Long-Term Seeding Outcomes in Slash Piles and Skid Trails after Conifer Removal


Abstract
1. Introduction
2. Materials and Methods
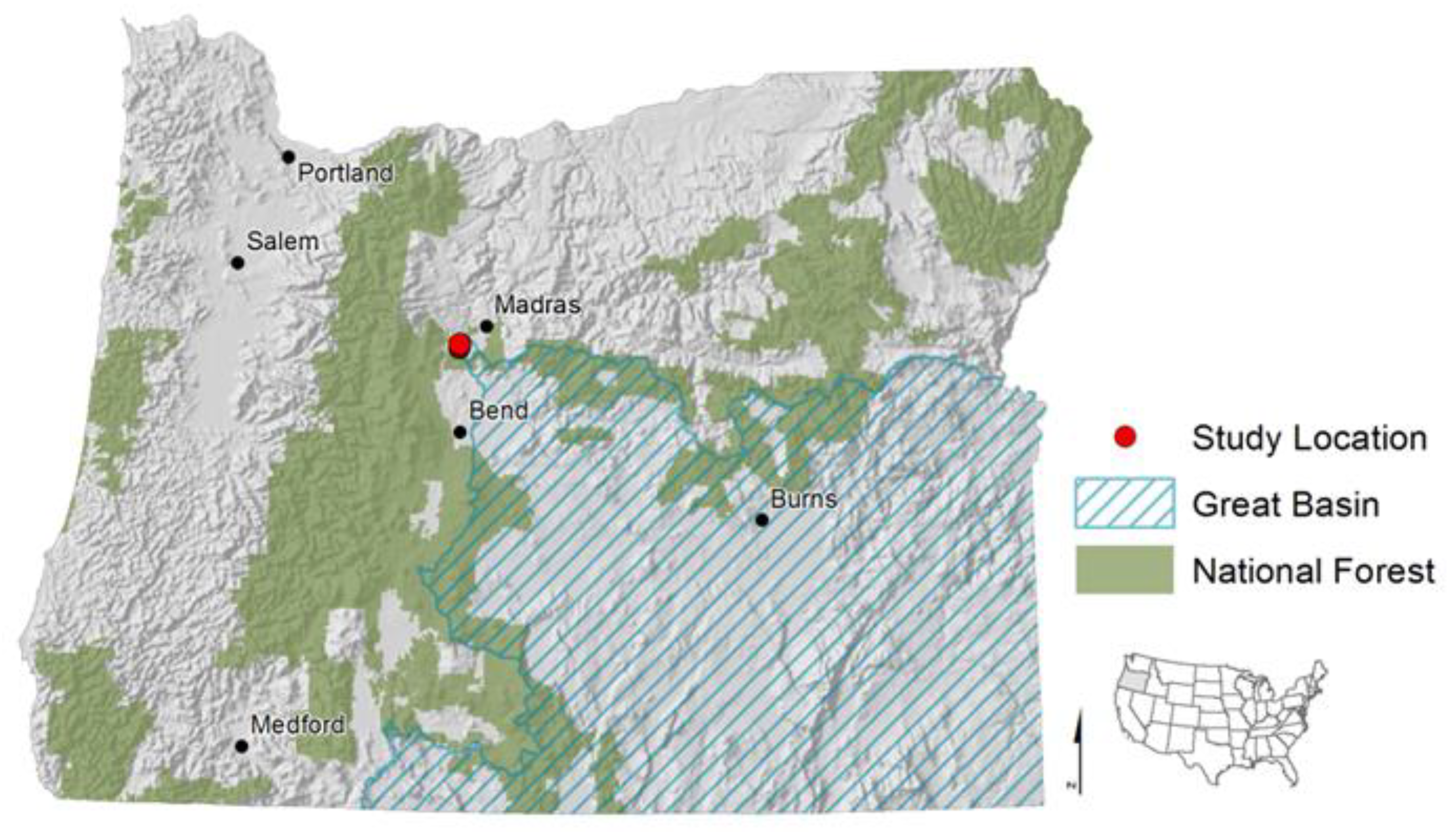
2.1. Study Area and Seeding Treatments
2.2. Sampling
2.3. Data Analysis
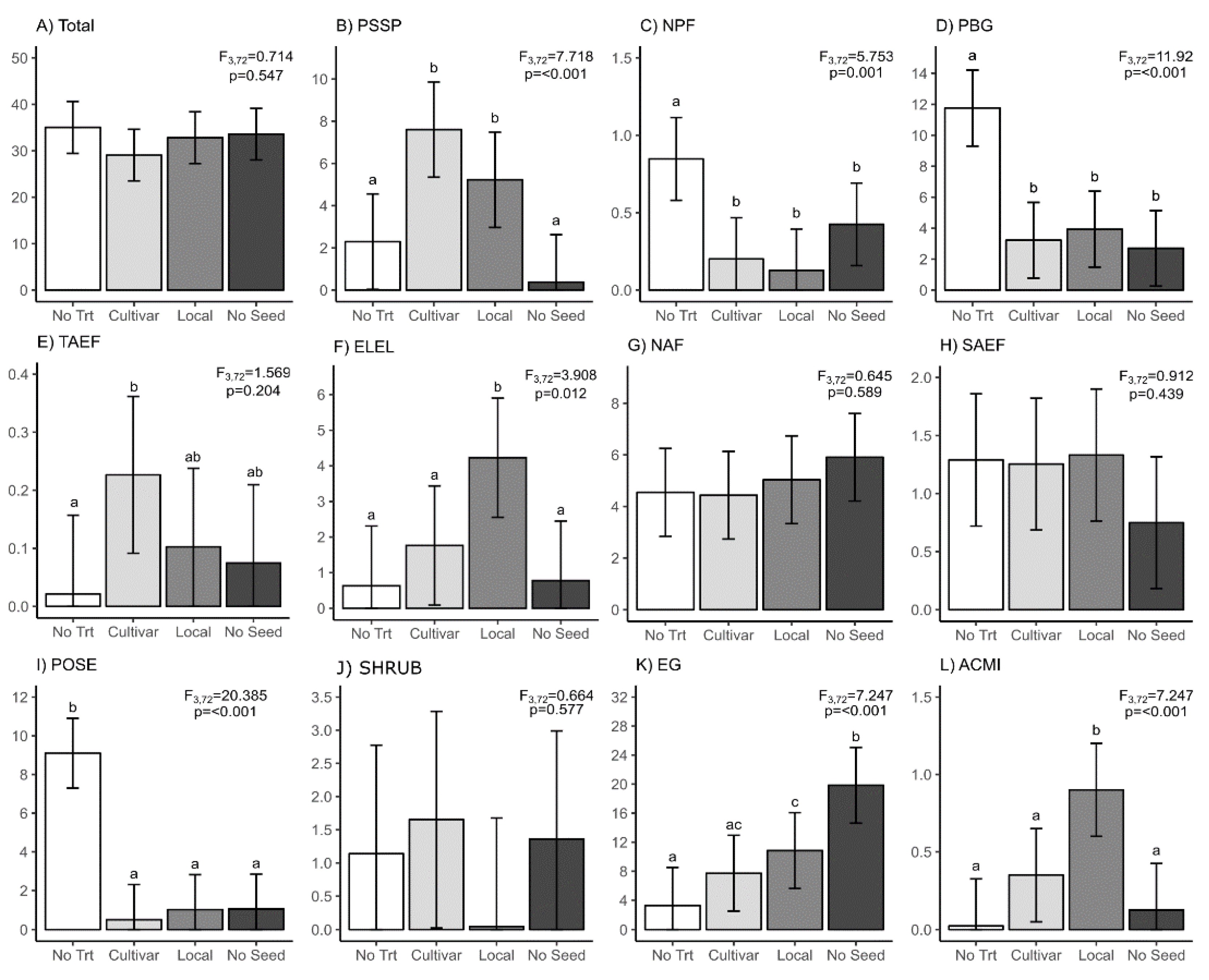
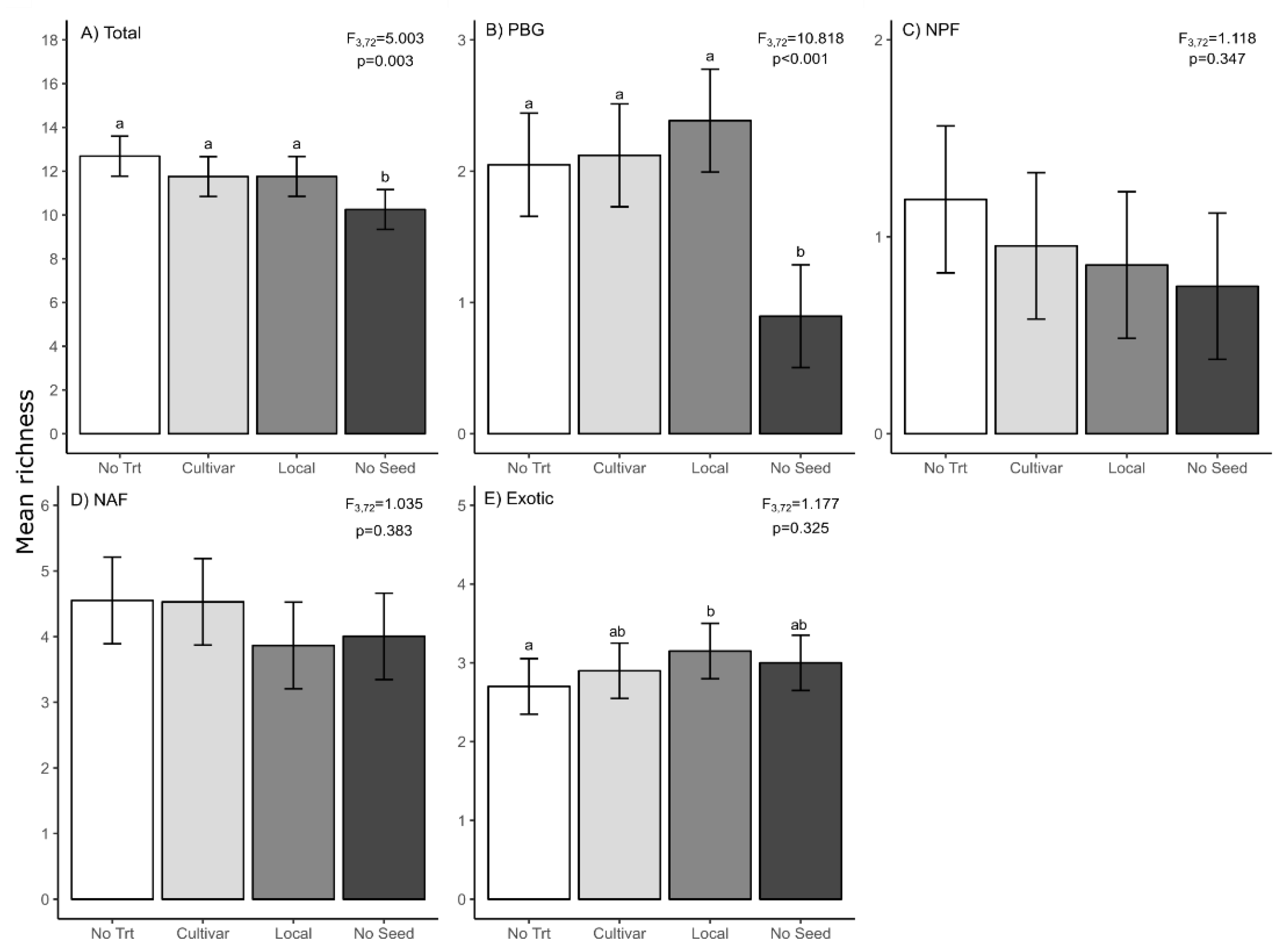
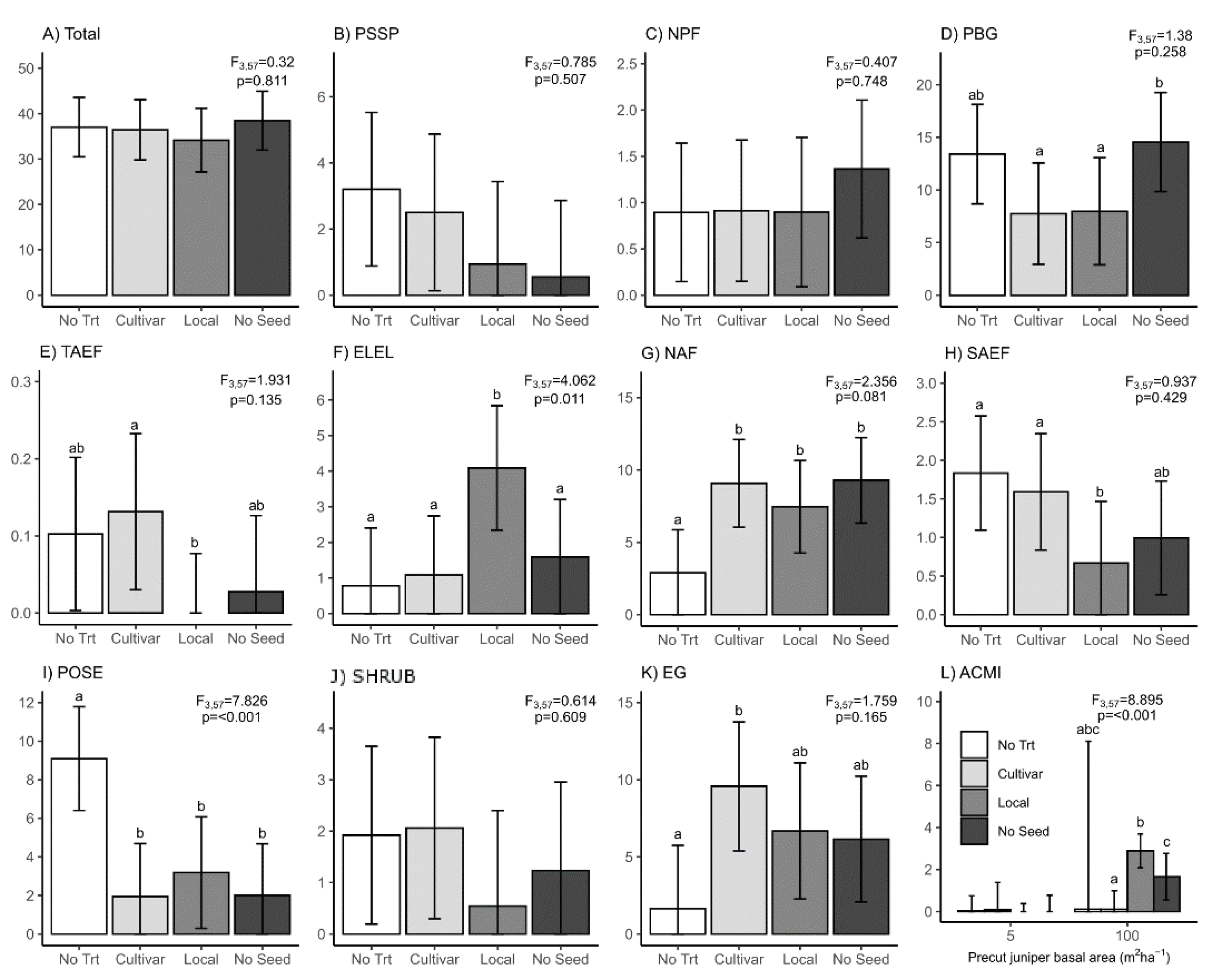
3. Results
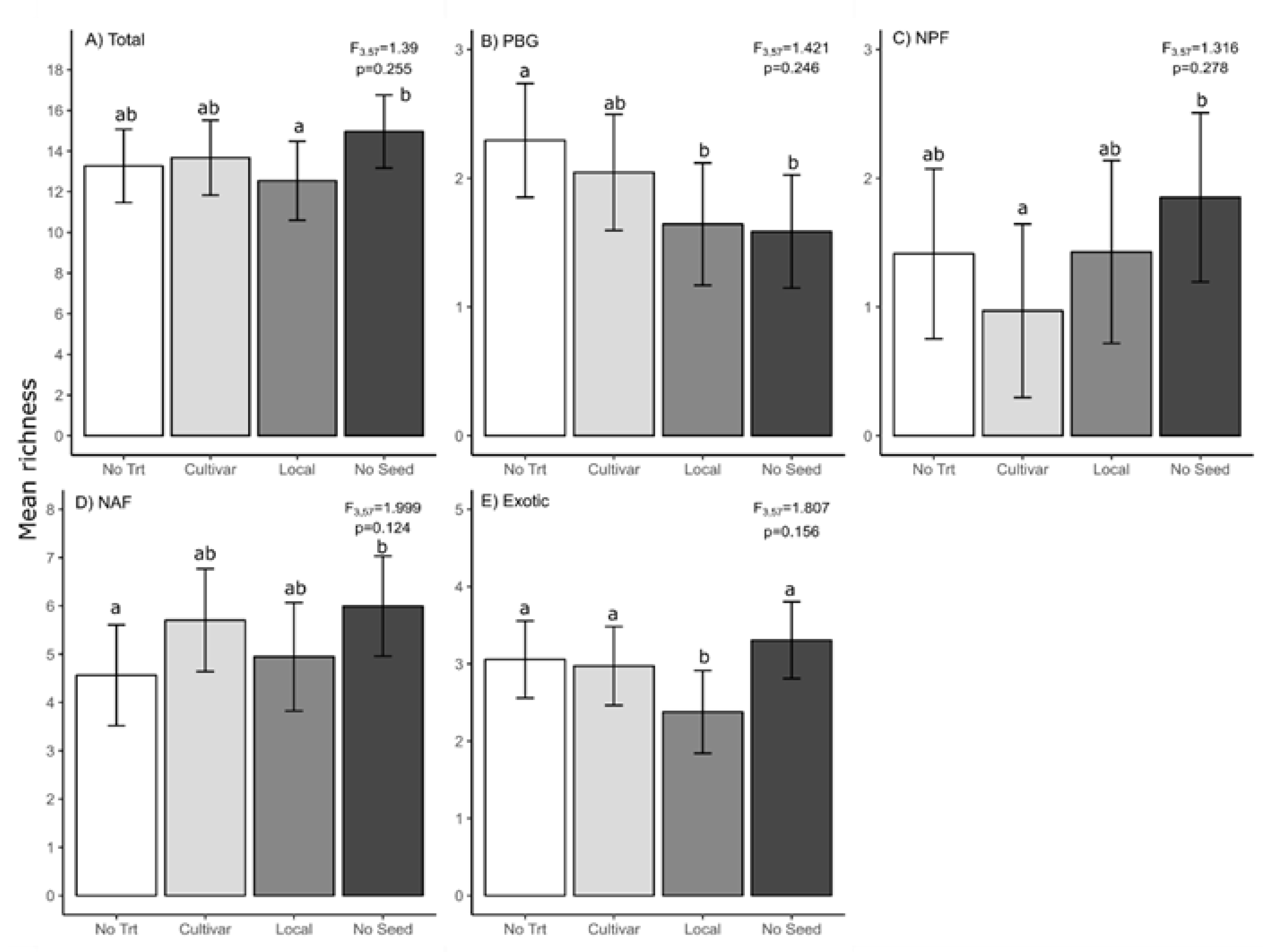
3.1. Slash Piles
3.2. Skid Trails
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Halpern, C.B.; Lutz, J.A. Canopy closure exerts weak controls on understory dynamics: A 30-year study of overstory–understory interactions. Ecol. Monogr. 2013, 83, 221–237. [Google Scholar] [CrossRef]
- Staver, A.C.; Archibald, S.; Levin, S. Tree cover in sub-Saharan Africa: Rainfall and fire constrain forest and savanna as alternative stable states. Ecology 2011, 92, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Lubetkin, K.C.; Westerling, A.L.; Kueppers, L.M. Climate and landscape drive the pace and pattern of conifer encroachment into subalpine meadows. Ecol. Appl. 2017, 27, 1876–1887. [Google Scholar] [CrossRef] [PubMed]
