Xylem Phenology and Growth Response of European Beech, Silver Fir and Scots Pine along an Elevational Gradient during the Extreme Drought Year 2018

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
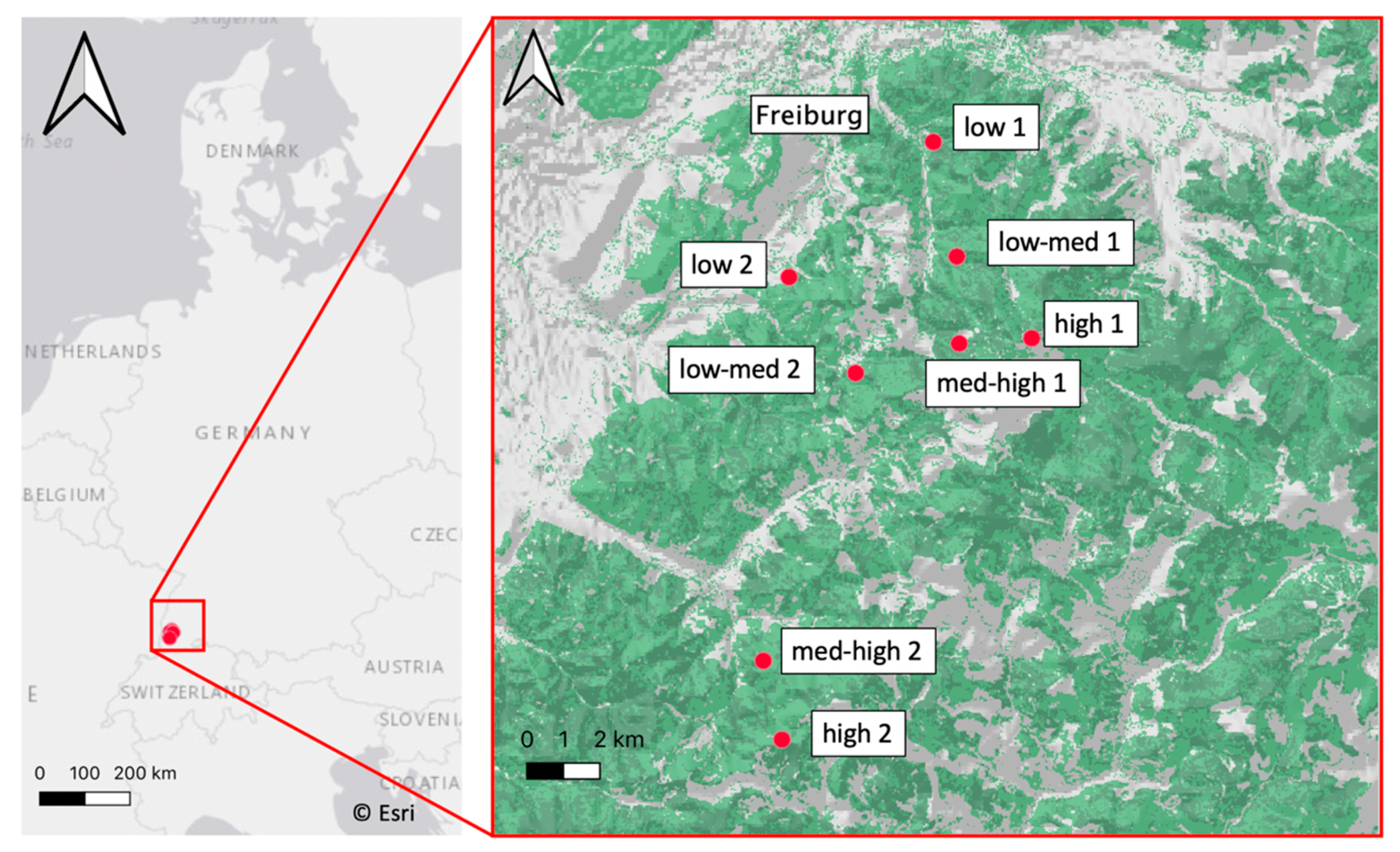
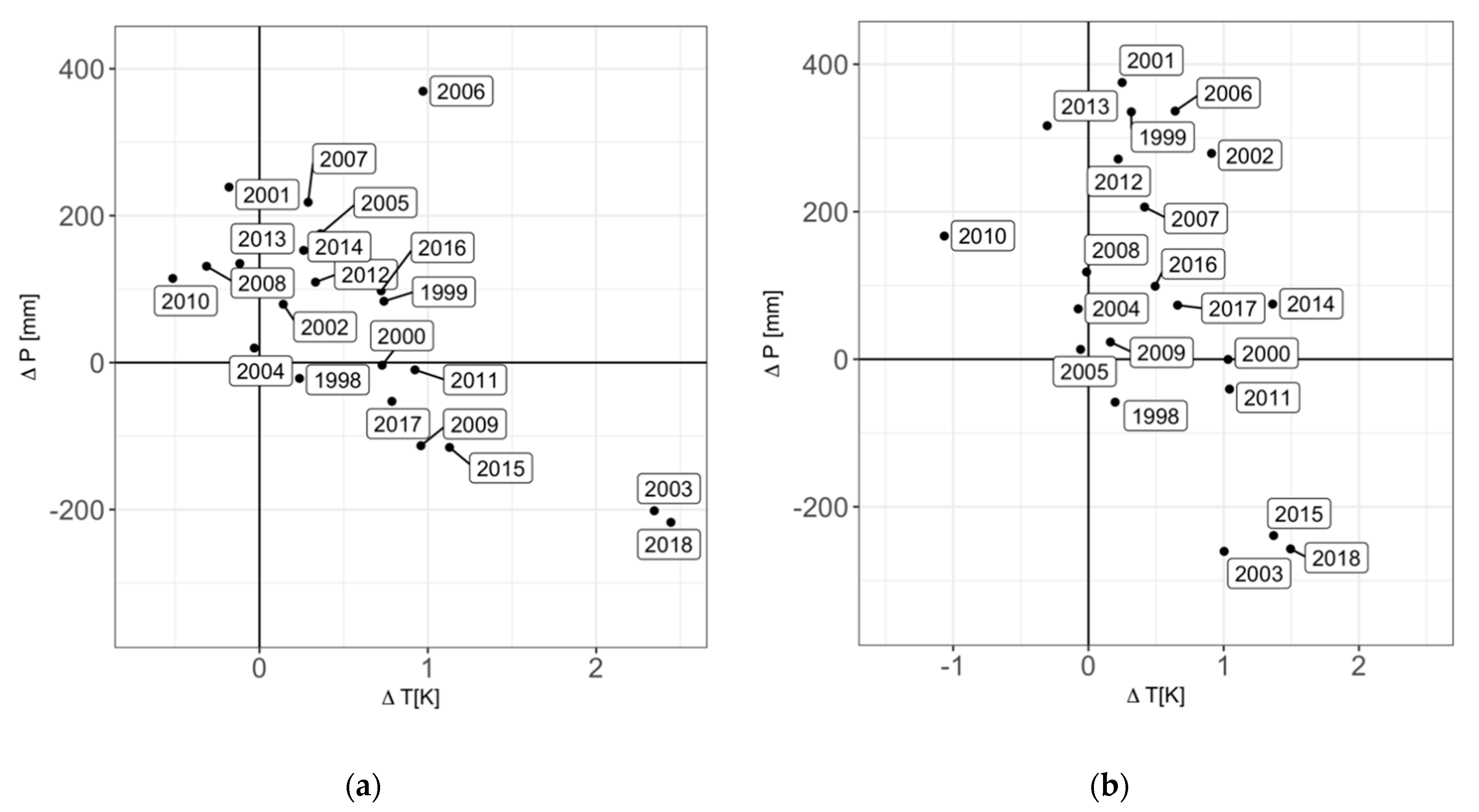
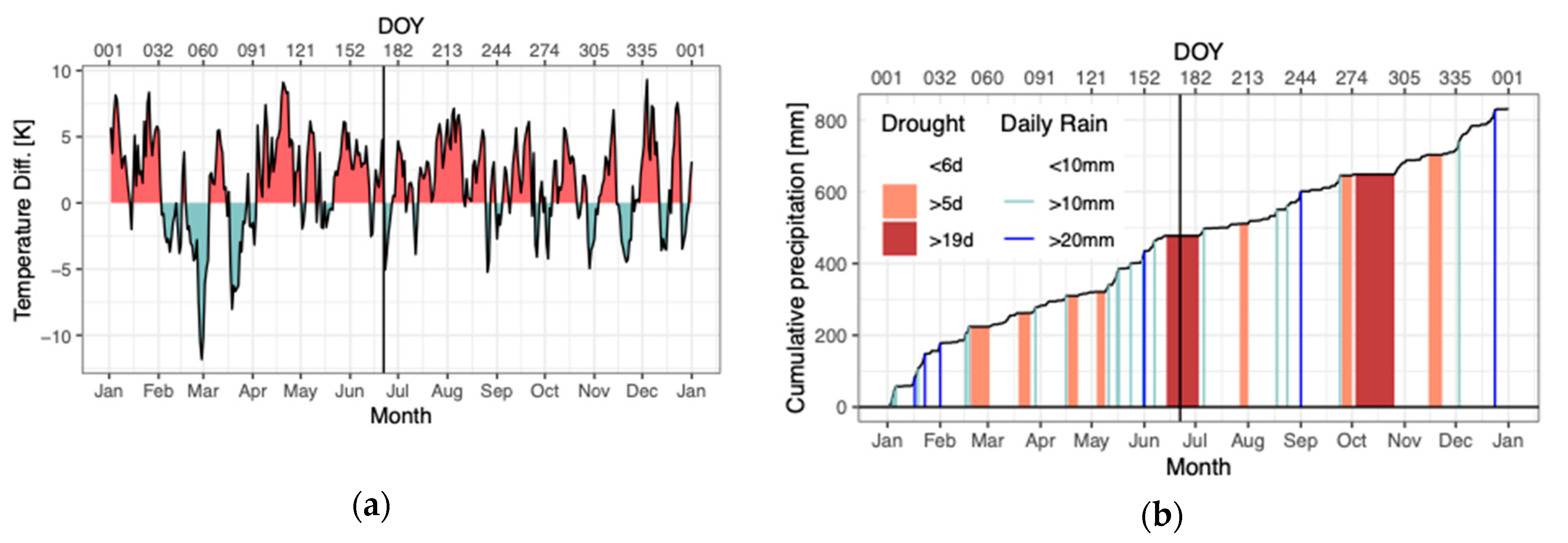
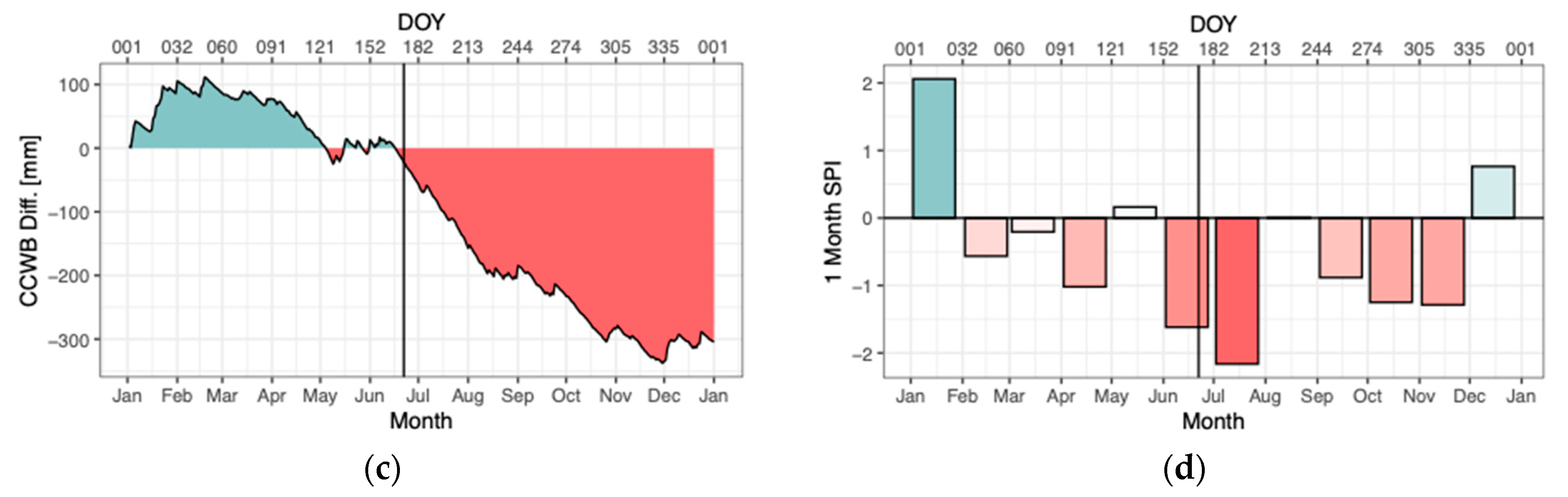
2.1. Research Design and Climatic Conditions
2.2. Field, Laboratory and Statistical Analysis Methods
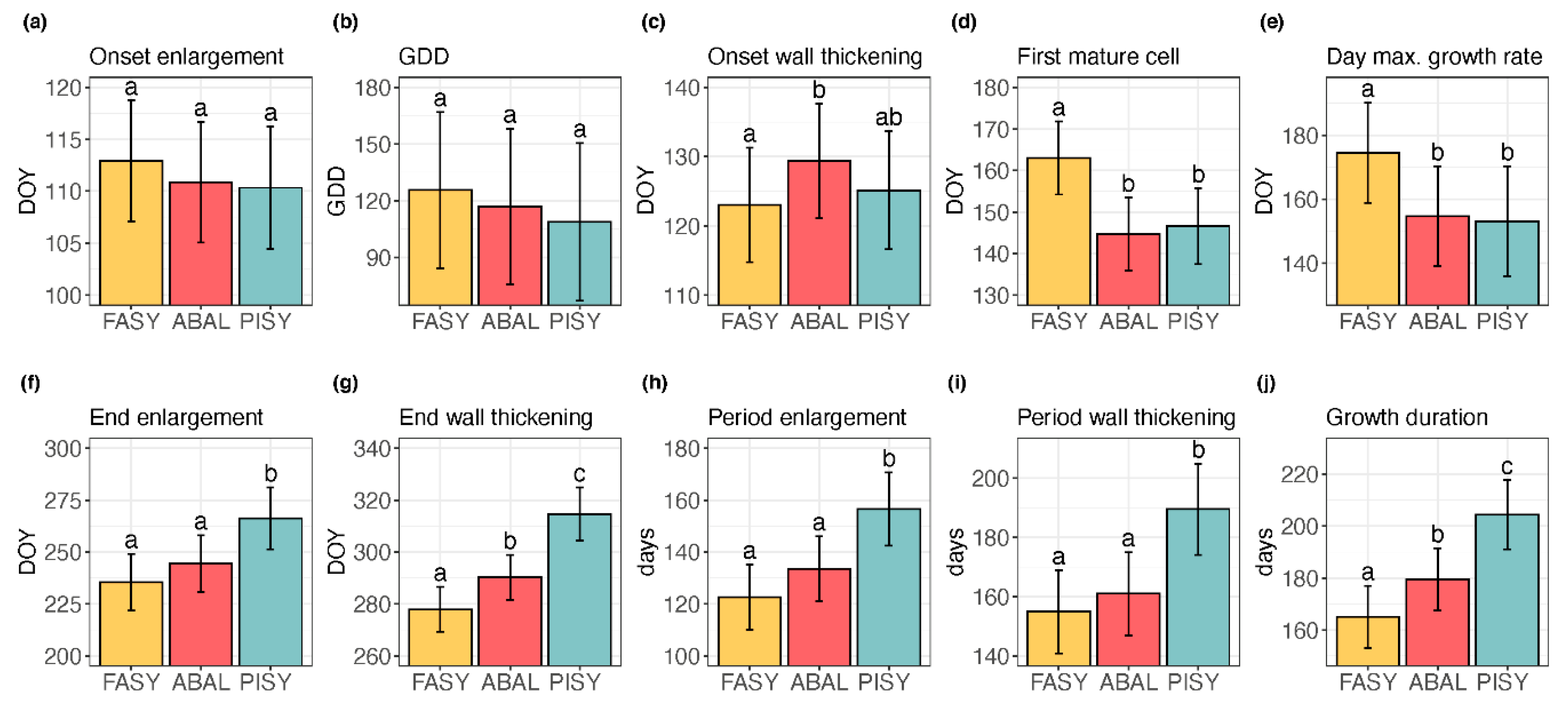
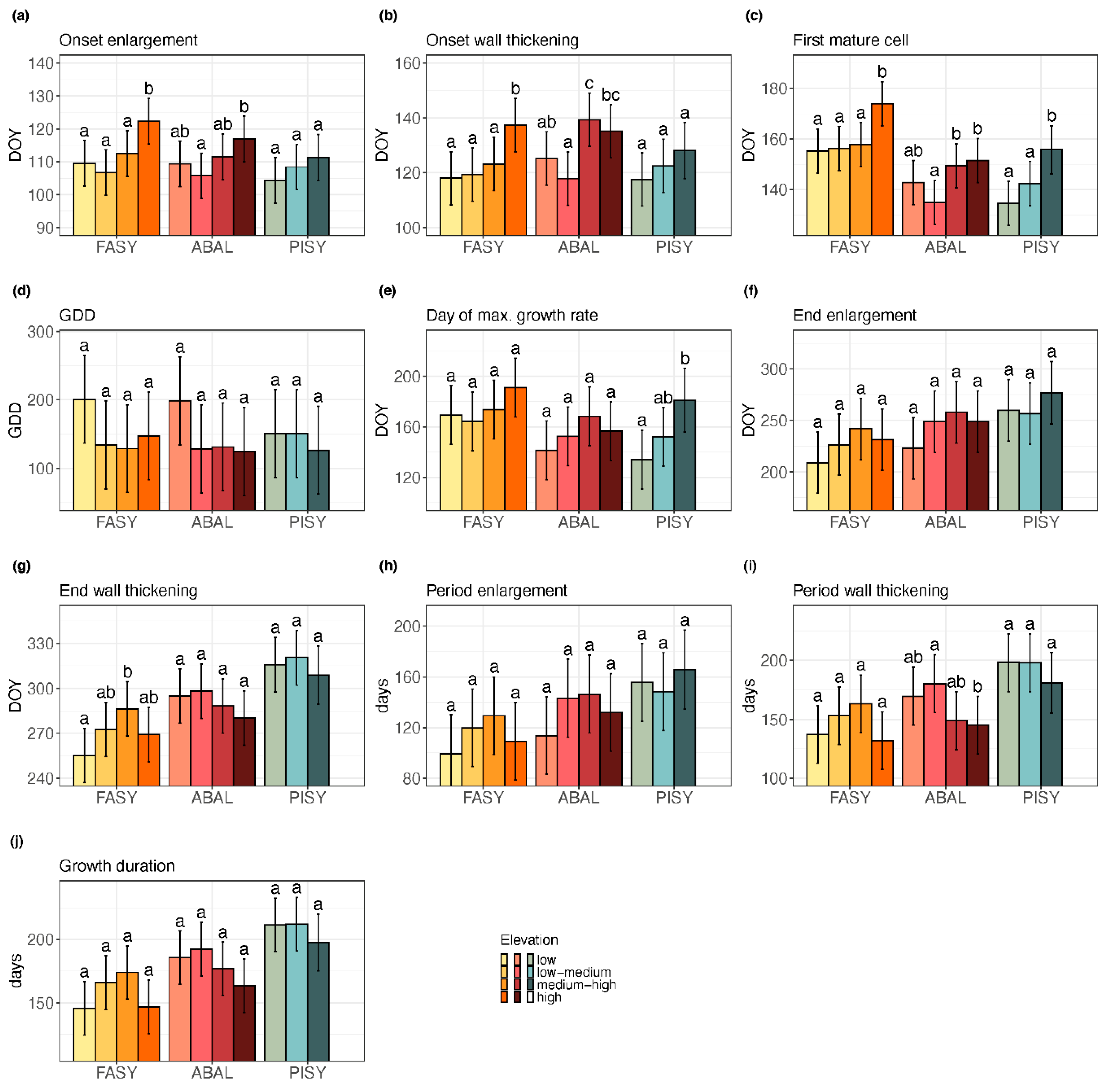
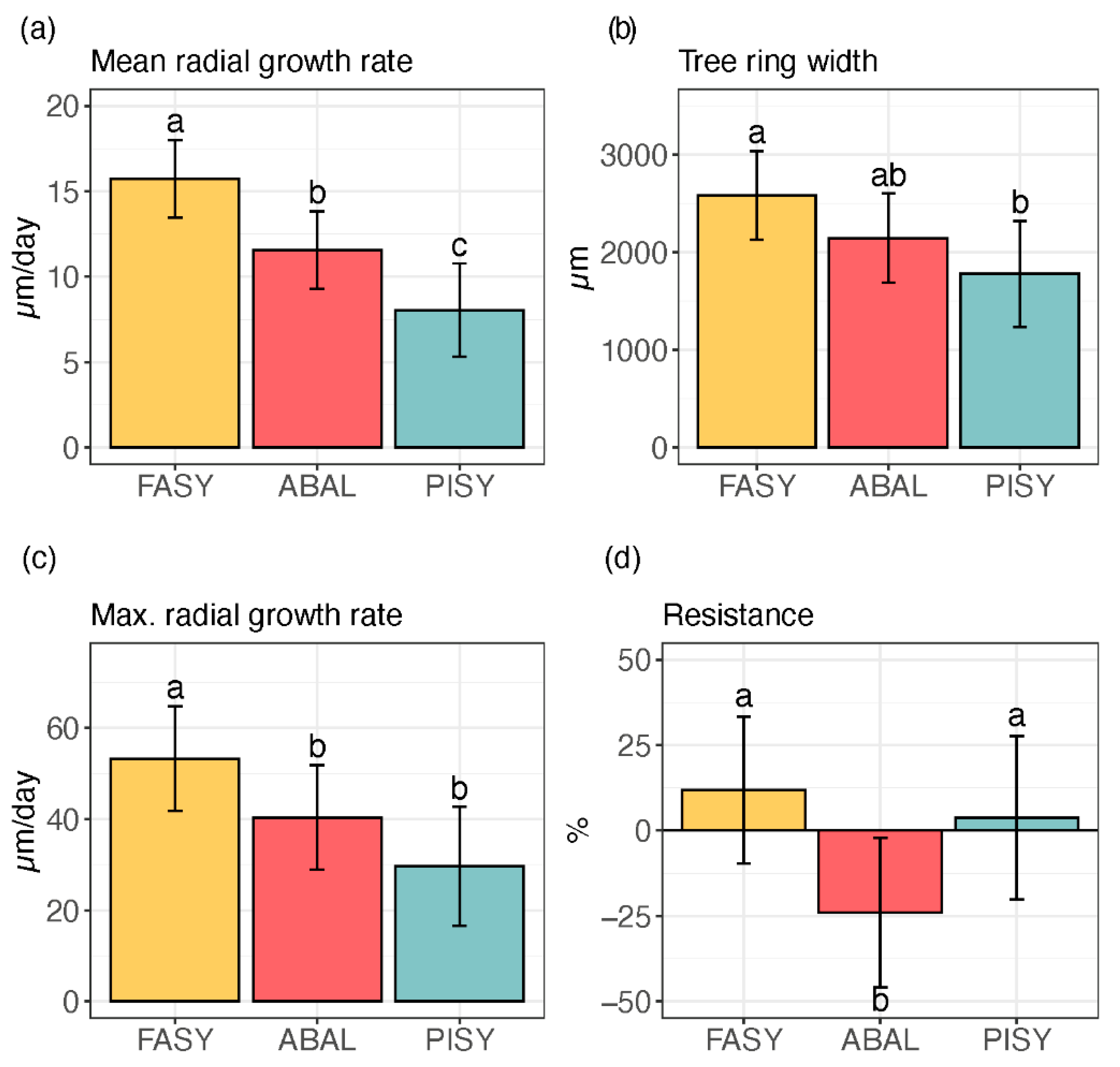
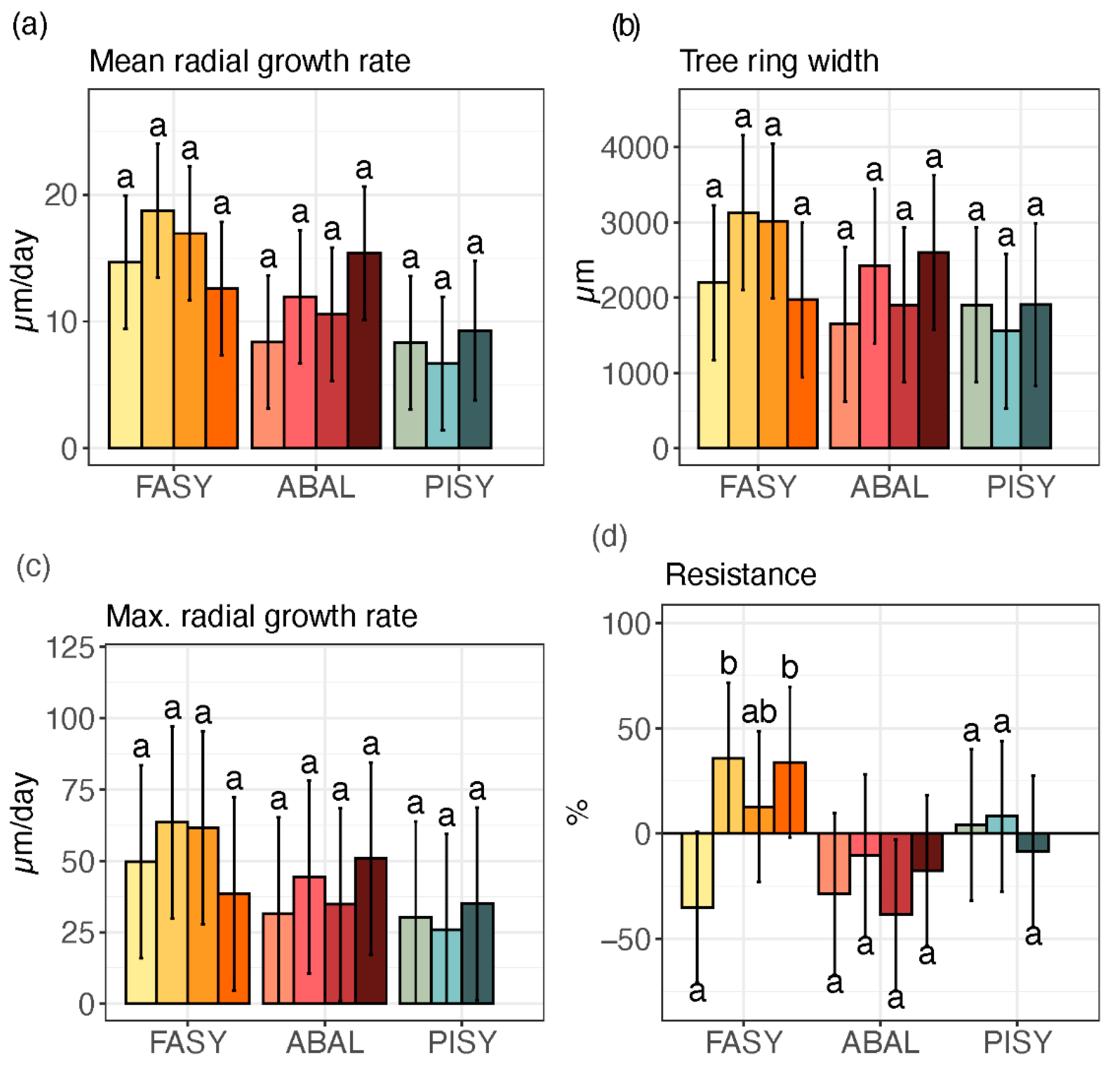
3. Results
3.1. Xylem Phenology
3.1.1. The Species Effect
3.1.2. The Elevational Gradient
3.2. Xylem Growth
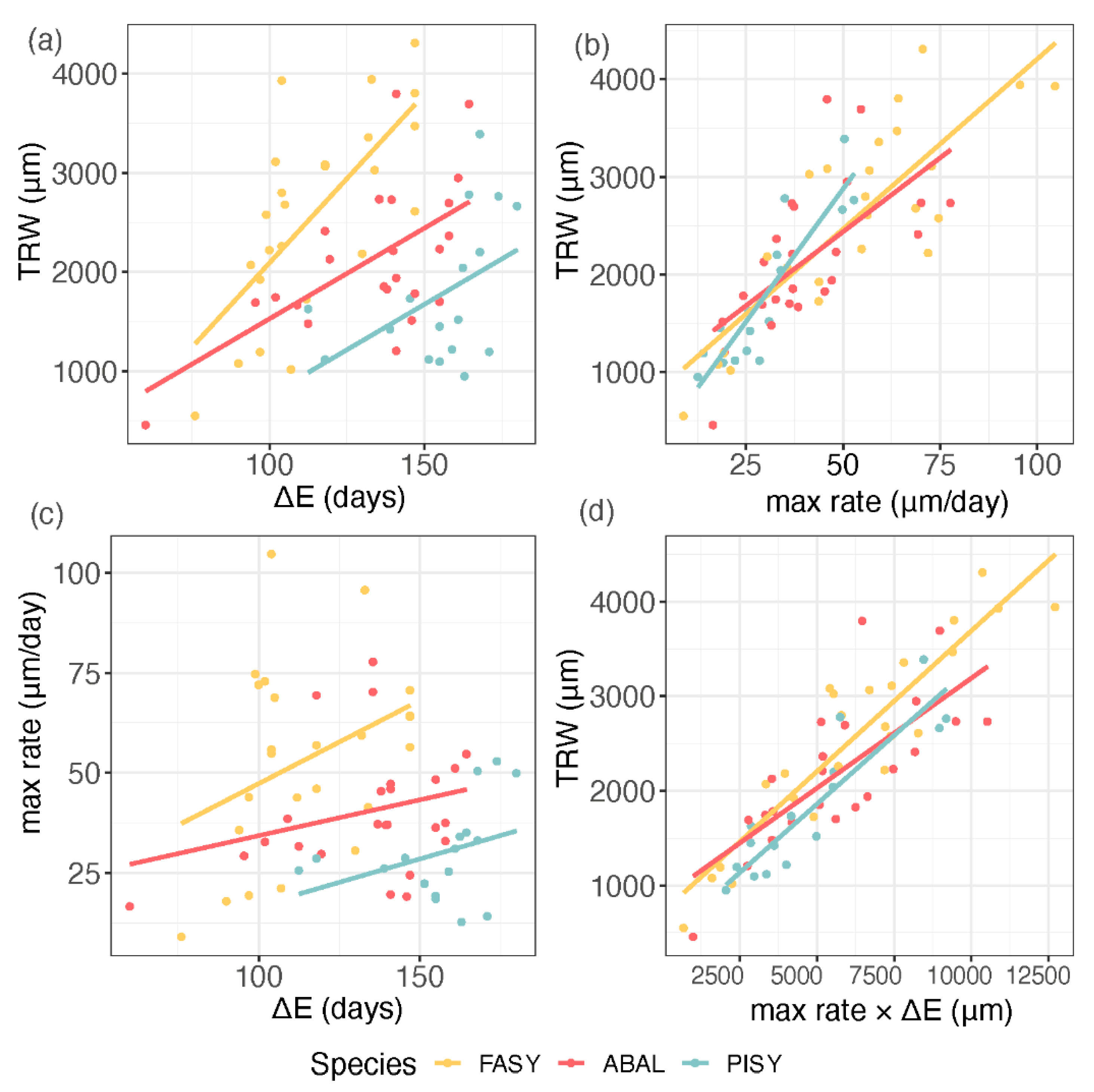
3.3. Interaction between Tree-Ring Growth and Phenology
4. Discussion
4.1. Onset of Cell Differentiation Phases and Thermal Accumulation in the Hot Spring 2018
4.2. Cessation and Duration of Cell Differentiation after the Summer Drought 2018
4.3. Tree-Ring Width in Relation to Xylem Phenology
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.G.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Skiadaresis, G.; Schwarz, J.A.; Bauhus, J. Groundwater Extraction in Floodplain Forests Reduces Radial Growth and Increases Summer Drought Sensitivity of Pedunculate Oak Trees (Quercus robur L.). Front. For. Glob. Chang. 2019, 2. [Google Scholar] [CrossRef]
- Buras, A.; Rammig, A.; Zang, C.S. Quantifying impacts of the 2018 drought on European ecosystems in comparison to 2003. Biogeosciences 2020, 17, 1655–1672. [Google Scholar] [CrossRef]
- Yuan, W.; Zheng, Y.; Piao, S.; Ciais, P.; Lombardozzi, D.; Wang, Y.; Ryu, Y.; Chen, G.; Dong, W.; Hu, Z.; et al. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 2019, 5, eaax1396. [Google Scholar] [CrossRef]
- Schuldt, B.; Buras, A.; Arend, M.; Vitasse, Y.; Beierkuhnlein, C.; Damm, A.; Gharun, M.; Grams, T.E.E.; Hauck, M.; Hajek, P.; et al. A first assessment of the impact of the extreme 2018 summer drought on Central European forests. Basic Appl. Ecol. 2020. [Google Scholar] [CrossRef]
- Kovats, R.S.; Valentini, R.; Bouwer, L.M.; Georgopoulou, E.; Jacob, D.; Martin, E.; Rounsevell, M.; Soussana, J.-F. Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Clim. Chang. 2014, 2014, 1267–1326. [Google Scholar]
- Rathgeber, C.B.K.; Fonti, P.; Shishov, V.V.; Rozenberg, P. Wood formation and tree adaptation to climate. Ann. For. Sci. 2019, 76, 10–12. [Google Scholar] [CrossRef]
