
Decision Support Tool for Tree Species Selection in Forest Regeneration Based on Harvester Data

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Stands
2.2. Estimation of Localized Growth
2.3. Creation of Micro-Stands
2.4. Simulation of Future Stand Development
3. Results
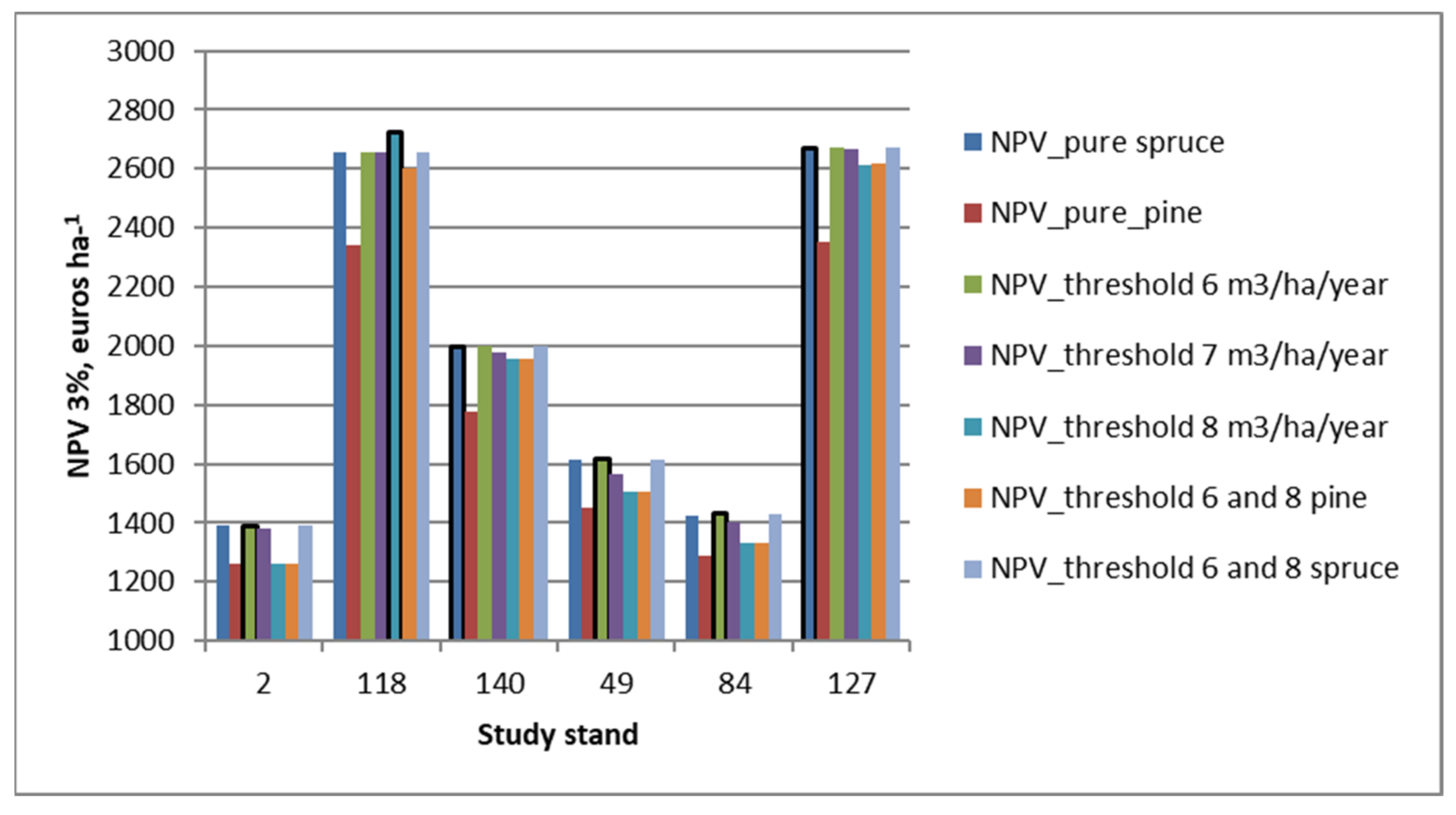
3.1. Comparison of Different Micro-Stand Solutions
3.2. Economic Effects Related to Micro-Stand-Level Forest Management
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kovacsova, P.; Antalova, M. Precision Forestry—Definition and Technologies. Sumar. List 2010, 134, 603–611. [Google Scholar]
- Ackerman, P.A.; Norihiro, J.; Ham, H.; Brewer, J.C. Producing More from Less. Towards optimising value in the bio-economy from data driven decisions. In Proceedings of the 4th Precision Forestry Symposium, Stellenbosch University, Stellenbosch, South Africa, 28 February–2 March 2017. [Google Scholar]
- Zhang, N.Q.; Wang, M.H.; Wang, N. Precision agriculture—A worldwide overview. Comput. Electron. Agric. 2002, 36, 113–132. [Google Scholar] [CrossRef]
- Georgi, C.; Spengler, D.; Itzerott, S.; Kleinschmit, B. Automatic delineation algorithm for site-specific management zones based on satellite remote sensing data. Precis. Agric. 2018, 19, 684–707. [Google Scholar] [CrossRef] [Green Version]
- Tompalski, P.; Coops, N.C.; White, J.C.; Wulder, M.A. Enhancing Forest Growth and Yield Predictions with Airborne Laser Scanning Data: Increasing Spatial Detail and Optimizing Yield Curve Selection through Template Matching. Forests 2016, 7, 255. [Google Scholar] [CrossRef] [Green Version]
- Mohamedou, C.; Tokola, T.; Eerikäinen, K. LiDAR-based TWI and terrain attributes in improving parametric predictor for tree growth in southeast Finland. Int. J. Appl. Earth Obs. Geoinf. 2017, 62, 183–191, ISSN 0303-2434. [Google Scholar] [CrossRef]
