The Spread and Role of the Invasive Alien Tree Quercus rubra (L.) in Novel Forest Ecosystems in Central Europe


Abstract
1. Introduction
2. Materials and Methods
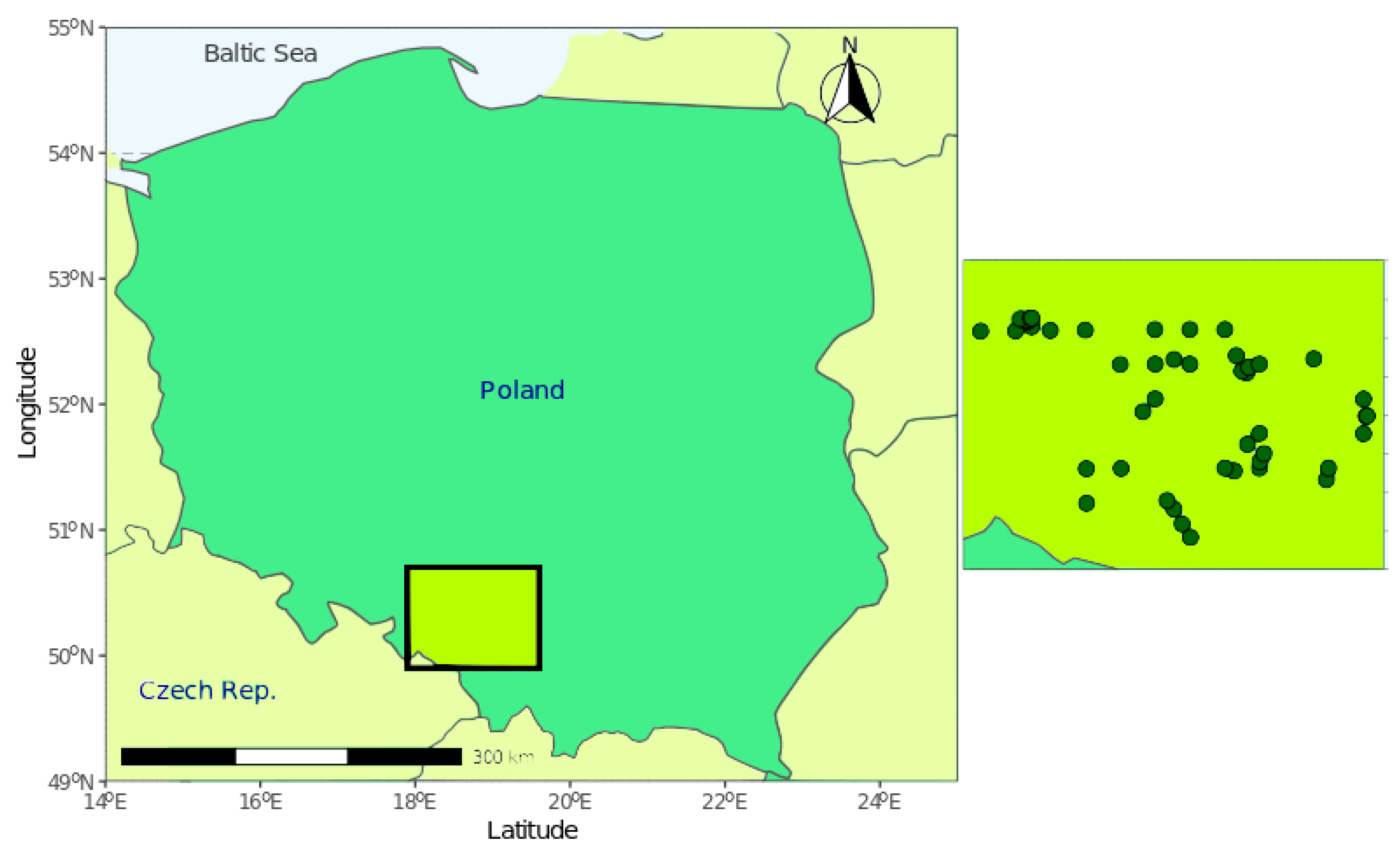
2.1. Study Area
2.2. Data Collection
- Forest interiors: dense patches of a forest community at least 50 m away from any disturbed sites, e.g., forest roadsides and margins;
- Forest roadsides: non-forest habitats that are associated with transport routes (forest paths, edges of roads, and railways). In addition, fringes of drainage and fire-control ditches were included;
- Forest margins: non-forest habitats of a natural or semi-natural origin (borders of a forest area, contact zones with meadows, grasslands and scrub communities, or a contact zone with arable fields). These habitats differ from forest roadsides, due to the fact that the forest habitat is present only on one side;
- Cutting areas: clearings, forest meadows, woodland glades, felling sites, and ruderal sites (dumping sites).
