Regeneration Dynamics Following the Formation of Understory Gaps in a Slovakian Beech Virgin Forest


Abstract
1. Introduction
2. Material and Methods
2.1. Study Site
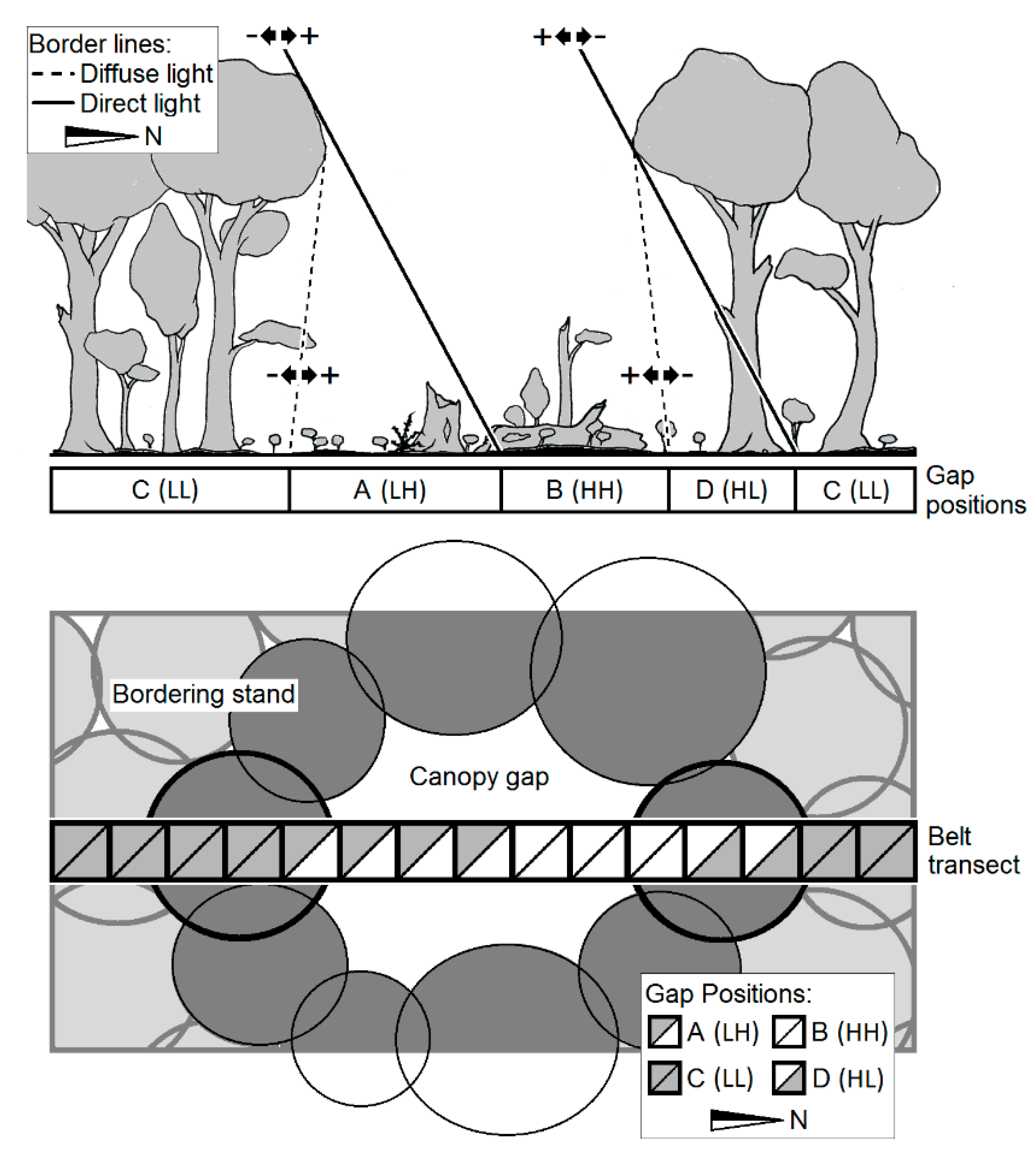
2.2. Study Design
2.3. Field Methods
2.4. Data Analysis
3. Results
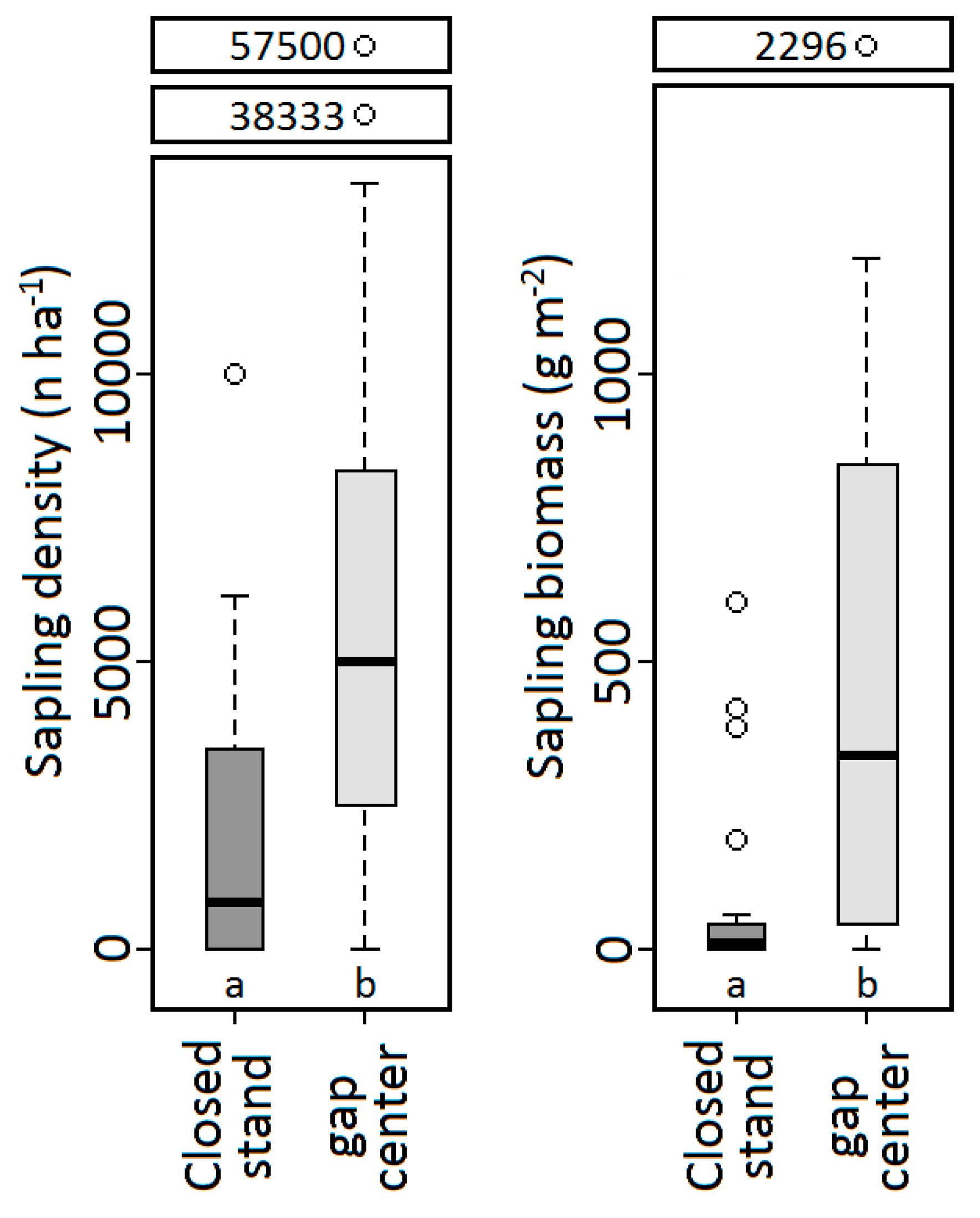
3.1. Comparison of Gaps with Closed-Canopy Conditions
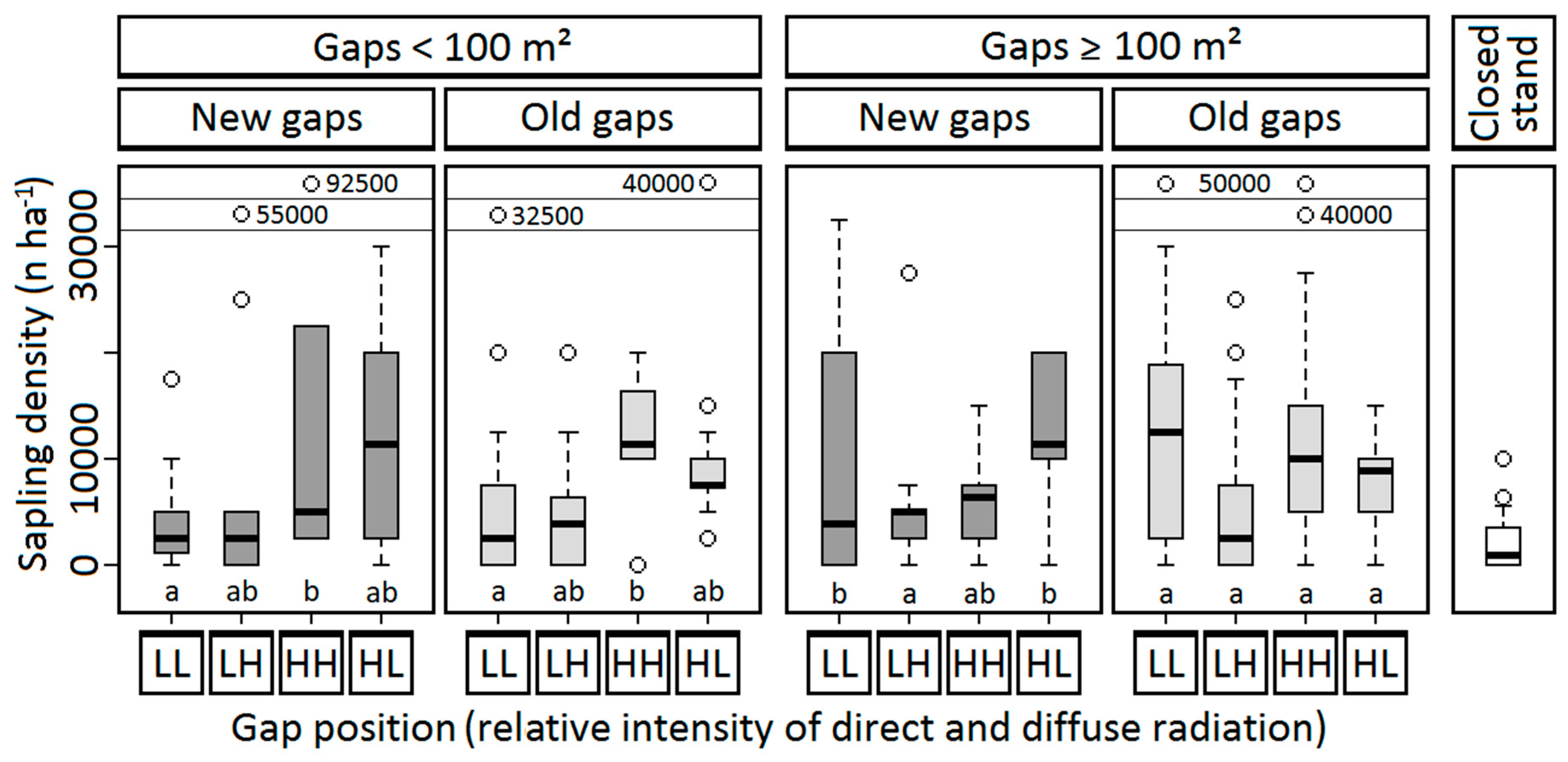
3.2. Spatial Differences in Regeneration Structure
3.2.1. Sapling Presence or Absence
3.2.2. Sapling Density
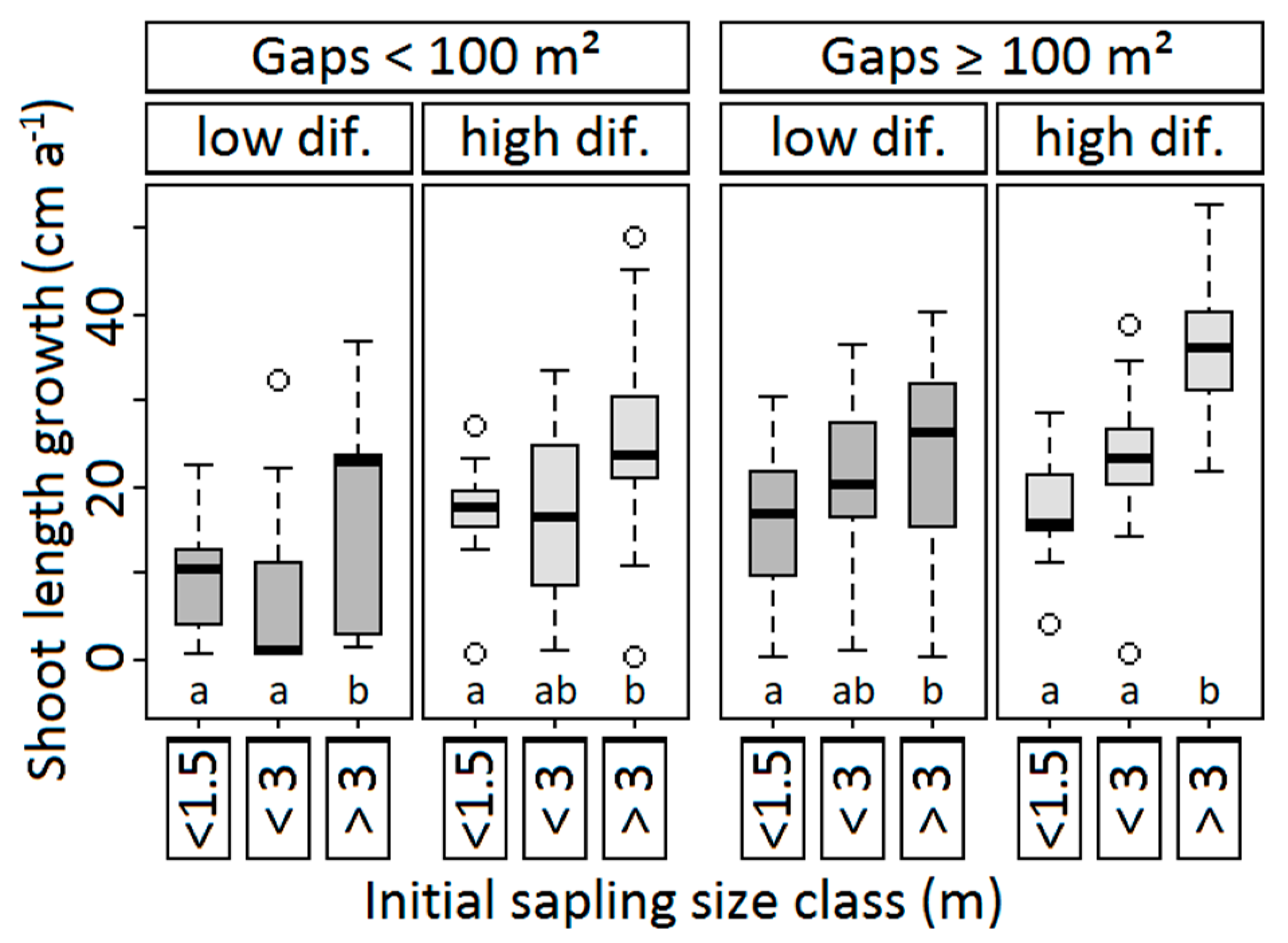
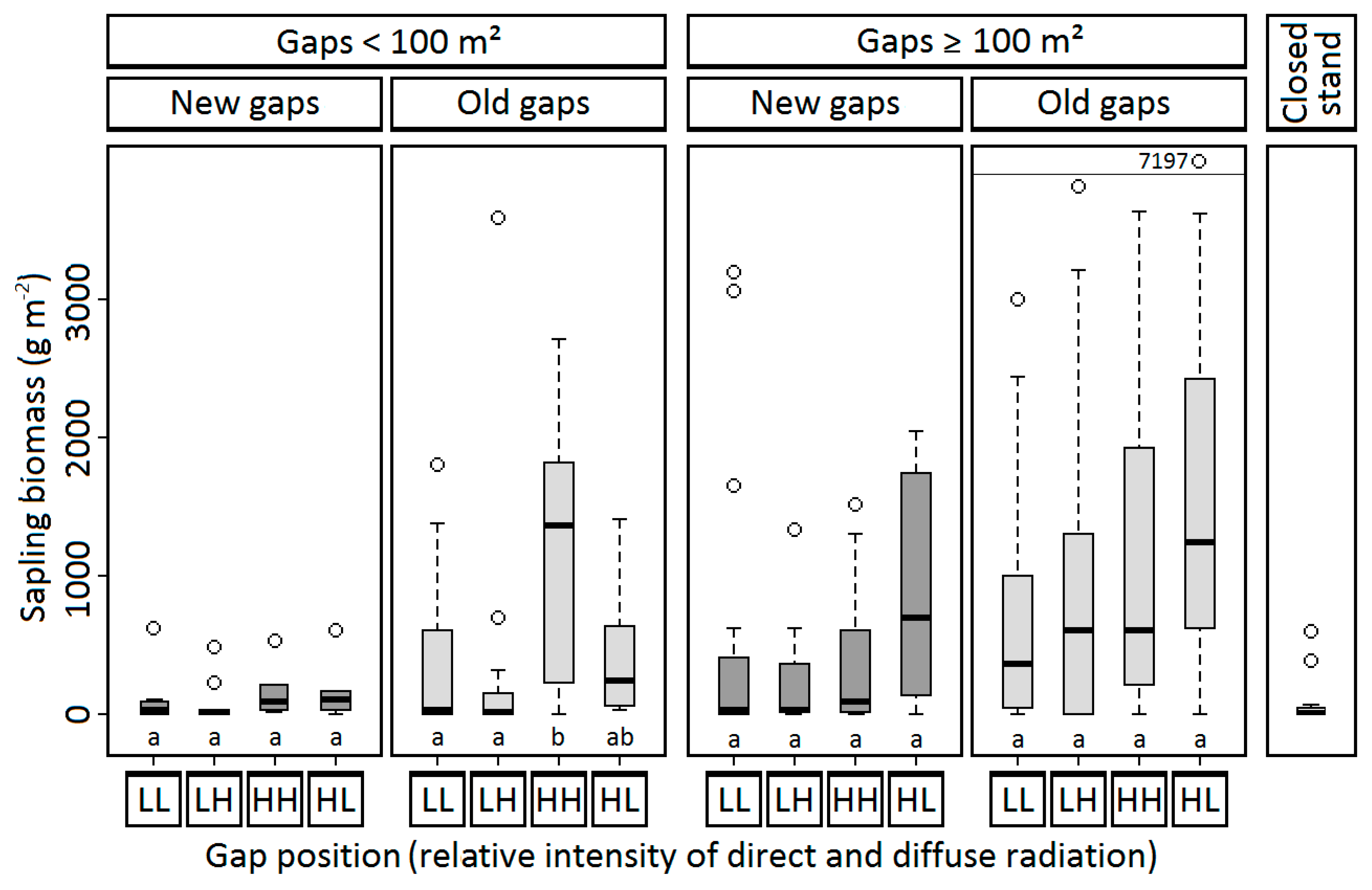
3.2.3. Shoot Length Growth and Sapling Height
3.2.4. Sapling Biomass
4. Discussion
4.1. Comparing Beech Regeneration in Gaps and Closed-Canopy Patches
4.2. The Role of Gaps for Beech Regeneration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ammer, C.; Fichtner, A.; Fischer, A.; Gossner, M.M.; Meyer, P.; Seidl, R.; Thomas, F.M.; Annighöfer, P.; Kreyling, J.; Ohse, B.; et al. Key ecological research questions for Central European forests. Basic Appl. Ecology 2018, 32, 3–25. [Google Scholar] [CrossRef]
- Ricard, J.P.; Messier, C.; Delagrange, S.; Beaudet, M. Do understory sapling respond to both light and below-ground competition? A field experiment in a north-eastern American hardwood forest and a literature review. Ann. For. Sci. 2003, 60, 749–756. [Google Scholar] [CrossRef]
