Role of Root and Stem Base Fungi in Fraxinus angustifolia (Vahl) Dieback in Croatian Floodplain Forests


Abstract
1. Introduction
2. Materials and Methods
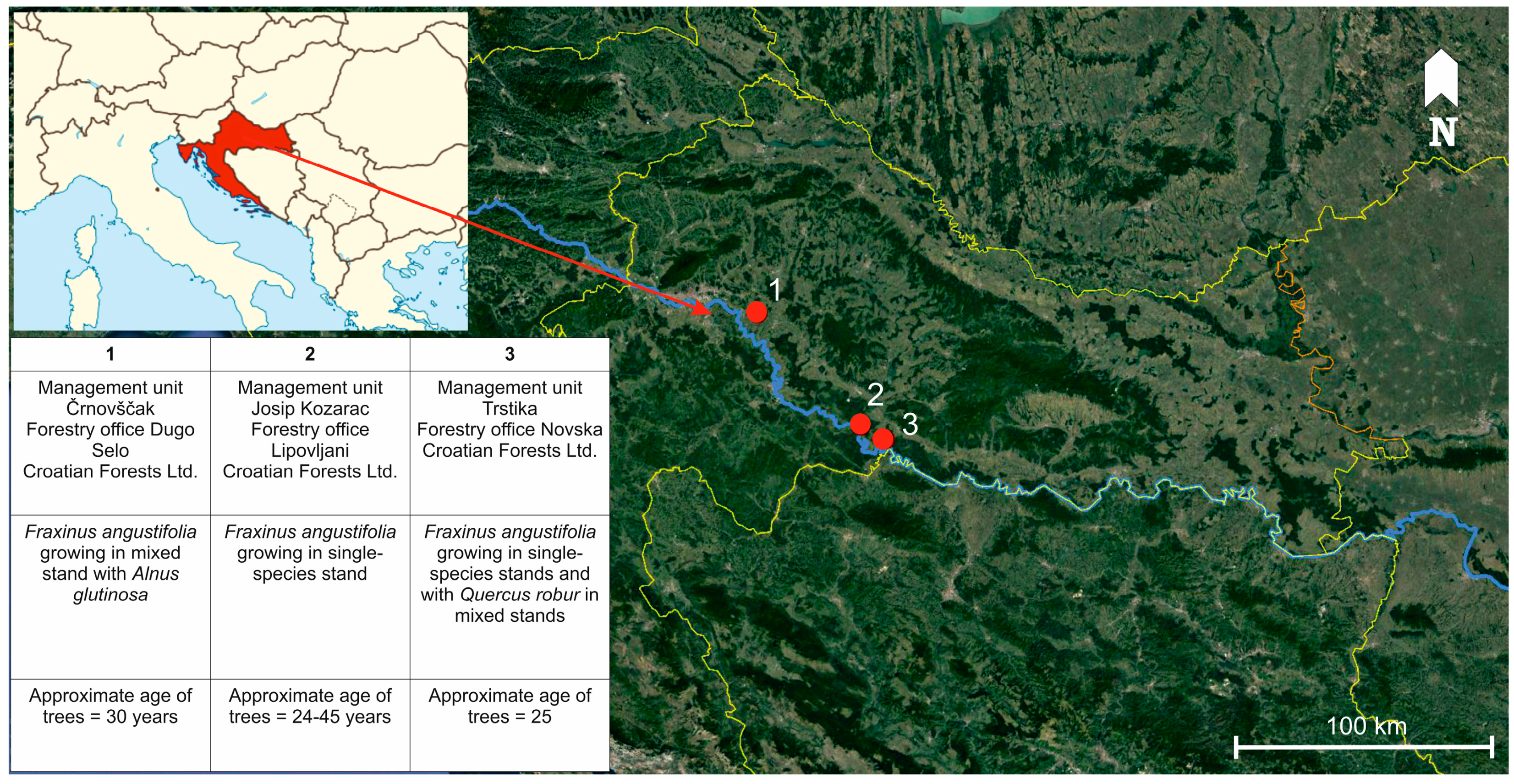
2.1. Field Sampling
2.2. Isolation of Fungi from Tissue Samples
2.3. Identification of Isolated Fungal Taxa
2.4. Statistical Analysis
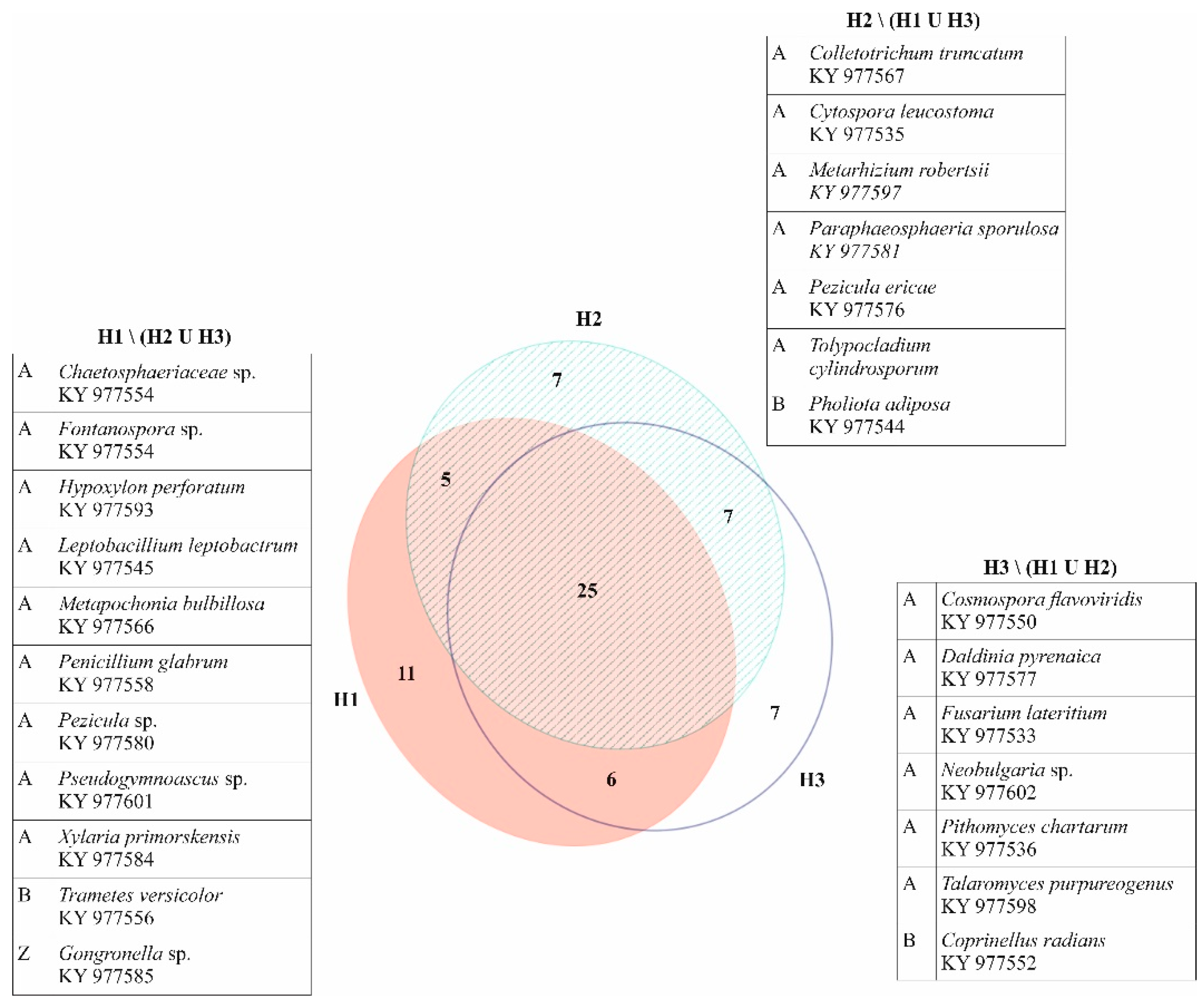
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FRAXIGEN. Ash Species in Europe: Biological Characteristics and Practical Guidelines for Sustainable Use; Oxford Forestry Institute, University of Oxford: Oxford, UK, 2005; pp. 1–128. [Google Scholar]
- Čavlović, J. Prva nacionalna inventura šuma Republike Hrvatske; Ministarstvo Regionalnog Razvoja šumarstva i Vodnoga Gospodarstva i Šumarski Fakultet Sveučilišta u Zagrebu: Zagreb, Hrvatska, 2010; pp. 1–300.