- Schriver, M.; Sherriff, R.L.; Varner, J.M.; Quinn-Davidson, L.; Valachovic, Y. Age and stand structure of oak woodlands along a gradient of conifer encroachment in northwestern California. Ecosphere 2018, 9, e02446. [Google Scholar] [CrossRef]
- Hagmann, R.K.; Franklin, J.F.; Johnson, K.N. Historical structure and composition of ponderosa pine and mixed-conifer forests in south-central Oregon. For. Ecol. Manag. 2013, 304, 492–504. [Google Scholar] [CrossRef]
- Miller, R.F.; Rose, J.A. Historic expansion of Juniperus occidentalis (western juniper) in southeastern Oregon. Great Basin Nat. 1995, 55, 37–45. [Google Scholar]
- Tausch, R.J.; West, N.E.; Nabi, A. Tree age and dominance patterns in Great Basin pinyon-juniper woodlands. Rangel. Ecol. Manag. J. Range Manag. Arch. 1981, 34, 259–264. [Google Scholar] [CrossRef]
- Romme, W.H.; Allen, C.D.; Balley, J.D.; Baker, W.L.; Bestelmeyer, B.T.; Brown, P.M.; Eisenhart, K.S.; Floyd, M.L.; Huffman, D.W.; Jacobs, B.F.; et al. Historical and modern disturbance regimes, stand structures, and landscape dynamics in pinon-puniper vegetation of the western United States. Rangel. Ecol. Manag. 2009, 62, 203–222. [Google Scholar] [CrossRef]
- Burkhardt, J.W.; Tisdale, E. Nature and successional status of western juniper vegetation in Idaho. J. Range Manag. 1969, 22, 264–270. [Google Scholar] [CrossRef]
- Eddleman, L.E. Establishment and stand development of western juniper in central Oregon. In Proceedings of the Pinyon-Juniper Conference; INT-GTR-215; USDA Forest Service, Intermountain Research Station: Ogden, UT, USA, 1986; pp. 255–259. [Google Scholar]
- Miller, R.F.; Rose, J.A. Fire history and western juniper encroachment in sagebrush steppe. J. Range Manag. 1999, 52, 550–559. [Google Scholar] [CrossRef]
- Miller, R.F.; Angell, R.F.; Eddleman, L.E. Water use by western juniper. In Proceedings of the Pinyon-Juniper Conference, Sun Vally, Reno, NV, USA, 13–16 January 1986; pp. 418–422. [Google Scholar]