- Hagemeier, M.; Leuschner, C. Leaf and crown optical properties of five early-, mid- and late-successional temperate tree species and their relation to the sapling’s light demand. Forests 2019, 10, 925. [Google Scholar] [CrossRef]
- Frelich, L.E. Forest Dynamics and Disturbance Regimes: Studies from Temperate Evergreen-deciduous Forests; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Forests. Vegetation Ecology of Central Europe; Springer Nature: Cham, Switzerland, 2017; Volume I. [Google Scholar]
- Ammer, C.; Stimm, B.; Mosandl, R. Ontogenetic variation in the relative influence of light and belowground resources on European beech seedling growth. Tree Physiol. 2008, 28, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.; Collet, C.; Madsen, P.; Nakashizuka, T.; Nyland, R.D.; Sagheb-Talebi, K. Beech regeneration research: From ecological to silvicultural aspects. For. Ecol. Manag. 2010, 259, 2172–2182. [Google Scholar] [CrossRef]
- Burschel, P.; Schmalz, J. Die Bedeutung des Lichtes für die Entwicklung junger Buchen. Allg. Forst Jagdztg. 1965, 136, 193–210. [Google Scholar]
- Szwagrzyk, J.; Szewczyk, J.; Bodziarczyk, J. Dynamics of seedling banks in beech forest: Results of a 10-year study on germination, growth and survival. For. Ecol. Manag. 2001, 141, 237–250. [Google Scholar] [CrossRef]
