A Comparative Evaluation of Three Stem Profile Equations for Three Precious Tree Species in Southern China


Abstract
1. Introduction
2. Materials and Methods
2.1. Data Source and Processing
2.2. Model Construction and Test Method
2.2.1. Basic Model
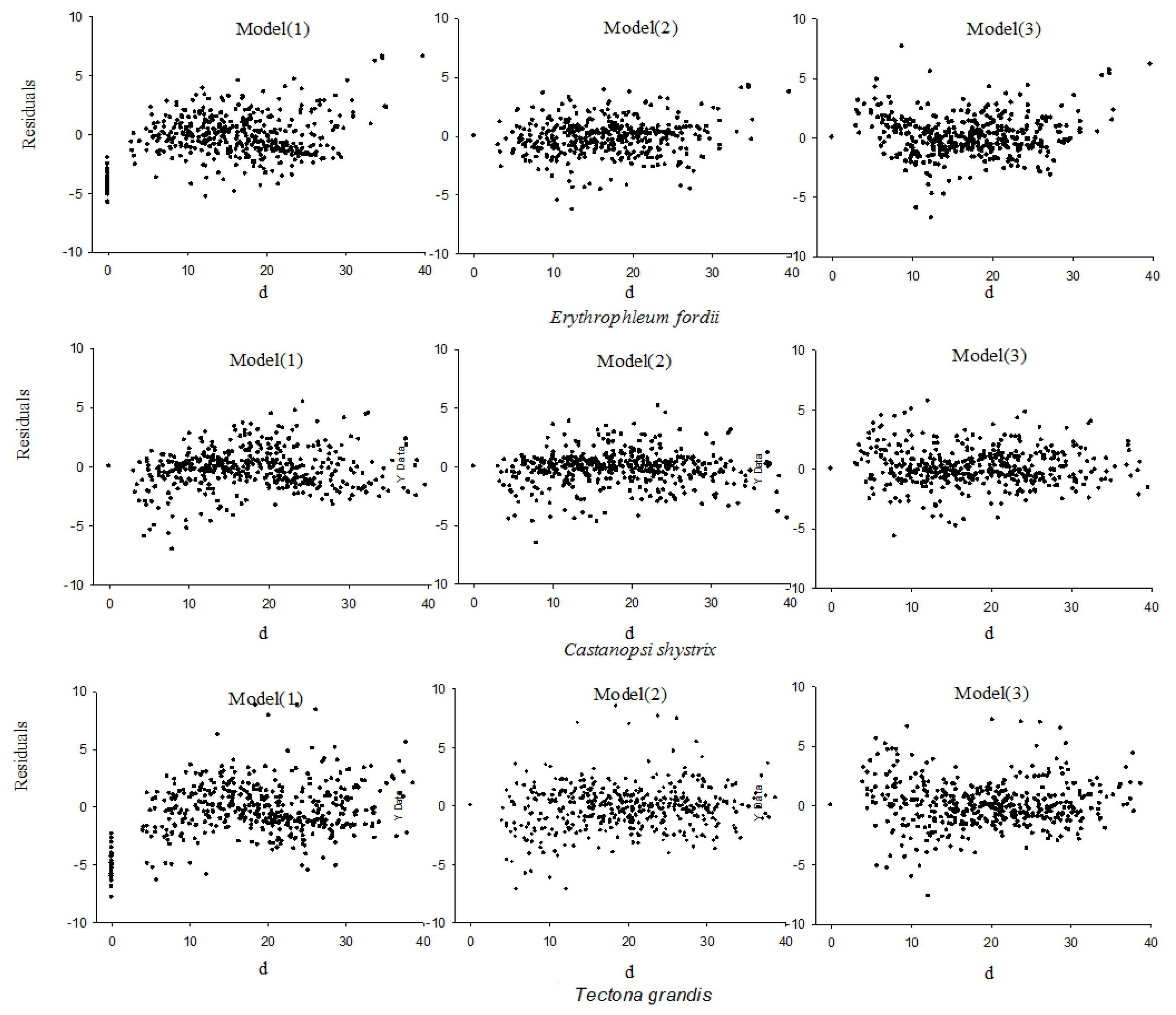
2.2.2. Model Evaluation and Test Indicators
3. Results
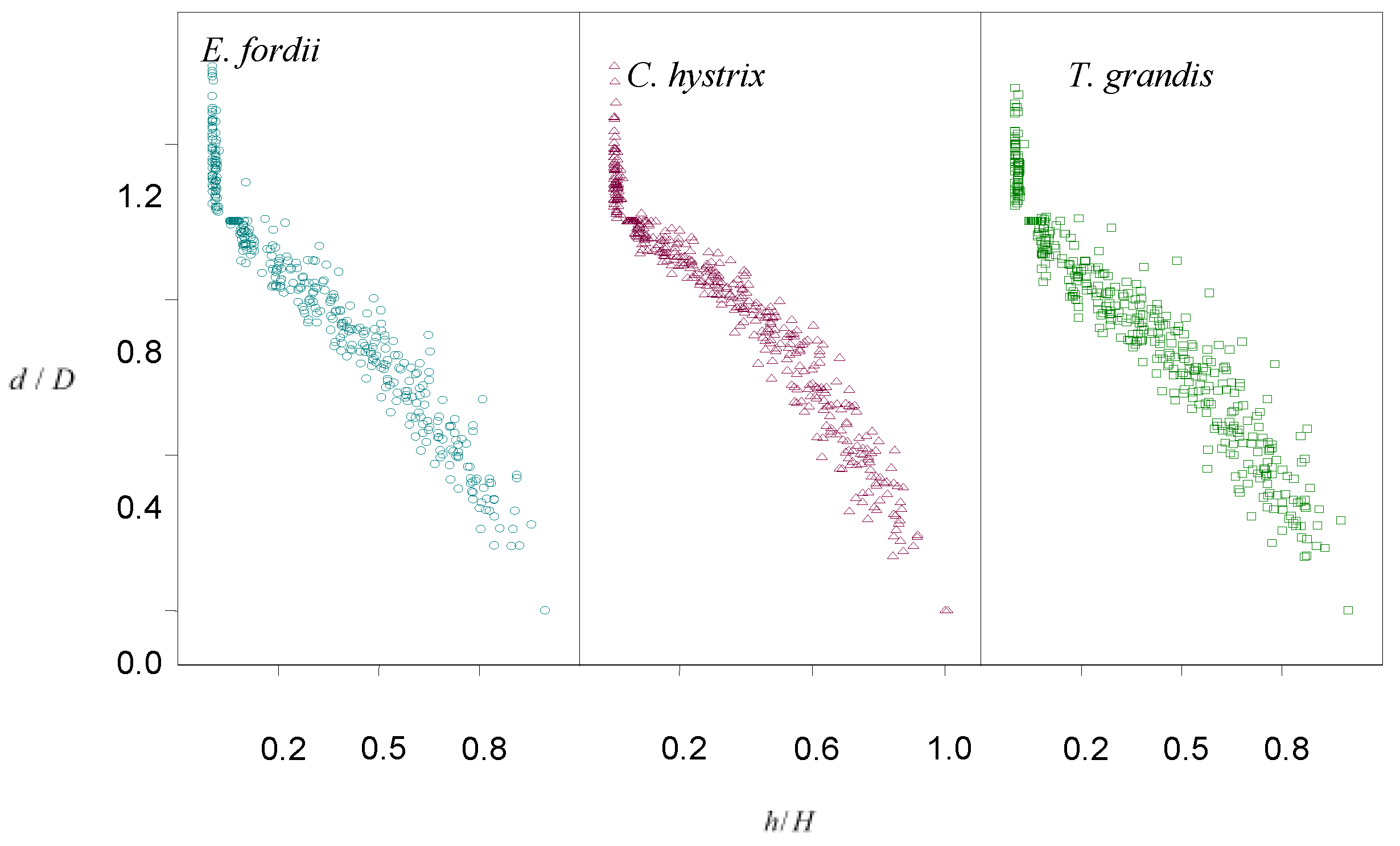
3.1. Selection of the Stem Curve Models for Three Tree Species
3.2. Parameter Estimation
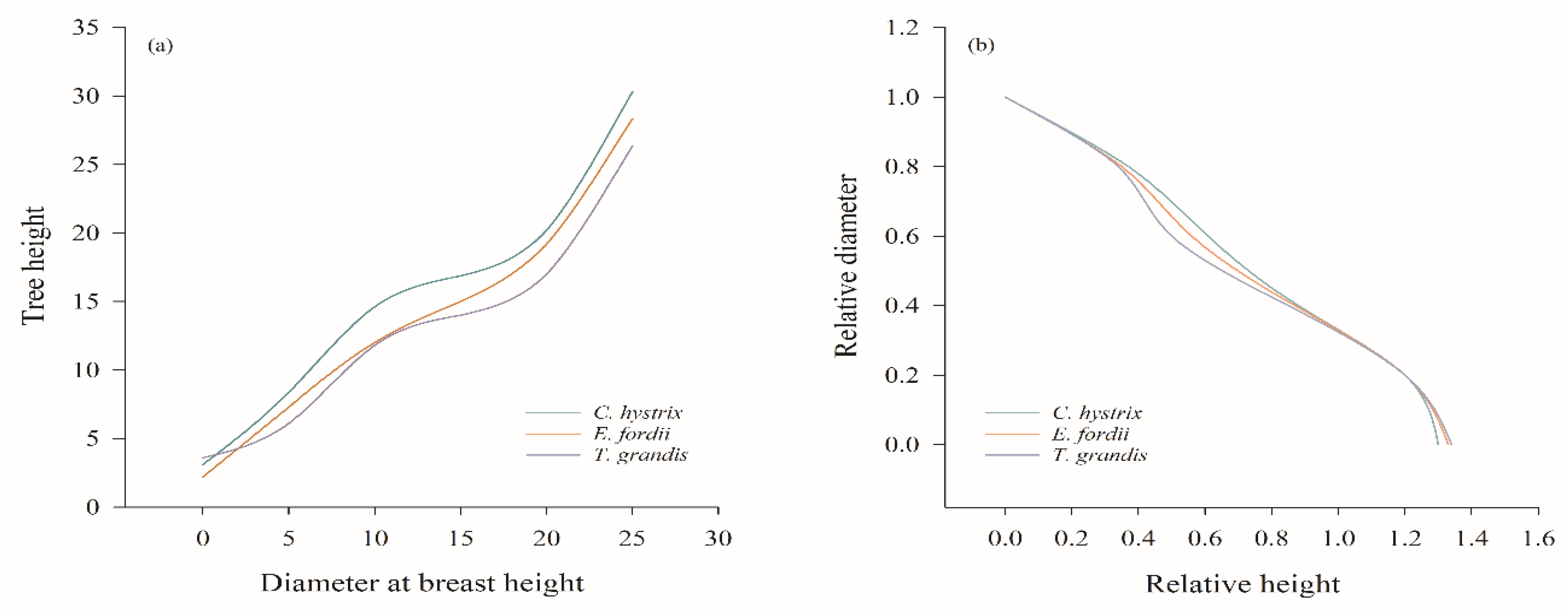
3.3. Stem Analysis of Three Invaluable Tree Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bai, J.P. Investigation Report of rare species in Guangdong, Fujian. Shanxi For. Sci. Technol. 2009, 38, 51–52. [Google Scholar]
- Chen, X.T. Study on Development Prospects of rare species Plantation in Guangxi. For. Sci. Technol. 2012, 37, 53–55. [Google Scholar]
- Shook, S.R.; Zhang, Y.; Wagner, F.G. Wood products cooperatives: Overview and exploratory analysis. For. Prod. J. 2001, 51, 25–33. [Google Scholar]
- Bowers, S. Increased value through optimal bucking. West. J. Appl. For. 1998, 13, 85–89. [Google Scholar] [CrossRef]
- Li, R.; Weiskittel, A.R. Comparison of model forms for estimating stem taper and volume in the primary conifer species of the North American Acadian Region. Ann. For. Sci. 2010, 67, 1–16. [Google Scholar] [CrossRef]
- Scolforo, H.F.; McTague, J.P.; Burkhart, H.; Roise, J.; Carneiro, R.L.; Stape, J.L. Generalized stem taper and tree volume equations applied to eucalyptus of varying genetics in Brazil. Can. J. For. Res. 2019, 49, 447–462. [Google Scholar] [CrossRef]
