
On the Consequences of Using Moving Window Segmentation to Analyze the Structural Stand Heterogeneity and Debatable Patchiness of Old-Growth Temperate Forests


Abstract
1. Introduction
2. Materials and Methods
2.1. Patch Classification
2.2. Simulation Study
2.3. Randomness vs. Patchiness—Empirical Examples
3. Results
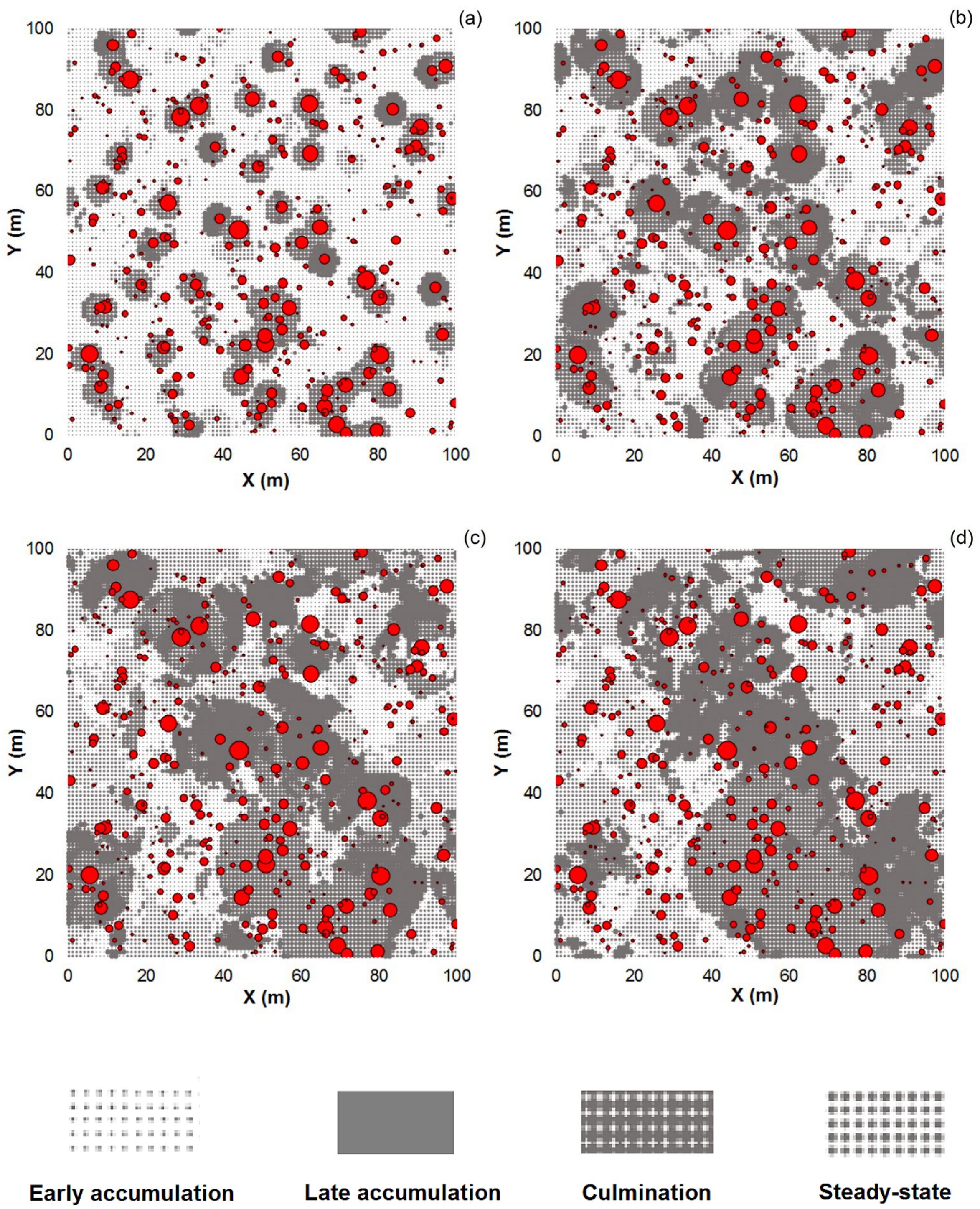
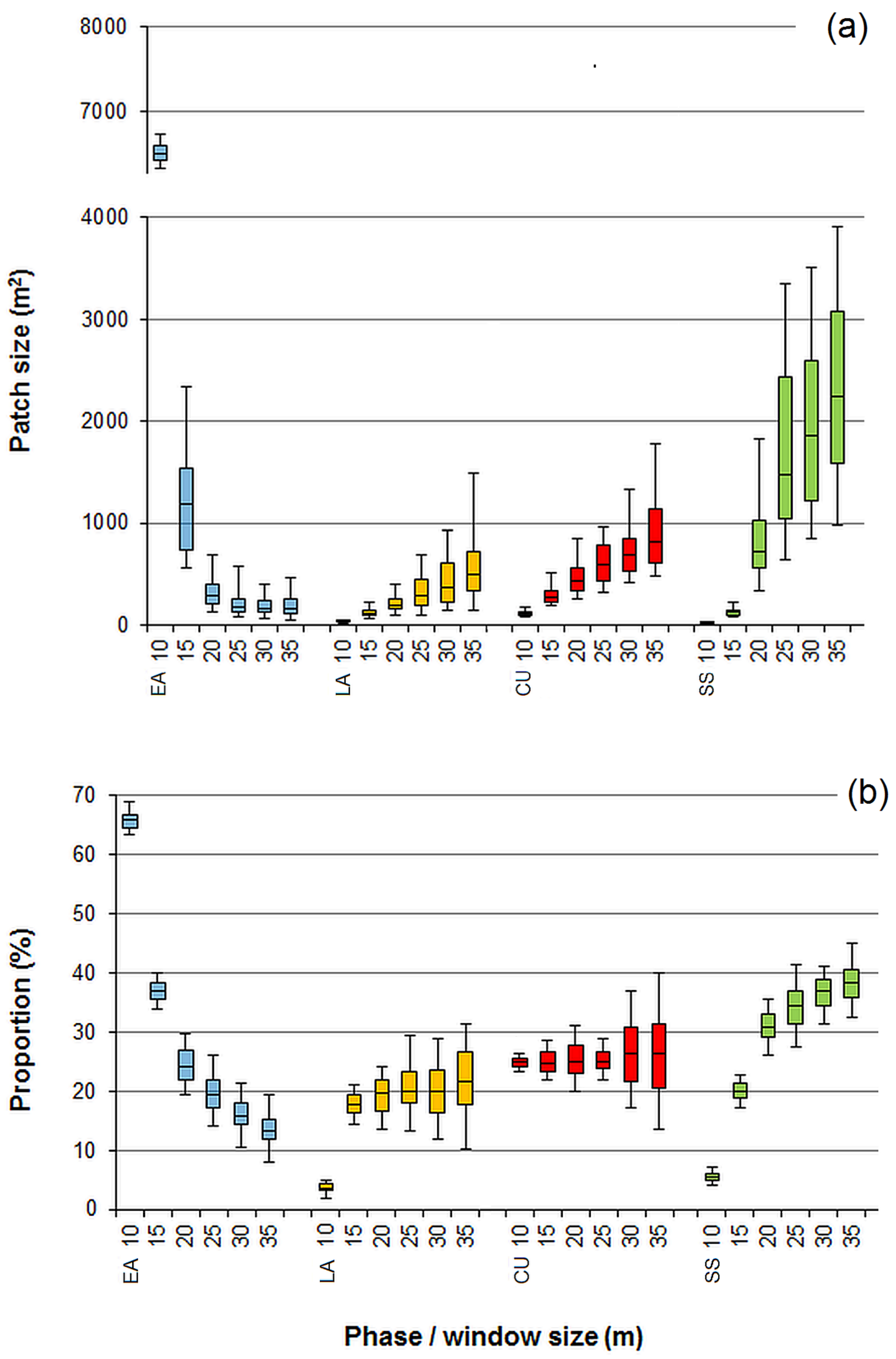
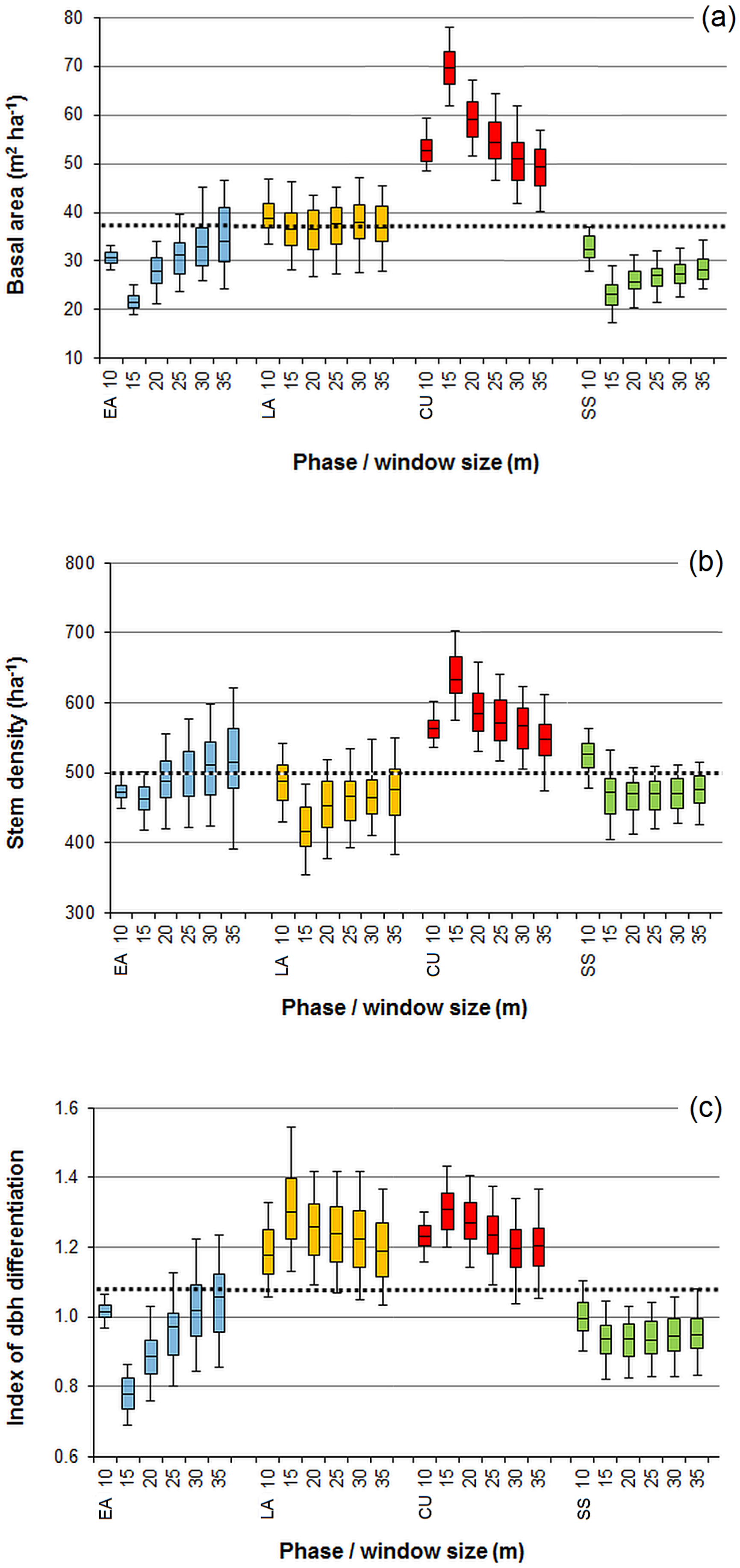
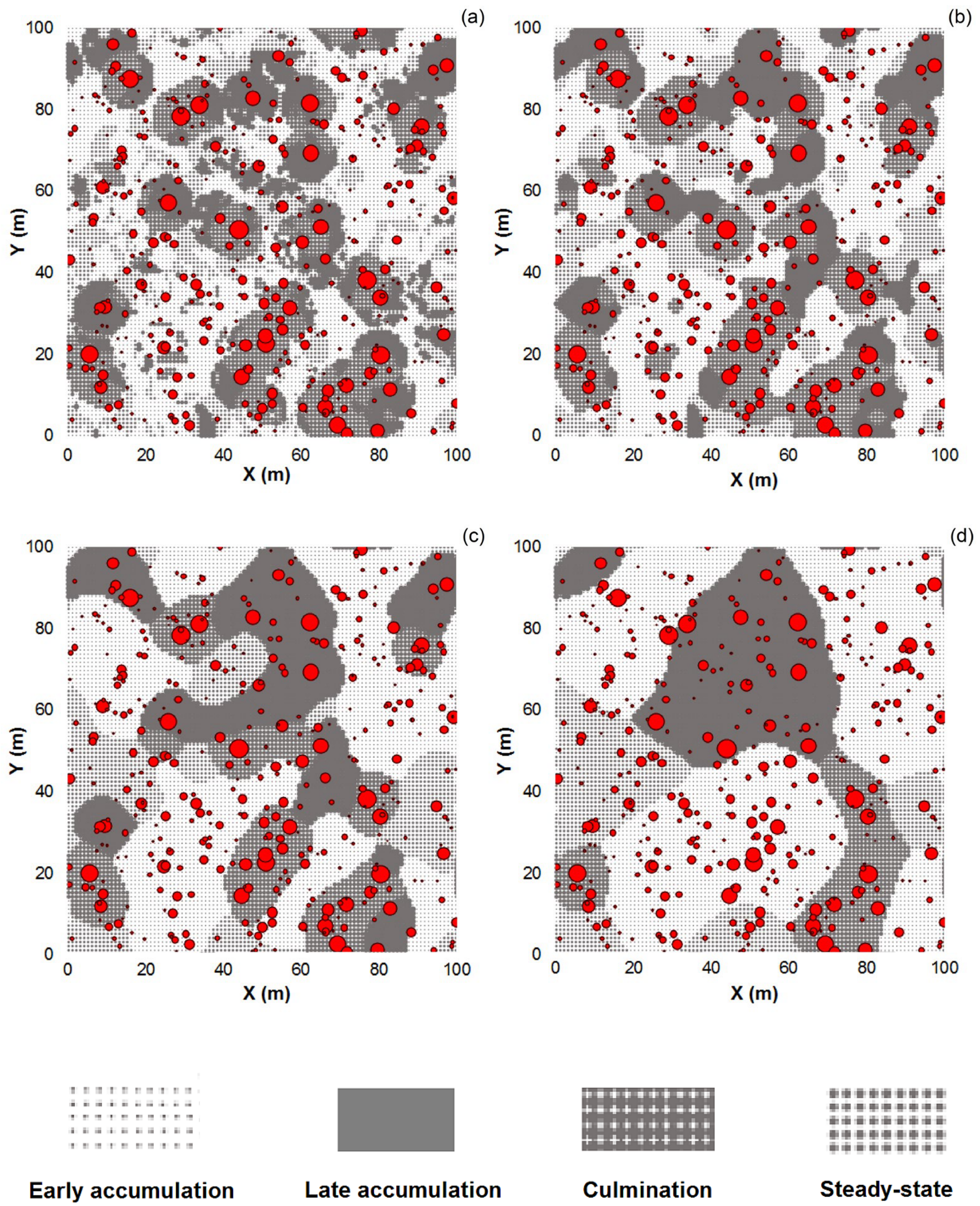
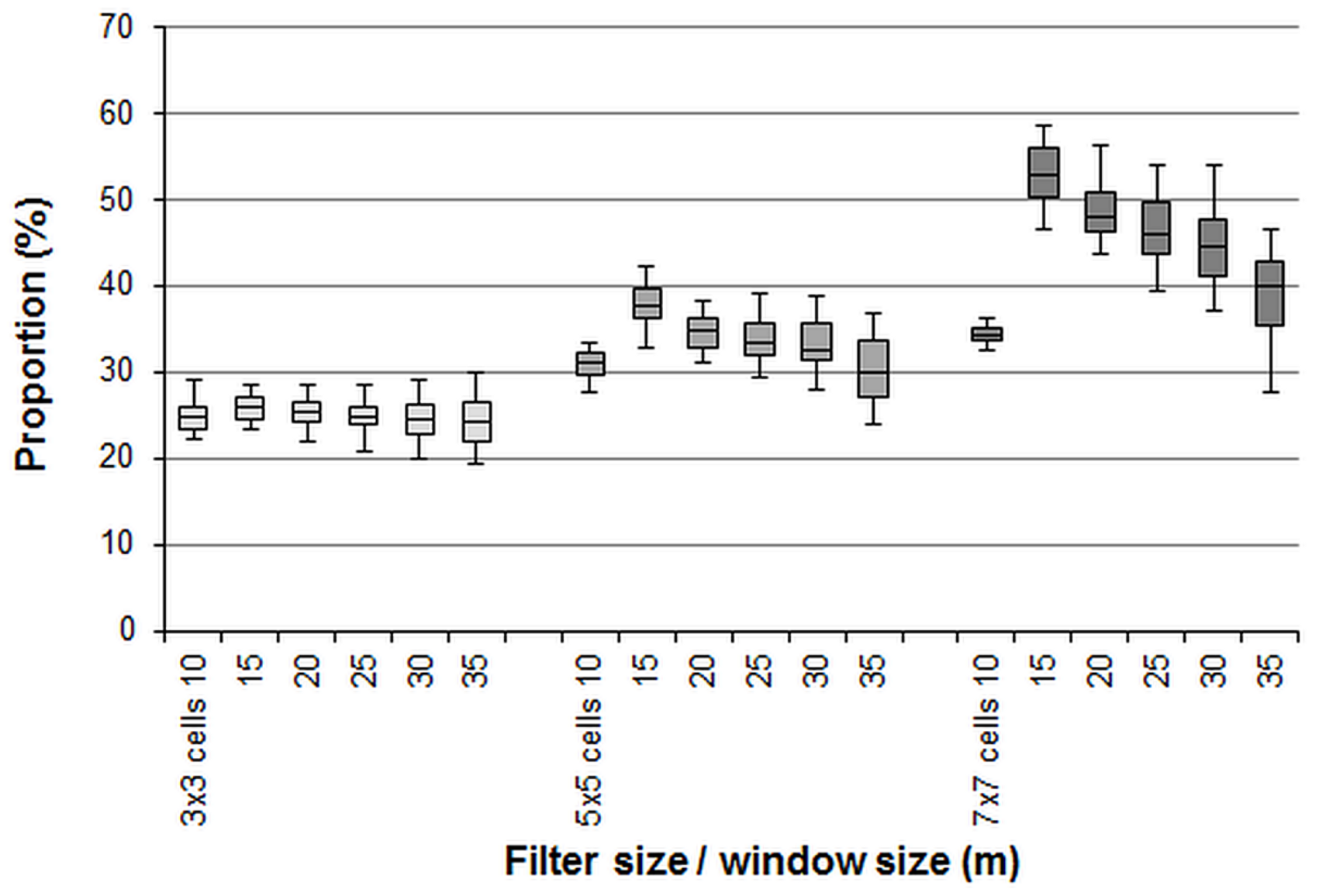
3.1. Effect of the Moving Window Size
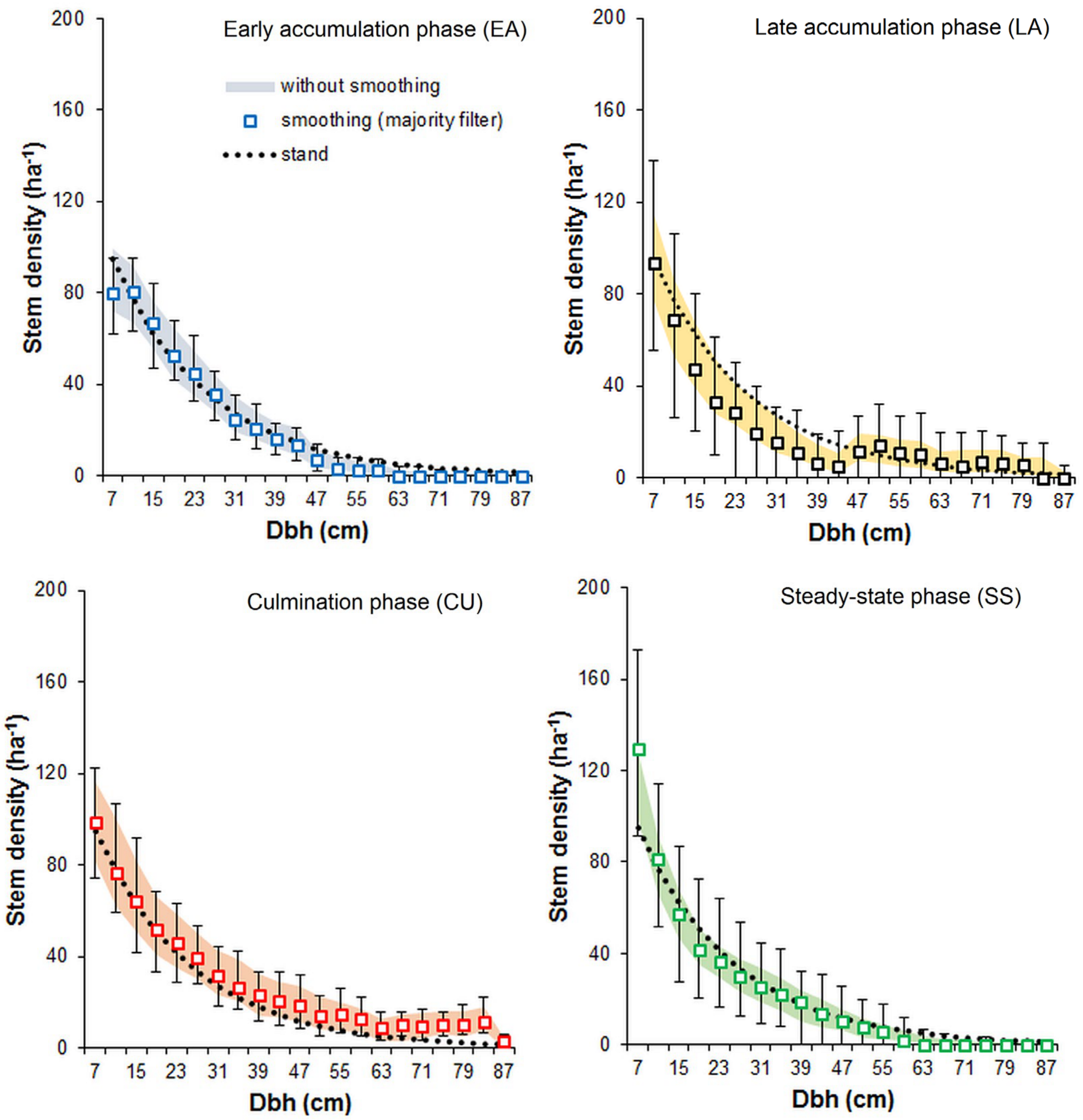
3.2. Effect of Noise Smoothing
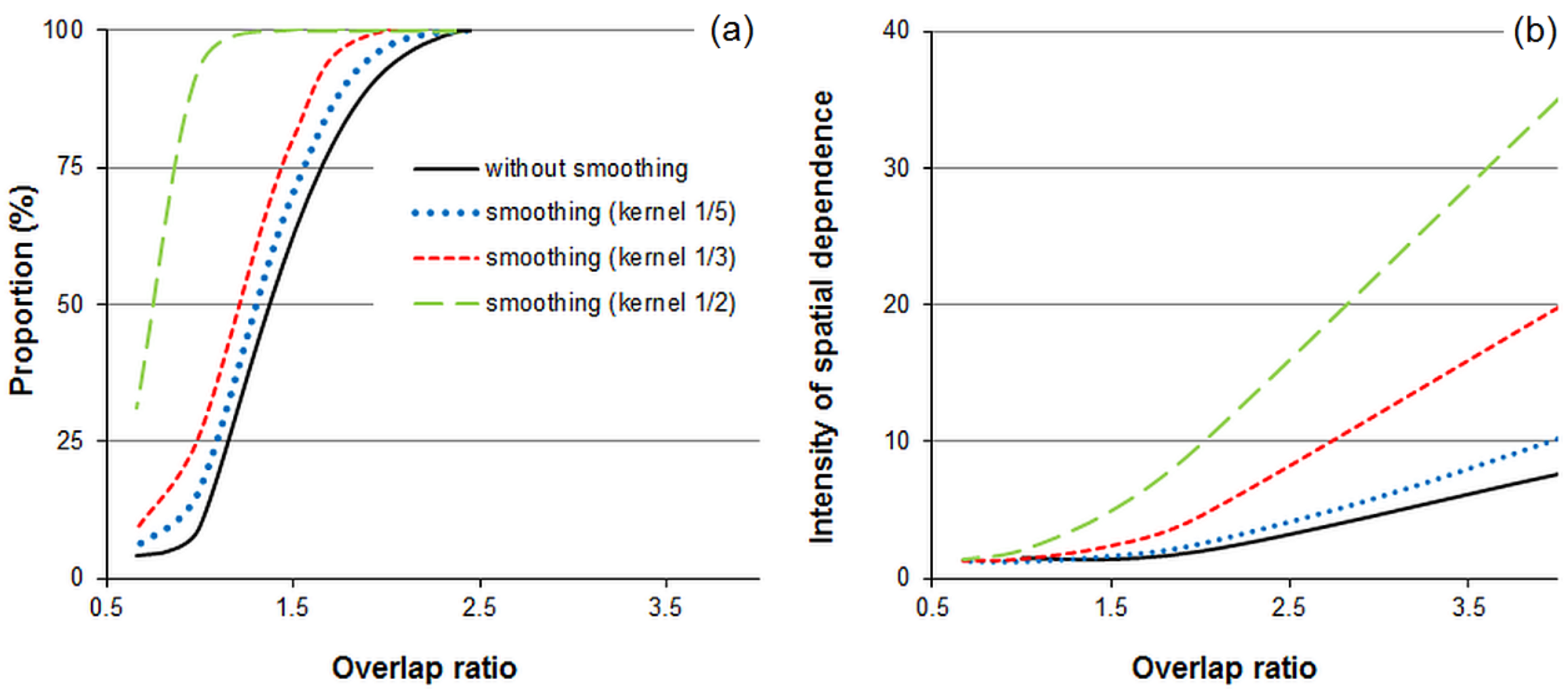
3.3. Effect of Overlapping Windows on the Spatial Dependence of the Phase Classification Outputs
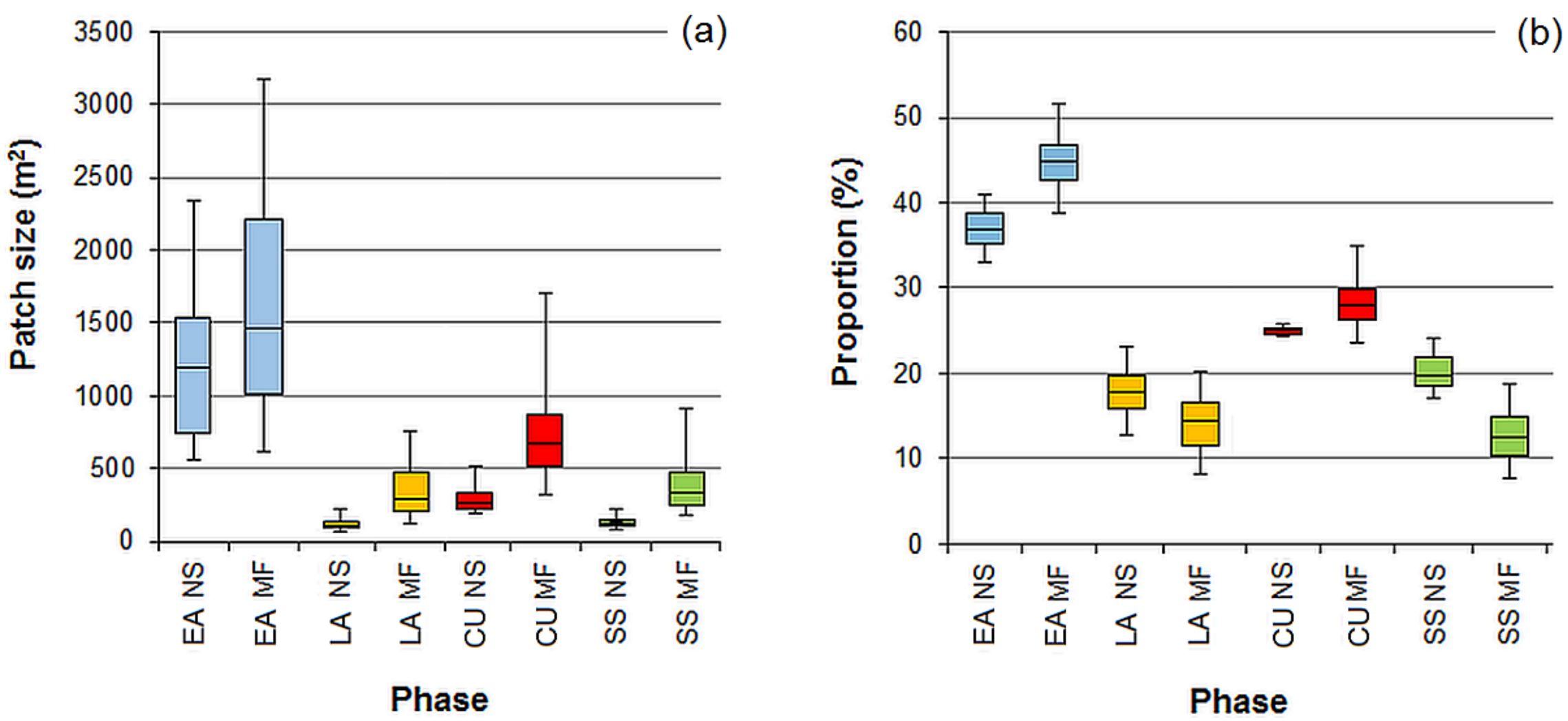
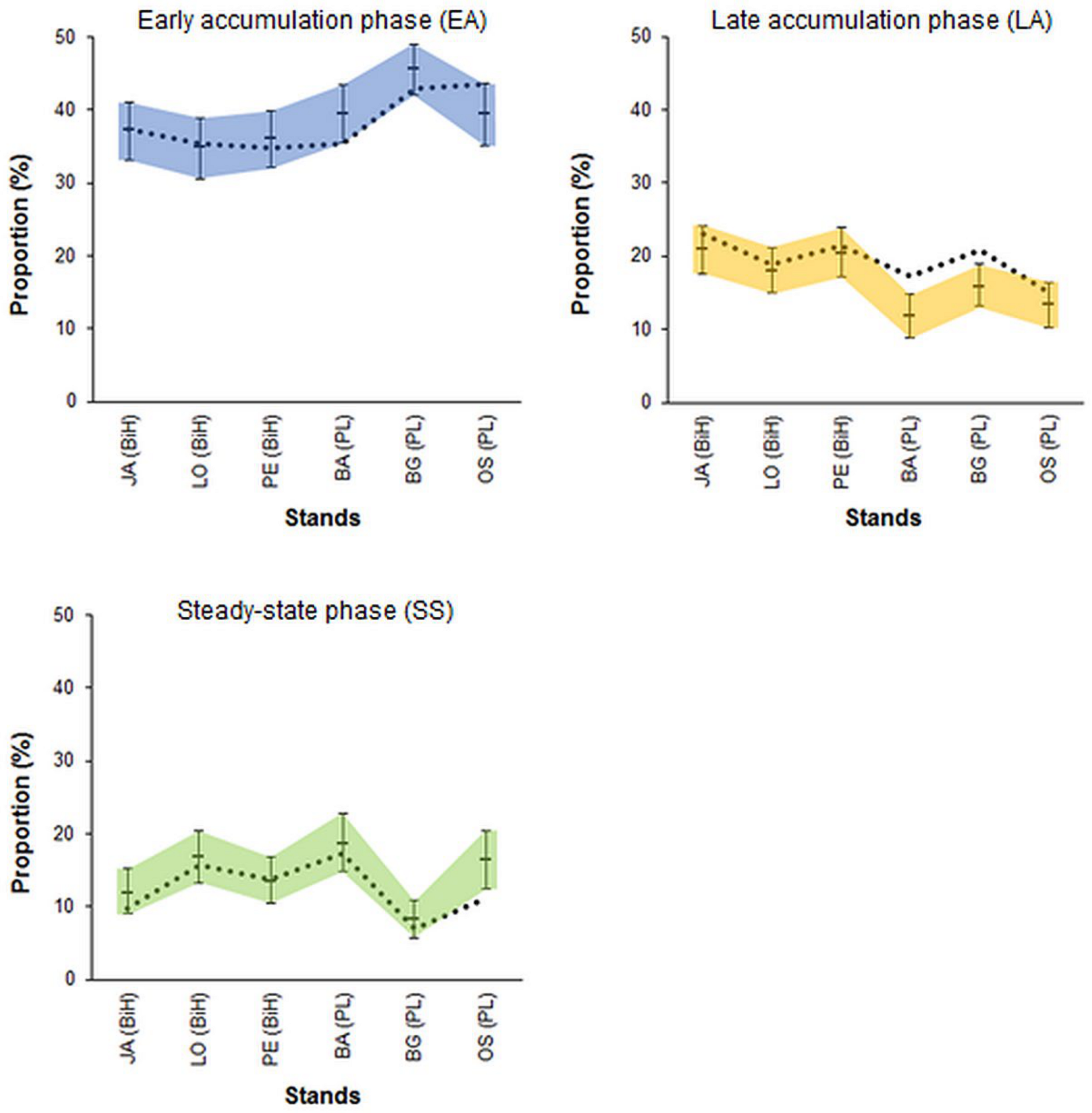
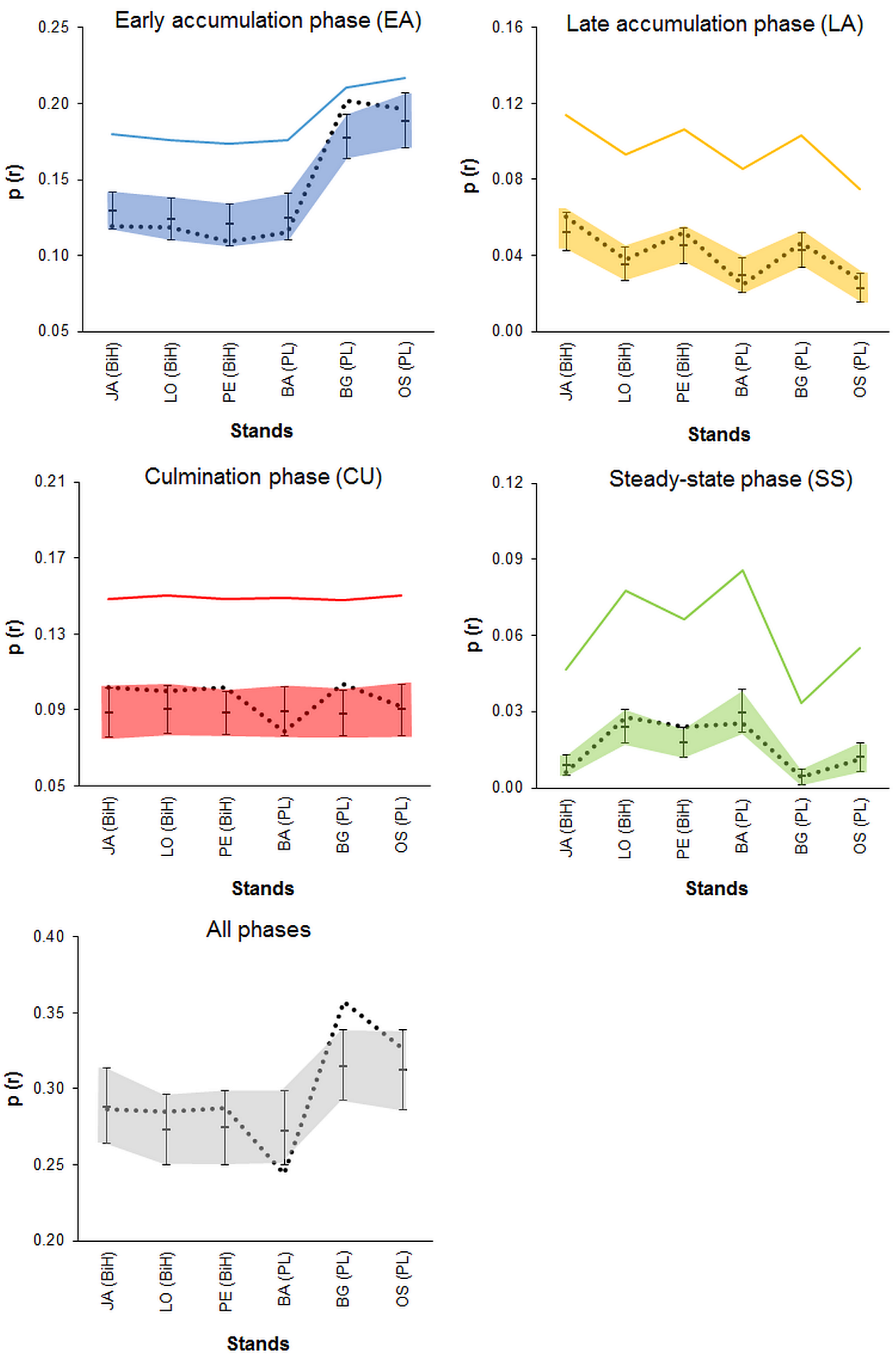
3.4. Randomness vs. Patchiness-Empirical Examples
4. Discussion
4.1. Remarks on the Phase Classification Protocol
4.2. Limitations of Using Moving Windows with Overlap for Patch Classification
4.3. Randomness vs. Patchiness-Empirical Examples
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, C.; Law, R.; Illian, J.B.; Burslem, D.F.R.P. Linking ecological processes with spatial and non-spatial patterns in plant communities. J. Ecol. 2011, 99, 1402–1414. [Google Scholar] [CrossRef]
- Illian, J.B.; Burslem, D.F.R.P. Improving the usability of spatial point process methodology: An interdisciplinary dialogue between statistics and ecology. AStA Adv. Stat. Anal. 2017, 101, 495–520. [Google Scholar] [CrossRef]
- Velázquez, E.; Martínez, I.; Getzin, S.; Moloney, K.A.; Wiegand, T. An evaluation of the state of spatial point pattern analysis in ecology. Ecography 2016, 39, 1042–1055. [Google Scholar] [CrossRef]
- Law, R.; Illian, J.B.; Burslem, D.F.R.P.; Gratzer, G.; Gunatilleke, C.V.S.; Gunatilleke, I.A.U.N. Ecological information from spatial patterns of plants: Insights from point process theory. J. Ecol. 2009, 97, 616–628. [Google Scholar] [CrossRef]
- Watt, A.S. Pattern and process in the plant community. J. Ecol. 1947, 35, 1–22. [Google Scholar] [CrossRef]
- Remmert, H. The Mosaic-Cycle Concept of Ecosystems; Remmert, H., Ed.; Springer Verlag: New York, NY, USA, 1991; pp. 1–21. [Google Scholar]
- Firm, D.; Nagel, T.A.; Diaci, J. Disturbance history and dynamics of an old-growth mixed species mountain forest in the Slovenian Alps. For. Ecol. Manag. 2009, 257, 1893–1901. [Google Scholar] [CrossRef]
- Hobi, M.L.; Commarmot, B.; Bugmann, H. Pattern and process in the largest primeval beech forest of Europe (Ukrainian Carpathians). J. Veg. Sci. 2015, 26, 323–336. [Google Scholar] [CrossRef]
- Feldmann, E.; Drößler, L.; Hauck, M.; Kucbel, S.; Pichler, V.; Leuschner, C. Canopy gap dynamics and tree understory release in a virgin beech forest, Slovakian Carpathians. For. Ecol. Manag. 2018, 415/416, 38–46. [Google Scholar] [CrossRef]