2.3. Statistical Analysis
3. Results
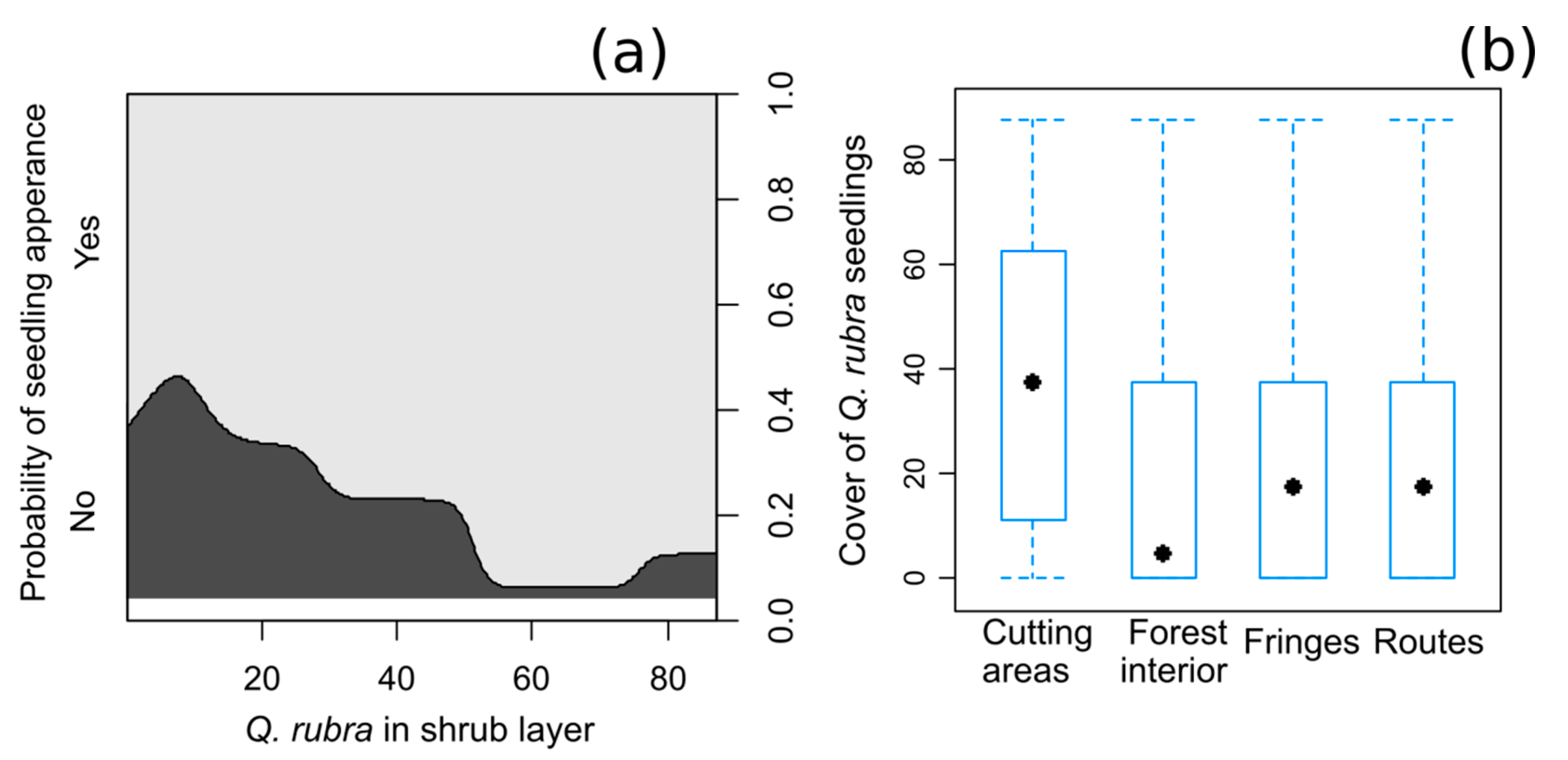
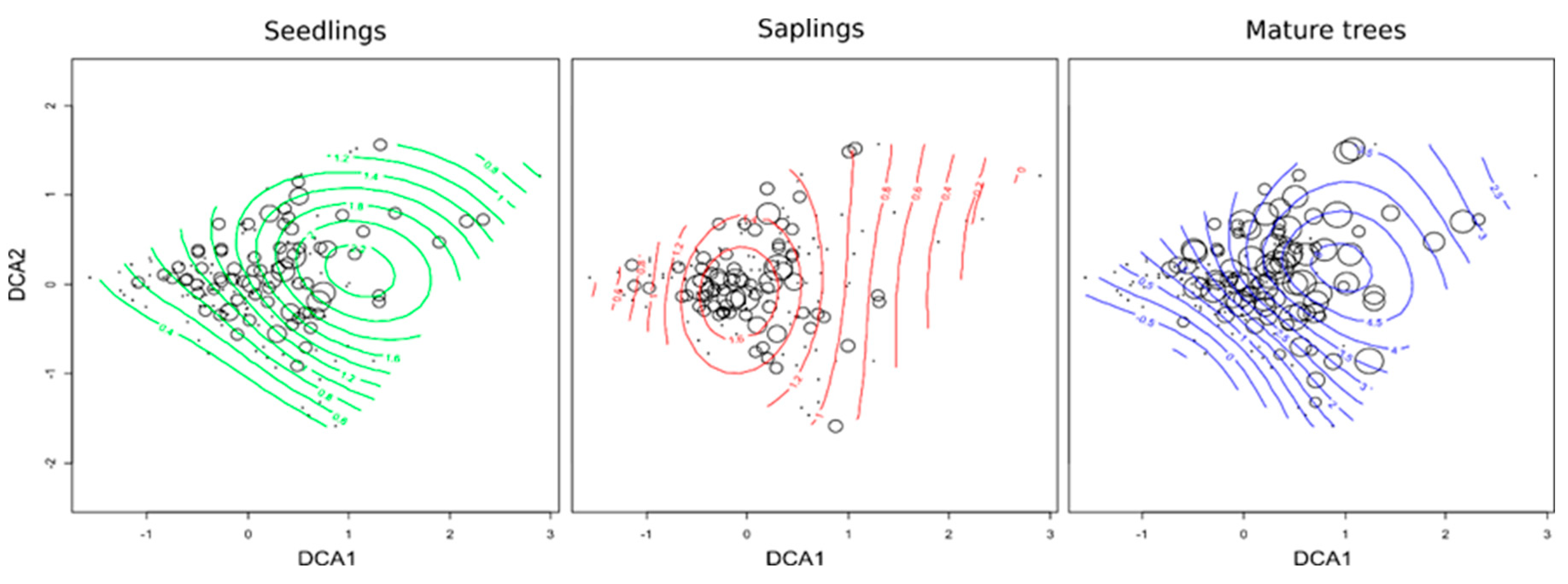
3.1. The Factors that Control the Regeneration and Abundance of Q. rubra Seedlings
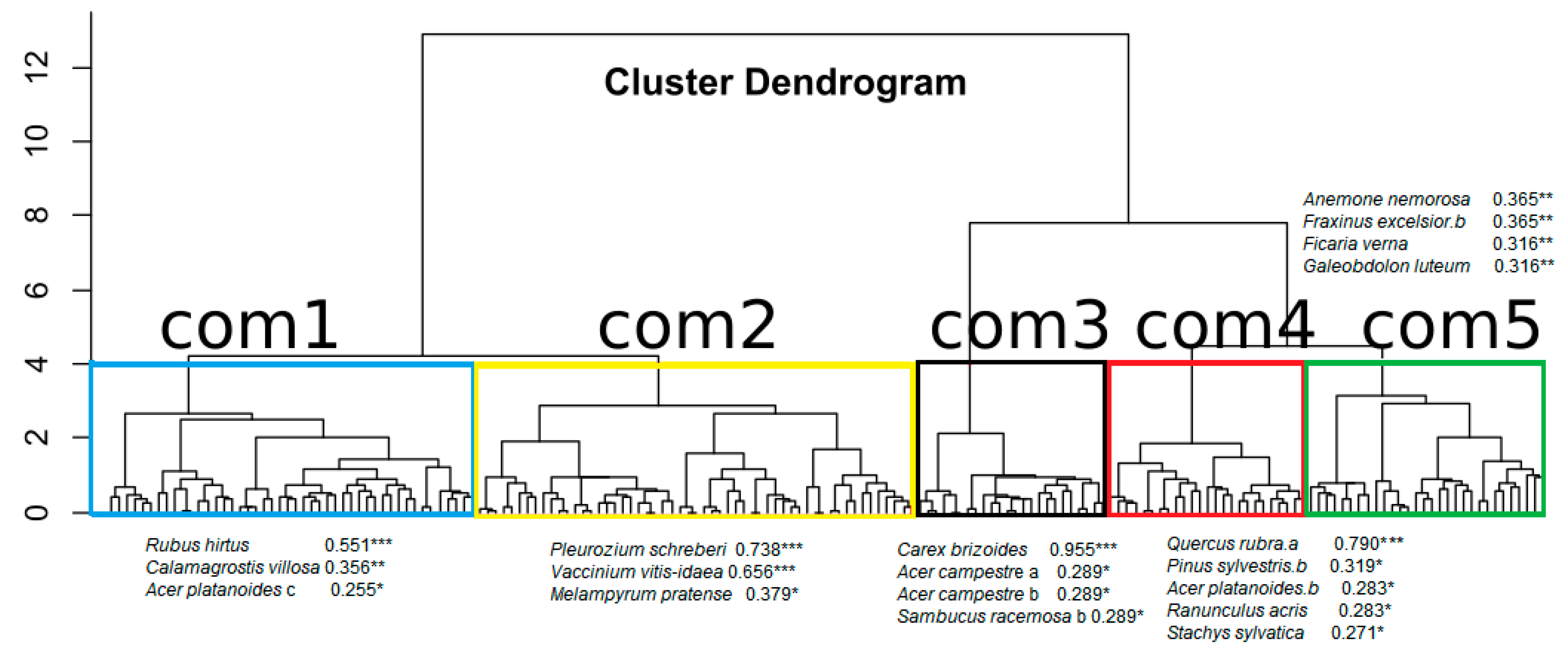
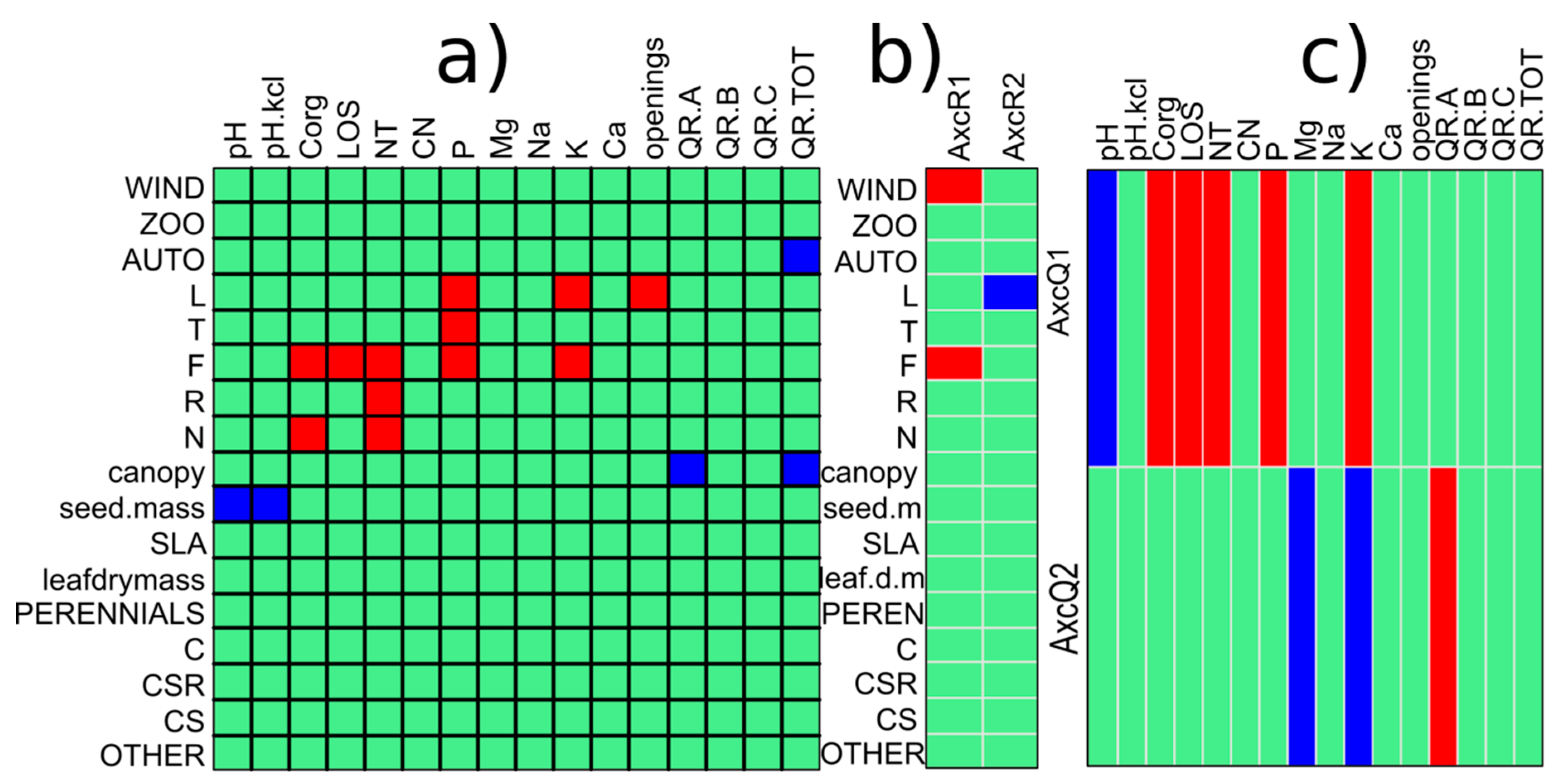
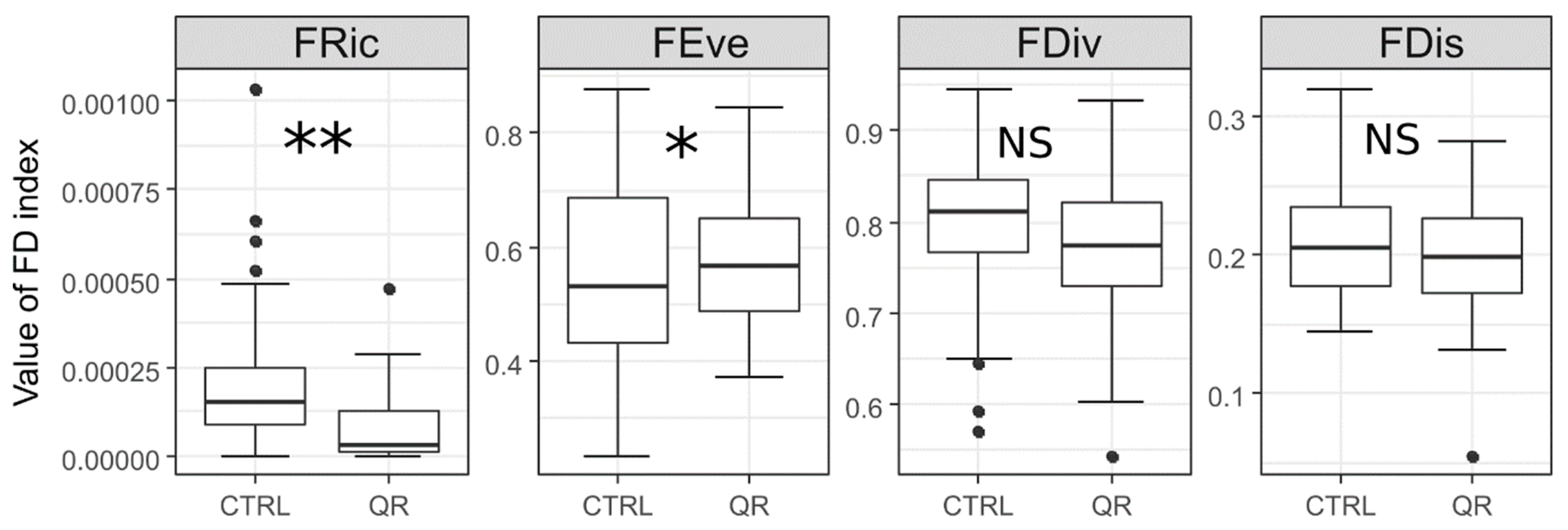
3.2. Functional Diversity of the Forest Communities That Have Q. rubra
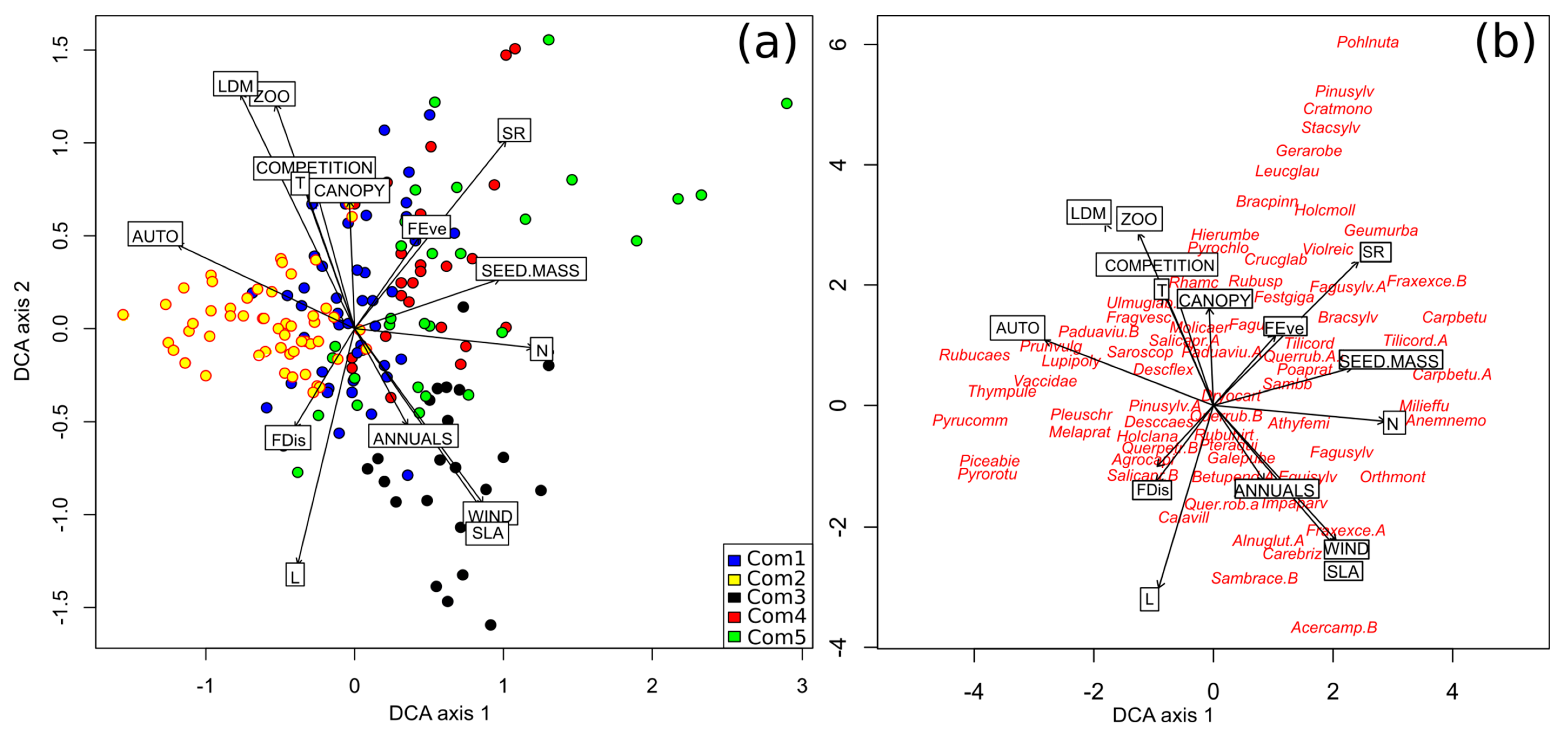
3.3. The Impact of Q. rubra on Plant Traits and the Functional Diversity of the Forest Communities
3.4. A Comparison of the Functional Diversity of the Invaded and Non-Invaded Forest Communities
4. Discussion
4.1. Influence of Q. rubra in the Overstorey on the Presence and Abundance of Seedlings
4.2. Differences among the Types of Forests That Have a Share of Q. rubra
4.3. Role of Red Oak in the Decrease of the Functional Diversity of Forest Communities
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response Variable | Type of Model | Model | AICc | Δ |
|---|---|---|---|---|
| QR | GLM (base model) | ~Mature trees * Saplings + Habitat + Forest site | 647.5 | 3.68 |
| GLM (best-fit model) | ~Mature trees * Saplings + Habitat | 644.23 | 0.0 | |
| QC | LMM (base model) | ~Mature trees * Saplings + Habitat + Forest site + (1|complex) | 4947.88 | 11.35 |