- Packalén, P.; Suvanto, A.; Maltamo, M. A Two Stage Method to Estimate Species-specific Growing Stock. Photogramm. Eng. Remote. Sens. 2009, 75, 1451–1460. [Google Scholar] [CrossRef]
- Coops, N.C. Characterizing Forest Growth and Productivity Using Remotely Sensed Data. Curr. For. Rep. 2015, 1, 195–205. [Google Scholar] [CrossRef]
- Kukkonen, M.; Kotivuori, E.; Maltamo, M.; Korhonen, L.; Packalen, P. Volumes by tree species can be predicted using photogrammetric UAS data, Sentinel-2 images and prior field measurements. Silva Fenn. 2021, 55. [Google Scholar] [CrossRef]
- Söderberg, J. A Method for Using Harvester Data in Airborne Laser Prediction of Forest Variables in Mature Coniferous Stands. Master’s Thesis, Dept. of Forest Resource Management, University of Agricultural Sciences, Umeå, Sweden, 2015. [Google Scholar]
- Holmgren, J.; Barth, A.; Larsson, H.; Olsson, H. 2012. Prediction of stem attributes by combining airborne laser scanning and measurements from harvesters. Silva Fenn. 2012, 46, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Möller, J.J.; Bhuiyan, N.; Arlinger, J.; Hannarup, B.; Nordström, M. Estimating thinning results based on standardised harvester data. In Proceedings of the 4th Precision Forestry Symposium, Stellenbosch University, Stellenbosch, South Africa, 28 February–2 March 2017. [Google Scholar]
- Olivera, A.; Visser, R. Development of forest-yield maps generated from Global Navigation Satellite System (GNSS)-enabled harvester StanForD files: Preliminary concepts. N. Z. J. For. Sci. 2016, 46. [Google Scholar] [CrossRef] [Green Version]
- Cajander, A.K. The theory of forest types. Acta For. Fenn. 1926, 29, 108. [Google Scholar] [CrossRef]
- Skogforsk. Available online: https://www.skogforsk.se/english/projects/stanford/ (accessed on 15 December 2019).