- Orman, O.; Dobrowolska, D. Gap dynamics in the Western Carpathian mixed beech old-growth forests affected by spruce bark beetle outbreak. Eur. J. For. Res. 2017, 136, 571–581. [Google Scholar] [CrossRef]
- Brang, P.; Spathelf, P.; Larsen, J.B.; Bauhus, J.; Boncina, A.; Chauvin, C.; Drössler, L.; Garcia-Guemes, C.; Heiri, C.; Kerr, G.; et al. Suitability of close-to-nature silviculture for adapting temperate European forests to climate change. Forestry 2014, 87, 492–503. [Google Scholar] [CrossRef]
- Petritan, I.C.; Commarmot, B.; Hobi, M.L.; Petritan, A.M.; Bigler, C.H.; Abrudan, I.V.; Rigling, A. Structural patterns of beech and silver fir suggest stability and resilience of the virgin forest Sinca in the Southern Carpathians, Romania. For. Ecol. Manag. 2015, 356, 184–195. [Google Scholar] [CrossRef]
- Janík, D.; Král, K.; Adam, D.; Hort, L.; Samonil, P.; Unar, P.; Vrška, T.; McMahon, S. Tree spatial patterns of Fagus sylvatica expansion over 37 years. For. Ecol. Manag. 2016, 375, 134–145. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Leibundgut, H. Europäische Urwälder; Paul Haupt: Bern, Switzerland, 1993. [Google Scholar]
- Korpel’, Š. Die Urwälder der Westkarpaten; Gustav Fisher Verlag: Stuttgart, Germany, 1995. [Google Scholar]
- Meyer, P. Bestimmung der Waldentwicklungsphasen und der Texturdiversität in Naturwäldern. Allg. Forst u. J. Ztg 1999, 170, 203–211. [Google Scholar]
- Emborg, J.; Christensen, M.; Heilmann-Clausen, J. The structural dynamics of Suserup Skov, a near-natural temperate deciduous forest in Denmark. For. Ecol. Manag. 2000, 126, 173–189. [Google Scholar] [CrossRef]
- Tabaku, V. Struktur von Buchen-Urwäldern in Albanien im Vergleich mit deutschen Buchen-Naturwaldreservaten und—Wirtschaftswäldern; Cuvillier Verlag: Göttingen, Germany, 2000. [Google Scholar]
- Drössler, L.; Meyer, P. Waldentwicklungsphasen in zwei Buchen-Urwaldreservaten in der Slowakei. Forstarchiv 2006, 77, 155–161. [Google Scholar]
- Podlaski, R. Dynamics in Central European near-natural Abies-Fagus forests: Does the mosaic-cycle approach provide an appropriate model? J. Veg. Sci. 2008, 19, 173–182. [Google Scholar] [CrossRef]
- Král, K.; Vrška, T.; Hort, L.; Adam, D.; Šamonil, P. Development phase in a temperate natural spruce-fir-beech forest: Determination by a supervised classification method. Eur. J. For. Res. 2010, 129, 339–351. [Google Scholar] [CrossRef]
- Winter, S.; Brambach, F. Determination of a common forest life cycle assessment method for biodiversity evaluation. For. Ecol. Manag. 2011, 262, 2120–2132. [Google Scholar] [CrossRef]
- Král, K.; Shue, J.; Vrška, T.; Gonzalez-Akre, E.B.; Parker, G.G.; McShea, W.J.; McMahon, S.M. Fine-scale patch mosaic of developmental stages in Northeast American secondary temperate forests: The European perspective. Eur. J. For. Res. 2016, 135, 981–996. [Google Scholar] [CrossRef]
- Rock, J.; Badeck, F.W.; Harmon, M.E. Estimating decomposition rate constants for European tree species from literature sources. Eur. J. For. Res. 2008, 127, 301–313. [Google Scholar] [CrossRef]