| LMM (best-fit model) | ~Mature trees + Saplings + Habitat + Forest site + (1|complex) | 4936.53 | 0.0 |
| Estimate | Std. Error | Z-Value | p-Value | |
|---|---|---|---|---|
| (Intercept) | 1.7277033 | 0.7706944 | 2.242 | 0.025 |
| Mature trees | −0.0011236 | 0.0106095 | −0.106 | NS |
| Saplings | 0.021347 | 0.0076399 | 2.794 | 0.0052 |
| Habitat (forest interior) | −1.1109173 | 0.7816578 | −1.421 | NS |
| Habitat (forest margins) | −1.4408269 | 0.8467675 | −1.702 | NS |
| Habitat (transport routes) | −1.6333783 | 0.7752345 | −2.107 | 0.0351 |
| Mature trees × saplings | 0.0008691 | 0.0005176 | 1.679 | NS |
| Estimate | Std. Error | DF | T-Value | p-Value | |
|---|---|---|---|---|---|
| (Intercept) | 26.69208 | 6.99856 | 500.61002 | 3.814 | 0.000154 |
| Mature trees | 0.45198 | 0.07334 | 525.6401 | 6.163 | <0.0001 |
| Saplings | 0.41774 | 0.05835 | 526.11664 | 7.16 | <0.0001 |
| Habitat (forest interior) | −17.3853 | 6.44975 | 511.40766 | −2.696 | 0.00726 |
| Habitat (forest margins) | −12.17581 | 7.43697 | 512.87867 | −1.637 | NS |
| Habitat (transport routes) | −18.61828 | 6.38134 | 509.44177 | −2.918 | 0.003683 |
| Forest site (deciduous) | 7.14314 | 4.40885 | 391.81312 | 1.62 | NS |
| Forest site (mixed coniferous) | 5.50261 | 3.33939 | 516.07134 | 1.648 | NS |