- Badeck, F.-W.; Bondeau, A.; Bottcher, K.; Doktor, D.; Lucht, W.; Schaber, J.; Sitch, S. Responses of spring phenology to climate change. New Phytol. 2004, 162, 295–309. [Google Scholar] [CrossRef]
- Donnelly, A.; Jones, M.; Sweeney, J. A review of indicators of climate change for use in Ireland. Int. J. Biometeorol. 2004, 49. [Google Scholar] [CrossRef]
- Gričar, J.; Prislan, P.; Gryc, V.; Vavrčík, H.; De Luis, M.; Čufar, K. Plastic and locally adapted phenology in cambial seasonality and production of xylem and phloem cells in Picea abies from temperate environments. Tree Physiol. 2014, 34, 869–881. [Google Scholar] [CrossRef]
- Vieira, J.; Moura, M.; Nabais, C.; Freitas, H.; Campelo, F. Seasonal adjustment of primary and secondary growth in maritime pine under simulated climatic changes. Ann. For. Sci. 2019, 76, 84. [Google Scholar] [CrossRef]
- Plomion, C.; Leprovost, G.; Stokes, A. Wood Formation in Trees Wood Formation in Trees. Plant Physiol. 2001, 127, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Chmielewski, F.M.; Rotzer, T. Response of tree phenology to climate change across Europe. Agric. For. Meteorol. 2001, 108, 101–112. [Google Scholar] [CrossRef]
- Čufar, K.; Prislan, P.; De Luis, M.; Gričar, J. Tree-ring variation, wood formation and phenology of beech (Fagus sylvatica) from a representative site in Slovenia, SE Central Europe. Trees-Struct. Funct. 2008, 22, 749–758. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Gričar, J.; Seo, J.W.; Rathgeber, C.B.K.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Moser, L.; Fonti, P.; Buntgen, U.; Esper, J.; Luterbacher, J.; Franzen, J.; Frank, D. Timing and duration of European larch growing season along altitudinal gradients in the Swiss Alps. Tree Physiol. 2010, 30, 225–233. [Google Scholar] [CrossRef]
- Martinez del Castillo, E.; Longares, L.A.; Gričar, J.; Prislan, P.; Gil-Pelegrín, E.; Čufar, K.; de Luis, M. Living on the Edge: Contrasted Wood-Formation Dynamics in Fagus sylvatica and Pinus sylvestris under Mediterranean Conditions. Front. Plant Sci. 2016, 7, 370. [Google Scholar] [CrossRef]
- Saderi, S.; Rathgeber, C.B.K.; Rozenberg, P.; Fournier, M. Phenology of wood formation in larch (Larix decidua Mill.) trees growing along a 1000-m elevation gradient in the French Southern Alps. Ann. For. Sci. 2019, 76, 89. [Google Scholar] [CrossRef]