- Feldmann, E.; Glatthorn, J.; Hauck, M.; Leuschner, C. A novel empirical approach for determining the extension of forest development stages in temperate old-growth forests. Eur. J. For. Res. 2018, 137, 321–335. [Google Scholar] [CrossRef]
- Illian, J.; Penttinen, A.; Stoyan, H.; Stoyan, D. Statistical Analysis and Modelling of Spatial Point Patterns; John Wiley & Sons: West Sussex, UK, 2008. [Google Scholar]
- Diggle, P.J. Statistical Analysis of Spatial and Spatio-Temporal Point Patterns; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Baddeley, A.; Rubak, E.; Turner, R. Spatial Point Patterns: Methodology and Applications with R; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Flechter, R.; Fortin, M.F. Spatial Ecology and Conservation Modeling. Applications with R; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Szwagrzyk, J.; Czerwczak, M. Spatial patterns of trees in natural forests of East-Central Europe. J. Veg. Sci. 1993, 4, 469–476. [Google Scholar] [CrossRef]
- Von Oheimb, G.; Westphal, C.; Tempel, H.; Härdtle, W. Structural pattern of a near-natural beech forest (Fagus sylvatica) (Serrahn, north-east Germany). For. Ecol. Manag. 2005, 212, 253–263. [Google Scholar] [CrossRef]
- Paluch, J. The spatial pattern of a natural European beech (Fagus sylvatica L.)—Silver fir (Abies alba Mill.) forest: A patch-mosaic perspective. For. Ecol. Manag. 2007, 253, 161–170. [Google Scholar] [CrossRef]
- Šebková, B.; Šamonil, P.; Janik, D.; Adam, D.; Král, K.; Vrška, T.; Hort, L.; Unar, P. Spatial and volume patterns of an unmanaged submontane mixed forest in Central Europe: 160 years of spontaneous dynamics. For. Ecol. Manag. 2011, 262, 873–885. [Google Scholar] [CrossRef]
- Paluch, J.; Kołodziej, Z.; Pach, M.; Jastrzębski, R. Spatial variability of close-to primeval Fagus-Abies-Picea forests in the Western Carpathians (Central Europe): A step towards a generalised pattern. Eur. J. For. Res. 2015, 134, 235–246. [Google Scholar] [CrossRef]
- Penttinen, A.; Stoyan, D.; Henttonen, H.M. Marked point processes in forest statistics. For. Sci. 1992, 38, 806–824. [Google Scholar] [CrossRef]
- Hung, C.C.; Song, E.; Lan, Y. Image Texture Analysis; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Král, K.; Valtera, M.; Janik, D.; Šamonil, P.; Vrška, T. Spatial variability of general stand characteristics in central European beech−dominated natural stands—Effects of scale. For. Ecol. Manag. 2014, 328, 353–364. [Google Scholar] [CrossRef]
- Staudhammer, C.L.; LeMay, V.M. Introduction and evaluation of possible indices of stand structural diversity. Can. J. For. Res. 2001, 31, 1105–1115. [Google Scholar] [CrossRef]
- Gavrikov, V.; Stoyan, D. The use of marked point processes in ecological and environmental forest studies. Environ. Ecol. Stat. 1995, 2, 331–344. [Google Scholar] [CrossRef]
- Stupar, V.; Milanović, D. Istorijat zaštite prirode na području Nacionalnog parka Sutjeska. Glasnik Šumarskog Fakulteta Univerziteta u Banjoj Luci 2017, 26, 113–128. [Google Scholar] [CrossRef]