- Varjo, J. Latvan hukkapuun pituusmallit männylle, kuuselle ja koivulle metsurimittausta varten. [Models of top part of pine, spruce and birch for lumberjack measurement]. Puutavaran mittauksen kehittämistutkimuksia 1989–93. Metsäntutkimuslaitoksen Tiedonantoja 1995, 558, 69. [Google Scholar]
- Gustavsen, H.G. Talousmetsien kasvupaikkaluokittelu valtapituuden avulla. [Site index curves for conifer stands in Finland]. Folia For. 1980, 454, 1–31. [Google Scholar]
- Vuokila, Y.; Väliaho, H. Viljeltyjen havumetsiköiden kasvatusmallit. [Growth and yield models for conifer cultures in Finland]. Commun. Inst. For. Fenn. 1980, 99, 1–48. [Google Scholar]
- Salminen, H.; Lehtonen, M.; Hynynen, J. Reusing legacy FORTRAN in the MOTTI growth and yield simulator. Comput. Electron. Agric. 2005, 9, 103–113. [Google Scholar] [CrossRef]
- Siipilehto, J.; Ojansuu, R.; Miina, J.; Hynynen, J.; Valkonen, S.; Saksa, T. Metsikön Varhaiskehityksen Kuvaus MOTTI-Ohjelmistossa [Early Development of Young Stands in Motti Software]. Metlan Työraportteja 2014, 286. Available online: http://www.metla.fi/julkaisut/workingpapers/2014/mwp286.pdf (accessed on 15 December 2019). (In Finnish)