| Forest site (mixed deciduous) | 3.42899 | 3.78507 | 456.864 | 0.906 | NS |
| Variables | DCA1 | DCA2 | R2 | p-Value |
|---|---|---|---|---|
| FRic | −0.68392 | −0.72956 | 0.0619 | 0.005 |
| FEve | 0.65932 | 0.75186 | 0.0803 | 0.001 |
| FDiv | 0.62502 | 0.78061 | 0.0256 | NS |
| FDis | −0.60482 | −0.79636 | 0.0825 | 0.001 |
| COMPETITION | −0.32302 | 0.94639 | 0.1093 | 0.001 |
| WIND | 0.67593 | −0.73696 | 0.3022 | 0.001 |
| ZOO | −0.40045 | 0.91632 | 0.323 | 0.001 |
| AUTO | −0.93397 | 0.35736 | 0.3022 | 0.001 |
| L | −0.2882 | −0.95757 | 0.3265 | 0.001 |
| T | −0.40783 | 0.91306 | 0.1267 | 0.001 |
| F | 0.06275 | −0.99803 | 0.0509 | 0.01 |
| R | 0.7079 | 0.70631 | 0.3828 | 0.001 |
| N | 0.99632 | −0.08573 | 0.269 | 0.001 |
| CANOPY | −0.04716 | 0.99889 | 0.0878 | 0.001 |
| SEED.MASS | 0.96458 | 0.26381 | 0.191 | 0.001 |
| SLA | 0.65848 | −0.7526 | 0.3045 | 0.001 |
| Leaf Dry Mass | −0.51739 | 0.85575 | 0.406 | 0.001 |
| PERENNIALS | 0.72815 | 0.68542 | 0.0074 | NS |
| ANNUALS | 0.56488 | −0.82517 | 0.0741 | 0.003 |
| RLQ Axes | Axis 1 | Axis 2 |
|---|---|---|
| Eigenvalues | 0.375 | 0.184 |
| Covariance | 0.613 | 0.429 |
| Correlation | 0.19 | 0.208 |
| Total variance (%) | 46.105 | 22.666 |
| R/RLQ (%) | 0.806 | 0.706 |
| L/RLQ (%) | 0.218 | 0.269 |
| Q/RLQ (%) | 0.825 | 0.793 |

References
