Mediterranean Heathland as a Key Habitat for Fire Adaptations: Evidence from an Experimental Approach

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Species
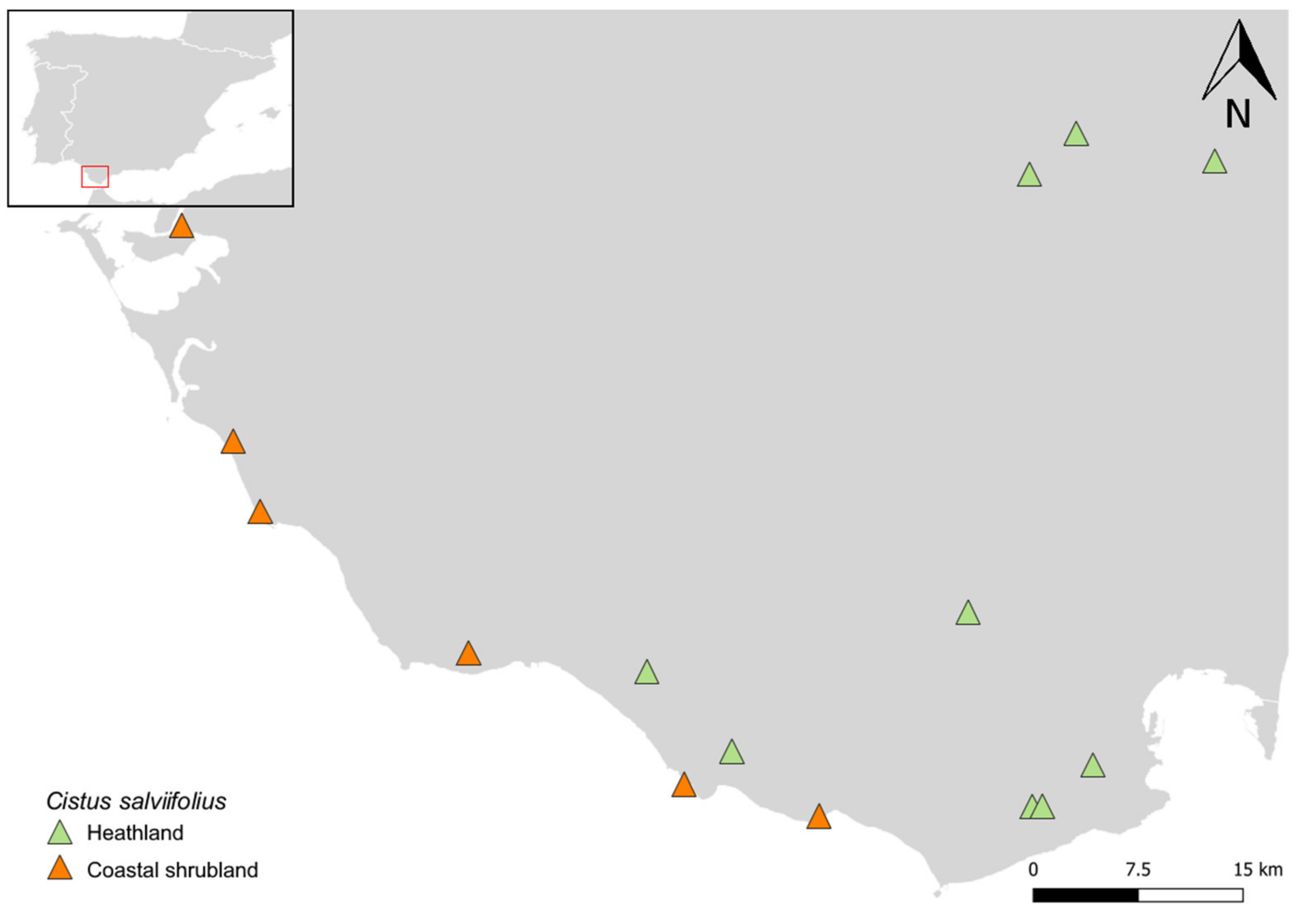
2.2. Study Area, Populations and Seed Collection
2.3. Seed Size Measurement and Germination Experiment
2.4. Statistical Analyses
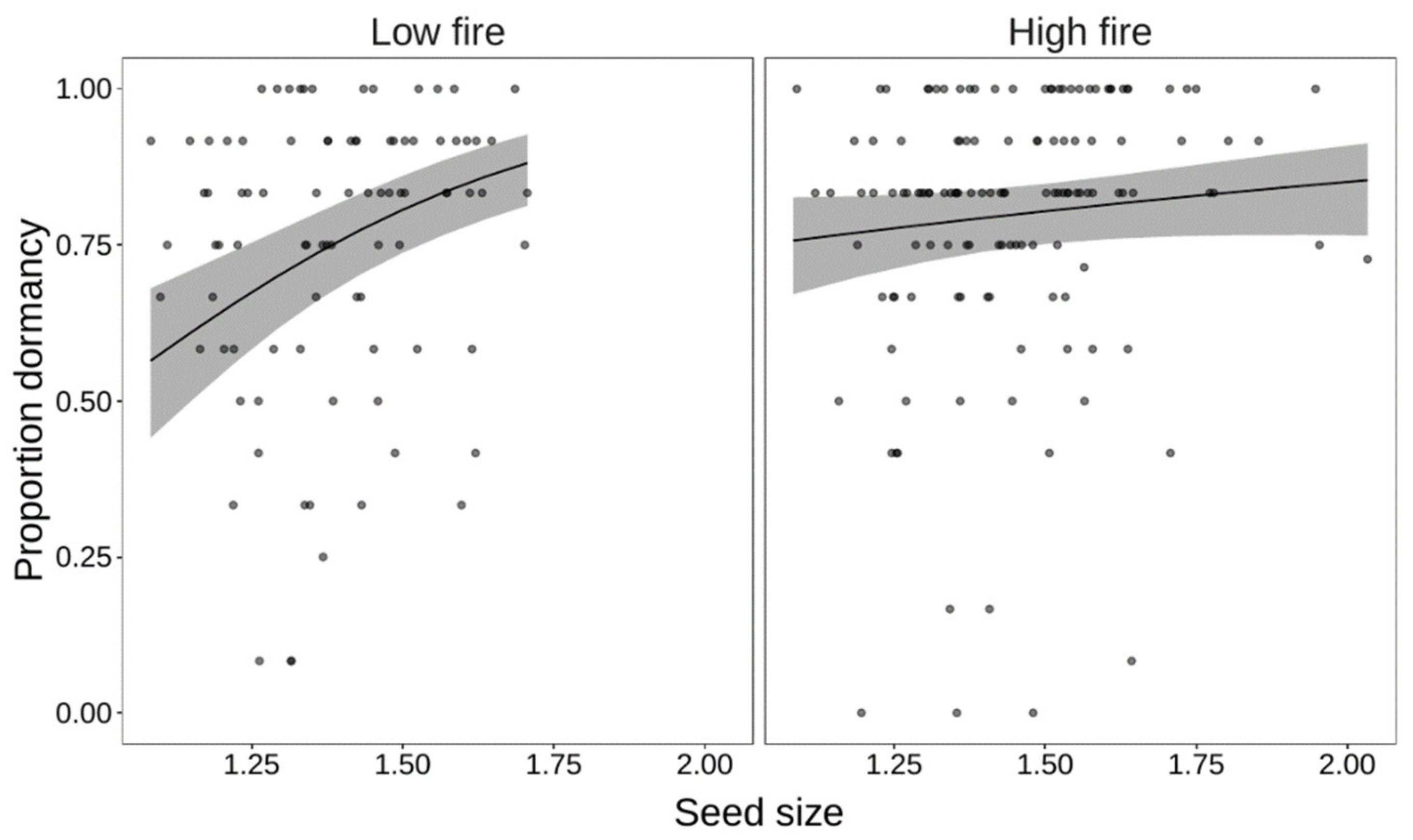
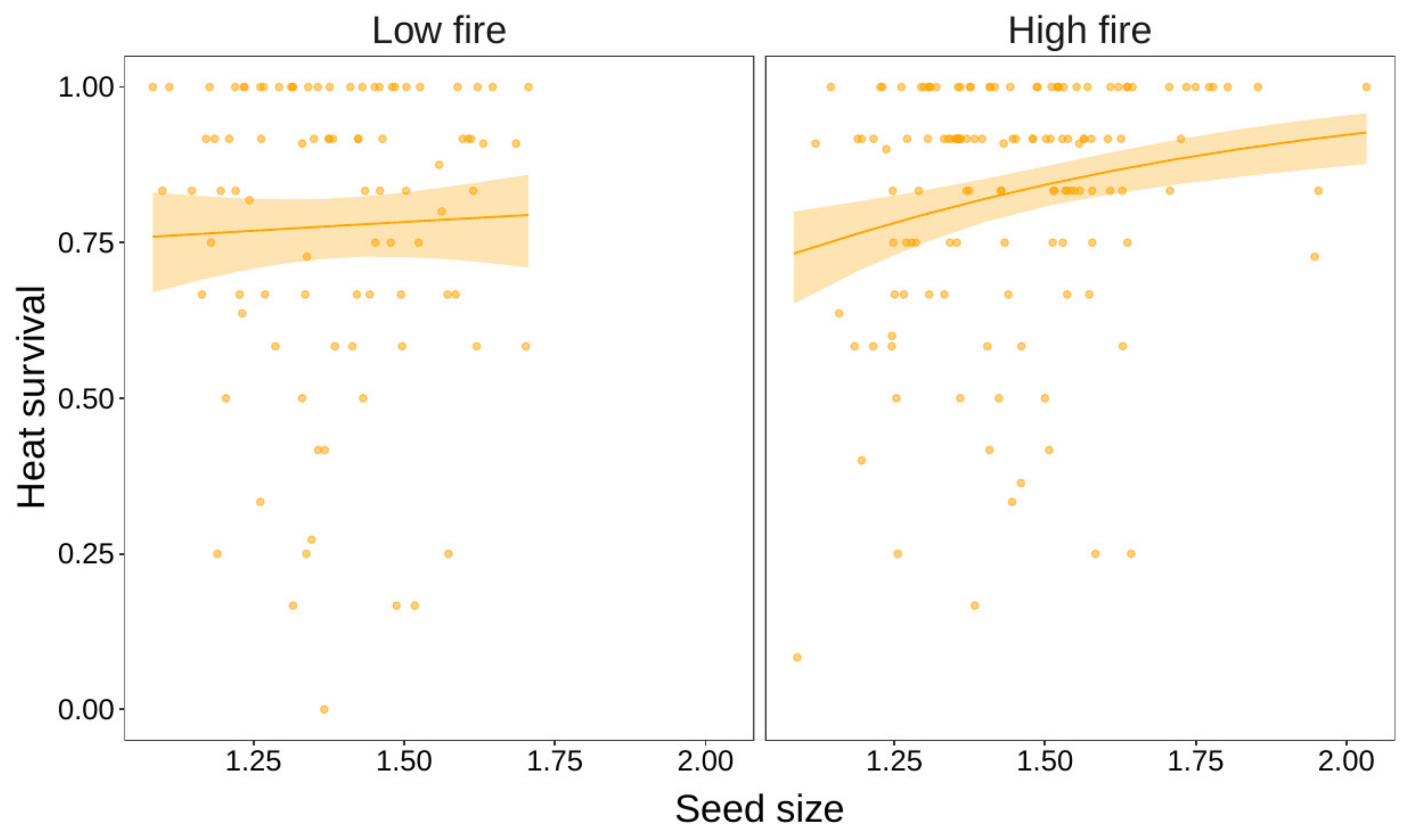
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cowling, R.M.; Rundel, P.W.; Lamont, B.B.; Arroyo, M.K.; Arianoutsou, M. Plant diversity in Mediterranean-climate regions. Trends Ecol. Evol. 1996, 11, 362–366. [Google Scholar] [CrossRef]
- Keeley, J.E.; Bond, W.J.; Bradstock, R.A.; Pausas, J.G.; Rundel, P.W. Fire in Mediterranean Ecosystems: Ecology, Evolution and Management, 1st ed.; Cambridge University Press: New York, NY, USA, 2012; 515p. [Google Scholar]
- Keeley, J.E.; Pausas, J.G.; Rundel, P.W.; Bond, W.L.; Bradstock, R.A. Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 2011, 16, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Mooney, H.A.; Dunn, E.L. Convergent evolution of Mediterranean-climate evergreen sclerophyll shrubs. Evolution 1970, 24, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Paula, S.; Naulin, P.I.; Arce, C.; Galaz, C.; Pausas, J.G. Lignotubers in Mediterranean Basin plants. Plant Ecol. 2016, 217, 661–676. [Google Scholar] [CrossRef]
- Pausas, J.G. Bark thickness and fire regime. Funct. Ecol. 2015, 29, 315–327. [Google Scholar] [CrossRef]
- Lamont, B.B.; Le Maitre, D.C.; Cowling, R.M.; Enright, N.J. Canopy seed storage in woody plants. Bot. Rev. 1991, 57, 277–317. [Google Scholar] [CrossRef]
- Keeley, J.E.; Fotheringham, C.J. Role of fire in regeneration from seed. In Seeds: The Ecology of Regeneration in Plant Communities, 1st ed.; Fenner, M., Ed.; CAB International: Oxford, UK, 2000; pp. 311–330. [Google Scholar]