- Collet, C.; Chenost, C. Using competition and light estimates to predict diameter and height growth of naturally regenerated beech seedlings growing under changing canopy conditions. Forestry 2006, 79, 489–502. [Google Scholar] [CrossRef]
- Petritan, A.M.; von Lüpke, B.; Petritan, I.C. Effects of shade on growth and mortality of maple (Acer pseudoplatanus), ash (Fraxinus excelsior) and beech (Fagus sylvatica) saplings. Forestry 2007, 80, 397–412. [Google Scholar] [CrossRef]
- Petritan, A.M.; von Lüpke, B.; Petritan, I.C. Influence of light availability on growth, leaf morphology and plant architecture of beech (Fagus sylvatica L.), maple (Acer pseudoplatanus L.) and ash (Fraxinus excelsior L.) saplings. Eur. J. For. Res. 2009, 128, 61–74. [Google Scholar] [CrossRef]
- Hunter, M.L., Jr. Wildlife, Forests, and Forestry. Principles of Managing Forests for Biological Diversity; Prentice Hall: Upper Saddle River, NJ, USA, 1990. [Google Scholar]
- Leibundgut, H. Europäische Urwälder—Wegweiser Zur Naturnahen Waldwirtschaft; Paul Haupt: Bern, Switzerland, 1993. [Google Scholar]
- Korpeĺ, Š. Die Urwälder der Westkarpaten; Gustav Fischer Verlag: Stuttgart, Germany, 1995. [Google Scholar]
