Long-Term Effects of Fuels Treatments, Overstory Structure, and Wildfire on Tree Regeneration in Dry Forests of Central Washington


Abstract
1. Introduction
2. Materials and Methods
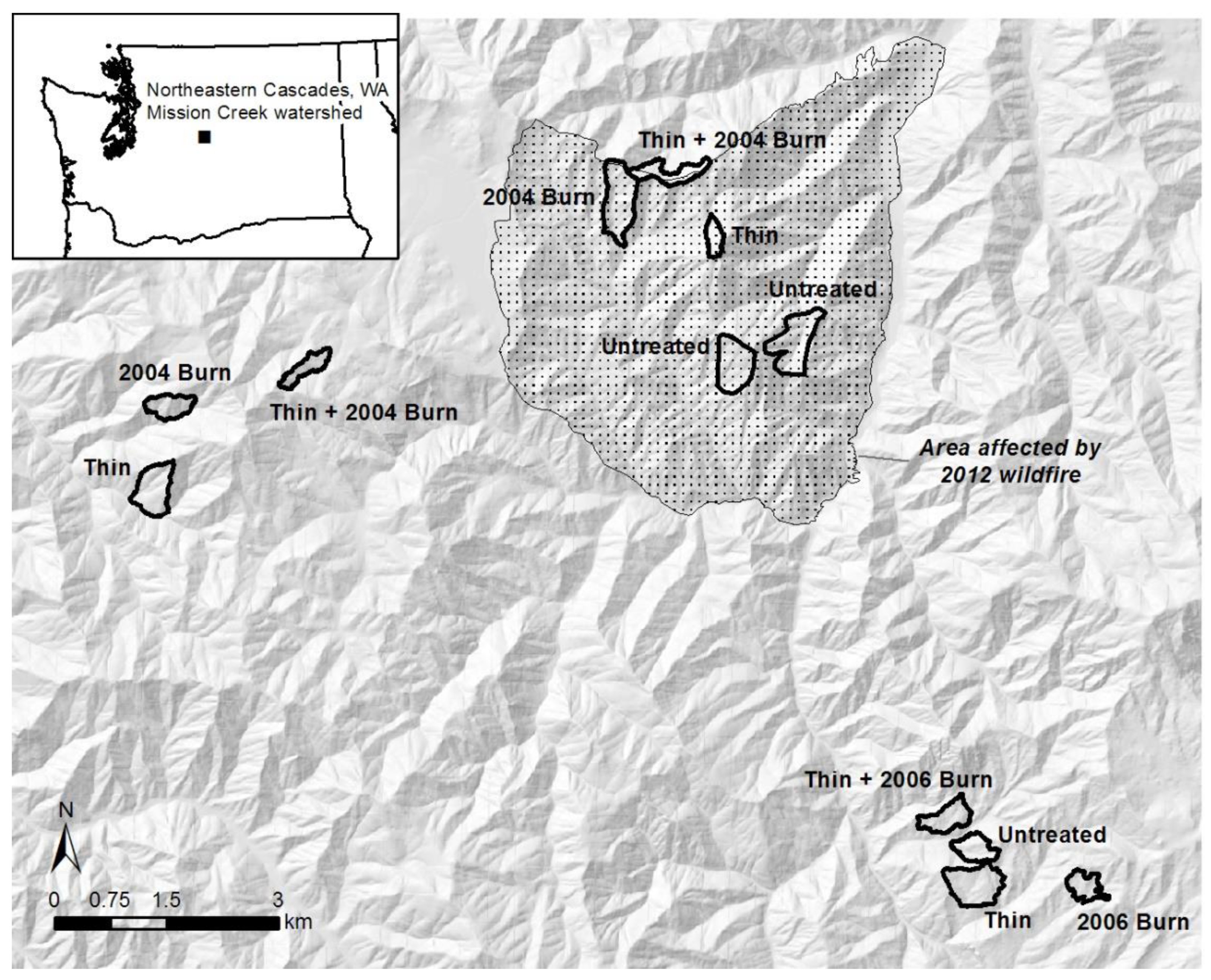
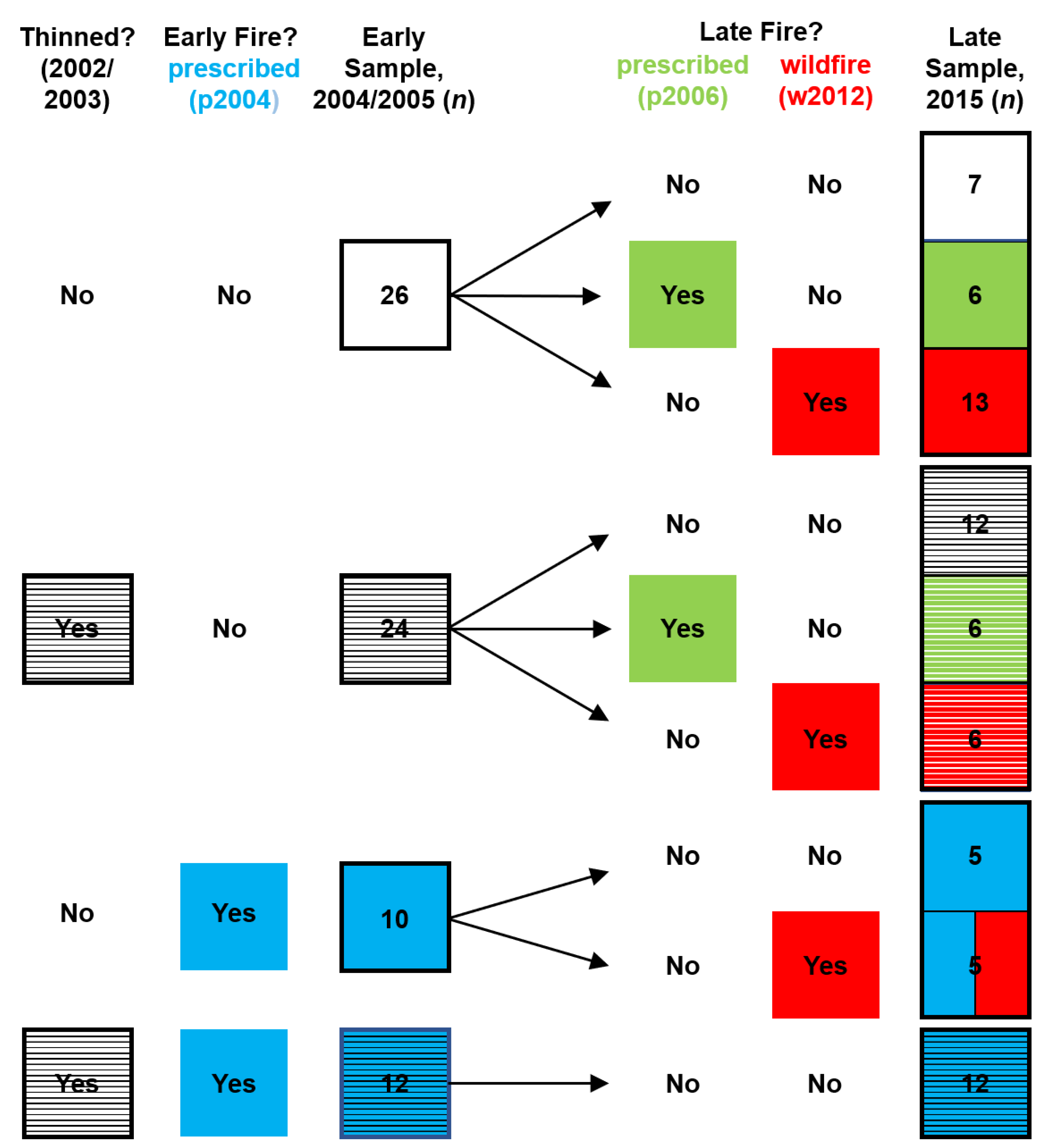
2.1. Study Sites and Treatments
2.2. Data Collection and Processing
2.2.1. Overstory Structure
2.2.2. Regeneration
2.3. Statistical Analyses
OA × Late fire + Thin × Early fire + Thin × Late fire + Early fire × Late fire
3. Results
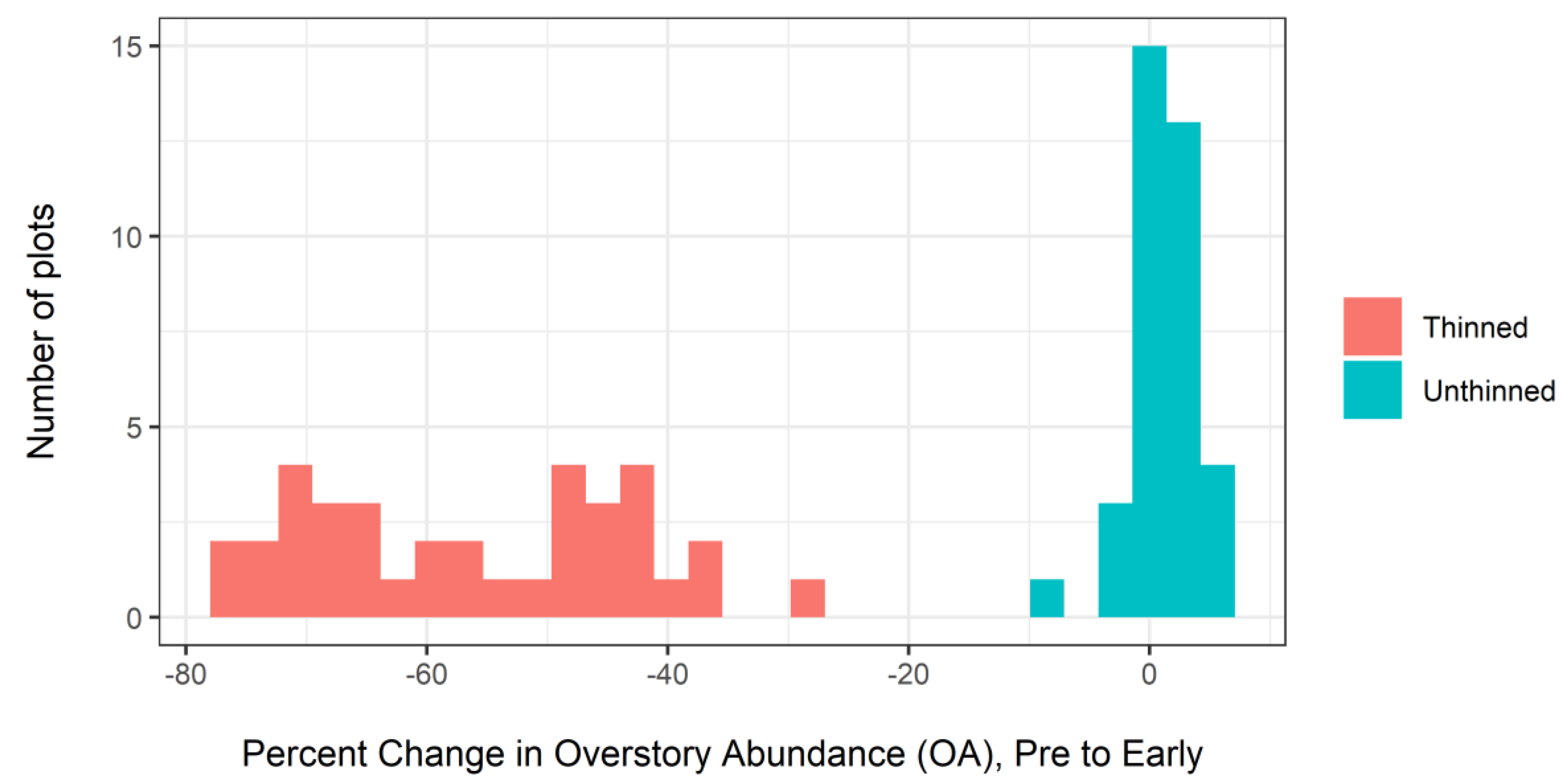
3.1. Overstory Dynamics: Early–Late Interval (2004–2015)
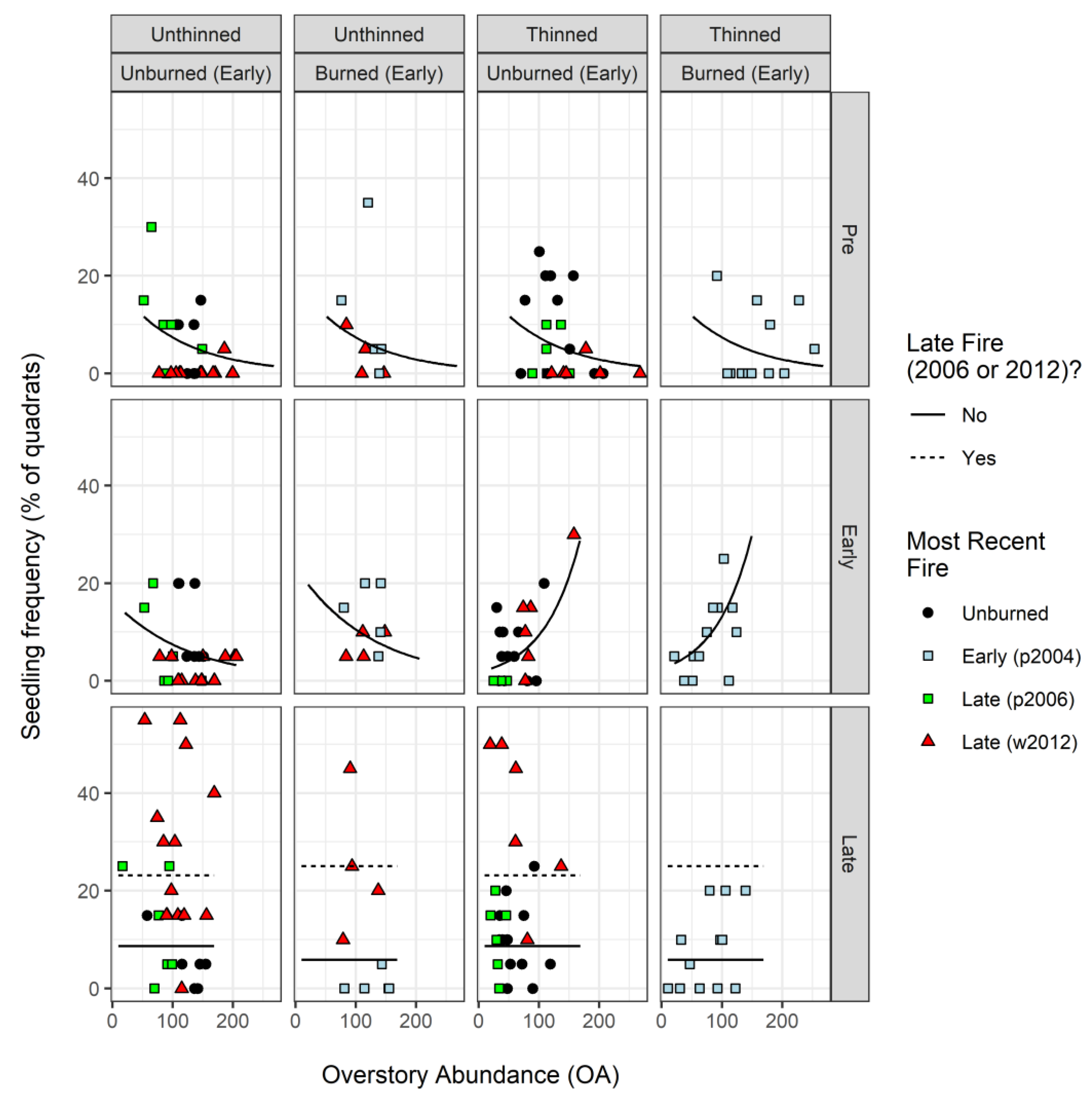
3.2. Frequency of Seedlings (<1 m Tall): Pre, Early, and Late Samples (2000/2001, 2005, 2015)
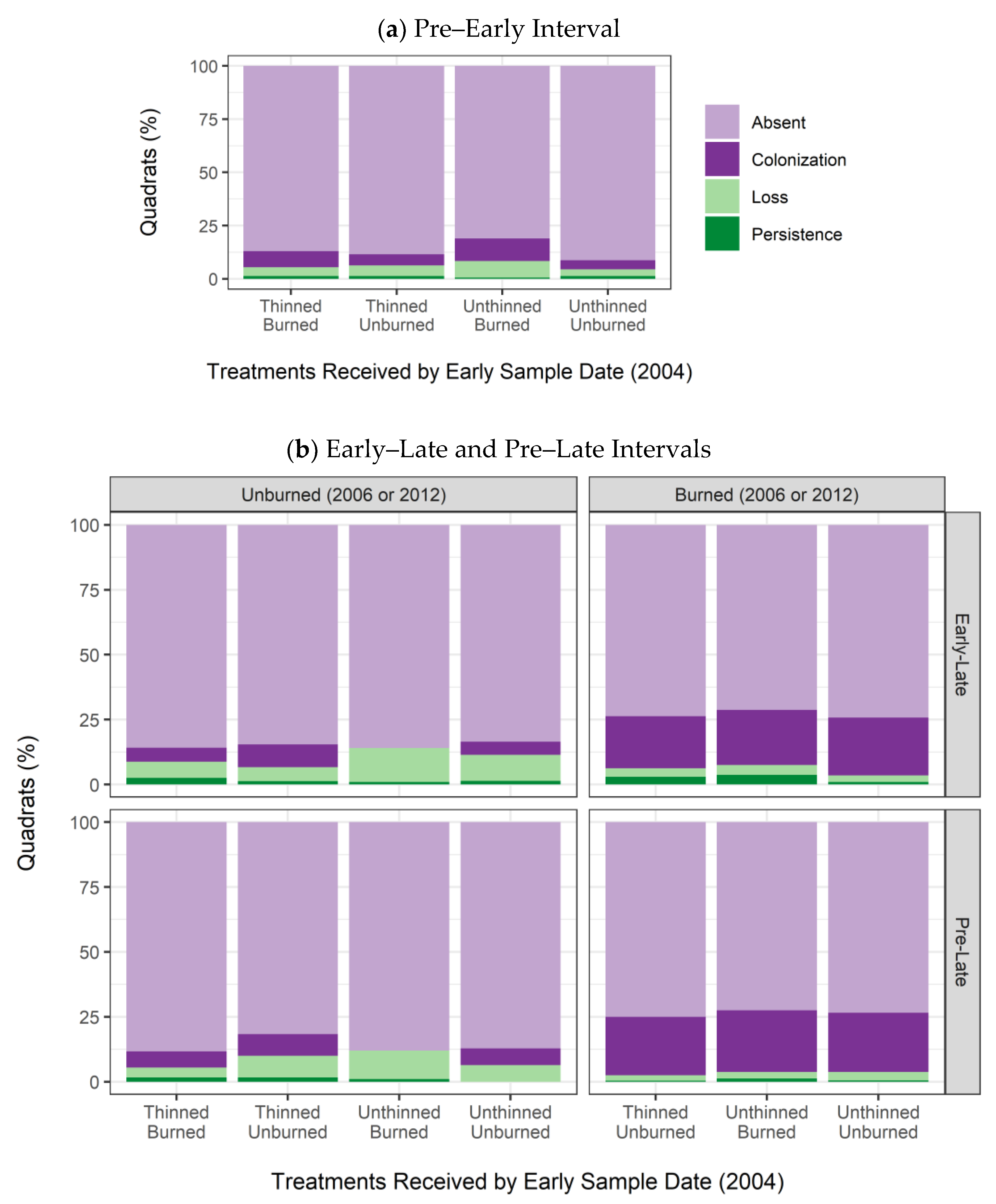
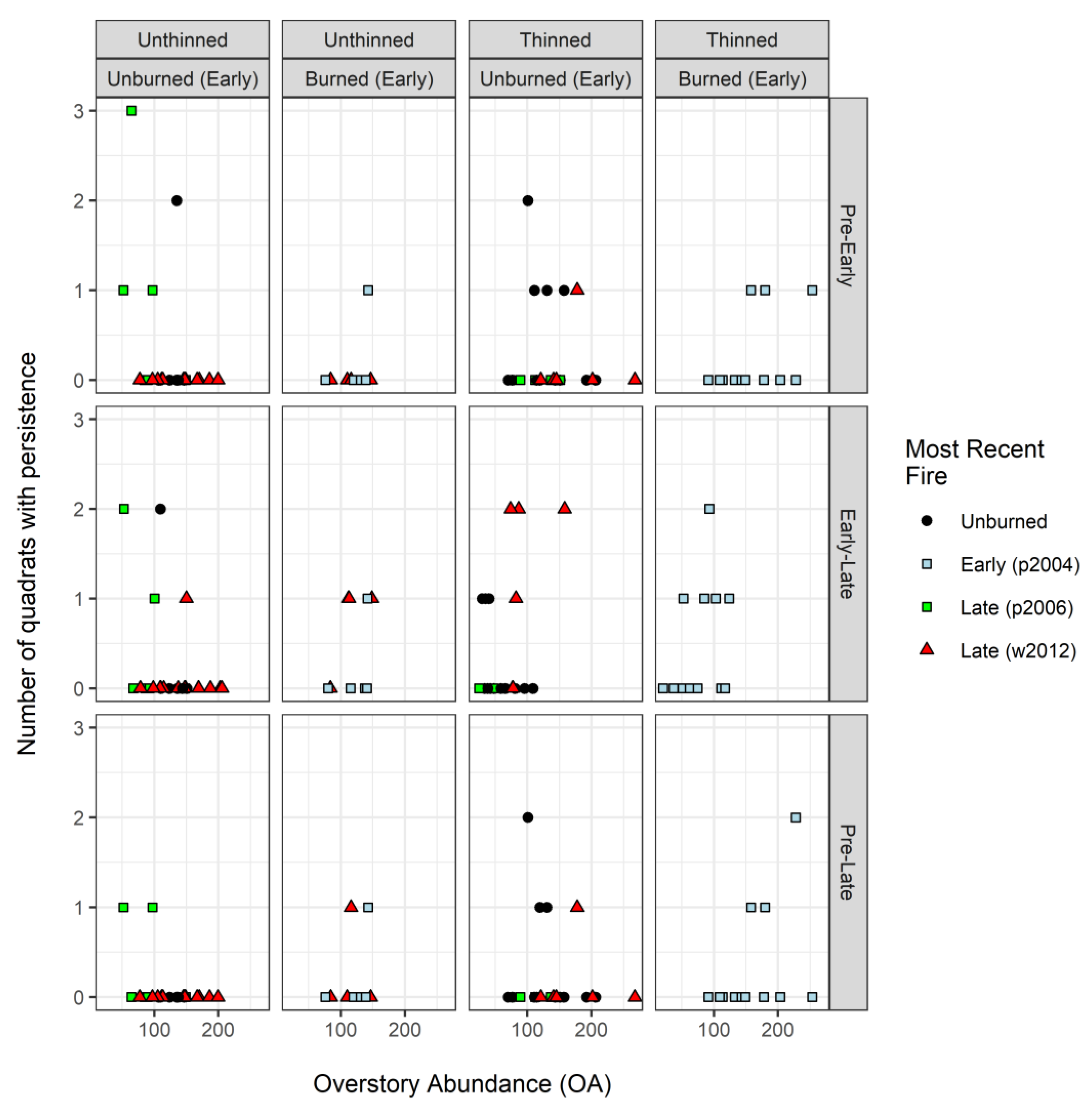
3.3. Regeneration Dynamics: Colonization and Persistence of Seedlings (<1 m Tall)
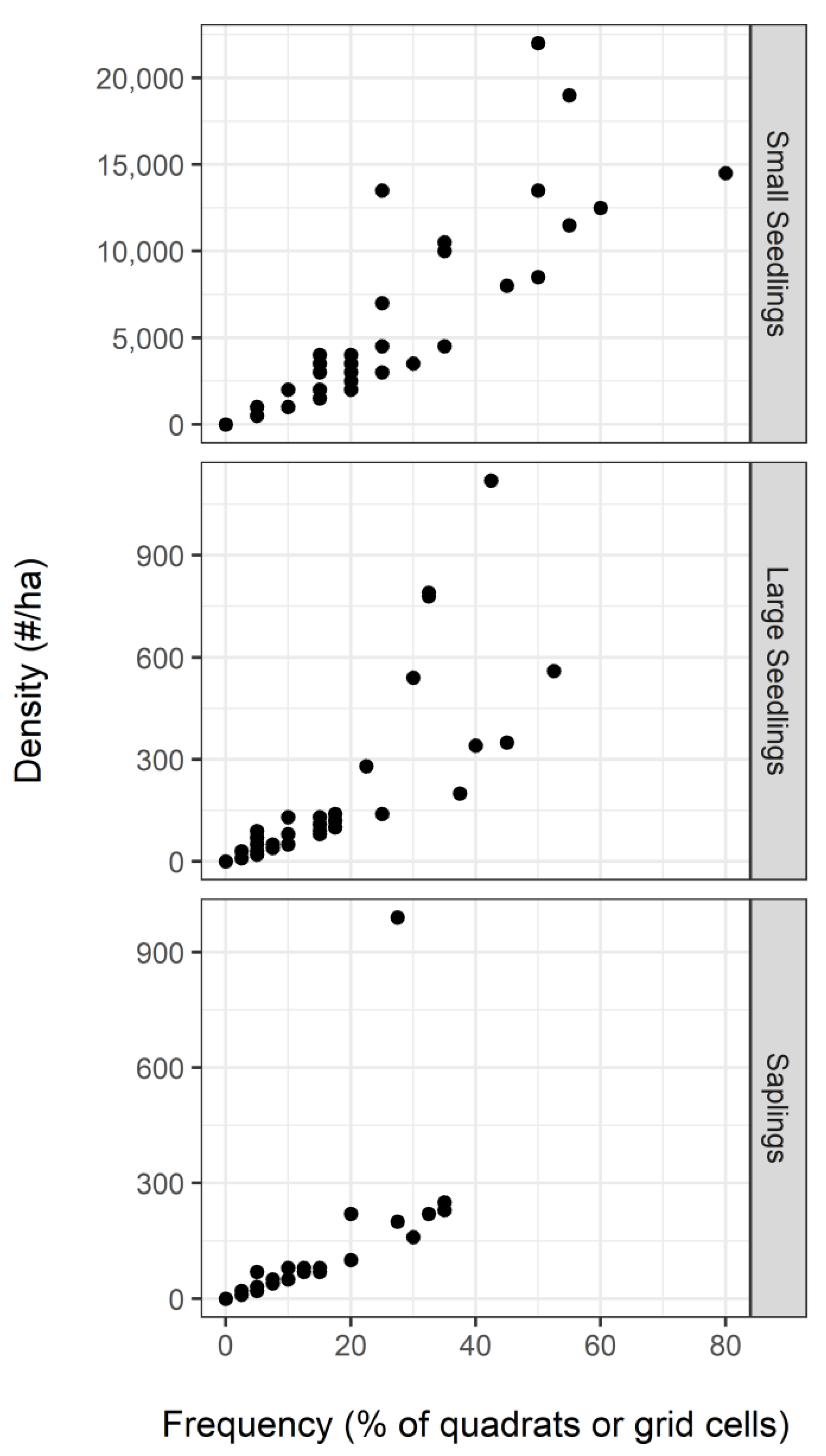
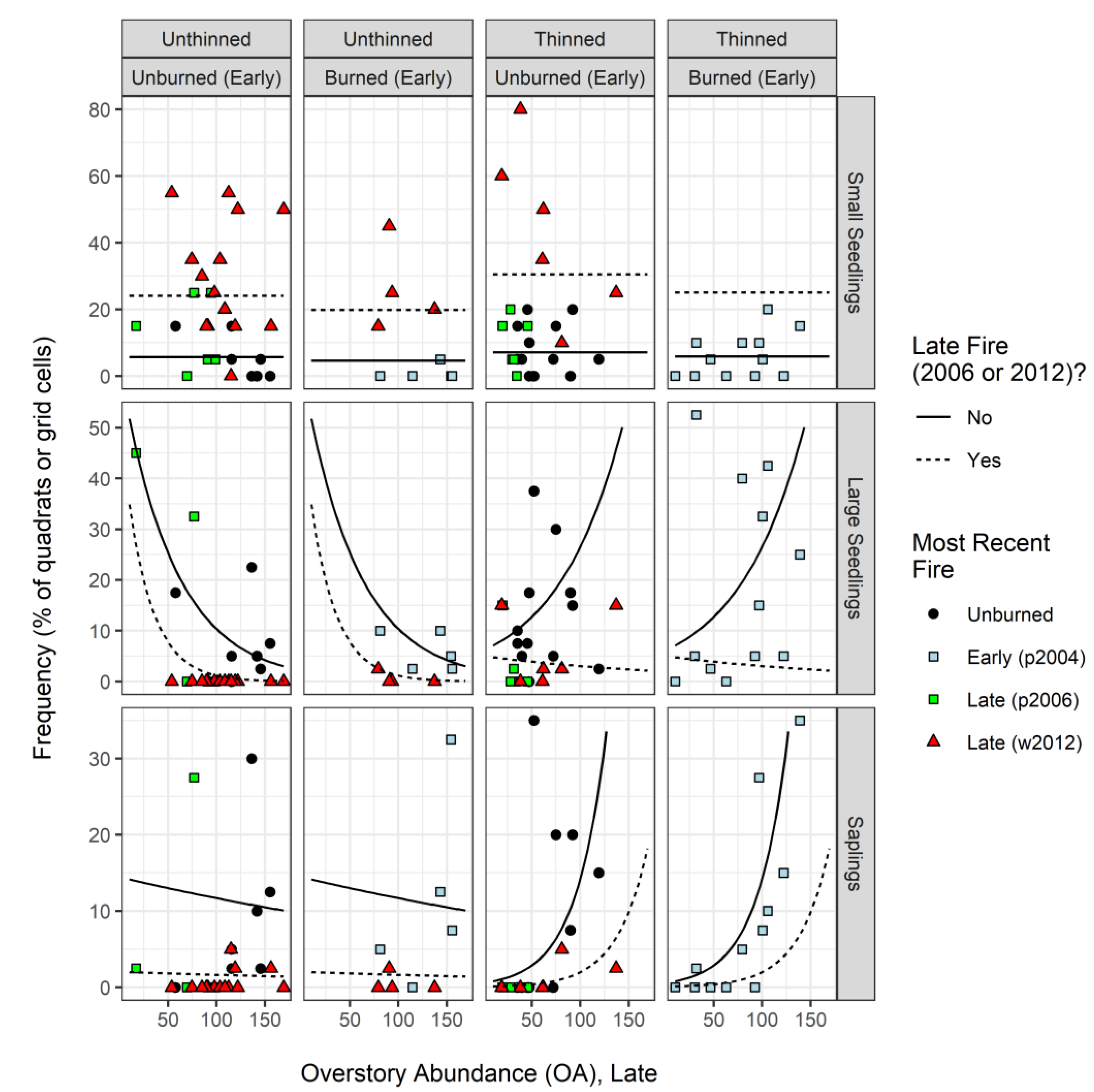
3.4. Frequency of Regeneration by Height Class: Late Sample (2015)
4. Discussion
4.1. Seedling Relationships with Overstory Structure and Disturbance
4.2. Responses to Thinning and Fire History among Regeneration Height Classes
4.3. Implications of Fuels Treatments and Wildfire for Structural Change and Stand Development
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
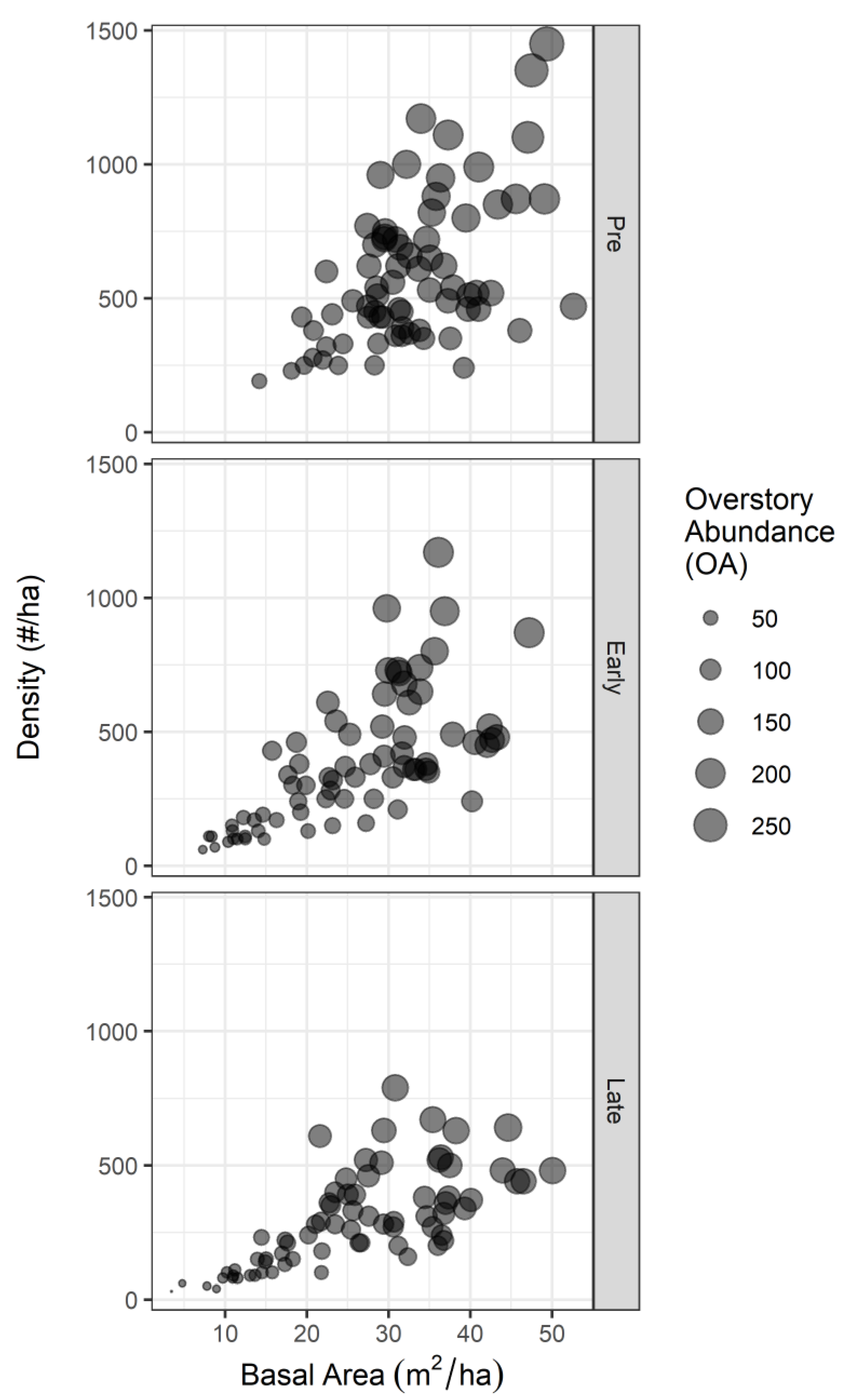
Appendix A.1. Relationships among Overstory Abundance (OA), Density, and Basal Area

Appendix A.2. Pre-Treatment Vegetation Structure and Environment

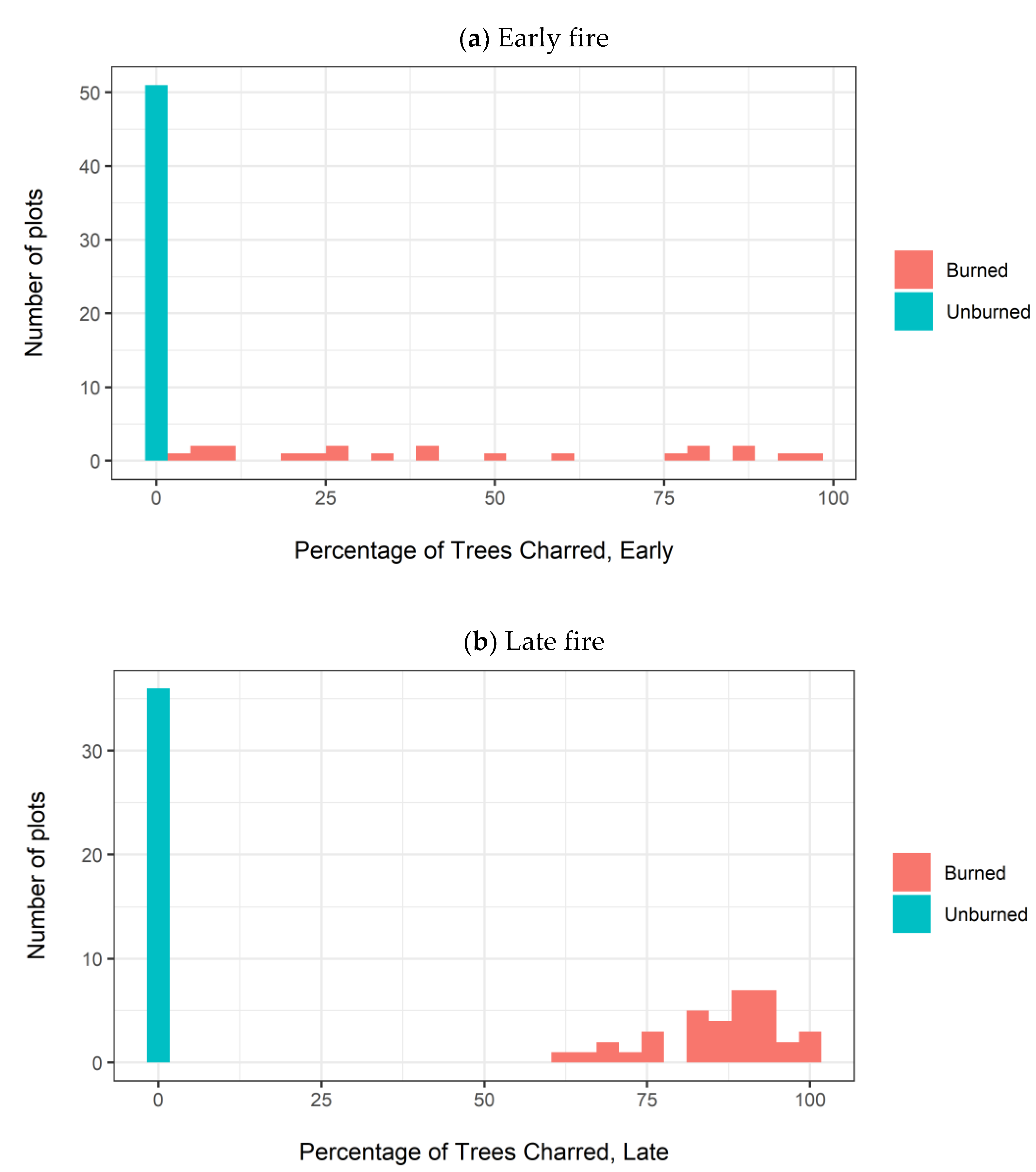
Appendix A.3. Thinning Intensity and Burn Severity


Appendix A.4. Detailed Model Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Response | Sampling Date or Interval | Table |
|---|---|---|---|
| Overstory dynamics | Ingrowth | Early–Late | A2 |
| Mortality | Early–Late | A2 | |
| BAI | Early–Late | A3 | |
| Seedling frequency (<1 m tall) | Seedlings | Pre | A4 |