- Pausas, J.G.; Keeley, J.E. Evolutionary ecology of resprouting and seeding in fire-prone ecosystems. N. Phytol. 2014, 204, 55–65. [Google Scholar] [CrossRef]
- Flematti, G.R.; Dixon, K.W.; Smith, S.M. What are karrikins and how were they ’discovered’ by plants? BMC Biol. 2015, 13, 108. [Google Scholar] [CrossRef]
- Moreira, B.; Pausas, J.G. Tanned or burned: The role of fire in shaping physical seed dormancy. PLoS ONE 2012, 7, e51523. [Google Scholar] [CrossRef]
- Bell, D.T.; Williams, D.S. Tolerance of thermal shock in seeds. Aust. J. Bot. 1998, 46, 221–233. [Google Scholar] [CrossRef]
- Gómez-González, S.; Torres-Díaz, C.; Bustos-Schindler, C.; Gianoli, E. Anthropogenic fire drives the evolution of seed traits. Proc. Natl. Acad. Sci. USA 2011, 108, 18743–18747. [Google Scholar] [CrossRef] [PubMed]
- Tangney, R.; Merritt, D.J.; Fontaine, J.B.; Miller, B.P. Seed moisture content as a primary trait regulating the lethal temperature thresholds of seeds. J. Ecol. 2019, 107, 1093–1105. [Google Scholar] [CrossRef]
- Hanley, M.; Unna, J.; Darvill, B. Seed size and germination response: A relationship for fire-following plant species exposed to thermal shock. Oecologia 2003, 134, 18–22. [Google Scholar] [PubMed]
- Tavşanoğlu, C.; Çatav, Ş.S. Seed size explains within-population variability in post-fire germination of Cistus salviifolius. Ann. Bot. Fenn. 2012, 49, 331–340. [Google Scholar] [CrossRef]
- Liyanage, G.S.; Ooi, M.K. Seed size-mediated dormancy thresholds: A case for the selective pressure of fire on physically dormant species. Biol. J. Linn. Soc. 2018, 123, 135–143. [Google Scholar] [CrossRef]
- Delgado, J.A.; Serrano, J.M.; López, F.; Acosta, F.J. Heat shock, mass-dependent germination, and seed yield as related components of fitness in Cistus ladanifer. Environ. Exp. Bot. 2001, 46, 11–20. [Google Scholar] [CrossRef]