- Weber, E. Invasive Plant Species of the World: A Reference Guide to Environmental Weeds; CABI Publishing: Wallingford, UK, 2003. [Google Scholar]
- Křivanek, M.; Pyšek, P. Forestry and horticulture as pathways of plant invasions: A database of alien woody plants in the Czech Republic. In Plant Invasions: Human Perception, Ecological Impacts and Management; Tokarska-Guzik Brock, B.J.H., Brundu, G., Child, L., Daehler, C.C., Pyšek, P.P., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2008; pp. 213–218. [Google Scholar]
- Pyšek, P.; Sádlo, J.; Mandák, B. Alien flora of the Czech Republic, its composition, structure and history. In Plant Invasions: Ecological Threats and Management Solutions; Child, L.E., Brock, J.H., Brundu, G., Prach, K., Pysek, P., Wade, P.M., Williamson, M., Eds.; Edited Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 113–130. [Google Scholar]
- Zelnik, I. The presence of invasive alien plant species in different habitats: Case study from Slovenia. Acta Biol. Slovenica 2012, 55, 25–38. [Google Scholar]
- Tokarska-Guzik, B.; Dajdok, Z.; Zając, M.; Zając, A.; Urbisz, A.; Danielewicz, W.; Hołdyński, C. Rośliny Obcego Pochodzenia w Polsce ze Szczególnym Uwzględnieniem Gatunków Inwazyjnych; Generalna Dyrekcja Ochrony Środowiska: Warszawa, Poland, 2012; ISBN 978-83-62940-33-2.