| Early | A4 | ||
| Late | A4 | ||
| Seedling dynamics | Colonization | Pre–Early | A5 |
| Early–Late | A5 | ||
| Pre–Late | A5 | ||
| Regeneration frequency (by height class) | Small seedlings | Late | A6 |
| Large seedlings | Late | A6 | |
| Saplings | Late | A6 |
| Response | Term | Coefficient | Deviance | p | |
|---|---|---|---|---|---|
| Ingrowth | Intercept | −2.293 | |||
| Overstory Abundance (OA) | 0.146 | 0.13 | 0.723 | ||
| Thin | 1.999 | 5.56 | 0.018 | * | |
| Early Fire (EF) | −4.817 | 2.06 | 0.151 | ||
| Late Fire (LF) | 1.849 | 3.36 | 0.067 | ||
| OA x Thin | −0.059 | 1.89 | 0.169 | ||
| OA x EF | 0.398 | 1.17 | 0.28 | ||
| OA x LF | −0.158 | 0.14 | 0.704 | ||
| Thin x EF | 1.761 | 4.08 | 0.043 | * | |
| Thin x LF | −2.498 | 1.13 | 0.289 | ||
| EF x LF | −1.125 | 0.42 | 0.515 | ||
| Mortality | Intercept | 0.643 | |||
| Overstory Abundance (OA) | 0.098 | 76.33 | <0.001 | *** | |
| Thin | −2.481 | 0.55 | 0.459 | ||
| Early Fire (EF) | 0.154 | 2.47 | 0.116 | ||
| Late Fire (LF) | 0.025 | 33.16 | <0.001 | *** | |
| OA x Thin | 0.136 | 1.37 | 0.241 | ||
| OA x EF | −0.107 | 9.99 | 0.002 | ** | |
| OA x LF | 0.064 | 3.27 | 0.071 | ||
| Thin x EF | 2.28 | 1.02 | 0.313 | ||
| Thin x LF | 1.945 | 2.95 | 0.086 | ||
| EF x LF | 1.029 | 2.32 | 0.128 |
| Term | Coefficient | Sums of Squares | F Value | p | |
|---|---|---|---|---|---|
| Intercept | 0.236 | ||||
| Overstory Abundance (OA) | −0.008 | 1.22 | 6.0 | 0.017 | * |
| Thin | −0.055 | 0.21 | 1.0 | 0.317 | |
| Early Fire (EF) | −0.2 | 0.86 | 4.2 | 0.045 | * |
| Late Fire (LF) | 0.024 | 1.84 | 9.0 | 0.004 | ** |
| OA x Thin | 0.035 | 0.65 | 3.2 | 0.079 | |
| OA x EF | 0.038 | 1.75 | 8.6 | 0.005 | ** |
| OA x LF | −0.042 | 0.09 | 0.4 | 0.517 | |
| Thin x EF | −0.374 | 0.32 | 1.6 | 0.217 | |
| Thin x LF | −0.14 | 0.07 | 0.3 | 0.559 | |
| EF x LF | 0.23 | 0.08 | 0.4 | 0.537 |
| Sampling Date | Term | Coefficient | Deviance | p | |
|---|---|---|---|---|---|
| Pre | Intercept | 1.339 | |||