- Kozak, A.; Munro, D.O.; Smith, J.H.G. Taper functions and their application in forest inventory. For. Chron. 1969, 45, 278–283. [Google Scholar] [CrossRef]
- Özçelik, R.; Brooks, J.R.; Jiang, L. Modeling stem profile of Lebanon cedar, Brutian pine, and Cilicica fir in Southern Turkey using nonlinear mixed-effects models. Eur. J. For. Res. 2011, 130, 613–621. [Google Scholar] [CrossRef]
- Poudel, K.P.; Temesgen, H.; Gray, A.N. Estimating upper stem diameters and volume of Douglas-fir and Western hemlock trees in the Pacific northwest. For. Ecosyst. 2018, 5, 3–14. [Google Scholar] [CrossRef]
- Amidon, E.L. A general taper functional form to predict bole volume for five mixed-conifer species in Califonia. For. Sci. 1984, 30, 166–171. [Google Scholar]
- Max, T.A.; Burkhart, H.E. Segmented polynomial regression applied to taper equations. For. Sci. 1976, 22, 283–289. [Google Scholar]
- Cao, Q.V.; Burkhart, H.E.; Max, T.A. Evaluations of two methods for cubic volume prediction of loblolly pine to any merchantable limit. For. Sci. 1980, 26, 71–80. [Google Scholar]
- Brooks, J.R.; Jiang, L.; Ozçelik, R. Compatible stem volume and taper equations for Brutian pine, Cedar of Lebanon, and Cilicica fir in Turkey. For. Ecol. Manag. 2008, 256, 147–151. [Google Scholar] [CrossRef]
- Jiang, L.C.; Liu, R.L. A Stem Taper Model with Nonlinear Mixed Effects for Dahurian larch. Sci. Silvae Sin. 2011, 47, 101–106. [Google Scholar]
- Jiang, L.; Liu, R. Segmented taper equations with crown ratio and stand density for Dahurian Larch (Larix gmelinii) in Northeastern China. J. For. Res. 2011, 22, 347–352. [Google Scholar] [CrossRef]
- Vasilescu, M.M.; Teresneu, C.C.; Dinulica, F. A rapid method for estimating the median diameter of the stem profile of Norway spruce (Picea abies Karst) trees. iFor. Biogeosci. For. 2017, 10, e1–e6. [Google Scholar] [CrossRef]