- Ruprecht, E.; Fenesi, A.; Fodor, E.I.; Kuhn, T.; Tökölyi, J. Shape determines fire tolerance of seeds in temperate grasslands that are not prone to fire. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 397–404. [Google Scholar] [CrossRef]
- Gómez-González, S.; Ojeda, F.; Torres-Morales, P.; Palma, J.E. Seed pubescence and shape modulate adaptative responses to fire cues. PLoS ONE 2016, 11, e0159655. [Google Scholar] [CrossRef]
- Cochrane, A.; Yates, C.J.; Hoyle, G.L.; Nicotra, A.B. Will among-population variation in seed traits improve the chance of species persistence under climate change? Glob. Ecol. Biogeogr. 2015, 24, 12–24. [Google Scholar] [CrossRef]
- Moreira, B.; Tavsanoglu, Ç.; Pausas, J.G. Local versus regional intraspecific variability in regeneration traits. Oecologia 2012, 168, 671–677. [Google Scholar] [CrossRef]
- Leonard, J.; West, A.G.; Ojeda, F. Differences in germination response to smoke and temperature cues in ’pyrophyte’ and ’pyrofuge’ forms of Erica coccinea (Ericaceae). Int. J. Wild. Fire 2018, 27, 562–568. [Google Scholar] [CrossRef]
- Ojeda, F.; Pausas, J.G.; Verdú, M. Soil shapes community structure through fire. Oecologia 2010, 163, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Cowling, R.M.; Kirkwood, D.; Midgley, J.J.; Pierce, S.M. Invasion and persistence of bird-dispersed, subtropical thicket and forest species in fire-prone coastal fynbos. J. Veg. Sci. 1997, 8, 475–488. [Google Scholar] [CrossRef]
- Kraaij, T.; van Wilgen, B.W. Drivers, ecology, and management of fire in fynbos. In Fynbos: Ecology, Evolution, and Conservation of a Megadiverse Region, 1st ed.; Allsopp, N., Colville, J.F., Verboom, G.A., Eds.; Oxford University Press: Oxford, UK, 2014; pp. 47–72. [Google Scholar]
- Naveh, Z. The evolutionary significance of fire in the Mediterranean region. Vegetatio 1975, 29, 199–208. [Google Scholar] [CrossRef]