| Overstory Abundance (OA) | −0.095 | 5.03 | 0.025 | * | |
| Early | Intercept | 1.171 | |||
| Overstory Abundance (OA) | −0.087 | 1.45 | 0.228 | ||
| Thin | −2.053 | 1.56 | 0.212 | ||
| Early Fire (EF) | 0.166 | 4.7 | 0.030 | * | |
| OA x Thin | 0.249 | 16.68 | <0.001 | *** | |
| OA x EF | 0.041 | 1.21 | 0.271 | ||
| Thin x EF | −0.488 | 1.04 | 0.309 | ||
| Late | Intercept | 1.287 | |||
| Overstory Abundance (OA) | −0.089 | 0.53 | 0.468 | ||
| Thin | −0.34 | 3.52 | 0.061 | ||
| Early Fire (EF) | −3.764 | 6.06 | 0.014 | * | |
| Late Fire (LF) | 0.407 | 26.19 | <0.001 | *** | |
| OA x Thin | 0.048 | 2.34 | 0.126 | ||
| OA x EF | 0.154 | 0.17 | 0.676 | ||
| OA x LF | 0.073 | 2.64 | 0.104 | ||
| Thin x EF | 2.311 | 0.48 | 0.488 | ||
| Thin x LF | 0.013 | 0.42 | 0.517 | ||
| EF x LF | 2.308 | 4.97 | 0.026 | * |
| Sampling Interval | Term | Coefficient | Deviance | p | |
|---|---|---|---|---|---|
| Pre-Early | Intercept | −2.527 | |||
| Overstory Abundance (OA) | −0.046 | 10.44 | 0.001 | *** | |
| Thin | −2.721 | 0.63 | 0.426 | ||
| Early Fire (EF) | 1.15 | 7.2 | 0.007 | ** | |
| OA x Thin | 0.195 | 12.25 | <0.001 | *** | |
| OA x EF | −0.02 | 2.25 | 0.134 | ||
| Thin x EF | −0.689 | 1.45 | 0.228 | ||
| Early-Late | Intercept | −2.394 | |||
| Overstory Abundance (OA) | −0.037 | 6.22 | 0.013 | * | |
| Thin | −0.307 | 0 | 0.99 | ||
| Early Fire (EF) | −38.482 | 11.62 | 0.001 | *** | |
| Late Fire (LF) | 0.712 | 29.79 | <0.001 | *** | |
| OA x Thin | 0.095 | 2.6 | 0.107 | ||
| OA x EF | 0.148 | 0.75 | 0.387 | ||
| OA x LF | 0.054 | 3.58 | 0.059 | ||
| Thin x EF | 36.583 | 0.43 | 0.513 | ||
| Thin x LF | −0.284 | 1.19 | 0.275 | ||
| EF x LF | 36.802 | 6.81 | 0.009 | ** | |
| Pre-Late | Intercept | −2.138 | |||
| Overstory Abundance (OA) | −0.043 | 2.97 | 0.085 | ||
| Thin | −0.583 | 4.87 | 0.027 | * | |
| Early Fire (EF) | −36.185 | 12.03 | 0.001 | *** | |
| Late Fire (LF) | 0.612 | 30.73 | <0.001 | *** | |
| OA x Thin | 0.069 | 2.54 | 0.111 | ||