- Potočić, N.; Seletković, I.; Jakovljević, T.; Marjanović, H.; Indir, K.; Medak, J.; Ognjenović, M.; Zorić, N. Oštećenost šumskih ekosustava Republike Hrvatske; Izvješće za 2019. godinu; Nacionalni Koordinacijski Centar za Procjenu i Motrenje Utjecaja Atmosferskog Onečišćenja i drugih čimbenika na šumske Ekosustave, Hrvatski šumarski institut: Jastrebarsko, Croatia, 2020; pp. 1–91. [Google Scholar]
- Kräutler, K.; Kirisits, T. The ash dieback pathogen Hymenoscyphus pseudoalbidus is asociated with leaf symptoms on ash species (Fraxinus spp.). J. Agric. Ext. Rural Dev. 2012, 9, 261–265. [Google Scholar]
- Gross, A.; Holdenrieder, O.; Pautasso, M.; Queloz, V.; Sieber, T.N. H ymenoscyphus pseudoalbidus, the causal agent of E uropean ash dieback. Mol. Plant Pathol. 2013, 15, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, T.; Holdenrieder, O. The teleomorph of Chalara fraxinea, the causal agent of ash dieback. For. Pathol. 2009, 39, 304–308. [Google Scholar] [CrossRef]
- Barić, L.; Diminić, D. Prvi nalaz patogene gljive Chalara fraxinea Kowalski na običnom jasenu (Fraxinus excelsior L.) u Gorskom kotaru. In Proceedings of the 54. Seminar Biljne Zaštite, Opatija, Hrvatska, 9–12 February 2010; p. 33. [Google Scholar]
- Barić, L.; Županić, M.; Pernek, M.; Diminić, D. First records of Chalara fraxinea in Croatia—A new agent of ash dieback (Fraxinus spp.). Šumarski List 2012, 461–469. [Google Scholar]
- Diminić, D. Nova bolest jasena (Fraxinus spp.) u Hrvatskoj. In Proceedings of the Proizvodnja Hrane i šumarstvo—Temelj Razvoja Istočne Hrvatske, Osijek, Hrvatska, 14–15 June 2015; pp. 363–373. [Google Scholar]
- Milotić, M.; Kranjec, J.; Diminić, D. Current status of ash dieback disease Hymenoscyphus fraxineus in Croatia. In Proceedings of the Natural Resources, Green Technology & Sustainable Development—GREEN/2, Zagreb, Hrvatska, 5–7 February 2016; p. 124. [Google Scholar]
- Bakys, R.; Vasaitis, R.; Barklund, P.; Thomsen, I.M.; Stenlid, J. Occurrence and pathogenicity of fungi in necrotic and non-symptomatic shoots of declining common ash (Fraxinus excelsior) in Sweden. Eur. J. For. Res. 2008, 128, 51–60. [Google Scholar] [CrossRef]
- Przybył, K. Fungi associated with necrotic apical parts of Fraxinus excelsior shoots. For. Pathol. 2002, 32, 387–394. [Google Scholar] [CrossRef]
- Davydenko, K.; Vasaitis, R.; Stenlid, J.; Menkis, A. Fungi in foliage and shoots of Fraxinus excelsior in eastern Ukraine: A first report on Hymenoscyphus pseudoalbidus. For. Pathol. 2013, 43, 462–467. [Google Scholar] [CrossRef]
- Pukacki, P.M.; Przybył, K. Frost Injury as a Possible Inciting Factor in Bud and Shoot Necroses of Fraxinus excelsior L. J. Phytopathol. 2005, 153, 512–516. [Google Scholar] [CrossRef]
- Kowalski, T.; Kraj, W.; Bednarz, B. Fungi on stems and twigs in initial and advanced stages of dieback of European ash (Fraxinus excelsior) in Poland. Eur. J. For. Res. 2016, 135, 565–579. [Google Scholar] [CrossRef]
- Hauptman, T.; Piškur, B.; De Groot, M.; Ogris, N.; Ferlan, M.; Jurc, D. Temperature effect on Chalara fraxinea: Heat treatment of saplings as a possible disease control method. For. Pathol. 2013, 43, 360–370. [Google Scholar] [CrossRef]
- Kowalski, T.; Bartnik, C. Morphologial variation in colonies of Chalara fraxinea isolated from ash (Fraxinus excelsior L.) stems with symptoms of dieback and effects of temperature on colony growth and structure. Acta Agrobot. 2012, 63, 99–106. [Google Scholar] [CrossRef]