- Knapp, P.A.; Soulé, P.T. Recent Juniperus occidentalis (western juniper) expansion on a protected site in central Oregon. Glob. Chang. Biol. 1998, 4, 347–357. [Google Scholar] [CrossRef]
- Ko, D.W.; Sparrow, A.D.; Weisberg, P.J. Land-use legacy of historical tree harvesting for charcoal production in a semi-arid woodland. For. Ecol. Manag. 2011, 261, 1283–1292. [Google Scholar] [CrossRef]
- Suring, L.H.; Rowland, M.M.; Wisdom, M.J. Identifying species of conservation concern. In Habitat Threats in the Sagebrush Ecosystem: Methods of Regional Assessment and Applications in the Great Basin; Wisdom, M.J., Rowland, M.M., Suring, L.H., Eds.; Alliance Communications Group: Lawrence, KS, USA, 2005; pp. 150–162. [Google Scholar]
- Wisdom, M.J.; Rowland, M.M.; Suring, L.H.; Schueck, L.; Meinke, C.; Knick, S. Evaluating species of conservation concern at regional scales. In Habitat Threats in the Sagebrush Ecosystem: Methods of Regional Assessment and Applications in the Great Basin; Wisdom, M.J., Rowland, M.M., Suring, L.H., Eds.; Alliance Communications Group: Lawrence, KS, USA, 2005; pp. 5–24. [Google Scholar]
- Miller, R.F.; Bates, J.D.; Svejcar, T.J.; Pierson, F.B.; Eddleman, L.E. Biology, Ecology, and Management of Western Juniper (Juniperus Occidentalis) in Southeastern Oregon; 152; Oregon State University, Agricultural Experiment Station: Corvallis, OR, USA, 2005. [Google Scholar]
- Kormos, P.R.; Marks, D.; Pierson, F.B.; Williams, C.J.; Hardegree, S.P.; Havens, S.; Hedrick, A.; Bates, J.D.; Svejcar, T.J. Ecosystem water availability in juniper versus sagebrush snow-dominated rangelands. Rangel. Ecol. Manag. 2017, 70, 116–128. [Google Scholar] [CrossRef]
- Miller, R.F.; Chambers, J.C.; Evers, L.; Williams, C.J.; Snyder, K.A.; Roundy, B.A.; Pierson, F.B. The Ecology, History, Ecohydrology, and Management of Pinyon and Juniper Woodlands in the Great Basin and Northern Colorado Plateau of the Western United States; Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2019; p. 284.