- Lundqvist, S.-O.; Seifert, S.; Grahn, T.; Olsson, L.; García-Gil, M.R.; Karlsson, B.; Seifert, T. Age and weather effects on between and within ring variations of number, width and coarseness of tracheids and radial growth of young Norway spruce. Eur. J. For. Res. 2018, 137, 719–743. [Google Scholar] [CrossRef]
- Rathgeber, C.B.K.; Cuny, H.E.; Fonti, P. Biological Basis of Tree-Ring Formation: A Crash Course. Front. Plant Sci. 2016, 7, 734. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; de Luis, M.; Smith, K.T.; Čufar, K. Phenological variation in xylem and phloem formation in Fagus sylvatica from two contrasting sites. Agric. For. Meteorol. 2013, 180, 142–151. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Morin, H.; Saracino, A.; Motta, R.; Borghetti, M. Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length. New Phytol. 2006, 170, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Rathgeber, C.B.K.; Rossi, S.; Bontemps, J.D. Cambial activity related to tree size in a mature silver-fir plantation. Ann. Bot. 2011, 108, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carrer, M. Age-dependent xylogenesis in timberline conifers. New Phytol. 2008, 177, 199–208. [Google Scholar] [CrossRef]
- Vitasse, Y.; Delzon, S.; Bresson, C.C.; Michalet, R.; Kremer, A. Altitudinal differentiation in growth and phenology among populations of temperate-zone tree species growing in a common garden. Can. J. For. Res. 2009, 39, 1259–1269. [Google Scholar] [CrossRef]
- Cuny, H.E.; Rathgeber, C.B.K.; Kiessé, T.S.; Hartmann, F.P.; Barbeito, I.; Fournier, M. Generalized additive models reveal the intrinsic complexity of wood formation dynamics. J. Exp. Bot. 2013, 64, 1983–1994. [Google Scholar] [CrossRef]
- Klimo, E.; Hager, H.; Kulhavy, J. Spruce Monocultures in Central Europe—Problems and Prospects; European Forest Institute: Joensuu, Finland, 2000. [Google Scholar]
- Pugh, T.A.M.; Lindeskog, M.; Smith, B.; Poulter, B.; Arneth, A.; Haverd, V.; Calle, L. Role of forest regrowth in global carbon sink dynamics. Proc. Natl. Acad. Sci. USA 2019, 116, 4382–4387. [Google Scholar] [CrossRef]
- Bouriaud, O.; Popa, I. Comparative dendroclimatic study of Scots pine, Norway spruce, and silver fir in the Vrancea Range, Eastern Carpathian Mountains. Trees-Struct. Funct. 2009, 23, 95–106. [Google Scholar] [CrossRef]
- van der Maaten-Theunissen, M.; Kahle, H.-P.; van der Maaten, E. Drought sensitivity of Norway spruce is higher than that of silver fir along an altitudinal gradient in southwestern Germany. Ann. For. Sci. 2013, 70, 185–193. [Google Scholar] [CrossRef]