- Grosdidier, M.; Ioos, R.; Marçais, B. Do higher summer temperatures restrict the dissemination of Hymenoscyphus fraxineus in France? For. Pathol. 2018, 48, e12426. [Google Scholar] [CrossRef]
- Przybył, K. Mycobiota of thin roots showing decay of Fraxinus excelsior L. young trees. Dendrobiology 2002, 48, 65–69. [Google Scholar]
- Kowalski, T.; Bilanski, P.; Kraj, W. Pathogenicity of fungi associated with ash dieback towards Fraxinus excelsior. Plant Pathol. 2017, 66, 1228–1238. [Google Scholar] [CrossRef]
- Kowalski, T.; Łukomska, A. The studies on ash dying (Fraxinus excelsior L.) in the Włoszczowa Forest Unit stands. Acta Agrobot. 2012, 58, 429–440. [Google Scholar] [CrossRef]
- Kowalski, T.; Holdenrieder, O. Pathogenicity of Chalara fraxinea. For. Pathol. 2009, 39, 1–7. [Google Scholar] [CrossRef]
- Bakys, R.; Vasaitis, R.; Barklund, P.; Ihrmark, K.; Stenlid, J. Investigations concerning the role of Chalara fraxineain declining Fraxinus excelsior. Plant Pathol. 2009, 58, 284–292. [Google Scholar] [CrossRef]
- Chandelier, A.; Gerarts, F.; Martin, G.S.; Herman, M.; Delahaye, L. Temporal evolution of collar lesions associated with ash dieback and the occurrence of Armillariain in Belgian forests. For. Pathol. 2016, 46, 289–297. [Google Scholar] [CrossRef]
- Langer, G. Collar rots in forests of Northwest Germany affected by ash dieback. Balt. For. 2017, 23, 4–19. [Google Scholar]
- Enderle, R.; Sander, F.; Metzler, B. Temporal development of collar necroses and butt rot in association with ash dieback. Iforest—Biogeosciences For. 2017, 10, 529–536. [Google Scholar] [CrossRef]
- Marçais, B.; Husson, C.; Godart, L.; Caël, O. Influence of site and stand factors on Hymenoscyphus fraxineus—induced basal lesions. Plant Pathol. 2016, 65, 1452–1461. [Google Scholar] [CrossRef]
- Husson, C.; Caël, O.; Grandjean, J.P.; Nageleisen, L.M.; Marçais, B. Occurrence of Hymenoscyphus pseudoalbidus on infected ash logs. Plant Pathol. 2012, 61, 889–895. [Google Scholar] [CrossRef]
- Skovsgaard, J.P.; Thomsen, I.M.; Skovgaard, I.M.; Martinussen, T. Associations among symptoms of dieback in even-aged stands of ash (Fraxinus excelsiorL.). For. Pathol. 2010, 40, 7–18. [Google Scholar] [CrossRef]
- Bakys, R.; Vasiliauskas, A.; Ihrmark, K.; Stenlid, J.; Menkis, A.; Vasaitis, R. Root rot, associated fungi and their impact on health condition of declining Fraxinus excelsior stands in Lithuania. Scand. J. For. Res. 2010, 26, 128–135. [Google Scholar] [CrossRef]
- Lygis, V.; Vasiliauskas, R.; Larsson, K.-H.; Stenlid, J. Wood-inhabiting fungi in stems of Fraxinus excelsior in declining ash stands of northern Lithuania, with particular reference to Armillaria cepistipes. Scand. J. For. Res. 2005, 20, 337–346. [Google Scholar] [CrossRef]
- Enderle, R.; Peters, F.; Nakou, A.; Metzler, B. Temporal development of ash dieback symptoms and spatial distribution of collar rots in a provenance trial of Fraxinus excelsior. Eur. J. For. Res. 2013, 132, 865–876. [Google Scholar] [CrossRef]
- Hauptman, T.; Ogris, N.; De Groot, M.; Piškur, B.; Jurc, D. Individual resistance of Fraxinus angustifolia clones to ash dieback. For. Pathol. 2016, 46, 269–280. [Google Scholar] [CrossRef]