- Wagenbrenner, J.W.; MacDonald, L.H.; Coats, R.N.; Robichaud, P.R.; Brown, R.E. Effects of post-fire salvage logging and a skid trail treatment on ground cover, soils, and sediment production in the interior western United States. For. Ecol. Manag. 2015, 335, 176–193. [Google Scholar] [CrossRef]
- Rhoades, C.C.; Fornwalt, P.J. Pile burning creates a fifty-year legacy of openings in regenerating lodgepole pine forests in Colorado. For. Ecol. Manag. 2015, 336, 203–209. [Google Scholar] [CrossRef]
- Rhoades, C.C.; Meier, A.; Rebertus, A. Soil properties in fire-consumed log burnout openings in a Missouri oak savanna. For. Ecol. Manag. 2004, 192, 277–284. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Esquilín, A.E.J.; Stromberger, M.E.; Massman, W.J.; Frank, J.M.; Shepperd, W.D. Microbial community structure and activity in a Colorado Rocky Mountain forest soil scarred by slash pile burning. Soil Biol. Biochem. 2007, 39, 1111–1120. [Google Scholar] [CrossRef]
- Busse, M.D.; Shestak, C.J.; Hubbert, K.R. Soil heating during burning of forest slash piles and wood piles. Int. J. Wildland Fire 2013, 22, 786–796. [Google Scholar] [CrossRef]
- Halpern, C.B.; Antos, J.A.; Beckman, L.M. Vegetation recovery in slash-pile scars following conifer removal in a grassland-restoration experiment. Restor. Ecol. 2014, 22, 731–740. [Google Scholar] [CrossRef]
- DeSandoli, L.; Turkington, R.; Fraser, L. Restoration of slash pile burn scars to prevent establishment and propagation of non-native plants. Can. J. For. Res. 2016, 46, 1042–1050. [Google Scholar] [CrossRef]
- Korb, J.E.; Johnson, N.C.; Covington, W.W. Slash pile burning effects on soil biotic and chemical properties and plant establishment: Recommendations for amelioration. Restor. Ecol. 2004, 12, 52–62. [Google Scholar] [CrossRef]
- Hebel, C.L.; Smith, J.E.; Cromack, K.J., Jr. Invasive plant species and soil microbial response to wildfire burn severity in Cascade Range of Oregon. Appl. Soil. Ecol. 2009, 42, 150–159. [Google Scholar] [CrossRef]
- Kerns, B.; Day, M. Fuel reduction, seeding, and vegetation in a juniper woodland. Rangel. Ecol. Manag. 2014, 67, 667–679. [Google Scholar] [CrossRef]
- Robichaud, P.R.; Beyers, J.L.; Neary, D.G. Evaluating The Effectiveness Of Postfire Rehabilitation Treatments; RMRS-GTR-63; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2000; p. 85.