- Yan, R.H.; Wu, F.Z. Research on commodity volume estimation system variant. J. Nanjing For. Univ. 1992, 16, 31–37. [Google Scholar]
- Muhairwe, C.K. Tree form and taper variation over time for interior lodgepole pine. Can. J. For. Res. 1994, 24, 1904–1913. [Google Scholar] [CrossRef]
- Kozak, A. A variable-exponent taper equation. Can. J. For. Res. 1988, 18, 1363–1368. [Google Scholar] [CrossRef]
- Valentine, H.T.; Gregoire, T.G. A switching model of bole taper. Can. J. For. Res. 2001, 31, 1400–1409. [Google Scholar] [CrossRef]
- Wang, M.L. Theoretical bucking: Taper equations and merchantable volume tables. For. Res. 1998, 11, 271–276. [Google Scholar]
- Hu, C.X.; Yang, S.L.; Jia, W.W. Stem profile model and variable-exponent of Larix gmelinii plantation. Chin. J. Appl. Ecol. 2011, 22, 1695–1701. [Google Scholar]
- Lee, W.-K.; Seo, J.-H.; Son, Y.-M.; Lee, K.-H.; von Gadow, K. Modeling stem profiles for Pinus densiflora in Korea. For. Ecol. Manag. 2003, 172, 69–77. [Google Scholar] [CrossRef]
- Jiang, J.; Lu, Y.C.; Wang, L.; Cai, D.; Jia, H.; Ming, A.; Chen, B. Facilitation by Tree Species in Variable Retention Harvesting for the Restoration of Monoculture Plantations in Southern China. Trop. Conserv. Sci. 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Pang, L.F. Stand Harvest Analysis and Information Management System for Target Tree; Chinese Academy of Forestry: Beijing, China, 2015. [Google Scholar]
- Tang, S.Z.; Lang, K.J.; Li, H.K. Statistics and Computation of Biomathematical Models (Forstat Course); Science Press: Beijing, China, 2009; pp. 56–79. [Google Scholar]