- Orlikowski, L.B.; Ptaszek, M.; Rodziewicz, A.; Nechwatal, J.; Thinggaard, K.; Jung, T. Phytophthora root and collar rot of mature Fraxinus excelsior in forest stands in Poland and Denmark. For. Pathol. 2011, 41, 510–519. [Google Scholar] [CrossRef]
- Akilli, S.; Serçe, Ç.U.; Katırcıoğlu, Y.Z.; Maden, S.; Katircioğlu, Y.Z. Phytophthora dieback on narrow leaved ash in the Black Sea region of Turkey. For. Pathol. 2013, 43, 252–256. [Google Scholar] [CrossRef]
- Schumacher, J.; Kehr, R.; Leonhard, S. Mycological and histological investigations of Fraxinus excelsior nursery saplings naturally infected by Chalara fraxinea. For. Pathol. 2009, 40, 419–429. [Google Scholar] [CrossRef]
- Anić, I. Uspijevanje i pomlađivanje sastojina poljskog jasena (Fraxinus angustifolia Vahl) u Posavini. Doctoral Thesis, University of Zagreb, Faculty of Forestry, Zagreb, Croatia, 2001. [Google Scholar]
- Eichhorn, J.; Roskams, P.; Potočić, N.; Timmermann, V.; Ferretti, M.; Mues, V.; Szepesi, A.; Durrant, D.; Seletković, I.; Schröck, H.-W.; et al. Part IV: Visual Assessment of Crown Condition and Damaging Agents. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Coordinating Centre, Ed.; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2016; 49p. [Google Scholar]
- Greenfield, M.; Pareja, R.; Ortiz, V.; Jiménez, M.I.G.; Vega, F.E.; Parsa, S. A novel method to scale up fungal endophyte isolations. Biocontrol. Sci. Technol. 2015, 25, 1–6. [Google Scholar] [CrossRef]
- Hoff, J.A.; Klopfenstein, N.; McDonald, G.I.; Tonn, J.R.; Kim, M.-S.; Zambino, P.J.; Hessburg, P.F.; Rogers, J.D.; Peever, T.L.; Carris, L.M. Fungal endophytes in woody roots of Douglas-fir (Pseudotsuga menziesii) and ponderosa pine (Pinus ponderosa). For. Pathol. 2004, 34, 255–271. [Google Scholar] [CrossRef]
- Ježić, M.; Krstin, L.; Rigling, D.; Ćurković-Perica, M. High diversity in populations of the introduced plant pathogen, Cryphonectria parasitica, due to encounters between genetically divergent genotypes. Mol. Ecol. 2011, 21, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Kranjec, J.; Diminić, D.; Hegol, M.; Milotić, M. Gljivama slični organizmi u tlu odumirućih sastojina poljskog jasena (Fraxinus angustifolia Vahl). Šumarski List 2017, 141, 122. [Google Scholar] [CrossRef][Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Philologenetics. In PCR Protocols; Elsevier BV: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Sørensen, T. A method of establishing groups of equal amplitude in plant sociology based on similarity of species and its application to analyses of the vegetation on Danish commons. Biolog. Skri. 1948, 5, 1–34. [Google Scholar]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples; Version 9. User’s Guide and Application; University of Colorado Museum of Natural History : Boulder, CO, USA, 2013. [Google Scholar]
- Micallef, L.; Rodgers, P. eulerAPE: Drawing Area-Proportional 3-Venn Diagrams Using Ellipses. PLoS ONE 2014, 9, e101717. [Google Scholar] [CrossRef]
- Keča, N.; Kirisits, T.; Menkis, A. First report of the invasive ash dieback pathogen Hymenoscyphus fraxineus on Fraxinus excelsior and F. angustifolia in Serbia. Balt. For. 2017, 23, 56–59. [Google Scholar]
- Kirisits, T.; Matlakova, M.; Halmschlager, E.; Lakatos, F.; Mottinger-Kroupa, S. Chalara fraxineaassociated with dieback of narrow-leafed ash (Fraxinus angustifolia). Plant Pathol. 2010, 59, 411. [Google Scholar] [CrossRef]