- Beyers, J.L. Postfire seeding for erosion control: Effectiveness and impacts on native plant communities. Conserv. Biol. 2004, 18, 947–956. [Google Scholar] [CrossRef]
- Urza, A.K.; Weisberg, P.J.; Chambers, J.C.; Board, D.; Flake, S.W. Seeding native species increases resistance to annual grass invasion following prescribed burning of semiarid woodlands. Biol. Invasions 2019, 21, 1993–2007. [Google Scholar] [CrossRef]
- DiTomaso, J.M. Invasive weeds in rangelands: Species, impacts, and management. Weed Sci. 2000, 48, 255–265. [Google Scholar] [CrossRef]
- Davies, K.W. Medusahead dispersal and establishment in sagebrush steppe plant communities. Rangel. Ecol. Manag. 2008, 61, 110–115. [Google Scholar] [CrossRef]
- Davies, K.W. Revegetation of medusahead-invaded sagebrush steppe. Rangel. Ecol. Manag. 2010, 63, 564–571. [Google Scholar] [CrossRef]
- Abella, S.R.; Craig, D.J.; Smith, S.D.; Newton, A.C. Identifying native vegetation for reducing exotic species during the restoration of desert ecosystems. Restor. Ecol. 2012, 20, 781–787. [Google Scholar] [CrossRef]
- Herron, C.M.; Jonas, J.L.; Meiman, P.J.; Paschke, M.W. Using native annual plants to restore post-fire habitats in western North America. Int J. Wildland Fire 2013, 22, 815–821. [Google Scholar] [CrossRef]
- Bischoff, A.; Steinger, T.; Müller-Schärer, H. The importance of plant provenance and genotypic diversity of seed material used for ecological restoration. Restor. Ecol. 2010, 18, 338–348. [Google Scholar] [CrossRef]
- Smith, S.L.; Sher, A.A.; Grant III, T.A. Genetic diversity in restoration materials and the impacts of seed collection in Colorado’s restoration plant production industry. Restor. Ecol. 2007, 15, 369–374. [Google Scholar] [CrossRef]
- Gustafson, D.; Gibson, D.; Nickrent, D. Competitive relationships of Andropogon gerardii (Big Bluestem) from remnant and restored native populations and select cultivated varieties. Funct. Ecol. 2004, 18, 451–457. [Google Scholar] [CrossRef]
- Bates, J.D.; Miller, R.F.; Svejcar, T. Long-term successional trends following western juniper cutting. Rangel. Ecol. Manag. 2005, 58, 533–541. [Google Scholar] [CrossRef]
- USDA Forest Service. Crooked River National Grassland Vegetation Management/Grazing Final Environmental Impact Statement; USDA Forest Service, Ochoco National Forest, Crooked River National Grassland: Madras, OR, USA, 2004.
- ESRI. ArcMap; 9.2; Environmental Systems Research Institute, Inc.: Redlands, CA, USA, 2006. [Google Scholar]
- Aleksoff, K.C. Achillea millefolium. In Fire Effects Information System; USDA Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory: Missoula, MT, USA, 1999. [Google Scholar]
- Sheley, R.L.; Mangold, J.; Goodwin, K.; Marks, J. Revegetation Guidelines for the Great Basin: Considering Invasive Weeds; ARS-168; USDA-ARS: Washington, DC, USA, 2008; p. 60.
- Davies, K.; Dean, A. Prescribed summer fire and seeding applied to restore juniper-encroached and exotic annual grass-invaded sagebrush steppe. Rangel. Ecol. Manag. 2019, 72, 635–639. [Google Scholar] [CrossRef]
- Meyers, S.C.; Jaster, T.; Mitchell, K.E.; Hardison, L.K. (Eds.) Flora of Oregon. Volume 1: Pteridophytes, Gymnosperms, and Monocots; Botanical Research Institute of Texas Press: Fort Worth, TX, USA, 2015; p. 591. [Google Scholar]
- Hitchcock, C.L.; Cronquist, A. Flora of the Pacific Northwest; University of Washington Press: Seattle, WA, USA, 1973. [Google Scholar]
- R Development Core Team. R. A Language and Environment for Statistical Computing; 3.5.2; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. Nlme: Linear and Nonlinear Mixed Effects Models. 3.1.111; R Package. 2015. Available online: https://cran.r-project.org/web/packages/nlme/index.html (accessed on 8 June 2020).
- Lenth, R.; Singmann, H.; Love, J. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. 1.3.1; R Package. 2018. Available online: https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 8 June 2020).
- Rinella, M.J.; Mangold, J.M.; Espeland, E.K.; Sheley, R.L.; Jacobs, J.S. Long-term population dynamics of seeded plants in invaded grasslands. Ecol. Appl. 2012, 22, 1320–1329. [Google Scholar] [CrossRef]
- Knutson, K.C.; Pyke, D.A.; Wirth, T.A.; Arkle, R.S.; Pilliod, D.S.; Brooks, M.L.; Chambers, J.C.; Grace, J.B. Long-term effects of seeding after wildfire on vegetation in Great Basin shrubland ecosystems. J. Appl. Ecol. 2014, 51, 1414–1424. [Google Scholar] [CrossRef]
- Ott, J.E.; Kilkenny, F.F.; Summers, D.D.; Thompson, T.W. Long-term vegetation recovery and invasive annual suppression in native and introduced postfire seeding treatments. Rangel. Ecol. Manag. 2019, 72, 640–653. [Google Scholar] [CrossRef]
- Creech, M.N.; Katherine Kirkman, L.; Morris, L.A. Alteration and recovery of slash pile burn sites in the restoration of a fire-maintained ecosystem. Restor. Ecol. 2012, 20, 505–516. [Google Scholar] [CrossRef]
- Copeland, S.M.; Munson, S.M.; Bradford, J.B.; Butterfield, B.J.; Gunnell, K.L. Long-term plant community trajectories suggest divergent responses of native and non-native perennials and annuals to vegetation removal and seeding treatments. Restor. Ecol. 2019, 27, 821–831. [Google Scholar] [CrossRef]
- Bates, J.D.; Miller, R.F.; Svejcar, T.J. Understory dynamics in cut and uncut western juniper woodlands. J. Range Manag. 2000, 53, 119–126. [Google Scholar] [CrossRef]
- Davies, K.W.; Johnson, D.D. Established perennial vegetation provides high resistance to reinvasion by exotic annual grasses. Rangel. Ecol. Manag. 2017, 70, 748–754. [Google Scholar] [CrossRef]
- Aubry, C.; Shoal, R.; Erickson, V. Grass Cultivars: Their Origins, Development, and Use on National Forests and Grasslands in the Pacific Northwest; USDA Forest Service: Washington, DC, USA, 2005.