- Drößler, L.; von Lüpke, B. Bestandesstruktur, Verjüngung und Standortfaktoren in zwei Buchenurwald-reservaten der Slowakei. Allg. Forst Jagdztg. 2006, 178, 121–135. [Google Scholar]
- Nagel, T.A.; Svoboda, M.; Rugani, T.; Diaci, J. Gap regeneration and replacement patterns in an old-growth Fagus–Abies forest of Bosnia–Herzegovina. Plant Ecol. 2010, 208, 307–318. [Google Scholar] [CrossRef]
- Motta, R.; Berretti, R.; Castagneri, D.; Dukić, V.; Garbarino, M.; Govedar, Z.; Lingua, E.; Maunaga, Z.; Meloni, F. Toward a definition of the range of variability of central European mixed Fagus–Abies–Picea forests: The nearly steady-state forest of Lom (Bosnia and Herzegovina). Can. J. For. Res. 2011, 41, 1871–1884. [Google Scholar] [CrossRef]
- Petritan, A.M.; Biris, I.A.; Merce, O.; Turcu, D.O.; Petritan, I.C. Structure and diversity of a natural temperate sessile oak (Quercus petraea L.)–European Beech (Fagus sylvatica L.) forest. For. Ecol. Manag. 2012, 280, 140–149. [Google Scholar] [CrossRef]
- Hobi, M.L.; Commarmot, B.; Bugmann, H. Pattern and process in the largest primeval beech forest of Europe (Ukrainian Carpathians). J. Veg. Sci. 2015, 26, 323–336. [Google Scholar] [CrossRef]
- Peters, R. Beech forests. Geobotany, 24; Springer: Berlin/Heidelberg, Germany, 1997; 170 p. [Google Scholar]