- Bohn, U.; Neuhäusl, R.; Gollub, G.; Hettwer, C.; Neuhäuslova, Z.; Schlüter, H.; Weber, H. Map of the Natural Vegetation of Europe 1: 2,500,000, Map and Explanatory Text; Landwirtschaftsverlag: Münster, Germany, 2003. [Google Scholar]
- Kandler, O. Development of the Recent Episode of Tannensterben (Fir Decline) in Eastern Bavaria and the Bavarian Alps. In Forest Decline in the Atlantic and Pacific Region; Hüttl, R.F., Mueller-Dombois, D., Eds.; Springer: Berlin/Heidelberg, Germany, 1993; pp. 216–226. [Google Scholar]
- Larsen, J.B. Das Tannensterben: Eine neue Hypothese zur Klärung des Hintergrundes dieser rätselhaften Komplexkrankheit der Weißtanne (Abies alba Mill.). Forstwiss. Cent. 1986, 105, 381–396. [Google Scholar] [CrossRef]
- Meining, S.; Puhlmann, H.; Augustin, N. Waldzustandsbericht 2016 für Baden-Württemberg; Forstl. Versuchs- und Forschungsanstalt Baden-württemb: Breisgau, Germany, 2016. [Google Scholar]
- BMEL. Wald und Rohholzpotenzial der Nächsten 40 Jahre; BMEL: Berlin, Germany, 2016. [Google Scholar]
- Dolos, K.; Märkel, U. Modellierung der Klimatischen Standorteignung Forstlich Relevanter Baumarten; LUBW Klimopass-Berichte: Karlsruhe, Germany, 2016. [Google Scholar]
- Antonucci, S.; Rossi, S.; Lombardi, F.; Marchetti, M.; Tognetti, R. Influence of climatic factors on silver fir xylogenesis along the Italian Peninsula. IAWA J. 2019, 40, 259–275. [Google Scholar] [CrossRef]
- Cuny, H.E.; Rathgeber, C.B.K.; Lebourgeois, F.; Fortin, M.; Fournier, M. Life strategies in intra-annual dynamics of wood formation: Example of three conifer species in a temperate forest in north-east France. Tree Physiol. 2012, 32, 612–625. [Google Scholar] [CrossRef] [PubMed]
- Reif, A.; Brucker, U.; Kratzer, R.; Schmiedinger, A.; Bauhus, J. Waldbau und Baumartenwahl in Zeiten des Klimawandels aus Sicht des Naturschutzes. BfN-Skripten 2010, 272, 125. [Google Scholar]
- Lebourgeois, F.; Rathgeber, C.B.K.; Ulrich, E. Sensitivity of French temperate coniferous forests to climate variability and extreme events (Abies alba, Picea abies and Pinus sylvestris). J. Veg. Sci. 2010, 21, 364–376. [Google Scholar] [CrossRef]
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Graf Pannatier, E.; Rigling, A. Drought alters timing, quantity, and quality of wood formation in Scots pine. J. Exp. Bot. 2011, 62, 2763–2771. [Google Scholar] [CrossRef] [PubMed]