- Kerns, B.K.; Tortorelli, C.; Day, M.A.; Nietupski, T.; Barros, A.M.G.; Kim, J.B.; Krawchuk, M.A. Invasive grasses: A new perfect storm for forested ecosystems? For. Ecol. Manag. 2020, 463. [Google Scholar] [CrossRef]
- Bradley, B.A.; Curtis, C.A.; Fusco, E.J.; Abatzoglou, J.T.; Balch, J.K.; Dadashi, S.; Tuanmu, M.-N. Cheatgrass (Bromus tectorum) distribution in the intermountain Western United States and its relationship to fire frequency, seasonality, and ignitions. Biol. Invasions 2018, 20, 1493–1506. [Google Scholar] [CrossRef]
- Knapp, P.A. Cheatgrass (Bromus tectorum L.) dominance in the Great Basin Desert-history, persistence, and influences to human activities. Glob. Environ. Chang. Hum. Policy Dimens. 1996, 6, 37–52. [Google Scholar] [CrossRef]
- Davies, G.; Bakker, J.; Dettweiler-Robinson, E.; Dunwiddie, P.W.; Hall, S.; Downs, J.; Evans, J. Trajectories of change in sagebrush steppe vegetation communities in relation to multiple wildfires. Ecol. Appl. 2012, 22, 1562–1577. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Activity | Date |
|---|---|
| Control units established | April 2008 |
| Slash-pile plot locations established | April 2008 |
| Slash-pile pretreatment data collected | Spring/Summer 2008 |
| Juniper thinned | Spring–Fall 2008 |
| Slash piled by fire crew | Fall 2008 |
| Trees removed from site | Fall 2008–Summer 2009 |
| Skid trail plots established (no pretreat data) | May 2009 |
| Piles burned | December 2009 |
| Piles seeded | December 2009 |
| Skid trails raked and seeded | December 2009 |
| Data collected (published by Kerns and Day, 2014) | Summer 2011 |
| Data collected | Summer 2017 |
| Functional Group | Code | Dominant Species |
|---|---|---|
| Total plant cover | Total | Sandberg bluegrass (Poa secunda), Thurber’s needlegrass (Achnatherum thurberianum), Idaho fescue (Festuca idahoensis Elmer) |
| Perennial large bunchgrasses | PBG | No seeded species, Thurber’s needlegrass, Idaho fescue |
| Shallow, early season grass | POSE | Sandberg bluegrass |
| Perennial Forbs | NPF | Low pussytoes (Antennaria dimorpha (Nutt.) Torr. & A. Gray), velvet lupine (Lupinus leucophyllus Douglas ex Lindl.) |
| Native annuals | NAF | Pacific popcornflower (Plagiobothrys tenellus (Nutt. ex Hook.), small fescue (Vulpia microstachys (Nutt.) Munro) |
| Exotic grasses | EG | Cheatgrass (Bromus tectorum) |
| Tall annual exotic forbs | TAEF | Tall tumblemustard (Sisymbrium altissimum L.), littlepod false flax (Camelina microcarpa Andrz. ex DC.) |
| Small annual exotic forbs | SAEF | Jagged chickweed (Holosteum umbellatum L.), spring draba (Draba verna L.) |
| Shrubs | SHRUB | Antelope bitterbrush (Purshia tridentate (Pursh) DC), Wyoming sagebrush (Artemisia tridentata ssp. wyomingensis), green rabbit brush (Chrysothamnus viscidiflorus(Hook.) Nutt.) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerns, B.K.; Day, M.A.; Ikeda, D. Long-Term Seeding Outcomes in Slash Piles and Skid Trails after Conifer Removal. Forests 2020, 11, 839. https://doi.org/10.3390/f11080839
Kerns BK, Day MA, Ikeda D. Long-Term Seeding Outcomes in Slash Piles and Skid Trails after Conifer Removal. Forests. 2020; 11(8):839. https://doi.org/10.3390/f11080839
Chicago/Turabian StyleKerns, Becky K., Michelle A. Day, and Dana Ikeda. 2020. "Long-Term Seeding Outcomes in Slash Piles and Skid Trails after Conifer Removal" Forests 11, no. 8: 839. https://doi.org/10.3390/f11080839
APA StyleKerns, B. K., Day, M. A., & Ikeda, D. (2020). Long-Term Seeding Outcomes in Slash Piles and Skid Trails after Conifer Removal. Forests, 11(8), 839. https://doi.org/10.3390/f11080839