- Schelhaas, M.J.; Nabuurs, G.J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Tabaku, V.; Meyer, P. Lückenmuster albanischer und mitteleuropäischer Buchenwälder unterschiedlicher Nutzungsintensität. Forstarchiv 1999, 70, 87–97. [Google Scholar]
- Drößler, L.; von Lüpke, B. Canopy gaps in two virgin beech forest reserves in Slovakia. J. For. Sci. 2005, 51, 446–457. [Google Scholar] [CrossRef]
- Zeibig, A.; Diaci, J.; Wagner, S. Gap disturbance patterns of a Fagus sylvatica virgin forest remnant in the mountain vegetation belt of Slovenia. For. Snow Landsc. Res. 2005, 79, 69–80. [Google Scholar]
- Splechtna, B.E.; Gratzer, G. Natural disturbances in Central European forests: Approaches and preliminary results from Rothwald, Austria. For. Snow Landsc. Res. 2005, 79, 57–67. [Google Scholar]
- Nagel, T.A.; Svoboda, M. Gap disturbance regime in an old-growth Fagus-Abies forest in the Dinaric Mountains, Bosnia-Herzegovina. Can. J. For. Res. 2008, 38, 2728–2737. [Google Scholar] [CrossRef]
- Kenderes, K.; Král, K.; Vrška, T.; Standovár, T. Natural gap dynamics in a Central European mixed beech-spruce-fir old-growth forest. Ecoscience 2009, 16, 39–47. [Google Scholar] [CrossRef]
- Kucbel, S.; Jaloviar, P.; Saniga, M.; Vencurik, J.; Klimaš, V. Canopy gaps in an old-growth fir-beech forest remnant of Western Carpathians. Eur. J. For. Res. 2010, 129, 249–259. [Google Scholar] [CrossRef]
