutase: A Stable Biochemical Marker for Abiotic Stress Tolerance in Higher Plants. In Abiotic and Biotic Stress in Plants; De Oliveira, A., Ed.; IntechOpen: London, UK, 2018; ISBN 978-1-78923-812-9. [Google Scholar]
- Leggatt, C.W. Catalase Activity as a Measure of Seed Viability. Master’s Thesis, University of Alberta, 1928. [Google Scholar]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Van Breusegem, F.; Noctor, G. Catalase Function in Plants: A Focus on Arabidopsis Mutants as Stress-Mimic Models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [Green Version]
- Palma, J.M.; Gupta, D.K.; Corpas, F.J. Hydrogen Peroxide and Nitric Oxide Generation in Plant Cells: Overview and Queries. In Nitric Oxide and Hydrogen Peroxide Signaling in Higher Plants; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–16. ISBN 978-3-030-11129-8. [Google Scholar]
- Palma, J.M.; Mateos, R.M.; López-Jaramillo, J.; Rodríguez-Ruiz, M.; González-Gordo, S.; Lechuga-Sancho, A.M.; Corpas, F.J. Plant Catalases as NO and H2S Targets. Redox Biol. 2020, 34, 101525. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, A.; Steinger, T.; Müller-Schärer, H. The Importance of Plant Provenance and Genotypic Diversity of Seed Material Used for Ecological Restoration. Restor. Ecol. 2010, 18, 338–348. [Google Scholar] [CrossRef]
- Bharuth, V.; Naidoo, C.; Pammenter, N.W.; Lamb, J.M.; Moodley, T. Responses to Chilling of Recalcitrant Seeds of Ekebergia Capensis from Different Provenances. S. Afr. J. Bot. 2020, 130, 8–24. [Google Scholar] [CrossRef]
- Atkinson, R.J.; Thomas, E.; Roscioli, F.; Cornelius, J.P.; Zamora-Cristales, R.; Franco Chuaire, M.; Alcázar, C.; Mesén, F.; Lopez, H.; Ipinza, R.; et al. Seeding Resilient Restoration: An Indicator System for the Analysis of Tree Seed Systems. Diversity 2021, 13, 367. [Google Scholar] [CrossRef]
- Buras, A.; Sass-Klaassen, U.; Verbeek, I.; Copini, P. Provenance Selection and Site Conditions Determine Growth Performance of Pedunculate Oak. Dendrochronologia 2020, 61, 125705. [Google Scholar] [CrossRef]
- Arend, M.; Kuster, T.; Günthardt-Goerg, M.S.; Dobbertin, M. Provenance-Specific Growth Responses to Drought and Air Warming in Three European Oak Species (Quercus robur, Q. petraea and Q. pubescens). Tree Physiol. 2011, 31, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Matthews, S.; Powell, A.A.; Vigour, S. Electrical Conductivity Vigour Test: Physiological Basis and Use. Seed Test. Int. 2006, 131, 32–35. [Google Scholar]
- Gupta, R.; Deswal, R. Antifreeze Proteins Enable Plants to Survive in Freezing Conditions. J. Biosci. 2014, 39, 931–944. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalemba, E.M.; Wawrzyniak, M.K.; Suszka, J.; Chmielarz, P. Thermotherapy and Storage Temperature Manipulations Limit the Production of Reactive Oxygen Species in Stored Pedunculate Oak Acorns. Forests 2021, 12, 1338. https://doi.org/10.3390/f12101338
Kalemba EM, Wawrzyniak MK, Suszka J, Chmielarz P. Thermotherapy and Storage Temperature Manipulations Limit the Production of Reactive Oxygen Species in Stored Pedunculate Oak Acorns. Forests. 2021; 12(10):1338. https://doi.org/10.3390/f12101338
Chicago/Turabian StyleKalemba, Ewa Marzena, Mikołaj Krzysztof Wawrzyniak, Jan Suszka, and Paweł Chmielarz. 2021. "Thermotherapy and Storage Temperature Manipulations Limit the Production of Reactive Oxygen Species in Stored Pedunculate Oak Acorns" Forests 12, no. 10: 1338. https://doi.org/10.3390/f12101338
APA StyleKalemba, E. M., Wawrzyniak, M. K., Suszka, J., & Chmielarz, P. (2021). Thermotherapy and Storage Temperature Manipulations Limit the Production of Reactive Oxygen Species in Stored Pedunculate Oak Acorns. Forests, 12(10), 1338. https://doi.org/10.3390/f12101338