- Hynynen, J.; Salminen, H.; Ahtikoski, A.; Huuskonen, S.; Ojansuu, R.; Siipilehto, J.; Lehtonen, M.; Eerikäinen, K. Long-term impacts of forest management on biomass supply and forest resource development: A scenario analysis for Finland. Eur. J. For. Res. 2015, 134, 415–431. [Google Scholar] [CrossRef]
- Hynynen, J.; Salminen, H.; Ahtikoski, A.; Huuskonen, S.; Ojansuu, R.; Siipilehto, J.; Lehtonen, M.; Rummukainen, A.; Kojola, S.; Eerikäinen, K. Scenario Analysis for the Biomass Supply Potential and the Future Development of Finnish Forest Resources, Working Papers of the Finnish Forest Research Institute 302. Metla 2014. Available online: http://www.metla.fi/julkaisut/workingpapers/2014/mwp302-en.htm (accessed on 15 December 2019).
- Hynynen, J.; Ojansuu, R.; Hökkä, H.; Siipilehto, J.; Salminen, H.; Haapala, P. Models for predicting stand development in MELA System. Finn. For. Res. Inst. Res. Pap. 835 2002, 1–116. [Google Scholar]
- Huuskonen, S.; Haikarainen, S.; Sauvula-Seppälä, T.; Salminen, H.; Lehtonen, M.; Siipilehto, J.; Ahtikoski, A.; Korhonen, K.T.; Hynynen, J. Benefits of juvenile stand management in Finland—Impacts on wood production based on scenario analysis. For. Int. J. For. Res. 2020, 93, 458–470. [Google Scholar] [CrossRef]
- Luke. Available online: https://statdb.luke.fi/PXWeb/pxweb/fi/LUKE/ (accessed on 15 December 2019).