- Bottero, A.; Garbarino, M.; Dukic, V.; Govedar, Z.; Lingua, E.; Nagel, T.A.; Motta, R. Gap-phase dynamics in the old-growth forest of Lom (Bosnia-Herzegovina). Silva Fennica 2011, 45, 875–887. [Google Scholar] [CrossRef]
- Petritan, A.M.; Nuske, R.S.; Petritan, I.C.; Tudose, N.C. Gap disturbance patterns in an old-growth sessile oak (Quercus petraea L.)–European beech (Fagus sylvatica L.) forest remnant in the Carpathian Mountains, Romania. For. Ecol. Manag. 2013, 308, 67–75. [Google Scholar] [CrossRef]
- Hobi, M.L.; Ginzler, C.; Commarmot, B.; Bugmann, H. Gap pattern of the largest primeval beech forest of Europe revealed by remote sensing. Ecosphere 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Feldmann, E.; Drößler, L.; Hauck, M.; Kucbel, S.; Pichler, V.; Leuschner, C. Canopy gap dynamics and tree understory release in a virgin beech forest, Slovakian Carpathians. For. Ecol. Manag. 2018, 415, 38–46. [Google Scholar] [CrossRef]
- Coates, K.D.; Burton, P.J. A gap-based approach for development of silvicultural systems to address ecosystem management objectives. For. Ecol. Manag. 1997, 99, 337–354. [Google Scholar] [CrossRef]
- Wiedemann, E. Über den künstlichen gruppenweisen Voranbau von Tanne und Buche. Allg. Forst Jagdztg. 1927, 103, 433–452. [Google Scholar]