- Trabaud, L. Postfire plant community dynamics in the Mediterranean Basin. In The Role of Fire in Mediterranean-type Ecosystems, 1st ed.; Moreno, J.M., Oechel, W.C., Eds.; Springer: New York, NY, USA, 1994; pp. 1–15. [Google Scholar]
- Pausas, J.G.; Verdú, M. Plant persistence traits in fire-prone ecosystems of the Mediterranean basin: A phylogenetic approach. Oikos 2005, 109, 196–202. [Google Scholar] [CrossRef]
- Verdú, M.; Pausas, J.G. Fire drives phylogenetic clustering in Mediterranean Basin woody plant communities. J. Ecol. 2007, 95, 1316–1323. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E.; Verdú, M. Inferring differential evolutionary processes of plant persistence traits in Northern Hemisphere Mediterranean fire-prone ecosystems. J. Ecol. 2006, 94, 31–39. [Google Scholar] [CrossRef]
- Gil-López, M.J.; Segarra-Moragues, J.G.; Ojeda, F. Floristic distinctiveness and endemic richness of woody plants highlight the biodiversity value of the herriza among all Mediterranean heathlands. Plant Ecol. Div. 2018, 11, 111–119. [Google Scholar] [CrossRef]
- Gil-López, M.; Segarra-Moragues, J.G.; Ojeda, F. Influence of habitat patchiness on diversity patterns of a habitat specialist plant community. J. Veg. Sci. 2017, 28, 436–444. [Google Scholar] [CrossRef]
- Santos, L.; Capelo, J.; Tavares, M. Germination patterns of soil seed banks in relation to fire in Portuguese littoral pine forest vegetation. Fire Ecol. 2010, 6, 1–15. [Google Scholar] [CrossRef]
- Thanos, C.A.; Georghiou, K. Ecophysiology of fire-stimulated seed germination in Cistus incanus ssp. creticus (L.) Hey wood and C. salvifolius L. Plant Cell Environ. 1988, 11, 841–849. [Google Scholar] [CrossRef]
- Céspedes, B.; Torres, I.; Luna, B.; Pérez, B.; Moreno, J.M. Soil seed bank, fire season, and temporal patterns of germination in a seeder-dominated Mediterranean shrubland. Plant Ecol. 2012, 213, 383–393. [Google Scholar] [CrossRef]