- Pautasso, M.; Aas, G.; Queloz, V.; Holdenrieder, O. European ash (Fraxinus excelsior) dieback—A conservation biology challenge. Boil. Conserv. 2013, 158, 37–49. [Google Scholar] [CrossRef]
- Coker, T.L.R.; Rozsypálek, J.; Edwards, A.; Harwood, T.P.; Butfoy, L.; Buggs, R.J.A. Estimating mortality rates of European ash (Fraxinus excelsior) under the ash dieback (Hymenoscyphus fraxineus) epidemic. Plants People Planet 2018, 1, 48–58. [Google Scholar] [CrossRef]
- Meyn, R.; Langer, G.J.; Gross, A.; Langer, E.J. Fungal colonization patterns in necrotic rootstocks and stem bases of dieback-affected Fraxinus excelsior L. For. Pathol. 2019, 49, e12520. [Google Scholar] [CrossRef]
- Żółciak, A.; Nowakowska, J.; Pacia, A.; Keča, N.; Oszako, T. Fungi isolated from shoots showing ash dieback in the Wolica Nature Reserve in Poland and artificially inoculated seedlings with Hymenoscyphus fraxineus. Folia For. Pol. 2019, 61, 42–50. [Google Scholar] [CrossRef]
- Haňáčková, Z.; Havrdová, L.; Černý, L.; Zahradník, D.; Koukol, O. Fungal Endophytes in Ash Shoots—Diversity and Inhibition of Hymenoscyphus fraxineus. Balt. For. 2017, 23, 89–106. [Google Scholar]
- Bakys, R.; Vasiliauskas, R.; Barklund, P.; Ihrmark, K.; Stenlid, J. Fungal attacks to root systems and crowns of declining Fraxinus excelsior. Aktuelt fra Skogforskningen 2006, 1-06, 71–72. [Google Scholar]
- Bengtsson, S.B.K.; Barklund, P.; Von Brömssen, C.; Stenlid, J. Seasonal Pattern of Lesion Development in Diseased Fraxinus excelsior Infected by Hymenoscyphus pseudoalbidus. PLoS ONE 2014, 9, e76429. [Google Scholar] [CrossRef]
- Muñoz, F.; Marçais, B.; Dufour, J.; Dowkiw, A. Rising Out of the Ashes: Additive Genetic Variation for Crown and Collar Resistance toHymenoscyphus fraxineusinFraxinus excelsior. Phytopathology 2016, 106, 1535–1543. [Google Scholar] [CrossRef]
- Milotić, M. The Role of Fungus Hymenoscyphus Fraxineus (T. Kowalski) Baral, Queloz & Hosoya in Ash Dieback (Fraxinus spp.) in the Republic of Croatia. Doctoral Thesis, University of Zagreb, Faculty of Forestry, Zagreb, Croatia, 2017. [Google Scholar]
- Thomas, H.E. Studies on Armillaria mellea (Vahl) Quel., infection, parasitism, and host resistance. J. Agric. Res. 1934, 48. [Google Scholar]
- Sipos, G.; Anderson, J.B.; Nagy, L.G. Armillaria. Curr. Boil. 2018, 28, R297–R298. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species–opportunistic, avirulent plant symbionts. Nat. Rev. Genet. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Elad, Y.; Chet, I.; Henis, Y. Degradation of plant pathogenic fungi by Trichoderma harzianum. Can. J. Microbiol. 1982, 28, 719–725. [Google Scholar] [CrossRef]
- Ragazzi, A.; Moricca, S.; Capretti, P.; Dellavalle, I.; Turco, E. Differences in composition of endophytic mycobiota in twigs and leaves of healthy and declining Quercus species in Italy. For. Pathol. 2003, 33, 31–38. [Google Scholar] [CrossRef]
- Gennaro, M.; Gonthier, P.; Nicolotti, G. Fungal Endophytic Communities in Healthy and Declining Quercus robur L. and Q. cerris L. Trees in Northern Italy. J. Phytopathol. 2003, 151, 529–534. [Google Scholar] [CrossRef]
- Giordano, L.; Gonthier, P.; Varese, G.C.; Miserere, L.; Nicolotti, G. Mycobiota inhabiting sapwood of healthy and declining Scots pine (Pinus sylvestris L.) trees in the Alps. Fungal Divers. 2009, 38, 69–83. [Google Scholar]