- Canham, C.D.; Denslow, J.S.; Platt, W.J.; Runkle, J.R.; Spies, T.A.; White, P.S. Light regimes beneath closed canopies and tree-fall gaps in temperate and tropical forests. Can. J. For. Res. 1990, 20, 620–631. [Google Scholar] [CrossRef]
- Wagner, S. Ökologische Untersuchungen zur Initialphase der Naturverjüngung in Eschen-Buchen-Mischbeständen; Schriften aus der Forstlichen Fakultät der Universität Göttingen und der Niedersächsischen Forstlichen Versuchsanstalt 129: Göttingen, Germany, 1999. [Google Scholar]
- Diaci, J. Regeneration dynamics in a Norway spruce plantation on a silver fir-beech forest site in the Slovenian Alps. For. Ecol. Manag. 2002, 161, 27–38. [Google Scholar] [CrossRef]
- Madsen, P.; Hahn, K. Natural regeneration in a beech-dominated forest managed by close-to-nature principles—A gap cutting based experiment. Can. J. For. Res. 2008, 38, 1716–1729. [Google Scholar] [CrossRef]
- Vilhar, U.; Roženbergar, D.; Simončič, P.; Diaci, J. Variation in irradiance, soil features and regeneration patterns in experimental forest canopy gaps. Ann. For. Sci. 2015, 72, 253–266. [Google Scholar] [CrossRef]
- Runkle, J.R. Synchrony of regeneration, gaps, and latitudinal differences in tree species diversity. Ecology 1989, 70, 546–547. [Google Scholar] [CrossRef]