| OA x EF | 0.137 | 1.49 | 0.223 | ||
| OA x LF | 0.049 | 0 | 0.945 | ||
| Thin x EF | 33.267 | 0.78 | 0.377 | ||
| Thin x LF | −0.513 | 3.97 | 0.046 | * | |
| EF x LF | 34.651 | 8.35 | 0.004 | ** |
| Height Class | Term | Coefficient | Deviance | p | |
|---|---|---|---|---|---|
| Small seedlings | Intercept | 1.196 | |||
| Overstory Abundance (OA) | −0.093 | 0.54 | 0.461 | ||
| Thin | −0.336 | 4.01 | 0.045 | * | |
| Early Fire (EF) | −3.573 | 7.69 | 0.006 | ** | |
| Late Fire (LF) | 0.411 | 32.87 | <0.001 | *** | |
| OA x Thin | 0.037 | 1.04 | 0.307 | ||
| OA x EF | 0.151 | 0.09 | 0.77 | ||
| OA x LF | 0.092 | 2.19 | 0.139 | ||
| Thin x EF | 2.134 | 0.18 | 0.676 | ||
| Thin x LF | 0.234 | 0.06 | 0.802 | ||
| EF x LF | 2.162 | 3.81 | 0.051 | ||
| Large seedlings | Intercept | 3.577 | |||
| Overstory Abundance (OA) | −0.187 | 2.15 | 0.142 | ||
| Thin | −2.997 | 7.63 | 0.006 | ** | |
| Early Fire (EF) | 0.951 | 1.97 | 0.161 | ||
| Late Fire (LF) | −0.221 | 8.01 | 0.005 | ** | |
| OA x Thin | 0.365 | 16.65 | <0.001 | *** | |
| OA x EF | −0.088 | 0.35 | 0.552 | ||
| OA x LF | −0.233 | 5.58 | 0.018 | * | |
| Thin x EF | −0.22 | 0.01 | 0.928 | ||
| Thin x LF | 0.464 | 0.33 | 0.568 | ||
| EF x LF | −1.133 | 0.68 | 0.409 | ||
| Saplings | Intercept | −1.512 | |||
| Overstory Abundance (OA) | 0.207 | 15.76 | <0.001 | *** | |
| Thin | −0.484 | 7.52 | 0.006 | ** | |
| Early Fire (EF) | 0.796 | 0.06 | 0.811 | ||
| Late Fire (LF) | 1.859 | 9.49 | 0.002 | ** | |
| OA x Thin | 0.264 | 7.9 | 0.005 | ** | |
| OA x EF | −0.047 | 1.79 | 0.181 | ||
| OA x LF | −0.27 | 0.11 | 0.738 | ||
| Thin x EF | −1.711 | 0 | 0.954 | ||
| Thin x LF | −2.633 | 1.71 | 0.191 | ||
| EF x LF | −1.469 | 0.8 | 0.371 |
References
- Everett, R.L.; Schellhaas, R.; Keenum, D.; Spurbeck, D.; Ohlson, P. Fire history in the ponderosa pine/Douglas-fir forests on the east slope of the Washington Cascades. For. Ecol. Manag. 2000, 129, 207–225. [Google Scholar] [CrossRef]