- Raatevaara, A.; Korpunen, H.; Tiitta, M.; Tomppo, L.; Kulju, S.; Antikainen, J.; Uusitalo, J. Electrical impedance and image analysis methods in detecting and measuring Scots pine heartwood from a log end during tree harvesting. Comput. Electron. Agric. 2020, 177. [Google Scholar] [CrossRef]
- Lindroos, O.; Ringdahl, O.; La Hera, P.; Hohnloser, P.; Hellström, T. Estimating the Position of the Harvester Head—A Key Step towards the Precision Forestry of the Future? Croat. J. For. Eng. 2015, 36, 147–167. [Google Scholar]
- Mäkinen, H.; Hynynen, J.; Isomäki, A. Intensive management of Scots pine stands in southern Finland: First empirical results and simulated further development. For. Ecol. Manag. 2005, 215, 37–50. [Google Scholar] [CrossRef]
- Frivold, L.H.; Frank, J. Growth of mixed birch-coniferous stands in relation to pure coniferous stands at similar sites in south-eastern Norway. Scand. J. For. Res. 2002, 17, 139–149. [Google Scholar] [CrossRef]
- Hynynen, J.; Repola, J.; Mielikäinen, K. The effects of species mixture on the growth and yield of mid-rotation mixed stands of Scots pine and silver birch. For. Ecol. Manag. 2011, 262, 1174–1183. [Google Scholar] [CrossRef]