- Riepšas, E.; Straigyte, L. Invasiveness and ecological effects of red oak (Quercus rubra L.) in Lithuanian forests. Balt. For. 2008, 2, 122–130. [Google Scholar]
- Vor, T. Natural regeneration of Quercus rubra L. (Red Oak) in Germany. In Biological Invasions from Ecology to Control. Neobiota 6; Nentwig, W., Bacher, S., Cock, M.J.W., Dietz, H., Gigon, A., Wittenberg, R., Eds.; TU Germany: Berlin, Germany, 2005; pp. 111–123. [Google Scholar]
- Medvecká, J.; Jarolímek, I.; Hegedüšová, K.; Škodová, I.; Bazalová, D.; Botková, K.; Šibíková, M. Forest habitat invasions—Who with whom, where and why. For. Ecol. Manag. 2018, 409, 468–478. [Google Scholar] [CrossRef]
- Oosterbaan, A.; Olsthorn, A.F.M. Control strategies for Prunus serotina and Quercus rubra as exotic tree species in the Netherlands. In Biological Invasions from Ecology to Control. Neobiota 6; Nentwig, W., Bacher, S., Cock, M.J.W., Dietz, H., Gigon, A., Wittenberg, R., Eds.; TU Germany: Berlin, Germany, 2005; pp. 177–181. [Google Scholar]
- Merceron, N.R.; De Langhe, A.; Dubois, H.; Garin, O.; Gerarts, F.; Jacquemin, F.; Balligand, B.; Otjacques, M.; Sabbe, T.; Servranckx, M.; et al. Removal of acorns of the alien oak Quercus rubra on the ground by scatter-hoarding animals in Belgian forests. Biotechnol. Agron. Soc. Environ. 2017, 21, 127–130. [Google Scholar] [CrossRef]
- Petkova, K.; Popov, E.; Tsvetkov, I. Bulgaria. In Non-Native Tree Species for European Forests: Experiences, Risks and Opportunities. FP 1403 NNEXT Country Reports, Joint Volume; Hasenauer, H., Gazda, A., Konnert, M., Lapin, K., (Frits) Mohren, G.M.J., Spiecker, H., Eds.; University of Natural Resources and Life Sciences: Vienna, Austria, 2006; pp. 40–60. [Google Scholar]
- Richardson, D.M.; Pyšek, P.; Carlton, J.T. A compendium of essential concepts and terminology in invasion ecology. In Fifty Years Invasion Ecology: The Legacy of Charles Elton; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011. [Google Scholar] [CrossRef]
- Chmura, D. Impact of alien tree species Quercus rubra L. on understorey environment and flora: A study of the Silesian Upland (Southern Poland). Pol. J. Ecol. 2013, 61, 431–442. [Google Scholar]
- Woziwoda, B.; Kopeć, D.; Witkowski, J. The negative impact of intentionally introduced Quercus rubra L. on a forest community. Acta Soc. Bot. Pol. 2014, 83, 39–49. [Google Scholar] [CrossRef]
- Gentili, R.; Ferrè, C.; Cardarelli, E.; Montagnani, C.; Bogliani, G.; Citterio, S.; Comolli, R. Comparing Negative Impacts of Prunus serotina, Quercus rubra and Robinia pseudoacacia on Native Forest Ecosystems. Forests 2019, 10, 842. [Google Scholar] [CrossRef]
- Woziwoda, B.; Potocki, M.; Sagan, J.; Zasada, M.; Tomusiak, R.; Wilczyński, S. Commercial forestry as a vector of alien tree species—The case of Quercus rubra L. introduction in Poland. Balt. For. 2014, 20, 131–141. [Google Scholar]
- Hereźniak, J. Amerykańskie drzewa i krzewy na ziemiach polskich. In Rośliny pochodzenia amerykańskiego zadomowione w Polsce; Ławrynowicz, M., Warcholińska, A.U., Eds.; Łódzkie Towarzystwo Naukowe. Szlakami Nauki: Łódź, Poland, 1992; pp. 97–150. [Google Scholar]
- Renou-Wilson, F.; Keane, M.; Farrell, E.P. Establishing oak woodland on cutaway peatlands: Effects of soil preparation and fertilization. For. Ecol. Manag. 2008, 255, 728–737. [Google Scholar] [CrossRef]
- Balandier, P.; Dupraz, C. Growth of widely spaced trees. A case study from young agroforestry plantations in France. Agrofor. Syst. 1999, 43, 151–167. [Google Scholar] [CrossRef]
- Lehmkuhler, J.W.; Felton, E.E.D.; Schmidt, D.A.; Bader, K.J.; Garrett, H.E.; Kerley, M.S. Tree protection methods during the silvopastoral-system establishment in midwestern USA: Cattle performance and tree damage. Agrofor. Syst. 2003, 59, 35–42. [Google Scholar] [CrossRef]
- Ferreiro-Dominguez, N.; Rigureiro-Rodrigez, A.; Mosquera-Losada, M.R. Response to sewage sludge fertilisation in a Quercus rubra L. silvopastoral system: Soil, plant biodiversity and tree pasture production. Agricult. Ecosys. Environ. 2011, 141, 49–57. [Google Scholar] [CrossRef]
- Garcia, D.; Banuelos, M.J.; Houle, G. Differential effects of acorn burial and litter cover on Quercus rubra recruitment AT the limit of its range in eastern North America. Can. J. Bot. 2002, 80, 1115–1120. [Google Scholar] [CrossRef]
- Nicolescu, N.V.; Vor, T.; Mason, W.L.; Bastien, J.C.; Brus, R.; Henin, J.M.; Kupka, I.; Lavnyy, V.; La Porta, N.; Mohren, F.; et al. Ecology and management of northern red oak (Quercus rubra L. syn. Q. borealis F. Michx.) in Europe: A review. Forestry 2018, 1–14. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Dyderski, M.K.; Horodecki, P.; Rawlik, K. Limited dispersal prevents Quercus rubra invasion in a 14-species common garden experiment. Divers. Distrib. 2018, 24, 403–414. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Jagodziński, A.M. Functional traits of acquisitive invasive woody species differ from conservative invasive and native species. NeoBiota 2019, 41, 91–113. [Google Scholar] [CrossRef]
- Kondracki, J. Geografia Regionalna Polski; PWN: Warszawa, Poland, 1998. [Google Scholar]
- Gilewska, S. Wyżyny Śląsko-Małopolskie. In Geomorfologia Polski; Klimaszewski, M., Ed.; PWN: Warszawa, Poland, 1972; pp. 2323–2339. [Google Scholar]
- Stupnicka, E. Geologia Regionalna; Polski. Wyd. Geol: Warszawa, Poland, 1989. [Google Scholar]
- Nowak, T.; Urbisz, A.; Kapusta, P.; Tokarska-Guzik, B. Distribution patterns and habitat preferences of mountain vascular plant species in the Silesian Uplands (Southern Poland). Pol. J. Ecol. 2011, 59, 219–234. [Google Scholar]
- Pyšek, P.; Prach, K.; Mandák, B. Invasions of alien plants into habitats of Central European landscape: An historical pattern. In Plant Invasions: Ecological Mechanisms and Human Responses; Starfinger, U., Kowarik, I., Williamson, M., Eds.; Backhuys Publishers: Leiden, The Netherlands, 1998; pp. 23–32. [Google Scholar]
- Pielech, R.; Malicki, M. Relacje między typem siedliskowym lasu a zbiorowiskiem roślinnym w warunkach górskich. Sylwan 2014, 158, 675–683. [Google Scholar] [CrossRef]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas mit den Alpen; Ulmer: Stuttgart, Germany, 2010. [Google Scholar]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; Van Groenendael, J.M.; Klimeš, L.; Klimešová, J.; et al. The LEDA Traitbase: A database of life-history traits of the Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Grime, J.P.; Hodgson, J.G.; Hunt, R. Comparative Plant Ecology: A Functional Approach to Common British Species; Castlepoint Press: Dalbeattie, UK, 2007. [Google Scholar]
- Brown, N.; Jennings, S.; Wheeler, P.; Nabe-Nielsen, J. An improved method for the rapid assessment of forest understorey light environments. J. Appl. Ecol. 2000, 37, 1044–1053. [Google Scholar] [CrossRef]
- Chmura, D.; Salachna, A.; Sierka, E. Porównanie oceny zwarcia drzewostanu za pomocą metody wizualnej i zwarciomierza. Sylwan 2016, 160, 475–481. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference. R Package Version 1.43.15. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 9 April 2020).