- Hessburg, P.F.; Smith, B.G.; Salter, R.B.; Ottmar, R.D.; Alvarado, E. Recent changes (1930s–1990s) in spatial patterns of interior northwest forests, USA. For. Ecol. Manag. 2000, 136, 53–83. [Google Scholar] [CrossRef]
- Hessburg, P.F.; Agee, J.K.; Franklin, J.F. Dry forests and wildland fires of the inland Northwest USA: Contrasting the landscape ecology of the pre-settlement and modern eras. For. Ecol. Manag. 2005, 211, 117–139. [Google Scholar] [CrossRef]
- Merschel, A.; Spies, T.; Heyerdahl, E. Mixed-conifer forests of central Oregon: Effects of logging and fire exclusion vary with environment. Ecol. Appl. 2014, 24, 1670–1688. [Google Scholar] [CrossRef] [PubMed]
- Haugo, R.; Zanger, C.; DeMeo, T.; Ringo, C.; Shlisky, A.; Blankenship, K.; Simpson, M.; Mellen-McLean, K.; Kertis, J.; Stern, M. A new approach to evaluate forest structure restoration needs across Oregon and Washington, USA. For. Ecol. Manag. 2015, 335, 37–50. [Google Scholar] [CrossRef]
- Brown, R.T.; Agee, J.K.; Franklin, J.F. Forest restoration and fire: Principles in the context of place. Conserv. Biol. 2004, 18, 903–912. [Google Scholar] [CrossRef]
- Martinson, E.J.; Omi, P.N. Fuel Treatments and Fire Severity: A Meta-Analysis; Res. Pap. RMRS-RP-103WWW; USDA/FS Rocky Mountain Res. Sta.: Fort Collins, CO, USA, 2013.
- Harrod, R.J.; Peterson, D.W.; Povak, N.A.; Dodson, E.K. Thinning and prescribed fire effects on overstory tree and snag structure in dry coniferous forests of the interior Pacific Northwest. For. Ecol. Manag. 2009, 258, 712–721. [Google Scholar] [CrossRef]
- McIver, J.D.; Stephens, S.L.; Agee, J.K.; Barbour, J.; Boerner, R.E.J.; Edminster, C.B.; Erickson, K.L.; Farris, K.L.; Fettig, C.J.; Fiedler, C.E.; et al. Ecological effects of alternative fuel-reduction treatments: Highlights of the national Fire and Fire Surrogate study (FFS). Int. J. Wildland Fire 2013, 22, 66–82. [Google Scholar] [CrossRef]