- Moricca, S.; Ginetti, B.; Ragazzi, A. Species- and organ-specificity in endophytes colonizing healthy and declining Mediterranean oaks. Phytopathol. Mediterr. 2012, 51, 587–598. [Google Scholar]
- O’Donnell, K. Molecular phylogeny of the Nectria haematococca-Fusarium solani species complex. Mycologia 2000, 92, 919–938. [Google Scholar] [CrossRef]
- Demirci, F.; Maden, S. A severe dieback of box elder (Acer negundo) caused byFusarium solani(Mart.) Sacc. in Turkey. Australas. Plant Dis. Notes 2006, 1, 13. [Google Scholar] [CrossRef]
- Chen, W.; Swart, W.J. First Report of Stem Canker of English Walnut Caused by Fusarium solani in South Africa. Plant Dis. 2000, 84, 592. [Google Scholar] [CrossRef]
- Abreo, E.; Sessa, L.; Bettucci, L.; Lupo, S.; Kopp, S.M. Phomopsis cotoneastri as a Pathogen Associated with Trunk Cankers and Death of Young Apple Trees cv. Cripps Pink. J. Phytopathol. 2012, 160, 434–436. [Google Scholar] [CrossRef]
- Herder, M.C. Identification of Ilyonectria Species Associated with and Determining Their Role in Avocado Decline. Doctoral Thesis, Lincoln University, Lincoln, New Zealand, 2014. [Google Scholar]
- Manici, L.M.; Kelderer, M.; Caputo, F.; Saccà, M.L.; Nicoletti, F.; Topp, A.R.; Mazzola, M. Involvement of Dactylonectria and Ilyonectria spp. in tree decline affecting multi-generation apple orchards. Plant Soil 2018, 425, 217–230. [Google Scholar] [CrossRef]
- Erper, I.; Agustí-Brisach, C.; Tunali, B.; Armengol, J. Characterization of root rot disease of kiwifruit in the Black Sea region of Turkey. Eur. J. Plant Pathol. 2013, 136, 291–300. [Google Scholar] [CrossRef]
- Marçais, B.; Caël, O.; Delatour, C. Interaction between root rot basidiomycetes and Phytophthora species on pedunculate oak. Plant Pathol. 2010, 60, 296–303. [Google Scholar] [CrossRef]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fungal Biol. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Salgado-Salazar, C.; Crouch, J.A. Genome resources for the stem and bark canker pathogens Corinectria fuckeliana, Neonectria hederae and N. punicea. Plant Dis. 2019, 103, 389–391. [Google Scholar] [CrossRef]
- Castlebury, L.A.; Rossman, A.Y.; Hyten, A.S. Phylogenetic relationships of Neonectria/Cylindrocarpon on Fagus in North America. Botany 2006, 84, 1417–1433. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Phylum | Fungal Taxon | GenBank Accession No. | H1 | H2 | H3 | Total |
|---|---|---|---|---|---|---|---|
| 1 | A | Alternaria sp. | KY977546 | 7 (8) | 2 (2) | 3 (4) | 12 (14) |
| 2 | A | Aspergillus ruber | KY977532 | 4 (5) | 3 (4) | 1 (2) | 8 (11) |
| 3 | A | Beauveria bassiana | KY977586 | 4 (4) | 1 (1) | 1 (1) | 6 (6) |
| 4 | A | Cladosporium cladosporioides | KY977538 | 18 (36) | 11 (19) | 8 (18) | 37 (73) |
| 5 | A | Clonostachys rosea | KY977607 | 6 (6) | 6 (6) | 5 (8) | 17 (20) |
| 6 | A | Diaporthe cotoneastri | KY977583 | 18 (26) | 13 (20) | 9 (13) | 40 (59) |
| 7 | A | Epicoccum nigrum | KY977575 | 4 (5) | 3 (3) | 1 (1) | 8 (9) |
| 8 | A | Fusarium solani | KY977568 | 10 (15) | 16 (42) | 10 (27) | 36 (84) |
| 9 | A | Fusarium sporotrichioides | KY977588 | 3 (3) | 2 (2) | 4 (4) | 9 (9) |
| 10 | A | Fusarium verticillioides | KY977579 | 3 (3) | 4 (4) | 1 (1) | 8 (8) |
| 11 | A | Hymenoscyphus fraxineus | KY977559 | 1 (1) | 4 (7) | 3 (16) | 8 (24) |