- Sabatini, F.M.; Burrascano, S.; Keeton, W.S.; Levers, C.; Lindner, M.; Pötzschner, F.; Verkerk, P.J.; Bauhus, J.; Buchwald, E.; Chaskovsky, O.; et al. Where are Europe’s last primary forests? Divers. Distrib. 2018, 24, 1426–1439. [Google Scholar] [CrossRef]
- Paluch, J.; Keren, S.; Govedar, Z. The Dinaric Mountains versus the Western Carpathians: Is structural heterogeneity similar in close-to-primeval Abies–Picea–Fagus forests? Eur. J. For. Res. 2020. [Google Scholar] [CrossRef]
- Paluch, J.; Bartkowicz, L. Przestrzenna zmienność struktury drzewostanu w wybranych lasach o charakterze pierwotnym w Karpatach Zachodnich i Górach Dynarskich. Sylwan 2020, 164, 91–101. [Google Scholar] [CrossRef]
- Leibundgut, H. Über Zweck und Methodik der Struktur- und Zuwachsanalyse von Urwäldern. Schweiz. Z. Forstwes. 1959, 110, 111–124. [Google Scholar]
- Leibundgut, H. Über die Dynamik europäischen Urwälder. Allg. Forst Ztg 1978, 24, 686–688. [Google Scholar]
- Mayer, H. Europäische Urwälder; Fischer Verlag: Stuttgart, Germany, 1986. [Google Scholar]
- Zenner, E.K.; Peck, J.E.; Hobi, M.L.; Commarmot, B. Validation of a classification protocol: Meeting the prospect requirement and ensuring distinctiveness when assigning forest development phases. Appl. Veg. Sci. 2016, 19, 541–552. [Google Scholar] [CrossRef]
- Goff, F.G.; West, D. Canopy-understory interaction effects on forest population structure. For. Sci. 1975, 21, 98–108. [Google Scholar] [CrossRef]
- Schütz, J.P. Der Plenterwald und Weitere Formen Strukturierter und Gemischter Wälder; Parey: Berlin, Germany, 2001. [Google Scholar]
- Schütz, J.P. Modelling the demographic sustainability of pure beech plenter forests in eastern Germany. Ann. For. Sci. 2006, 63, 93–100. [Google Scholar] [CrossRef]
- Schütz, J.P.; Saniga, M.; Diaci, J.; Vrška, T. Comparing close-to-nature silviculture with processes in pristine forests: Lessons from Central Europe. Ann. For. Sci. 2016, 73, 911–921. [Google Scholar] [CrossRef]
- Mayer, H. Gebirgswald-Schutzwaldpflege; Fischer Verlag: Stuttgart, Germany, 1976. [Google Scholar]
- Commarmot, B.; Bachofen, H.; Bundziak, Y.; Bürgi, A.; Ramp, B.; Shparyk, Y.; Sukhariuk, D.; Viter, R.; Zingg, A. Structures of virgin and managed beech forests in Uholka (Ukraine) and Sihlwald (Switzerland): A comparative study. For. Snow. Lands. Res. 2005, 79, 45–56. [Google Scholar]
- Motta, R.; Berretti, R.; Castagneri, D.; Dukič, V.; Garbarino, M.; Govedar, Z.; Lingua, E.; Maunaga, Z.; Meloni, F. Toward a definition of the range of variability of central European mixed Fagus-Abies-Picea forests: The nearly steady-state forest of Lom (Bosnia and Herzegovina). Can. J. For. Res. 2011, 41, 1871–1884. [Google Scholar] [CrossRef]
- Drössler, L.; Feldmann, E.; Glatthorn, J.; Annighöfer, P.; Kucbel, S.; Tabaku, V. What happens after the gap? Size distributions of patches with homogeneously sized trees in natural and managed beech forests in Europe. Open J. For. 2016, 6, 177–190. [Google Scholar] [CrossRef]