- Stephens, S.L.; McIver, J.D.M.; Boerner, R.E.J.; Fettig, C.J.; Joseph, B.; Hartsough, B.R.; Kennedy, P.L.; Schwilk, D.W. The effects of forest fuel-reduction treatments in the United States. BioScience 2012, 62, 549–560. [Google Scholar] [CrossRef]
- Prichard, S.J.; Povak, N.A.; Kennedy, M.C.; Peterson, D.W. Fuel treatment effectiveness in the context of landform, vegetation, and large, wind-driven wildfires. Ecol. Appl. 2020, 30, e02104. [Google Scholar] [CrossRef] [PubMed]
- Agee, J.K.; Skinner, C.N. Basic principles of forest fuel reduction treatments. For. Ecol. Manag. 2005, 211, 83–96. [Google Scholar] [CrossRef]
- Yocom, L. Fuel Treatment Longevity; Working Pap. No. 27; Ecological Restoration Institute: Flagstaff, AZ, USA, 2013. [Google Scholar]
- Agee, J.K. Fire Ecology of Pacific Northwest Forests; Island Press: Covelo, CA, USA, 1993. [Google Scholar]
- Francis, D.; Ex, S.; Hoffman, C. Stand composition and aspect are related to conifer regeneration densities following hazardous fuels treatments in Colorado, USA. For. Ecol. Manag. 2018, 409, 417–424. [Google Scholar] [CrossRef]
- Korb, J.E.; Fornwalt, P.J.; Stevens-Rumann, C.S. What drives ponderosa pine regeneration following wildfire in the western United States? For. Ecol. Manag. 2019, 454, 117663. [Google Scholar] [CrossRef]
- Stevens-Rumann, C.S.; Morgan, P. Tree regeneration following wildfires in the western US: A review. Fire Ecol. 2019, 15, 15. [Google Scholar] [CrossRef]
- Schwilk, D.W.; Keeley, J.E.; Knapp, E.E.; McIver, J.; Bailey, J.D.; Fettig, C.J.; Fiedler, C.E.; Harrod, R.J.; Moghaddas, J.J.; Outcalt, K.W.; et al. The national Fire and Fire Surrogate study: Effects of fuel reduction methods on forest vegetation structure and fuels. Ecol. Appl. 2009, 19, 285–304. [Google Scholar] [CrossRef] [PubMed]
- Haig, I.T.; Davis, K.P.; Weidman, R.H. Natural Regeneration in the Western White Pine Type; Tech. Bull. 767; USDA/FS, Northern Rocky Mountain Forest & Range Exp. Sta.: Washington, DC, USA, 1941.