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2018; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 9 April 2020).
- Oksanen, J. Multivariate Analysis of Ecological Communities in R: Vegan Tutorial. 2011. Available online: http://r-forge.r-project.org/projects/vegan (accessed on 9 April 2020).
- Cáceres, M.D.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Dray, S.; Choler, P.; Doledec, S.; Peres-Neto, P.R.; Thuiller, W.; Pavoine, S.; ter Braak, C.J.F. Combining the fourth-corner and the RLQ methods for assessing trait responses to environmental variation. Ecology 2014, 95, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Doledec, S.; Chessel, D.; ter Braak, C.J.F.; Champely, S. Matching species traits to environmental variables: A new three-table ordination method. Environ. Ecol. Stat. 1996, 3, 143–166. [Google Scholar] [CrossRef]
- Villéger, S.N.; Mason, W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Kotowski, W.; Beauchard, O.; Opdekamp, W.; Meire, P.; Van Diggelen, R. Waterlogging and canopy interact to control species recruitment in floodplains. Function. Ecol. 2010, 24, 918–926. [Google Scholar] [CrossRef]
- Thonicke, K.; Billing, M.; von Bloh, W.; Sakschewski, B.; Niinemets, Ü.; Peñuelas, J.; Cornelissen, J.H.C.; Onoda, Y.; van Bodegom, P.; Schaepman, M.E.; et al. Simulating functional diversity of European natural forests along climatic gradients. J. Biogeogr. 2020. [Google Scholar] [CrossRef]
- Chmura, D. Differences in invasiveness of alien woody plants in forest communities of the Silesian Upland (Southern Poland). In The Role of Geobotany in Biodiversity Conservation; Holeksa, J., Babczyńska-Sendek, B., Wika, S., Eds.; University of Silesia: Katowice, Poland, 2009; pp. 297–302. [Google Scholar]
- Gazda, A.; Szlaga, A. Obce gatunki drzewiaste w północnym kompleksie Puszczy Niepołomickiej. Sylwan 2008, 152, 58–67. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Chmura, D.; Dylewski, Ł.; Horodecki, P.; Jagodziński, A.M.; Pietras, M.; Robakowski, P.; Woziwoda, B. Biological Flora of the British Isles: Quercus rubra. J. Ecol. 2020, 291. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Jagodziński, A.M. Drivers of invasive tree and shrub natural regeneration in temperate forests. Biol. Invasions 2018, 20, 2363–2379. [Google Scholar] [CrossRef]
- Woziwoda, B.; Krzyżanowska, A.; Dyderski, M.K.; Jagodziński, A.M.; Stefańska-Krzaczek, E. Propagule pressure, presence of roads, and microsite variability influence dispersal of introduced Quercus rubra in temperate Pinus sylvestris forest. For. Ecol. Manag. 2018, 428, 35–45. [Google Scholar] [CrossRef]
- Major, K.C.; Nosko, P.; Kuehne, C.; Campbell, D.; Bauhus, J. Regeneration dynamics of non-native northern red oak (Quercus rubra L.) populations as influenced by environmental factors: A case study in managed hardwood forests of southwestern Germany. For. Ecol. Manag. 2013, 291, 144–153. [Google Scholar] [CrossRef]
- Kuehne, C.; Nosko, P.; Horwath, T.; Bauhus, J. A comparative study of physiological and morphological seedling traits associated with shade tolerance in introduced red oak (Quercus rubra) and native hardwood tree species in southwestern Germany. Tree Physiol. 2014, 34, 184–193. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Jagodziński, A.M. Low impact of disturbance on ecological success of invasive tree and shrub species in temperate forests. Plant Ecol. 2018, 219, 1369–1380. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Jagodziński, A.M. Seedling survival of Prunus serotina Ehrh., Quercus rubra L. and Robinia pseudoacacia L. in temperate forests of Western Poland. For. Ecol. Manag. 2019, 450, 117498. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Jagodziński, A.M. Similar Impacts of Alien and Native Tree Species on Understory Light Availability in a Temperate Forest. Forests 2019, 10, 951. [Google Scholar] [CrossRef]
- Parker, G.R.; Leopold, D.J.; Eichenberger, J.K. Tree dynamics in an old-growth, deciduous forest. For. Ecol. Manag. 1985, 11, 31–57. [Google Scholar] [CrossRef]
- Crow, T.R. Population dynamics and growth patterns for a cohort of northern red oak (Quercus rubra) seedlings. Oecologia 1992, 91, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Gazda, A.; Fijała, M. Obce gatunki drzewiaste w południowym kompleksie Puszczy Niepołomickiej. Sylwan 2010, 154, 333–340. [Google Scholar] [CrossRef]
- Dajdok, Z.; Krzysztofiak, A.; Romański, M.; Śliwiński, M. Rośliny inwazyjne w Wigierskim Parku Narodowym; Wigierski Park Narodowy: Krzywe, Poland, 2007. [Google Scholar]
- Chwistek, K. Kierunki i dynamika zmian składu gatunkowego i struktury drzewostanów Ojcowskiego Parku Narodowego w latach 19902-003. Prądnik. Prace Muz. Szafera 2007, 17, 95–111. [Google Scholar]
- Szczygielski, M. Zmiany charakterystyki fitosocjologicznej borów świeżych Peucedano-Pinetum w puszczach: Piskiej i augustowskiej na przestrzeni 50 lat. Studia i Materiały CEPL 2007, 9, 153–167. [Google Scholar]