- Parobeková, Z.; Pittner, J.; Kucbel, S.; Saniga, M.; Filípek, M.; Sedmáková, D.; Vencurik, J.; Jaloviar, P. Structural diversity in a mixed spruce-fir-beech old-growth forest remnant of the Western Carpathians. Forests 2018, 9, 379. [Google Scholar] [CrossRef]
- Paluch, J. The stochastic backward shifts model better corresponds to the fine-scale structural heterogeneity of old-growth Abies-Fagus-Picea forests compared to the ontogenic life cycle. For. Ecol. Manag. 2020. under review. [Google Scholar]
- Král, K.; Daněk, P.; Janík, D.; Krůček, M.; Vrška, T. How cyclical and predictable are Central European temperate forest dynamics in terms of development phases? J. Veg. Sci. 2018, 29, 84–97. [Google Scholar] [CrossRef]
- Bütler, R.; Patty, L.; Le Bayon, R.C.; Guenat, C.; Schlaepfer, R. Log decay of Picea abies in the Swiss Jura Mountains of Central Europe. For. Ecol. Manag. 2007, 242, 791–799. [Google Scholar] [CrossRef]
- Holeksa, J.; Zielonka, T.; Żywiec, M. Modeling the decay of coarse woody debris in a subalpine Norway spruce forest of the West Carpathians, Poland. Can. J. For. Res. 2008, 38, 415–428. [Google Scholar] [CrossRef]
- Lombardi, F.; Cherubini, P.; Lasserre, B.; Tognetti, R.; Marchetti, M. Tree rings used to assess time since death of deadwood of different decay classes in beech and silver fir forests in the central Apennines (Molise, Italy). Can. J. For. Res. 2008, 38, 821–833. [Google Scholar] [CrossRef]
- Müller-Using, S.; Bartsch, N. Decay dynamic of coarse and fine woody debris of a beech (Fagus sylvatica L.) forest in Central Germany. Eur. J. For. Res. 2009, 128, 287–296. [Google Scholar] [CrossRef]
- Přívětivý, T.; Adam, D.; Vrška, T. Decay dynamics of Abies alba and Picea abies deadwood in relation to environmental conditions. For. Ecol. Manag. 2018, 427, 250–259. [Google Scholar] [CrossRef]
- Jaworski, A. Dolnoreglowe Lasy o Charakterze Pierwotnym w Babiogórskim Parku Narodowym (Lata 1930–2006); Wydawnictwo Uniwersytetu Rolniczego w Krakowie: Kraków, Poland, 2016. [Google Scholar]
- Jaworski, A.; Paluch, J. Factors affecting the basal area increment of the primeval forests in the Babia Góra National Park, Southern Poland. Forstw. Cbl. 2002, 121, 97–108. [Google Scholar] [CrossRef]
- Jaworski, A.; Podlaski, R. Structure and dynamics of selected stands of primeval character in the Pieniny National Park. Dendrobiol. 2007, 58, 25–42. [Google Scholar]
- Zenner, E.K.; Peck, J.E.; Hobi, M.L.; Commarmot, B. The dynamics of structure across scale in a primaeval European beech stand. Forestry 2015, 88, 180–189. [Google Scholar] [CrossRef]
- Zenner, E.K.; Peck, J.E.; Hobi, M.L. Development phase convergence across scale in a primeval European beech (Fagus sylvatica L.) forest. For. Ecol. Manag. 2020, 460. [Google Scholar] [CrossRef]
- Alessandrini, A.; Biondi, F.; Di Filippo, A.; Ziaco, E.; Piovesan, G. Tree size distribution at increasing spatial scales converges to the rotated sigmoid curve in two old-growth beech stands of the Italian Apennines. For. Ecol. Manag. 2011, 262, 1950–1962. [Google Scholar] [CrossRef]