- Gray, A.N.; Spies, T.A. Microsite controls on tree seedling establishment in conifer forest canopy gaps. Ecology 1997, 78, 2458–2473. [Google Scholar] [CrossRef]
- Zald, H.S.J.; Gray, A.N.; North, M.; Kern, R.A. Initial tree regeneration responses to fire and thinning treatments in a Sierra Nevada mixed-conifer forest, USA. For. Ecol. Manag. 2008, 256, 168–179. [Google Scholar] [CrossRef]
- Wan, S.; Hui, D.; Luo, Y. Fire effects on nitrogen pools and dynamics in terrestrial ecosystems: A meta-analysis. Ecol. Appl. 2001, 11, 1349–1365. [Google Scholar] [CrossRef]
- Neary, D.G.; Ryan, K.C.; DeBano, L.F. (Eds.) Wildland Fire in Ecosystems: Effects of Fire on Soils and Water; Gen. Tech. Rep. RMRS-GTR-42-vol.4; USDA/FS, Rocky Mountain Res. Sta.: Ogden, UT, USA, 2005; (revised 2008).
- Fowells, H.A.; Stark, N.B. Natural Regeneration in Relation to Environment in the Mixed Conifer Forest Type of California; Res. Pap. PSW-24; USDA/FS, Pacific SW Forest & Range Exp. Sta: Berkeley, CA, USA, 1965.
- Kilgore, B.M. The ecological role of fire in Sierran conifer forests: Its application to National Park management. Quat. Res. 1973, 3, 496–513. [Google Scholar] [CrossRef]
- Krannitz, P.G.; Duralia, T.E. Cone and seed production in Pinus ponderosa: A review. West. N. Am. Nat. 2004, 64, 208–218. [Google Scholar]
- Welch, K.R.; Safford, H.D.; Young, T.P. Predicting conifer establishment post wildfire in mixed conifer forests of the North American Mediterranean-climate zone. Ecosphere 2016, 7, e01609. [Google Scholar] [CrossRef]
- Littlefield, C.E. Topography and post-fire climatic conditions shape spatio-temporal patterns of conifer establishment and growth. Fire Ecol. 2019, 15, 34. [Google Scholar] [CrossRef]
- Tappeiner, J.; Helms, J. Natural regeneration of Douglas fir and white fir on exposed sites in the Sierra Nevada of California. Am. Midland Nat. 1971, 86, 358–370. [Google Scholar]
- North, M.; Hurteau, M.; Fiegener, R.; Barbour, M. Influence of fire and El Niño on tree recruitment varies by species in Sierran mixed conifer. For. Sci. 2005, 51, 187–197. [Google Scholar]
- Oakley, B.B.; North, M.P.; Franklin, J.F. Facilitative and competitive effects of a N-fixing shrub on white fir saplings. For. Ecol. Manag. 2006, 233, 100–107. [Google Scholar] [CrossRef]
- McDonald, P.M. Shelterwood Cutting in a Young-Growth, Mixed Conifer Stand in North Central California; Res. Pap. PSW-117; USDA/FS, Pacific SW Forest & Range Exp. Sta: Berkeley, CA, USA, 1976.
- Burns, R.M.; Honkala, B.H. (Eds.) Silvics of North America: Volume 1. Conifers, Agric. Handb. 654; USDA/FS: Washington, DC, USA, 1990.
- van Mantgem, P.J.; Stephenson, N.L.; Keeley, J.E. Forest reproduction along a climatic gradient in the Sierra Nevada, California. For. Ecol. Manag. 2006, 225, 391–399. [Google Scholar] [CrossRef]
- Povak, N.A.; Churchill, D.J.; Cansler, A.; Hessburg, P.F.; Kane, V.R.; Kane, J.; Lutz, J.A.; Larson, A.J. Wildfire severity and postfire salvage effects on long-term forest regeneration. Ecosphere. in press.
- Conard, S.G.; Radosevich, S.R. Growth responses of white fir to decreased shading and root competition by montane chaparral shrubs. For. Sci. 1982, 28, 309–320. [Google Scholar]
- Helms, J.A.; Tappeiner, J.C. Silviculture in the Sierras, Chapter 15. In Sierra Nevada Ecosystem Project, Final Report to Congress, Assessments and Scientific Basis for Management Options; Centers for Water and Wildland Resources, University of California: Davis, CA, USA, 1996; Volume 2. [Google Scholar]
- Varner, J.M., III; Hiers, J.K.; Ottmar, R.D.; Gordon, D.R.; Putz, F.E.; Wade, D.D. Overstory tree mortality resulting from reintroducing fire to long-unburned longleaf pine forests: The importance of duff moisture. Can. J. For. Res. 2007, 37, 1349–1358. [Google Scholar] [CrossRef]
- Hood, S.M. Mitigating Old Tree Mortality in Long-Unburned, Fire-Dependent Forests: A Synthesis; Gen. Tech. Rep. RMRS-GTR-238; USDA/FS, Rocky Mountain Res. Sta.: Fort Collins, CO, USA, 2010.
- Gill, N.S.; Jarvis, D.; Veblen, T.T.; Pickett, S.T.A.; Kulakowsky, D. Is initial post-disturbance regeneration indicative of longer-term trajectories? Ecosphere 2017, 8, e01924. [Google Scholar] [CrossRef]
- Hessburg, P.F.; Salter, R.B.; Richmond, M.B.; Smith, B.G. Ecological subregions of the interior Columbia basin, USA. Appl. Veg. Sci. 2000, 3, 163–180. [Google Scholar] [CrossRef]
- Agee, J.K.; Lehmkuhl, J.F. Dry Forests of the Northeastern Cascades Fire and Fire Surrogate Project Site, Mission Creek, Okanogan-Wenatchee National Forest; Res. Pap. PNW-RP-577; USDA/FS, Pacific Northwest Res. Sta.: Portland, OR, USA, 2009.
- Lillybridge, T.R.; Kovalchik, B.L.; Williams, C.K.; Smith, B.G. Field Guide for Forested Plant Association of the Wenatchee National Forest; Gen. Tech. Report PNW-GTR-359; USDA/FS, Pacific Northwest Res. Sta.: Portland, OR, USA, 1995.
- Rossman, A.K.; Halpern, C.B.; Harrod, R.J.; Urgenson, L.S.; Peterson, D.W.; Bakker, J.D. Benefits of thinning and burning for understory diversity vary with spatial scale and time since treatment. For. Ecol. Manag. 2018, 419, 58–78. [Google Scholar] [CrossRef]
- Harrod, R.J.; McRae, B.H.; Hartl, W.E. Historical stand reconstruction in ponderosa pine forests to guide silvicultural prescriptions. For. Ecol. Manag. 1999, 114, 433–446. [Google Scholar] [CrossRef]
- Agee, J.K.; Lolley, M.R. Thinning and prescribed fire effects on fuels and potential fire behavior in an eastern Cascades forest, Washington, USA. Fire Ecol. 2006, 2, 3–19. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org (accessed on 12 June 2020).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Portugués, E.G. MSc in Big Data Analytics at Carlos III University of Madrid. Available online: https://bookdown.org/egarpor/PM-UC3M/ (accessed on 15 May 2020).
- Chiono, L.A.; O’Hara, K.L.; De Lasaux, M.J.; Nader, G.A.; Stephens, S.L. Development of vegetation and surface fuels following fire hazard reduction treatment. Forests 2012, 3, 700–722. [Google Scholar] [CrossRef]
- Fialko, K. Conifer Regeneration and Fuels Treatment Longevity in Dry Mixed-Conifer Forests of the Colorado Front Range. Master’s Thesis, Colorado State University, Fort Collins, CO, USA, 2018. [Google Scholar]
- Korb, J.E.; Stoddard, M.T.; Huffman, D.W. Effectiveness of restoration treatments for reducing fuels and increasing understory diversity in shrubby mixed-conifer forests of the Southern Rocky Mountains, USA. Forests 2020, 11, 508. [Google Scholar] [CrossRef]
- Harrod, R.J.; Fonda, R.W.; McGrath, M.K. Vegetation response to thinning and burning in a ponderosa pine forest, Washington. Northwest Sci. 2008, 82, 141–150. [Google Scholar] [CrossRef]
- Ryker, R.A. A Survey of Factors Affecting Regeneration of Rocky Mountain Douglas-Fir; Res. Pap. INT-174; USDA/FS: Ogden, UT, USA, 1975.