- Otręba, A.; Ferchmin, M. Obce gatunki drzew miarą przekształcenia przyrody Kampinoskiego Parku Narodowego. In Siedliska i gatunki wskaźnikowe w lasach. Studia i Materiały CEPL; Anderwald, D., Ed.; CEPL: Rogów, Poland, 2007; pp. 234–244. [Google Scholar]
- Chmura, D. Penetration and naturalisation of invasive alien plant species (neophytes) in woodlands of the Silesian Upland (southern Poland). Nat. Conserv. 2004, 60, 3–11. [Google Scholar]
- Chmura, D. Charakterystyka fitocenotyczna lasów z udziałem Quercus rubra L. na Wyżynie Śląskiej. Acta Bot. Siles. 2014, 10, 17–40. [Google Scholar]
- Zarzycki, W.; Wilczek, Z.; Woźnica, P.; Folcik, P.; Lewandowska, A.; Gancarek, M. Impact of northern red oak Quercus rubra L. on fern populations in different forest phytocoenoses. Sylwan 2015, 159, 658–665. [Google Scholar]
- Dyderski, M.K.; Jagodziński, A.M. Context-dependence of urban forest vegetation invasion level and alien species’ ecological success. Forests 2019, 10, 26. [Google Scholar] [CrossRef]
- Horodecki, P.; Jagodziński, A.M. Tree species effects on litter decomposition in pure stands on afforested post-mining sites. For. Ecol. Manag. 2017, 406, 1–11. [Google Scholar] [CrossRef]
- Hill, M.O.; Preston, C.D.; Roy, D.B. PLANTATT—Attributes of British and Irish Plants: Status, Size, Life History, Geography and Habitats; Centre for Ecology and Hydrology: Huntingdon, UK, 2004. [Google Scholar]
- Chytrý, M.; Tichý, L.; Dřevojan, P.; Sádlo, J.; Zelený, D. Ellenberg type indicator values for the Czech flora. Preslia 2018, 90, 83–103. [Google Scholar] [CrossRef]
- Chmura, D.; Sierka, E. The invasibility of deciduous forest communities after disturbance: A case study of Carex brizoides and Impatiens parviflora invasion. For. Ecol. Manag. 2007, 242, 487–495. [Google Scholar] [CrossRef]
- Dzwonko, Z.; Gawronski, S. The role of woodland fragments, soil types, and dominant species in secondary succession on the Western Carpathian foothills. Vegetatio 1994, 111, 149–160. [Google Scholar] [CrossRef]
- Olaczek, R. Kierunki degeneracji fitocenoz leśnych i metody ich badania. Phytocoenosis 1974, 3, 179–190. [Google Scholar]
- Łaska, G. The disturbance and vegetation dynamics: A review and an alternative framework. Plant Ecol. 2001, 157, 77–99. [Google Scholar] [CrossRef]






| Name | Type/Number of Attributes | Description of Variables |
|---|---|---|
| Ellenberg indicator values | Ordinal | L: light, T: temperature, F: moisture, R: soil reaction, N: nitrogen (0–9 scale) |
| Canopy (height of a plant) | Continuous/1 | Mean value [m] |
| Seed mass | Continuous/1 | 0: no seed, for >0 mean value [mg] |
| SLA (specific leaf area) | Continuous/1 | Mean value [mm2/g] |
| Leaf dry mass | Continuous/1 | Mean value [g] |
| Dispersal syndrome | Nominal/3 | WIND: anemochory, ZOO: zoochory, AUTO: autochory, barochory |
| PERENNIALS | Nominal/1 | herb perennials, woody perennials |
| Grime strategy | Nominal/4 | C: competitors, CSR: intermediate strategy, CR: competitive ruderal, OTHER: remaining strategies |
| Environmental variables | ||
| P, Mg, K, Ca, Na | Continuous | [mg/100 g] |
| pH | Ordinal | pH in (H2O, KCl) |
| Corg | Ordinal | Organic carbon |
| LOS | Percent | Loss on ignition |
| NT | Percent | Total nitrogen |
| CN | Continuous | Ration of carbon and nitrogen |
| Openings | Percent | Canopy openings were measured using a canopy scope |
| QR.A, QR.B, QR.C, QR.TOT | Percent | Percent of cover abundance Quercus rubra in each layer: A-tree, B-shrub, C-herb layer, TOT—combined cover |
| Type of Test | Statistics | p-Value | |
|---|---|---|---|
| Generalised Linear Model (Binomial Distribution) | Wald Χ2 | DF | |
| Mature trees | 2.182 | 1 | NS |
| Saplings | 25.3857 | 1 | <0.0001 |
| Habitat | 10.5820 | 3 | 0.01421 |
| Mature trees × saplings | 3.4618 | 1 | 0.0628 (NS) |
| Linear Mixed-Effects Model (Gaussian Distribution) | Wald Χ2 | DF | |
| Mature trees | 37.9835 | 1 | <0.0001 |
| Saplings | 51.2600 | 1 | <0.0001 |
| Habitat | 10.2400 | 3 | 0.01663 |
| Forest site | 3.8629 | 1 | NS |
| Spearman Rank Correlation | S statistics | rho | |
| Mature trees and seedlings | 20,222,514 | 0.21 | <0.0001 |
| Saplings and seedlings | 18,002,082 | 0.30 | <0.0002 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chmura, D. The Spread and Role of the Invasive Alien Tree Quercus rubra (L.) in Novel Forest Ecosystems in Central Europe. Forests 2020, 11, 586. https://doi.org/10.3390/f11050586
Chmura D. The Spread and Role of the Invasive Alien Tree Quercus rubra (L.) in Novel Forest Ecosystems in Central Europe. Forests. 2020; 11(5):586. https://doi.org/10.3390/f11050586
Chicago/Turabian StyleChmura, Damian. 2020. "The Spread and Role of the Invasive Alien Tree Quercus rubra (L.) in Novel Forest Ecosystems in Central Europe" Forests 11, no. 5: 586. https://doi.org/10.3390/f11050586
APA StyleChmura, D. (2020). The Spread and Role of the Invasive Alien Tree Quercus rubra (L.) in Novel Forest Ecosystems in Central Europe. Forests, 11(5), 586. https://doi.org/10.3390/f11050586