- Moretti, M.; Conedera, M.; Moresi, R.; Guisan, A. Modelling the influence of change in fire regime on the local distribution of a Mediterranean pyrophytic plant species (Cistus salviifolius) at its northern range limit. J. Biogeogr. 2006, 33, 1492–1502. [Google Scholar] [CrossRef]
- Demoly, J.P.; Montserrat, P. Cistus L. In Flora Ibérica, 1st ed.; Castroviejo, S., Aedo, C., Laínz, M., Muñoz Garmendia, F., Nieto Feliner, G., Paiva, J., Benedí, C., Eds.; Real Jardín Botánico, CSIC: Madrid, Spain, 1993; Volume 3, pp. 319–337. [Google Scholar]
- Valdés, B.; Talavera, S.; Fernández-Galiano, E. Flora Vascular de Andalucía Occidental II, 1st ed.; Ketres: Barcelona, Spain, 1987; p. 340. [Google Scholar]
- REDIAM. Red de Información Ambiental de Andalucía. Junta de Andalucía. Available online: http://www.juntadeandalucia.es (accessed on 14 June 2019).
- De Castro, R.; Simarro, M.E.; Priego, C.; Lafuente, R.; Sancho, A. Investigación social sobre los incendios forestales en Andalucía. In Proceedings of the Wildfire 2007 IV Conferencia Internacional sobre Incendios Forestales, Seville, Spain, 13–17 May 2007. [Google Scholar]
- Vuong, Q.H. Likelihood ratio tests for model selection and non-nested hypotheses. Econometrica 1989, 57, 307–333. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: http://www.R-project.org (accessed on 22 January 2020).
- Bates, D.; Sarkar, D.; Bates, M.D.; Matrix, L. The lme4 package. R Package Version 2007, 2, 74. [Google Scholar]
- Tavşanoğlu, Ç.; Gürkan, B. Post-fire dynamics of Cistus spp. in a Pinus brutia forest. Turkish J. Bot. 2005, 29, 337–343. [Google Scholar]
- Ojeda, F.; Marañón, T.; Arroyo, J. Postfire regeneration of a Mediterranean heathland in southern Spain. Int. J. Wild. Fire 1996, 6, 191–198. [Google Scholar] [CrossRef]
- Ojeda, F. Mediterranean heathland and fynbos: A neglected example of convergence between mediterranean climate regions (contributed case study). In The Biology of Mediterranean-Type Ecosystems, 1st ed.; Esler, K.J., Jacobsen, A.L., Pratt, R.B., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 160–163. [Google Scholar]