- Zenner, E.K.; Peck, J.E.; Sagheb-Talebi, K. Patchiness in old-growth oriental beech forests across development stages at multiple neighborhood scales. Eur. J. For. Res. 2019, 138, 739–752. [Google Scholar] [CrossRef]
- Zenner, E.K.; Sagheb-Talebi, K.; Akhavan, R.; Peck, J.E. Integration of small-scale canopy dynamics smoothes live-tree structural complexity across development stages in old growth Oriental beech (Fagus orientalis Lipsky) forests at the multi-gap scale. For. Ecol. Manag. 2015, 335, 26–36. [Google Scholar] [CrossRef]
- Nagel, T.A.; Svoboda, M. Gap disturbance regime in an old-growth Fagus–Abies forest in the Dinaric Mountains, Bosnia-Herzegovina. Can. J. For. Res. 2008, 38, 2728–2737. [Google Scholar] [CrossRef]
- Garbarino, M.; Mondino, E.B.; Lingua, E.; Nagel, T.A.; Dukić, V.; Govedar, Z.; Motta, R. Gap disturbances and regeneration patterns in a Bosnian old-growth forest: A multispectral remote sensing and and ground-based approach. Ann. For. Sci. 2012, 69, 617–625. [Google Scholar] [CrossRef]
- Drössler, L.; von Lüpke, B. Canopy gaps in two virgin beech forest reserves in Slovakia. J. For. Sci. 2005, 51, 446–457. [Google Scholar] [CrossRef]
- Zeibig, A.; Diaci, J.; Wagner, S. Gap disturbance patterns of a Fagus sylvatica virgin forest remnant in the mountain vegetation belt of Slovenia. For. Snow. Landsc. Res. 2005, 79, 69–80. [Google Scholar]
- Kenderes, K.; Král, K.; Vrška, T.; Standovár, T. Natural gap dynamics in a Central European mixed beech-spruce-fir old growth forest. Ecoscience 2009, 16, 39–47. [Google Scholar] [CrossRef]
- Kucbel, S.; Jaloviar, P.; Saniga, M.; Vencurik, J.; Klimaš, V. Canopy gaps in an old-growth fir-beech forest remnant of Western Carpathians. Eur. J. For. Res. 2010, 129, 249–259. [Google Scholar] [CrossRef]
- Bottero, A.; Garbarino, M.; Dukić, V.; Govedar, Z.; Lingua, E.; Nagel, T.A.; Motta, R. Gap-phase dynamics in the old-growth forest of Lom, Bosnia and Herzegovina. Silva. Fenn. 2011, 45, 875–887. [Google Scholar] [CrossRef]
- Hobi, M.L.; Ginzler, C.; Commarmot, B.; Bugmann, H. Gap pattern of the largest primeval beech forest of Europe revealed by remote sensing. Ecosphere 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Král, K.; Janik, D.; Vrška, T.; Adam, D.; Hort, L.; Unar, P.; Šamonil, P. Local variability of stand structural features in beech dominated natural forests of Central Europe: Implications for sampling. For. Ecol. Manag. 2010, 260, 2196–2203. [Google Scholar] [CrossRef]
- Lombardi, F.; Marchetti, M.; Corona, P.; Merlini, P.; Chirici, G.; Tognetti, R.; Burrascano, S.; Alivernini, A.; Puletti, N. Quantifying the effect of sampling plot size on the estimation of structural indicators in old-growth forest stands. For. Ecol. Manag. 2015, 346, 89–97. [Google Scholar] [CrossRef]
- Peck, J.E.; Commarmot, B.; Hobi, M.L.; Zenner, E.K. Should reference conditions be drawn from a single 10-ha plot? Assessing representativeness in a 10,000-ha old-growth European beech forest. Rest. Ecol. 2015, 23, 927–935. [Google Scholar] [CrossRef]