- Bigler, C.; Bräker, O.U.; Bugmann, H.; Dobbertin, M.; Rigling, A. Drought as an inciting mortality factor in scots pine stands of the Valais, Switzerland. Ecosystems 2006, 9, 330–343. [Google Scholar] [CrossRef]
- Swidrak, I.; Gruber, A.; Oberhuber, W. Xylem and phloem phenology in co-occurring conifers exposed to drought. Trees 2014, 28, 1161–1171. [Google Scholar] [CrossRef]
- del Castillo, E.M.; Prislan, P.; Gričar, J.; Gryc, V.; Merela, M.; Giagli, K.; de Luis, M.; Vavrčík, H.; Čufar, K. Emerging challenges for beech and co-occuring conifers in a changing climate context. Dendrochronologia 2018, 52, 1–10. [Google Scholar] [CrossRef]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas Mit den Alpen: In Ökologischer, Dynamischer und Historischer Sicht, 5th ed.; UTB: Stuttgart, Germany, 2010. [Google Scholar]
- Prislan, P.; Gričar, J.; Čufar, K.; de Luis, M.; Merela, M.; Rossi, S. Growing season and radial growth predicted for Fagus sylvatica under climate change. Clim. Chang. 2019, 153, 181–197. [Google Scholar] [CrossRef]
- Scharnweber, T.; Manthey, M.; Criegee, C.; Bauwe, A.; Schröder, C.; Wilmking, M. Drought matters—Declining precipitation influences growth of Fagus sylvatica L. and Quercus robur L. in north-eastern Germany. For. Ecol. Manag. 2011, 262, 947–961. [Google Scholar] [CrossRef]
- Giagli, K.; Gričar, J.; Vavrčík, H.; Menšík, L.; Gryc, V. The effects of drought on wood formation in Fagus sylvatica during two contrasting years. IAWA J. 2016, 37, 332–348. [Google Scholar] [CrossRef]
- van der Werf, G.W.; Sass-Klaassen, U.; Mohren, G.M.J. The impact of the 2003 summer drought on the intra-annual growth pattern of beech (Fagus sylvatica L.) and oak (Quercus robur L.) on a dry site in the Netherlands. Dendrochronologia 2007, 25, 103–112. [Google Scholar] [CrossRef]
- Kraus, C.; Zang, C.; Menzel, A. Elevational response in leaf and xylem phenology reveals different prolongation of growing period of common beech and Norway spruce under warming conditions in the Bavarian Alps. Eur. J. For. Res. 2016, 135, 1011–1023. [Google Scholar] [CrossRef]
- Legendre, L.; Legendre, P. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Jarvis, A.; Reuter, H.I.; Nelson, A.; Guevara, E. Hole-filled seamless SRTM data V4, International Centre for Tropical Agriculture (CIAT). 2008. Available online: http://srtm.csi.cgiar.org (accessed on 1 December 2020).