- Tasissa, G.; Burkhart, H.E.; Amateis, R.L. Volume and taper equations for thinned and unthinned loblolly pine trees in cotover, site-prepared plantations. South. J. Appl. For. 1997, 21, 146–152. [Google Scholar]
- Shahzad, M.K.; Hussain, A.; Jiang, L. A model form for stem taper and volume estimates of white birch (Betula platyphylla Sukaczev): A major commercial tree species of Northeast China. Can. J. For. Res. 2020, 50, 274–286. [Google Scholar]
- Arias-Rodil, M.; Diéguez-Aranda, U.; Puerta, F.R.; López-Sánchez, C.A.; Líbano, E.C.; Obregón, A.C.; Castedo-Dorado, F. Modelling and localizing a stem taper function for Pinus radiata in Spain. Can. J. For. Res. 2015, 45, 647–658. [Google Scholar] [CrossRef]
- Klos, R.J.; Wang, G.G.; Dang, Q.L.; East, E.W. Taper equations for five major commercial tree species in Manitoba, Canada. West. J. Appl. For. 2007, 22, 163–170. [Google Scholar] [CrossRef]
- Garber, S.M.; Maguire, D.A. Modeling stem taper of three central Oregon species using nonlinear mixed effects models and autoregressive error structures. For. Ecol. Manag. 2003, 179, 507–522. [Google Scholar] [CrossRef]
- Sharma, M.; Oderwald, R.G. Dimensionally compatible volume and taper equations. Can. J. For. Res. 2001, 31, 797–803. [Google Scholar] [CrossRef]
- Hussein, K.A.; Schmidt, M.; Kotz, H. Parameter parsimo nious taper functions for describing stem profiles. Sci. Silvae Sin. 2008, 44, 20–27. (In Chinese) [Google Scholar]
- Zhang, J.L.; Zhang, E.S.; Zhang, S.C.; Cui, L.-Z.; Yu, B.; Chen, D.-L. A study on the changing values of form index and geometrical solids of tree stem. J. Agric. Univ. Hebei 2007, 30, 66–70. (In Chinese) [Google Scholar]