- Carrer, M.; Castagneri, D.; Popa, I.; Pividori, M.; Lingua, E. Tree spatial patterns and stand attributes in temperate forests: The importance of plot size, sampling design, and null model. For. Ecol. Manag. 2018, 407, 125–134. [Google Scholar] [CrossRef]
- Grabarnik, P.; Myllymäki, M.; Stoyan, D. Correct testing of mark independence for marked point patterns. Ecol. Model. 2011, 222, 3888–3894. [Google Scholar] [CrossRef]
- Král, K.; McMahon, S.M.; Janík, D.; Adam, D.; Vrška, T. Patch mosaic of developmental stages in central European natural forests along vegetation gradient. For. Ecol. Manag. 2014, 330, 17–28. [Google Scholar] [CrossRef]
- Glatthorn, J.; Feldmann, E.; Tabaku, V.; Leuschner, C.; Meyer, P. Classifying development stages of primeval European beech forests: Is clustering a useful tool? BMC Ecol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.; Emborg, J.; Nielsen, A.B. The forest cycle of Suserup Skov: Revisited and revised. Ecol. Bull. 2007, 52, 33–42. [Google Scholar] [CrossRef]
- Hao, Z.; Zhang, J.; Song, B.; Ye, J.; Li, B. Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest. For. Ecol. Manag. 2007, 252, 1–11. [Google Scholar] [CrossRef]
- Larson, A.J.; Lutz, J.A.; Donato, D.C.; Freund, J.A.; Swanson, M.E.; HilleRisLambers, J.; Sprugel, D.G.; Franklin, J.F. Spatial aspects of tree mortality strongly differ between young and old-growth forests. Ecology 2015, 96, 2855–2861. [Google Scholar] [CrossRef]
- Bartkowicz, L.; Paluch, J. Co-occurrence of shade-tolerant and light-adapted tree species in uneven-aged deciduous forests of southern Poland. Eur. J. For. Res. 2019, 138, 15–30. [Google Scholar] [CrossRef]
- Liu, P.; Wang, W.; Bai, Z.; Guo, Z.; Ren, W.; Huang, J. Competition and facilitation co-regulate the spatial patterns of boreal tree species in Kanas of Xinjiang, northwest China. For. Ecol. Manag. 2020. [Google Scholar] [CrossRef]
- Shen, G.; He, F.; Waagepetersen, R.; Sun, I.; Hao, Z.; Chen, Z.; Yu, M. Quantifying effects of habitat heterogeneity and other clustering processes on spatial distributions of tree species. Ecology 2013, 94, 2436–2443. [Google Scholar] [CrossRef]
- Lin, Y.; Chang, L.; Yang, K.; Wang, H.; Sun, I. Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation. Oecologia 2011, 165, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Després, T.; Vítková, L.; Bače, R.; Čada, V.; Janda, P.; Mikoláš, M.; Schurman, J.S.; Trotsiuk, V.; Svoboda, M. Past disturbances and intraspecific competition as drivers of spatial pattern in primary spruce forests. Ecosphere 2017. [Google Scholar] [CrossRef]
- McIntire, E.J.B.; Fajardo, A. Beyond description: The active and effective way to infer processes from spatial patterns. Ecology 2009, 90, 46–56. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | JA | LO | PE | BA | BG | OS |
|---|---|---|---|---|---|---|
| Location | Dinaric Mts (Bosnia and Herzegovina) | Western Carpathians (Poland) | ||||
| Reserve | Janj | Lom | Perućica | Baniska | Żarnówka | Oszast |
| Coordinates | 44°08′ N 17°18′ E | 44°27′ N 16°29′ E | 43°18′ N 18°43′ E | 49º27′ N 20º37′ E | 49º35′ N 19º33′ E | 49°25′ N 19°11′ E |