- Schrödter, H. Verdunstung: Anwendungsorientierte Messverfahren und Bestimmungsmethoden; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Dietrich, H.; Wolf, T.; Kawohl, T.; Wehberg, J.; Kändler, G.; Mette, T.; Röder, A.; Böhner, J. Temporal and spatial high-resolution climate data from 1961 to 2100 for the German National Forest Inventory (NFI). Ann. For. Sci. 2019, 76, 6. [Google Scholar] [CrossRef]
- Nothdurft, A.; Wolf, T.; Ringeler, A.; Böhner, J.; Saborowski, J. Spatio-temporal prediction of site index based on forest inventories and climate change scenarios. For. Ecol. Manag. 2012, 279, 97–111. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.; Schelhaas, M.-J.; Nabuurs, G.-J.; Zimmermann, N. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Baier, P.; Pennerstorfer, J.; Schopf, A. PHENIPS—A comprehensive phenology model of Ips typographus (L.) (Col., Scolytinae) as a tool for hazard rating of bark beetle infestation. For. Ecol. Manag. 2007, 249, 171–186. [Google Scholar] [CrossRef]
- Pasho, E.; Camarero, J.J.; de Luis, M.; Vicente-Serrano, S.M. Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain. Agric. For. Meteorol. 2011, 151, 1800–1811. [Google Scholar] [CrossRef]
- Agnew, C.T. Using the SPI to Identify Drought. Drought Netw. News 2000, 12, 5–12. [Google Scholar]
- McKee, T.B.; Doesken, N.J.; Kleist, J. Drought monitoring with multiple time scales. In Proceedings of the Ninth Conference on Applied Climatology; American Meteorological Society: Boston, MA, USA, 1995; pp. 233–236. [Google Scholar]
- McMaster, G.S.; Wilhelm, W.W. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef]
- Rossi, S.; Anfodillo, T.; Menardi, R. Trephor: A new tool for sampling microcores from tree stems. IAWA J. 2006, 27, 89–97. [Google Scholar] [CrossRef]
- Gärtner, H.; Lucchinetti, S.; Schweingruber, F.H. New perspectives for wood anatomical analysis in dendrosciences: The GSL1-microtome. Dendrochronologia 2014, 32, 47–51. [Google Scholar] [CrossRef]
- Deslauriers, A.; Rossi, S.; Liang, E. Collecting and Processing Wood Microcores for Monitoring Xylogenesis. In Plant Microtechniques and Protocols; Springer International Publishing: Cham, Switzerland, 2015; pp. 417–429. ISBN 9783319199443. [Google Scholar]
- De Micco, V.; Carrer, M.; Rathgeber, C.B.K.; Camarero, J.J.; Voltas, J.; Cherubini, P.; Battipaglia, G. From xylogenesis to tree-rings: Wood traits to investigate tree response to environmental changes. IAWA J. 2019, 40, 155–182. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T. Assessment of cambial activity and xylogenesis by microsampling tree species: An example at the Alpine timberline. IAWA J. 2006, 27, 383–394. [Google Scholar] [CrossRef]
- Frankenstein, C.; Eckstein, D.; Schmitt, U. The onset of cambium activity—A matter of agreement? Dendrochronologia 2005, 23, 57–62. [Google Scholar] [CrossRef]
- Delpierre, N.; Lireux, S.; Hartig, F.; Camarero, J.J.; Cheaib, A.; Čufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Gričar, J.; et al. Chilling and forcing temperatures interact to predict the onset of wood formation in Northern Hemisphere conifers. Glob. Chang. Biol. 2019, 25, 1089–1105. [Google Scholar] [CrossRef]
- Stangler, D.F.; Mann, M.; Kahle, H.-P.; Rosskopf, E.; Fink, S.; Spiecker, H. Spatiotemporal alignment of radial tracheid diameter profiles of submontane Norway spruce. Dendrochronologia 2016, 37, 33–45. [Google Scholar] [CrossRef]
- Pya, N.; Wood, S.N. Shape constrained additive models. Stat. Comput. 2015, 25, 543–559. [Google Scholar] [CrossRef]
- R Core Team. R Programming Language. Available online: http://www.r-project.org (accessed on 1 December 2020).
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Schabenberger, O.; Pierce, F.J. Contemporary Statistical Models for the Plant and Soil Sciences; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.4.5. Available online: https://github.com/rvlenth/emmeans (accessed on 28 November 2020).
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Emerson, J.W.; Green, W.A.; Schloerke, B.; Crowley, J.; Cook, D.; Hofmann, H.; Wickham, H. The Generalized Pairs Plot. J. Comput. Graph. Stat. 2013, 22, 79–91. [Google Scholar] [CrossRef]
- Ren, P.; Rossi, S.; Camarero, J.J.; Ellison, A.M.; Liang, E.; Peñuelas, J. Critical temperature and precipitation thresholds for the onset of xylogenesis of Juniperus przewalskii in a semi-arid area of the north-eastern Tibetan Plateau. Ann. Bot. 2018, 121, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Gruber, A.; Strobl, S.; Veit, B.; Oberhuber, W. Impact of drought on the temporal dynamics of wood formation in Pinus sylvestris. Tree Physiol. 2010, 30, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Girard, M.J.; Morin, H. Lengthening of the duration of xylogenesis engenders disproportionate increases in xylem production. Glob. Chang. Biol. 2014, 20, 2261–2271. [Google Scholar] [CrossRef] [PubMed]
- Oribe, Y.; Funada, R.; Shibagaki, M.; Kubo, T. Cambial reactivation in locally heated stems of the evergreen conifer Abies sachalinensis (Schmidt) masters. Planta 2001, 212, 684–691. [Google Scholar] [CrossRef]
- Begum, S.; Kudo, K.; Rahman, M.H.; Nakaba, S.; Yamagishi, Y.; Nabeshima, E.; Nugroho, W.D.; Oribe, Y.; Kitin, P.; Jin, H.O.; et al. Climate change and the regulation of wood formation in trees by temperature. Trees-Struct. Funct. 2018, 32, 3–15. [Google Scholar] [CrossRef]
- Swidrak, I.; Gruber, A.; Kofler, W.; Oberhuber, W. Effects of environmental conditions on onset of xylem growth in Pinus sylvestris under drought. Tree Physiol. 2011, 31, 483–493. [Google Scholar] [CrossRef]
- Rossi, S.; Morin, H.; Deslauriers, A.; Plourde, P.Y. Predicting xylem phenology in black spruce under climate warming. Glob. Chang. Biol. 2011, 17, 614–625. [Google Scholar] [CrossRef]
- Rossi, S.; Anfodillo, T.; Čufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Frank, D.; Gričar, J.; Gruber, A.; Huang, J.G.; et al. Pattern of xylem phenology in conifers of cold ecosystems at the Northern Hemisphere. Glob. Chang. Biol. 2016, 22, 3804–3813. [Google Scholar] [CrossRef]
- Gričar, J.; Zupancic, M.; Čufar, K.; Koch, G.; Schmitt, U.; Oven, P. Effect of local heating and cooling on cambial activity and cell differentiation in the stem of Norway spruce (Picea abies). Ann. Bot. 2006, 97, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Gričar, J.; Zupančič, M.; Čufar, K.; Oven, P. Regular cambial activity and xylem and phloem formation in locally heated and cooled stem portions of Norway spruce. Wood Sci. Technol. 2007, 41, 463–475. [Google Scholar] [CrossRef]
- Oribe, Y.; Kubo, T. Effect of heat on cambial reactivation during winter dormancy in evergreen and deciduous conifers. Tree Physiol. 1997, 17, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Capdevielle-Vargas, R.; Estrella, N.; Menzel, A. Multiple-year assessment of phenological plasticity within a beech (Fagus sylvatica L.) stand in southern Germany. Agric. For. Meteorol. 2015, 211–212, 13–22. [Google Scholar] [CrossRef]