- Huuskonen, S.; Domisch, T.; Finér, L.; Hantula, J.; Hynynen, J.; Matala, J.; Miina, J.; Neuvonen, S.; Nevalainen, S.; Niemistö, P.; et al. What is the potential for replacing monocultures with mixed-species stands to enhance ecosystem services in boreal forests in Fennoscandia? For. Ecol. Manag. 2021, 479, 21. [Google Scholar] [CrossRef]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: From temperate to boreal forests. Glob. Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed]
- Felton, A.; Nilsson, U.; Sonesson, J.; Felton, A.M.; Roberge, J.-M.; Ranius, T.; Ahlström, M.; Bergh, J.; Björkman, C.; Boberg, J.; et al. Replacing monocultures with mixed-species stands: Ecosystem service implications of two production forest alternatives in Sweden. Ambio 2016, 45, 124–139. [Google Scholar] [CrossRef] [Green Version]
- Mina, M.; Huber, M.O.; Forrester, D.I.; Thüring, E.; Rohner, B. Multiple factors modulate tree growth complementarity in Central European mixed forests. J. Ecol. 2018, 106, 1106–1119. [Google Scholar] [CrossRef]
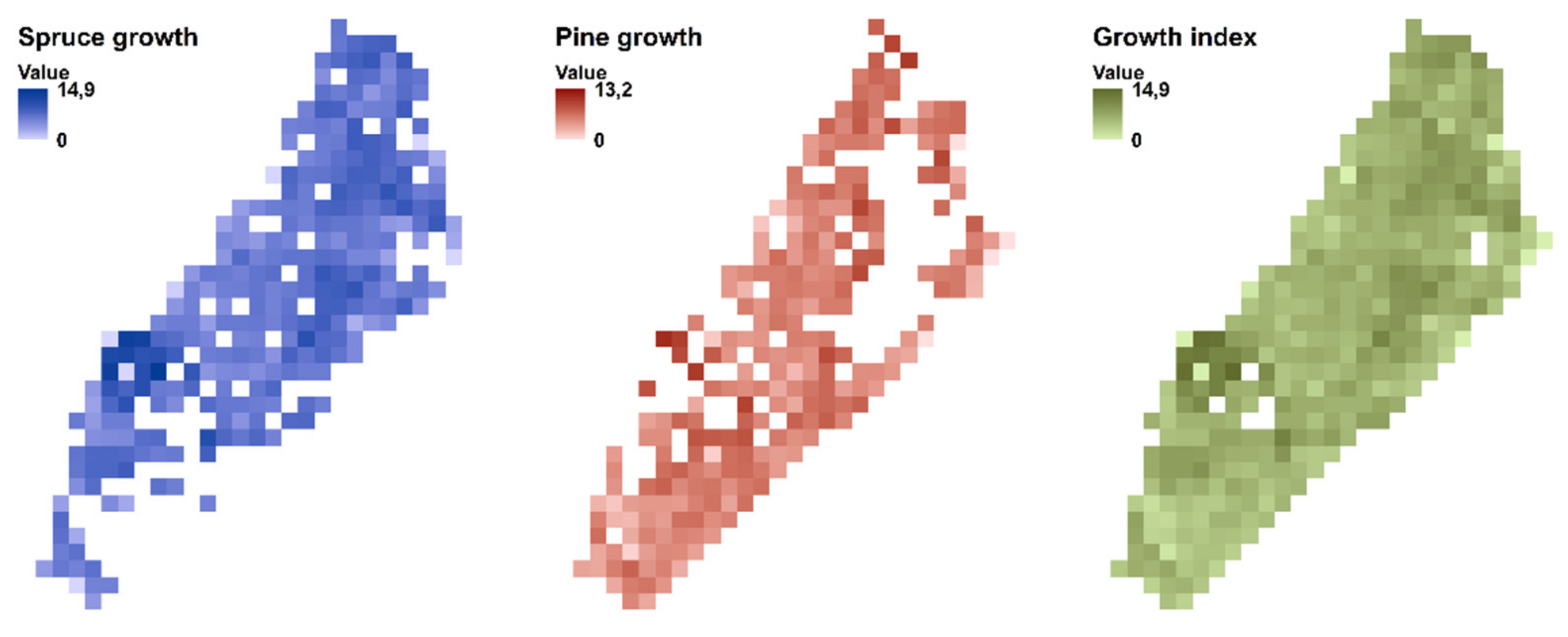


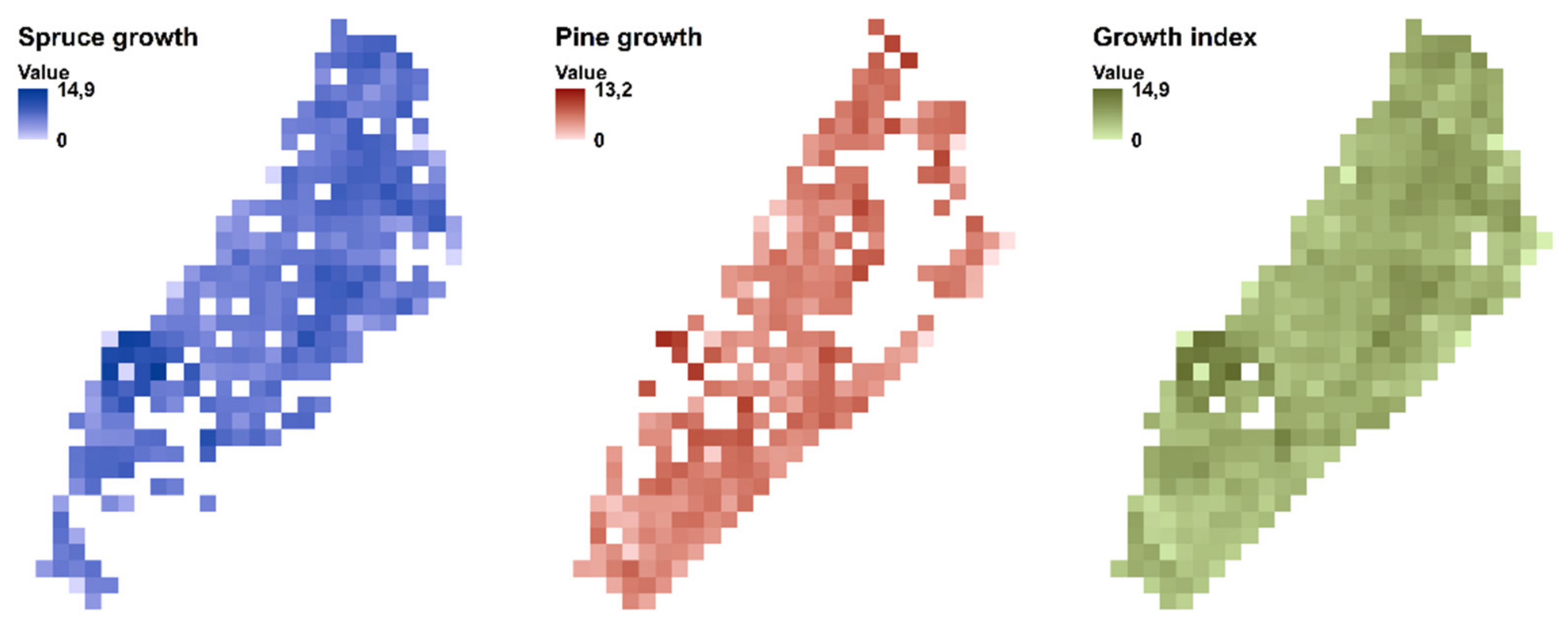{kind=link}
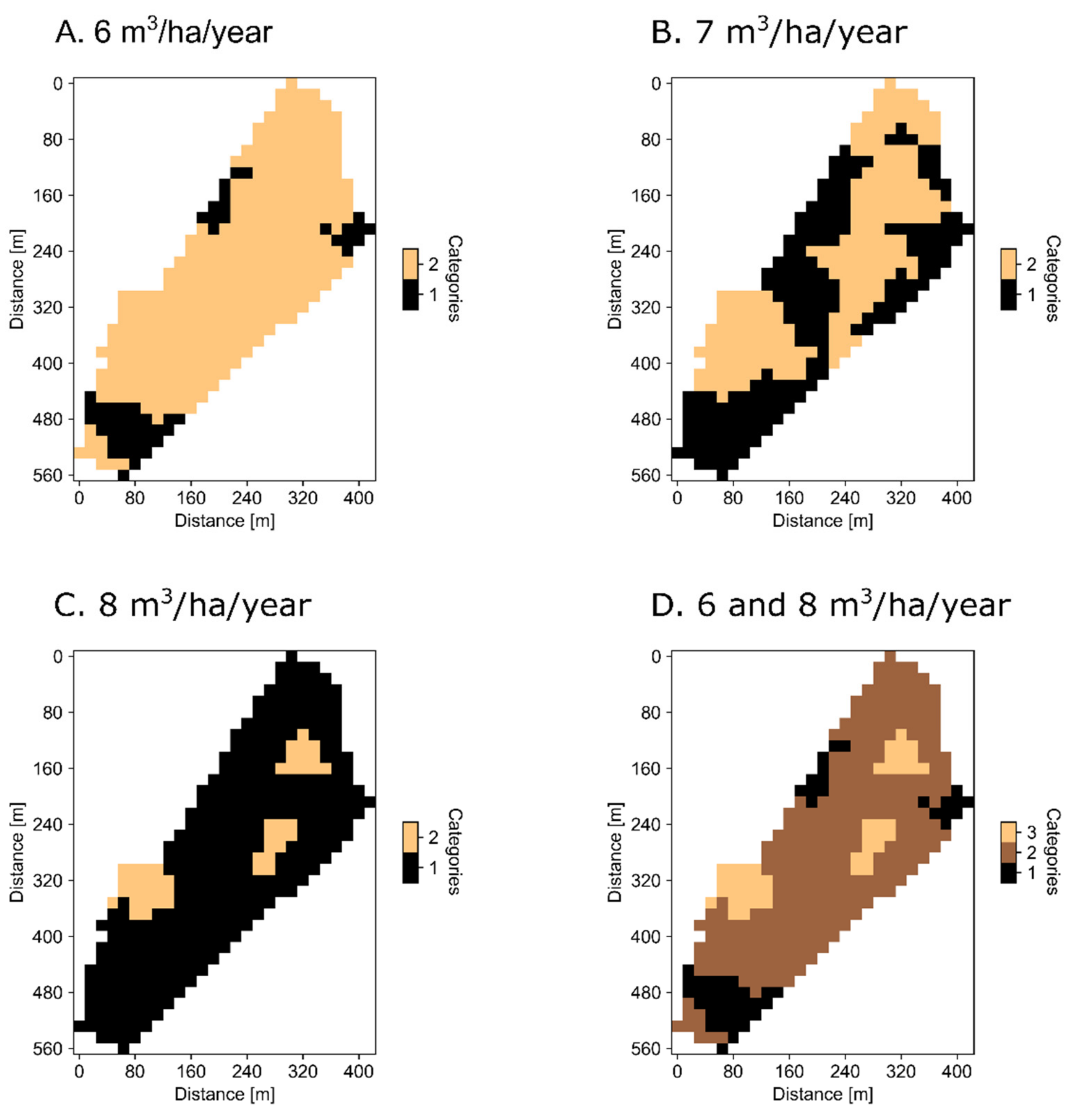
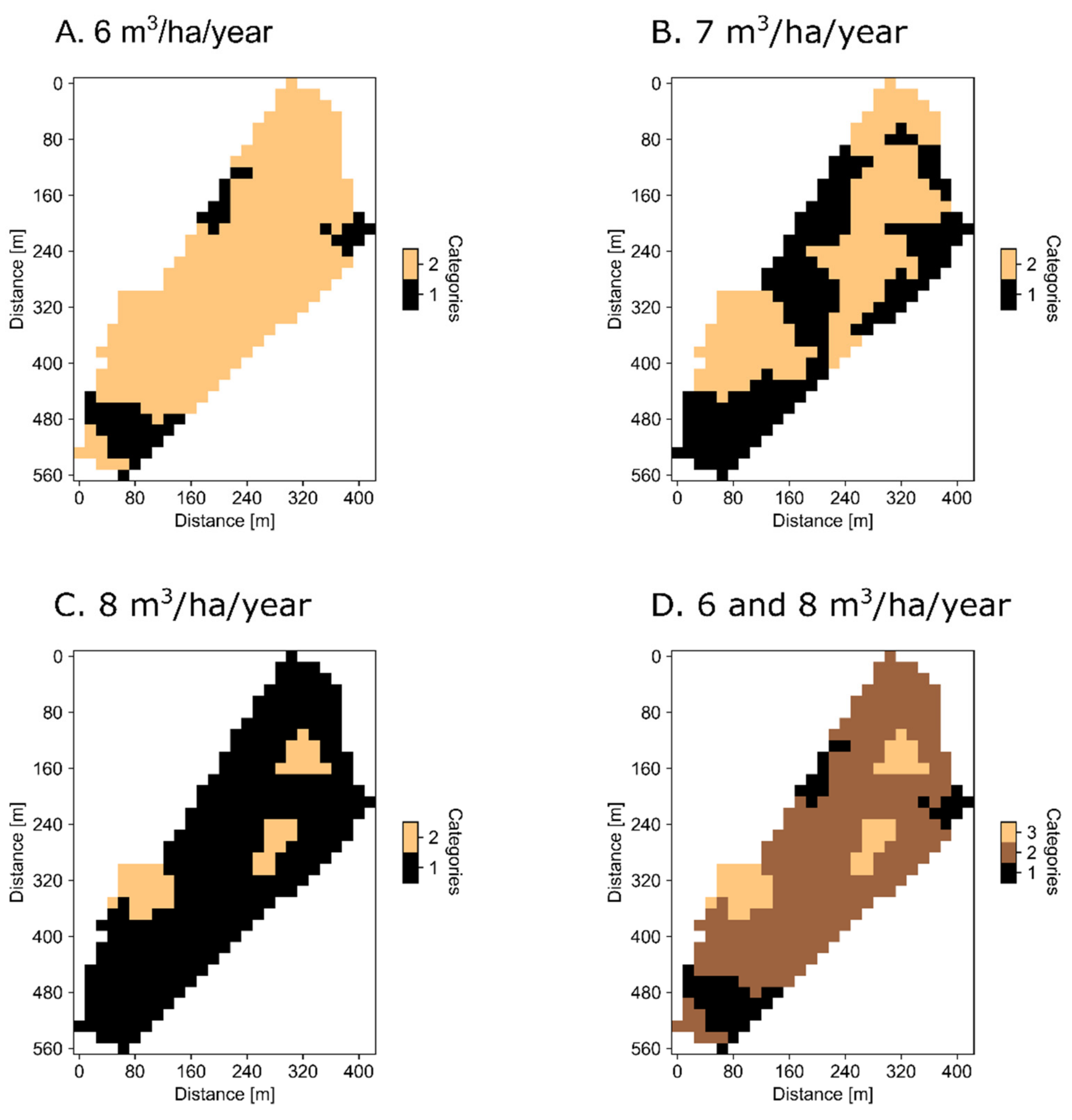{kind=link}
{kind=link}
| Stand Characteristics | |||||||
|---|---|---|---|---|---|---|---|
| Study Stand No. | Area, (ha) | Main Site Type | Mean Age (year) | Scots Pine, (m3/ha) | Norway, Spruce (m3/ha) | Birch, (m3/ha) | Total, (m3/ha) |