| Altitude (m a.s.l.) | 1260 –1400 | 1280 –1360 | 1420 –1480 | 900 –1050 | 930 –1020 | 1020 –1080 |
| Exposure | W | NE | SW | NW | N | NW |
| Inclination (º) | 0–10 | 10–25 | 10–20 | 10–20 | 10–20 | 15–25 |
| Bedrock | Dolomites | Limestones | Limestones | Flysch | Flysch | Flysch |
| Soils | Cambisols Rendzinas | Cambisols Chernozems Luvisols | Cambisols | Cambisols | Cambisols | Cambisols |
| Formal protection since (year) | 1951 | 1956 | 1952 | 1916 | 1936 | 1971 |
| Characteristics | JA | LO | PE | BA | BG | OS | ||
|---|---|---|---|---|---|---|---|---|
| Sample plots | 244 | 255 | 256 | 237 | 259 | 255 | ||
| Stem density | Total | N ha−1 | 569 | 532 | 430 | 682 | 288 | 464 |
| Abies | % | 16 | 27 | 63 | 13 | 16 | 4 | |
| Fagus | % | 72 | 55 | 32 | 84 | 65 | 80 | |
| Picea | % | 12 | 18 | 5 | 3 | 19 | 16 | |
| Basal area | Total | m2 ha−1 | 65.9 | 56.8 | 65.2 | 37.1 | 36.1 | 36.7 |
| Abies | % | 45 | 37 | 54 | 33 | 29 | 11 | |
| Fagus | % | 19 | 31 | 38 | 64 | 45 | 39 | |
| Picea | % | 36 | 32 | 8 | 3 | 26 | 50 | |
| Volume | m3 ha−1 | 1004 | 903 | 1134 | 478 | 522 | 574 | |
| Id | - | 1.67 | 1.89 | 1.26 | 1.10 | 1.72 | 1.09 | |
| BA75 | m2 ha−1 | 80.6 | 70.2 | 82.6 | 44.3 | 49.2 | 48.8 | |
| BAss | m2 ha−1 | 65.9 | 56.8 | 65.2 | 37.1 | 36.1 | 36.6 | |
| dbhBA | cm | 77 | 69 | 69 | 57 | 65 | 61 |
| Phase | JA | LO | PE | BA | BG | OS |
|---|---|---|---|---|---|---|
| Spatial Scale 800 m2 | ||||||
| All phases | n.s. | n.s. | n.s. | n.s. | A | A |
| Early accumulation | n.s. | n.s. | n.s. | n.s. | A | A |
| Late accumulation | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| Culmination | A | n.s. | A | S | A | n.s. |
| Steady-state | n.s. | n.s. | A | n.s. | n.s. | n.s. |
| Spatial Scale 1600 m2 | ||||||
| All phases | n.s. | n.s. | n.s. | S | A | n.s. |
| Early accumulation | n.s. | n.s. | n.s. | n.s. | A | n.s. |
| Late accumulation | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| Culmination | A | n.s. | A | n.s. | A | n.s. |
| Steady-state | n.s. | n.s. | A | n.s. | n.s. | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paluch, J. On the Consequences of Using Moving Window Segmentation to Analyze the Structural Stand Heterogeneity and Debatable Patchiness of Old-Growth Temperate Forests. Forests 2021, 12, 96. https://doi.org/10.3390/f12010096
Paluch J. On the Consequences of Using Moving Window Segmentation to Analyze the Structural Stand Heterogeneity and Debatable Patchiness of Old-Growth Temperate Forests. Forests. 2021; 12(1):96. https://doi.org/10.3390/f12010096
Chicago/Turabian StylePaluch, Jarosław. 2021. "On the Consequences of Using Moving Window Segmentation to Analyze the Structural Stand Heterogeneity and Debatable Patchiness of Old-Growth Temperate Forests" Forests 12, no. 1: 96. https://doi.org/10.3390/f12010096
APA StylePaluch, J. (2021). On the Consequences of Using Moving Window Segmentation to Analyze the Structural Stand Heterogeneity and Debatable Patchiness of Old-Growth Temperate Forests. Forests, 12(1), 96. https://doi.org/10.3390/f12010096