- Shearer, R.C. Silviculture. In Clearcutting and Fire in the Larch/Douglas-Fir Forests of Western Montana—A Multifaceted Research Summary; DeByle, N.V., Ed.; Gen. Tech. Rep. INT-99; USDA/FS, Intermountain Forest & Range Exp. Sta.: Ogden, UT, USA, 1981; pp. 27–31. [Google Scholar]
- Bonnet, V.H.; Schoettle, A.W.; Shepperd, W.D. Postfire environmental conditions influence the spatial pattern of regeneration for Pinus ponderosa. Can. J. For. Res. 2005, 35, 37–47. [Google Scholar] [CrossRef]
- Kerns, B.K.; Thies, W.G.; Niwa, C.G. Season and severity of prescribed burn in ponderosa pine forests: Implications for understory native and exotic plants. Ecoscience 2006, 13, 44–55. [Google Scholar] [CrossRef]
- Knapp, E.E.; Keeley, J.E. Heterogeneity in fire severity within early season and late season prescribed burns in a mixed-conifer forest. Int. J. Wildland Fire 2006, 15, 37–45. [Google Scholar] [CrossRef]
- Hatten, J.A.; Zabowski, D.; Ogden, A.; Thies, W. Soil organic matter in a ponderosa pine forest with varying seasons and intervals of prescribed burn. For. Ecol. Manag. 2008, 255, 2555–2565. [Google Scholar] [CrossRef]
- Westlind, D.J.; Kerns, B.K. Long-term effects of burn season and frequency on ponderosa pine forest fuels and seedlings. Fire Ecol. 2017, 13, 42–61. [Google Scholar] [CrossRef]
- Jones, J.R. Mixed Conifer Seedling Growth in Eastern Arizona; Res. Pap. RM-77; USDA/FS, Rocky Mountain Res. Sta.: Fort Collins, CO, USA, 1971.
- Strothman, R.O. Douglas-Fir Seedlings Plants by Four Methods…Results after 10 Years; Res. Note PSW-310; USDA/FS, Pacific SW Forest & Range Exp. Sta.: Berkeley, CA, USA, 1976.
- Hermann, R.K.; Lavender, D.P. Pseudotsuga menziesii. In Silvics of North America: Volume 1. Conifers; Burns, R.M., Honkala, B.H., Eds.; Agric. Handb. 654; USDA/FS: Washington, DC, USA, 1990. [Google Scholar]
- Schneider, W.G.; Knowe, S.A.; Harrington, T.B. Predicting survival of planted Douglas-fir and ponderosa pine seedlings on dry, low-elevation sites in southwestern Oregon. New For. 1998, 15, 139–159. [Google Scholar] [CrossRef]
- Rose, R.; Ketchum, J.S.; Hanson, D.E. Three-year survival and growth of Douglas-fir seedlings under various vegetation-free regimes. For. Sci. 1999, 45, 117–126. [Google Scholar]
- Krauch, H. Management of Douglas-Fir Timberland in the Southwest; Pap. 21; USDA/FS, Rocky Mountain Forest & Range Exp. Sta.: Fort Collins, CO, USA, 1956.
- Ryker, R.A.; Potter, D.R. Shade Increases First-Year Survival of Douglas-Fir Seedlings; Res. Pap. INT-119; USDA/FS: Ogden, UT, USA, 1970.
- Steinberg, P.D. Pseudotsuga menziesii var. glauca. In Fire Effects Information System; [Online]; USDA/FS, Rocky Mountain Res. Sta. Fire Sci. Lab., 2002. Available online: https://www.fs.fed.us/database/feis/plants/tree/psemeng/all.html (accessed on 10 May 2020).
- Keifer, M.B.; van Wagtendonk, J.W.; Buhler, M. Long-term surface fuel accumulation in burned and unburned mixed-conifer forests of the central and southern Sierra Nevada, CA, USA. Fire Ecol. 2006, 2, 53–72. [Google Scholar] [CrossRef]
- North, M.; Innes, J.; Zald, H. Comparison of thinning and prescribed fire restoration treatments to Sierran mixed-conifer historic conditions. Can. J. For. Res. 2007, 37, 331–342. [Google Scholar] [CrossRef]
- Rother, M.T.; Veblen, T.T.; Furman, L.G. A field experiment informs expected patterns of conifer regeneration after disturbance under changing climate conditions. Can. J. For. Res. 2015, 45, 1607–1616. [Google Scholar] [CrossRef]
- Rother, M.T.; Veblen, T.T. Limited conifer regeneration following wildfires in dry ponderosa pine forests of the Colorado Front Range. Ecosphere 2016, 7, e01594. [Google Scholar] [CrossRef]
- Warren, R.J. A test of temperature estimation from solar irradiation and a simple statistical method to integrate elevation into prediction models. Castanea 2010, 75, 67–77. [Google Scholar] [CrossRef]









| Category | Response | Definition | Sampling Date or Interval | Analysis Units |
|---|---|---|---|---|
| Overstory dynamics | Ingrowth | Recruitment into the overstory: trees reaching 7.6 cm DBH during interval | Early–Late | number/ha |
| Mortality | Trees that died during interval | Early–Late | number/ha | |
| Basal area increment (BAI) | Change in basal area during interval | Early–Late | m2/ha | |
| Seedling frequency (<1 m tall) | Seedlings | Conifers <1 m tall | Pre, Early, Late | proportion of quadrats |
| Seedling dynamics | Colonization | Quadrats with seedlings (<1 m tall) at the end but not start of interval | Pre–Early, Early–Late, Pre–Late | number of quadrats 1 |
| Persistence | Quadrats with seedlings (<1 m tall) at the start and end of interval | Pre–Early, Early–Late, Pre–Late | number of quadrats 1 | |
| Regeneration frequency (by height class) | Small seedlings | Conifers <0.3 m tall | Late | proportion of quadrats |
| Large seedlings | Conifers 0.3–1.37 m tall | Late | proportion of grid cells | |
| Saplings | Conifers >1.37 m tall and <7.6 cm DBH | Late | proportion of grid cells |
| Percentage of Explained Variability | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Category | Response | Sampling Date or Interval | R2 | OA | Thin | Early Fire (EF) | Late Fire (LF) | OA × Thin | OA × EF | OA × LF | Thin × EF | Thin × LF | EF × LF |
| Overstory dynamics | Ingrowth | Early–Late | 0.27 | 1 | 28 | 10 | 17 | 9 | 6 | 1 | 20 | 6 | 2 |
| Mortality | Early–Late | 0.64 | 57 | 0 | 2 | 25 | 1 | 7 | 2 | 1 | 2 | 2 | |
| BAI | Early–Late | 0.36 | 17 | 3 | 12 | 26 | 9 | 25 | 1 | 4 | 1 | 1 | |
| Seedling frequency (<1 m tall) | Seedlings | Pre | 0.07 | 100 | – | – | – | – | – | – | – | – | – |
| Early | 0.24 | 5 | 6 | 18 | – | 63 | 5 | – | 4 | – | – | ||
| Late | 0.38 | 1 | 7 | 13 | 55 | 5 | <1 | 6 | 1 | 1 | 10 | ||
| Seedling dynamics | Colonization | Pre–Early | 0.30 | 31 | 2 | 21 | – | 36 | 7 | – | 4 | – | – |
| Early–Late | 0.47 | 10 | <1 | 18 | 47 | 4 | 1 | 6 | 1 | 2 | 11 | ||
| Pre–Late | 0.49 | 4 | 7 | 18 | 45 | 4 | 2 | <1 | 1 | 6 | 12 | ||
| Persistence | Pre–Early | na | – | – | – | - | - | - | - | ||||
| Early–Late | na | – | – | – | – | – | – | ||||||
| Pre–Late | na | – | – | – | – | – | – | ||||||
| Regeneration frequency (by height class) | Small seedlings | Late | 0.41 | 1 | 8 | 15 | 63 | 2 | <1 | 4 | <1 | <1 | 7 |
| Large seedlings | Late | 0.39 | 5 | 18 | 5 | 18 | 38 | 1 | 13 | <1 | 1 | 2 | |
| Saplings | Late | 0.43 | 35 | 17 | <1 | 21 | 18 | 4 | <1 | <1 | 4 | 2 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossman, A.K.; Bakker, J.D.; Peterson, D.W.; Halpern, C.B. Long-Term Effects of Fuels Treatments, Overstory Structure, and Wildfire on Tree Regeneration in Dry Forests of Central Washington. Forests 2020, 11, 888. https://doi.org/10.3390/f11080888
Rossman AK, Bakker JD, Peterson DW, Halpern CB. Long-Term Effects of Fuels Treatments, Overstory Structure, and Wildfire on Tree Regeneration in Dry Forests of Central Washington. Forests. 2020; 11(8):888. https://doi.org/10.3390/f11080888
Chicago/Turabian StyleRossman, Allison K., Jonathan D. Bakker, David W. Peterson, and Charles B. Halpern. 2020. "Long-Term Effects of Fuels Treatments, Overstory Structure, and Wildfire on Tree Regeneration in Dry Forests of Central Washington" Forests 11, no. 8: 888. https://doi.org/10.3390/f11080888
APA StyleRossman, A. K., Bakker, J. D., Peterson, D. W., & Halpern, C. B. (2020). Long-Term Effects of Fuels Treatments, Overstory Structure, and Wildfire on Tree Regeneration in Dry Forests of Central Washington. Forests, 11(8), 888. https://doi.org/10.3390/f11080888