- Paniw, M.; Quintana-Ascencio, P.F.; Ojeda, F.; Salguero-Gómez, R. Interacting livestock and fire may both threaten and increase viability of a fire-adapted Mediterranean carnivorous plant. J. Appl. Ecol. 2017, 54, 1884–1894. [Google Scholar] [CrossRef]
- Delgado, J.A.; Serrano, J.M.; López, F.; Acosta, F.J. Seed size and seed germination in the Mediterranean fire-prone shrub Cistus ladanifer. Plant Ecol. 2008, 197, 269–276. [Google Scholar] [CrossRef]
- Escudero, A.; Núñez, Y.; Pérez-García, F. Is fire a selective force of seed size in pine species? Acta Oecol. 2000, 21, 245–256. [Google Scholar] [CrossRef]
- Thanos, C.A.; Georghiou, K.; Kadis, C.; Pantazi, C. Cistaceae: A plant family with hard seeds. Isr. J. Plant Sci. 1992, 41, 251–263. [Google Scholar]
- Pérez-García, F.; Escudero, A. Role of the seed coat in germination of Cistus populifolius L. Isr. J. Plant Sci. 1997, 45, 329–331. [Google Scholar] [CrossRef]
- Luna, B.; Chamorro, D.; Pérez, B. Effect of heat on seed germination and viability in species of Cistaceae. Plant Ecol. Div. 2019, 12, 151–158. [Google Scholar] [CrossRef]
- Grundy, A.C.; Mead, A.; Burston, S. Modelling the emergence response of weed seeds to burial depth: Interactions with seed density, weight and shape. J. Appl. Ecol. 2003, 40, 757–770. [Google Scholar] [CrossRef]
- Saatkamp, A.; Cochrane, A.; Commander, L.; Guja, L.K.; Jimenez-Alfaro, B.; Larson, J.; Nicotra, A.; Poschlod, P.; Silveira, F.A.O.; Cross, A.T.; et al. A research agenda for seed-trait functional ecology. N. Phytol. 2019, 221, 1764–1775. [Google Scholar] [CrossRef]
- Jiménez-Alfaro, B.; Silveira, F.A.; Fidelis, A.; Poschlod, P.; Commander, L.E. Seed germination traits can contribute better to plant community ecology. J. Veg. Sci. 2016, 27, 637–645. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | No. of Parameters | Deviance | χ2 | p |