- Patel, V.R.; Pramod, S.; Rao, K.S. Cambial activity, annual rhythm of xylem production in relation to phenology and climatic factors and lignification pattern during xylogenesis in drum-stick tree (Moringa oleifera). Flora-Morphol. Distrib. Funct. Ecol. Plants 2014, 209, 556–566. [Google Scholar] [CrossRef]
- Chen, L.; Rossi, S.; Deslauriers, A.; Liu, J. Contrasting strategies of xylem formation between black spruce and balsam fir in Quebec, Canada. Tree Physiol. 2019, 39, 747–754. [Google Scholar] [CrossRef]
- Huang, J.; Ma, Q.; Rossi, S.; Biondi, F.; Deslauriers, A.; Fonti, P.; Liang, E.; Mäkinen, H.; Oberhuber, W.; Rathgeber, C.B.K.; et al. Photoperiod and temperature as dominant environmental drivers triggering secondary growth resumption in Northern Hemisphere conifers. Proc. Natl. Acad. Sci. USA 2020, 117, 20645–20652. [Google Scholar] [CrossRef]
- Vitasse, Y.; Signarbieux, C.; Fu, Y.H. Global warming leads to more uniform spring phenology across elevations. Proc. Natl. Acad. Sci. USA 2018, 115, 1004–1008. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; de Luis, M.; Novak, K.; Martinez del Castillo, E.; Schmitt, U.; Koch, G.; Štrus, J.; Mrak, P.; Žnidarič, M.T.; et al. Annual Cambial Rhythm in Pinus halepensis and Pinus sylvestris as Indicator for Climate Adaptation. Front. Plant Sci. 2016, 7, 1923. [Google Scholar] [CrossRef]
- Malik, R.; Rossi, S.; Sukumar, R. Variations in the timing of different phenological stages of cambial activity in Abies pindrow (Royle) along an elevation gradient in the north-western Himalaya. Dendrochronologia 2020, 59, 125660. [Google Scholar] [CrossRef]
- Prislan, P.; Koch, G.; Čufar, K.; Gričar, J.; Schmitt, U. Topochemical investigations of cell walls in developing xylem of beech (Fagus sylvatica L.). Holzforschung 2009, 63. [Google Scholar] [CrossRef]
- Rossi, S.; Morin, H.; Deslauriers, A. Causes and correlations in cambium phenology: Towards an integrated framework of xylogenesis. J. Exp. Bot. 2012, 63, 2117–2126. [Google Scholar] [CrossRef]
- Rossi, S.; Anfodillo, T.; Čufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Frank, D.; Gričar, J.; Gruber, A.; King, G.; et al. A meta-analysis of cambium phenology and growth: Linear and non-linear patterns in conifers of the northern hemisphere. Ann. Bot. 2013, 112, 1911–1920. [Google Scholar] [CrossRef]
- Fonti, P.; Von Arx, G.; García-González, I.; Eilmann, B.; Sass-Klaassen, U.; Gärtner, H.; Eckstein, D. Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytol. 2010, 185, 42–53. [Google Scholar] [CrossRef]
- Prislan, P.; Čufar, K.; De Luis, M.; Gričar, J. Precipitation is not limiting for xylem formation dynamics and vessel development in European beech from two temperate forest sites. Tree Physiol. 2018, 38, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Sass, U.; Eckstein, D. The variability of vessel size in beech (Fagus sylvatica L.) and its ecophysiological interpretation. Trees 1995, 9, 247–252. [Google Scholar] [CrossRef]
- Wagenführ, R. Holzatlas; Fachbuchverlag: Leipzig, Germany, 1996. [Google Scholar]
- Cuny, H.E.; Rathgeber, C.B.K.; Frank, D.; Fonti, P.; Fournier, M. Kinetics of tracheid development explain conifer tree-ring structure. New Phytol. 2014, 203, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Andrianantenaina, A.; Rathgeber, C.B.K.; Pérez-de-Lis, G.; Cuny, H.E.; Ruelle, J. Quantifying intra-annual dynamics of carbon sequestration in the forming wood: A novel histologic approach. Ann. For. Sci. 2019, 76, 62. [Google Scholar] [CrossRef]
- Oladi, R.; Pourtahmasi, K.; Eckstein, D.; Bräuning, A. Seasonal dynamics of wood formation in Oriental beech (Fagus orientalis Lipsky) along an altitudinal gradient in the Hyrcanian forest, Iran. Trees-Struct. Funct. 2011, 25, 425–433. [Google Scholar] [CrossRef]
- Cuny, H.E.; Fonti, P.; Rathgeber, C.B.K.; Arx, G.; Peters, R.L.; Frank, D.C. Couplings in cell differentiation kinetics mitigate air temperature influence on conifer wood anatomy. Plant. Cell Environ. 2019, 42, 1222–1232. [Google Scholar] [CrossRef]
- Cuny, H.E.; Rathgeber, C.B.K. Xylogenesis: Coniferous Trees of Temperate Forests Are Listening to the Climate Tale during the Growing Season But Only Remember the Last Words! Plant Physiol. 2016, 171, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2003; ISBN 9783540435167. [Google Scholar]
- Fernández-de-Uña, L.; Rossi, S.; Aranda, I.; Fonti, P.; González-González, B.D.; Cañellas, I.; Gea-Izquierdo, G. Xylem and Leaf Functional Adjustments to Drought in Pinus sylvestris and Quercus pyrenaica at Their Elevational Boundary. Front. Plant Sci. 2017, 8, 1200. [Google Scholar] [CrossRef] [PubMed]
- Aranda, I.; Cano, F.J.; Gasco, A.; Cochard, H.; Nardini, A.; Mancha, J.A.; Lopez, R.; Sanchez-Gomez, D. Variation in photosynthetic performance and hydraulic architecture across European beech (Fagus sylvatica L.) populations supports the case for local adaptation to water stress. Tree Physiol. 2015, 35, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early warnings of tree dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef]
- Martínez-Sancho, E.; Vásconez Navas, L.K.; Seidel, H.; Dorado-Liñán, I.; Menzel, A. Responses of Contrasting Tree Functional Types to Air Warming and Drought. Forests 2017, 8, 450. [Google Scholar] [CrossRef]
- Lebourgeois, F.; Gomez, N.; Pinto, P.; Mérian, P. Mixed stands reduce Abies alba tree-ring sensitivity to summer drought in the Vosges mountains, western Europe. For. Ecol. Manag. 2013, 303, 61–71. [Google Scholar] [CrossRef]
- Liu, X.; Wang, C.; Zhao, J. Seasonal Drought Effects on Intra-Annual Stem Growth of Taiwan Pine along an Elevational Gradient in Subtropical China. Forests 2019, 10, 1128. [Google Scholar] [CrossRef]
- Michelot, A.; Simard, S.; Rathgeber, C.B.K.; Dufrêsne, E.; Damesin, C. Comparing the intra-annual wood formation of three European species (Fagus sylvatica, Quercus petraea and Pinus sylvestris) as related to leaf phenology and non-structural carbohydrate dynamics. Tree Physiol. 2012, 32, 1033–1045. [Google Scholar] [CrossRef]
- Jyske, T.; Mäkinen, H.; Kalliokoski, T.; Nöjd, P. Intra-annual tracheid production of Norway spruce and Scots pine across a latitudinal gradient in Finland. Agric. For. Meteorol. 2014, 194, 241–254. [Google Scholar] [CrossRef]
- Zhang, J.; Gou, X.; Manzanedo, R.D.; Zhang, F.; Pederson, N. Cambial phenology and xylogenesis of Juniperus przewalskii over a climatic gradient is influenced by both temperature and drought. Agric. For. Meteorol. 2018, 260–261, 165–175. [Google Scholar] [CrossRef]