- Bouriaud, O.; Stefan, G.; Saint-André, L. Comparing local calibration using random effects estimation and Bayesian calibrations: A case study with a mixed effect stem profile model. Ann. For. Sci. 2019, 76, 65. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Tree Species | DBH/cm | Height/m | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Number | Mean ± Standard Deviation (SD) | Max. | Min. | Variation Coefficient (CV) | Mean ± SD | Max. | Min. | CV | |
| E. fordii | 40 | 23.1 ± 3.5 | 30.1 | 9.2 | 0.21 | 17.7 ± 2.1 | 23.1 | 10.3 | 0.25 |
| C. hystrix | 40 | 22.2 ± 5.40 | 36.2 | 7.7 | 0.43 | 22.1 ± 4.1 | 29.4 | 11.2 | 0.18 |
| T. grandis | 40 | 24.3 ± 4.1 | 37.5 | 11.1 | 0.11 | 23.1 ± 2.6 | 26.5 | 9.40 | 0.27 |
| Model | E. fordii | C. hystrix | T. grandis | ||||||
|---|---|---|---|---|---|---|---|---|---|
| (1) | −0.412 | 4.151 | 2.287 | −0.241 | 2.223 | 1.733 | −0.398 | 6.4773 | 1.512 |
| (2) | −0.179 | 2.318 | 1.378 | −0.199 | 2.198 | 1.619 | −0.231 | 4.620 | 0.134 |
| (3) | 0.054 | 2.512 | 1.456 | 0.087 | 2.539 | 1.106 | 0.110 | 4.223 | 1.133 |
| Tree Species | Model | Parameter | Fitting Statistics | ||||||
|---|---|---|---|---|---|---|---|---|---|
| RSS | |||||||||
| E. fordii | (1) | 1.237 | −2.3098 | 1.0688 | 0.922 | 6.990 | |||
| (2) | −2.006 | 1.021 | 262.0113 | −0.712 | 0.0380 | 0.443 | 0.9123 | 5.11 | |
| (3) | 1.241 | −0.334 | 9.0421 | −9.618 | 4.5960 | 0.9321 | 4.360 | ||
| C. hystrix | (1) | 1.236 | −1.555 | 0.5712 | 0.962 | 5.515 | |||
| (2) | −2.458 | 1.7043 | 177.917 | −2.255 | 0.0374 | 0.119 | 0.961 | 2.14 | |
| (3) | 1.108 | −0.152 | 5.347 | −6.4546 | 1.2101 | 0.920 | 2.11 | ||
| T. grandis | (1) | 1.1124 | −2.123 | 1.578 | 0.9306 | 5.340 | |||