|---|---|---|---|---|
| Intercept | 4 | 2086.7 | ||
| Heat | 5 | 2041.8 | 44.91 | <0.001 |
| Heat + Size | 6 | 2041.6 | 0.21 | 0.645 |
| Heat + Habitat | 6 | 2040.5 | 1.29 | 0.255 |
| Heat × Size | 7 | 2013.2 | 28.63 | <0.001 |
| Heat × Habitat | 7 | 2036.7 | 5.16 | 0.075 |
| Heat × Size × Habitat | 11 | 2000.2 | 12.97 | 0.011 |
| Dormancy Strength | Heat Survival | |||||||
|---|---|---|---|---|---|---|---|---|
| No. of Parameters | Deviance | χ2 | p | No. of Parameters | Deviance | χ2 | p | |
| Intercept | 2 | 1182.8 | 2 | 1215.5 | ||||
| Habitat | 3 | 1164.1 | 1.69 | 0.194 | 3 | 1212.7 | 2.71 | 0.100 |
| Size | 3 | 1181.1 | 17.0 | <0.001 | 3 | 1203.8 | 11.68 | <0.001 |
| Size × Habitat | 5 | 1156.1 | 8.07 | 0.005 | 5 | 1199.0 | 4.80 | 0.090 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-González, S.; Paniw, M.; Durán, M.; Picó, S.; Martín-Rodríguez, I.; Ojeda, F. Mediterranean Heathland as a Key Habitat for Fire Adaptations: Evidence from an Experimental Approach. Forests 2020, 11, 748. https://doi.org/10.3390/f11070748
Gómez-González S, Paniw M, Durán M, Picó S, Martín-Rodríguez I, Ojeda F. Mediterranean Heathland as a Key Habitat for Fire Adaptations: Evidence from an Experimental Approach. Forests. 2020; 11(7):748. https://doi.org/10.3390/f11070748
Chicago/Turabian StyleGómez-González, Susana, Maria Paniw, Mario Durán, Sergio Picó, Irene Martín-Rodríguez, and Fernando Ojeda. 2020. "Mediterranean Heathland as a Key Habitat for Fire Adaptations: Evidence from an Experimental Approach" Forests 11, no. 7: 748. https://doi.org/10.3390/f11070748
APA StyleGómez-González, S., Paniw, M., Durán, M., Picó, S., Martín-Rodríguez, I., & Ojeda, F. (2020). Mediterranean Heathland as a Key Habitat for Fire Adaptations: Evidence from an Experimental Approach. Forests, 11(7), 748. https://doi.org/10.3390/f11070748