| 2 | 1.3 | MT * | 91 | 142 | 112 | 18 | 272 |
| 118 | 1.7 | MT | 67 | 93 | 121 | 31 | 245 |
| 140 | 5.1 | VT ** | 60 | 121 | 53 | 25 | 199 |
| 49 | 9.8 | MT | 78 | 65 | 106 | 12 | 183 |
| 84 | 12.8 | MT | 65 | 112 | 111 | 18 | 241 |
| 127 | 23.3 | MT | 65 | 182 | 113 | 8 | 303 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saksa, T.; Uusitalo, J.; Lindeman, H.; Häyrynen, E.; Kulju, S.; Huuskonen, S. Decision Support Tool for Tree Species Selection in Forest Regeneration Based on Harvester Data. Forests 2021, 12, 1329. https://doi.org/10.3390/f12101329
Saksa T, Uusitalo J, Lindeman H, Häyrynen E, Kulju S, Huuskonen S. Decision Support Tool for Tree Species Selection in Forest Regeneration Based on Harvester Data. Forests. 2021; 12(10):1329. https://doi.org/10.3390/f12101329
Chicago/Turabian StyleSaksa, Timo, Jori Uusitalo, Harri Lindeman, Esko Häyrynen, Sampo Kulju, and Saija Huuskonen. 2021. "Decision Support Tool for Tree Species Selection in Forest Regeneration Based on Harvester Data" Forests 12, no. 10: 1329. https://doi.org/10.3390/f12101329
APA StyleSaksa, T., Uusitalo, J., Lindeman, H., Häyrynen, E., Kulju, S., & Huuskonen, S. (2021). Decision Support Tool for Tree Species Selection in Forest Regeneration Based on Harvester Data. Forests, 12(10), 1329. https://doi.org/10.3390/f12101329