- Lupi, C.; Morin, H.; Deslauriers, A.; Rossi, S. Xylogenesis in black spruce: Does soil temperature matter? Tree Physiol. 2012, 32, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Ren, P.; Ziaco, E.; Rossi, S.; Biondi, F.; Prislan, P.; Liang, E. Growth rate rather than growing season length determines wood biomass in dry environments. Agric. For. Meteorol. 2019, 271, 46–53. [Google Scholar] [CrossRef]
- Kahle, H.-P. Impact of the drought in 2003 on intra- and inter-annual stem radial growth of beech and spruce along an altitudinal gradient in the Black Forest, Germany. Tree Rings Archeol. Clim. Ecol. 2006, 4, 151–163. [Google Scholar]
- Gričar, J.; Čufar, K. Seasonal dynamics of phloem and xylem formation in silver fir and Norway spruce as affected by drought. Russ. J. Plant Physiol. 2008, 55, 538–543. [Google Scholar] [CrossRef]
- Balducci, L.; Cuny, H.E.; Rathgeber, C.B.K.; Deslauriers, A.; Giovannelli, A.; Rossi, S. Compensatory mechanisms mitigate the effect of warming and drought on wood formation. Plant. Cell Environ. 2016, 39, 1338–1352. [Google Scholar] [CrossRef]
- Camarero, J.J.; Olano, J.M.; Parras, A. Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 2010, 185, 471–480. [Google Scholar] [CrossRef]
- Maxime, C.; Hendrik, D. Effects of climate on diameter growth of co-occurring Fagus sylvatica and Abies alba along an altitudinal gradient. Trees-Struct. Funct. 2011, 25, 265–276. [Google Scholar] [CrossRef]
- Mattes, A. Zuwachs und Konkurrenz in Buchen-/Eichenmischbeständen unter sich Ändernden Klimabedingungen. Ph.D. Thesis, Albert-Ludwigs-Universität Freiburg, Breisgau, Germany, 2014. [Google Scholar]
- Vitali, V.; Büntgen, U.; Bauhus, J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob. Chang. Biol. 2017, 23, 5108–5119. [Google Scholar] [CrossRef]
- Marion, L.; Gričar, J.; Oven, P. Wood formation in urban Norway maple trees studied by the micro-coring method. Dendrochronologia 2007, 25, 97–102. [Google Scholar] [CrossRef]
- Oribe, Y.; Funada, R.; Kubo, T. Relationships between cambial activity, cell differentiation and the localization of starch in storage tissues around the cambium in locally heated stems of Abies sachalinensis (Schmidt) Masters. Trees 2003, 17, 185–192. [Google Scholar] [CrossRef]
- Oberhuber, W.; Gruber, A. Climatic influences on intra-annual stem radial increment of Pinus sylvestris (L.) exposed to drought. Trees-Struct. Funct. 2010, 24, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Cantero, A.; Sánchez-Salguero, R.; Sánchez-Miranda, A.; Granda, E.; Serra-Maluquer, X.; Ibáñez, R. Forest growth responses to drought at short- and long-term scales in Spain: Squeezing the stress memory from tree rings. Front. Ecol. Evol. 2018, 6. [Google Scholar] [CrossRef]
- Eilmann, B.; Rigling, A. Tree-growth analyses to estimate tree species ’ drought tolerance. Tree Physiol. 2012, 32, 178–187. [Google Scholar] [CrossRef] [PubMed]
- John, R.; Grüner, J.; Seitz, G.; Delb, H. Buchen in Südwestdeutschland Leiden unter dem Trockenstress der Vorjahre; FVA Waldschutz-INFO: Freiburg, Germany, 2019. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevation | Stand Basal Area [m2/ha] | Species | Number of Trees | DBHob [cm] | Tree Height [m] | ||
|---|---|---|---|---|---|---|---|
| low: 450 m | 20.5 ± 1.2 | European beech | 6 | 47.7 | ±1.5 | 30.2 | ±1.8 |
| Scots pine | 6 | 41.3 | ±1.3 | 30.5 | ±1.4 | ||
| silver fir | 6 | 51.1 | ±2.1 | 25.0 | ±2.0 | ||
| low-med: 650 m | 24.4 ± 1.9 | European beech | 6 | 48.7 | ±3.4 | 30.4 | ±0.8 |
| Scots pine | 6 | 53.5 | ±3.5 | 30.7 | ±1.0 | ||
| silver fir | 6 | 58.8 | ±3.2 | 30.6 | ±1.2 | ||
| med-high: 850 m | 25.3 ± 1.4 | European beech | 6 | 35.9 | ±2.8 | 24.7 | ±1.8 |
| Scots pine | 6 | 42.2 | ±1.9 | 24.9 | ±0.5 | ||
| silver fir | 6 | 58.1 | ±1.8 | 26.5 | ±1.1 | ||
| high: 1100 m | 24.7 ± 1.9 | European beech | 6 | 31.9 | ±1.6 | 23.5 | ±2.3 |
| silver fir | 6 | 37.6 | ±4.4 | 22.6 | ±3.8 | ||
| Elevation | Plot | Tair [°C] | Tair Deviance in 2018 [K] | Annual P [mm] | P Deviance in 2018 [mm] |
|---|---|---|---|---|---|
| low: 450 m | 1 | 10.0 | +1.4 | 1072.3 | −241.3 |
| 2 | 9.1 | +1.4 | 1123.3 | −223.7 | |
| low-med: 650 m | 1 | 8.5 | +1.5 | 1281.8 | −197.5 |
| 2 | 8.9 | +1.5 | 1237.1 | −196.9 | |
| med-high: 850 m | 1 | 6.6 | +1.5 | 1478.0 | −181.6 |
| 2 | 7.4 | +1.5 | 1554.9 | −355.3 | |
| high: 1100 m | 1 | 5.9 | +1.5 | 1791.7 | −493.4 |
| 2 | 5.2 | +1.5 | 1616.7 | −167.2 |
| Response | Fixed Effects | p | Response | Fixed Effects | p |
|---|---|---|---|---|---|
| bE R2: 0.65 | Elevation | 0.009 | cE R2: 0.58 | Elevation | 0.080 |
| Species | 0.089 | Species | <0.001 | ||
| Species × Elevation | 0.051 | Species × Elevation | 0.433 | ||
| GDD R2: 0.54 | Elevation | 0.104 | cW R2: 0.65 | Elevation | 0.399 |
| Species | 0.244 | Species | <0.001 | ||
| Species × Elevation | 0.065 | Species × Elevation | 0.031 | ||
| bW R2: 0.56 | Elevation | 0.020 | ΔE R2: 0.58 | Elevation | 0.125 |
| Species | 0.007 | Species | <0.001 | ||
| Species × Elevation | 0.015 | Species × Elevation | 0.249 | ||
| bM R2: 0.66 | Elevation | 0.011 | ΔW R2: 0.63 | Elevation | 0.119 |
| Species | <0.001 | Species | <0.001 | ||
| Species × Elevation | 0.008 | Species × Elevation | 0.023 | ||
| tmax R2: 0.44 | Elevation | 0.101 | ΔX R2: 0.66 | Elevation | 0.105 |
| Species | 0.002 | Species | <0.001 | ||
| Species × Elevation | 0.131 | Species × Elevation | 0.053 |
| Response | Fixed Effects | p |
|---|---|---|
| mean rate R2: 0.52 | Elevation | 0.704 |
| Species | <0.001 | |
| Species × Elevation | 0.008 | |
| max rate R2: 0.52 | Elevation | 0.909 |
| Species | <0.001 | |
| Species × Elevation | 0.035 | |
| TRW R2: 0.34 | Elevation | 0.634 |
| Species | 0.009 | |
| Species × Elevation | 0.037 | |
| Resistance R2: 0.40 | Elevation | 0.159 |
| Species | <0.001 | |
| Species × Elevation | 0.067 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larysch, E.; Stangler, D.F.; Nazari, M.; Seifert, T.; Kahle, H.-P. Xylem Phenology and Growth Response of European Beech, Silver Fir and Scots Pine along an Elevational Gradient during the Extreme Drought Year 2018. Forests 2021, 12, 75. https://doi.org/10.3390/f12010075
Larysch E, Stangler DF, Nazari M, Seifert T, Kahle H-P. Xylem Phenology and Growth Response of European Beech, Silver Fir and Scots Pine along an Elevational Gradient during the Extreme Drought Year 2018. Forests. 2021; 12(1):75. https://doi.org/10.3390/f12010075
Chicago/Turabian StyleLarysch, Elena, Dominik Florian Stangler, Mona Nazari, Thomas Seifert, and Hans-Peter Kahle. 2021. "Xylem Phenology and Growth Response of European Beech, Silver Fir and Scots Pine along an Elevational Gradient during the Extreme Drought Year 2018" Forests 12, no. 1: 75. https://doi.org/10.3390/f12010075
APA StyleLarysch, E., Stangler, D. F., Nazari, M., Seifert, T., & Kahle, H.-P. (2021). Xylem Phenology and Growth Response of European Beech, Silver Fir and Scots Pine along an Elevational Gradient during the Extreme Drought Year 2018. Forests, 12(1), 75. https://doi.org/10.3390/f12010075