| 12 | A | Ilyonectria robusta | KY977528 | 20 (39) | 18 (34) | 18 (39) | 56 (112) |
| 13 | A | Nemania serpens | KY977591 | 2 (3) | 1 (1) | 2 (4) | 5 (8) |
| 14 | A | Neonectria punicea | KY977529 | 2 (2) | 1 (1) | 5 (5) | 8 (8) |
| 15 | A | Penicillium ochrochloron | KY977590 | 5 (5) | 6 (8) | 3 (4) | 14 (17) |
| 16 | A | Penicillium sp. 1 | KY977530 | 3 (3) | 3 (4) | 3 (3) | 9 (10) |
| 17 | A | Pezicula melanigena | KY977594 | 4 (8) | 6 (8) | 2 (2) | 12 (18) |
| 18 | A | Pseudeurotium bakeri | KY977542 | 4 (6) | 1 (2) | 5 (6) | 10 (14) |
| 19 | A | Talaromyces ohiensis | KY977604 | 10 (18) | 5 (6) | 5 (9) | 20 (33) |
| 20 | A | Talaromyces verruculosus | KY977539 | 6 (7) | 9 (11) | 4 (6) | 19 (24) |
| 21 | A | Trichoderma spp. | – | 20 (28) | 22 (49) | 19 (57) | 61 (134) |
| 22 | A | Dichotomopilus funicola | KY977574 | 1 (1) | 2 (2) | – | 3 (3) |
| 23 | A | Penicillium sp. 2 | KY977557 | 1 (1) | 1 (1) | – | 2 (2) |
| 24 | A | Podospora sp. | KY977549 | 1 (1) | 1 (1) | – | 2 (2) |
| 25 | A | Cadophora sp. | KY977578 | 1 (1) | – | 1 (8) | 2 (9) |
| 26 | A | Nemania sp. | KY977540 | 1 (1) | – | 2 (3) | 3 (4) |
| 27 | A | Paraphaeosphaeria neglecta | KY977541 | 1 (2) | – | 1 (1) | 2 (3) |
| 28 | A | Pleosporales sp. | KY977606 | 1 (1) | – | 1 (2) | 2 (3) |
| 29 | A | Tolypocladium album | KY977572 | 1 (1) | – | 1 (1) | 2 (2) |
| 30 | A | Xylaria polymorpha | KY977564 | 1 (1) | – | 1 (1) | 2 (2) |
| 31 | A | Cordyceps confragosa | KY977592 | – | 2 (2) | 2 (3) | 4 (5) |
| 32 | A | Dactylonectria vitis | KY977595 | – | 1 (1) | 2 (3) | 3 (4) |
| 33 | A | Dimorphospora foliicola | KY977582 | – | 1 (1) | 1 (2) | 2 (3) |
| 34 | A | Gliomastix murorum | KY977531 | – | 1 (1) | 1 (1) | 2 (2) |
| 35 | A | Microdiplodia sp. | KY977605 | – | 1 (1) | 1 (4) | 2 (5) |
| 36 | A | Neonectria sp. | KY977537 | – | 2 (2) | 1 (1) | 3 (3) |
| 37 | A | Penicillium manginii | KY977551 | – | 3 (3) | 1 (2) | 4 (5) |
| 38 | B | Armillaria spp. | KY977587 | 1 (5) | 5 (8) | 7 (22) | 13 (35) |
| 39 | B | Coprinellus micaceus | KY977600 | 3 (8) | 5 (11) | 5 (9) | 13 (28) |
| 40 | B | Ganoderma adspersum | KY977553 | 3 (3) | 7 (12) | 3 (5) | 13 (20) |
| 41 | B | Coprinellus disseminatus | KY977599 | 5 (5) | 2 (5) | – | 7 (10) |
| 42 | B | Psathyrella candolleana | KY977573 | 2 (2) | 2 (8) | – | 4 (10) |
| 43 | Z | Absidia sp. | KY977569 | 5 (6) | 7 (7) | 7 (8) | 19 (21) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kranjec Orlović, J.; Moro, M.; Diminić, D. Role of Root and Stem Base Fungi in Fraxinus angustifolia (Vahl) Dieback in Croatian Floodplain Forests. Forests 2020, 11, 607. https://doi.org/10.3390/f11060607
Kranjec Orlović J, Moro M, Diminić D. Role of Root and Stem Base Fungi in Fraxinus angustifolia (Vahl) Dieback in Croatian Floodplain Forests. Forests. 2020; 11(6):607. https://doi.org/10.3390/f11060607
Chicago/Turabian StyleKranjec Orlović, Jelena, Maja Moro, and Danko Diminić. 2020. "Role of Root and Stem Base Fungi in Fraxinus angustifolia (Vahl) Dieback in Croatian Floodplain Forests" Forests 11, no. 6: 607. https://doi.org/10.3390/f11060607
APA StyleKranjec Orlović, J., Moro, M., & Diminić, D. (2020). Role of Root and Stem Base Fungi in Fraxinus angustifolia (Vahl) Dieback in Croatian Floodplain Forests. Forests, 11(6), 607. https://doi.org/10.3390/f11060607