| (2) | −1.367 | 1.0861 | 49.6113 | −0.7689 | 0.13 | 0.478 | 0.877 | 2.98 | |
| (3) | 2.879 | −0.116 | 9.245 | −9.23 | 4.55 | 0.93 | 3.13 | ||
| Diameter Class (cm) | Height (m) | Percentage of Outturn (%) | |||
|---|---|---|---|---|---|
| Large Diameter Timber | Medium Size Timber | Small-Sized Timber | Total | ||
| 8 | 7 | 0 | 0 | 71.77 | 71.77 |
| 10 | 15 | 0 | 0 | 79.13 | 79.13 |
| 12 | 17 | 0 | 0 | 86.62 | 86.62 |
| 14 | 16 | 0 | 0 | 88.11 | 88.11 |
| 16 | 19 | 0 | 0 | 88.32 | 88.32 |
| 18 | 19 | 0 | 0 | 89.15 | 89.15 |
| 20 | 21 | 0 | 31.42 | 60.24 | 91.66 |
| 22 | 22 | 0 | 36.93 | 54.19 | 91.12 |
| 24 | 22 | 0 | 54.23 | 37.55 | 91.78 |
| 26 | 22 | 34.07 | 36.94 | 21.91 | 92.92 |
| 28 | 20 | 40.43 | 30.21 | 21.43 | 92.07 |
| 30 | 22 | 50.17 | 22.47 | 18.32 | 90.96 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, J.; Li, J.; Pang, L.; Ming, A. A Comparative Evaluation of Three Stem Profile Equations for Three Precious Tree Species in Southern China. Forests 2020, 11, 669. https://doi.org/10.3390/f11060669
Jiang J, Li J, Pang L, Ming A. A Comparative Evaluation of Three Stem Profile Equations for Three Precious Tree Species in Southern China. Forests. 2020; 11(6):669. https://doi.org/10.3390/f11060669
Chicago/Turabian StyleJiang, Jun, Jie Li, Lifeng Pang, and Angang Ming. 2020. "A Comparative Evaluation of Three Stem Profile Equations for Three Precious Tree Species in Southern China" Forests 11, no. 6: 669. https://doi.org/10.3390/f11060669
APA StyleJiang, J., Li, J., Pang, L., & Ming, A. (2020). A Comparative Evaluation of Three Stem Profile Equations for Three Precious Tree Species in Southern China. Forests, 11(6), 669. https://doi.org/10.3390/f11060669