- Whitmore, T.C. Canopy gaps and the two major groups of forest trees. Ecology 1989, 70, 536–538. [Google Scholar] [CrossRef]
- Diaci, J.; Rozenbergar, D.; Boncina, A. Interactions of light and regeneration in Slovenian Dinaric Alps: Patterns in virgin and managed forests. Natural Forests in the Temperate Zone of Europe—Values and Utilisation, 2005, Proceedings of the Conference Birmensdorf, Swiss Federal Research Institute WSL.; Rakhiv, Carpathian Biosphere Reserve, 13–17 October 2003, Mukachevo, Ukraine. Commarmot, B., Hamor, F.D., Eds.; 2005, pp. 154–160. Available online: https://www.academia.edu/13029495/Interactions_of_light_and_regeneration_in_Slovenian_Dinaric_Alps_patterns_in_virgin_and_managed_forests (accessed on 31 December 2005).
- Rozenbergar, D.; Mikac, S.; Anić, I.; Diaci, J. Gap regeneration patterns in relationship to light heterogeneity in two old-growth beech–fir forest reserves in South East Europe. Forestry 2007, 80, 431–443. [Google Scholar]
- Kenderes, K.; Mihók, B.; Standovár, T. Thirty years of gap dynamics in a Central European beech forest reserve. Forestry 2008, 81, 111–123. [Google Scholar] [CrossRef]
- Kucbel, S.; Saniga, M.; Jaloviar, P.; Vencurik, J. Stand structure and temporal variability in old-growth beech-dominated forests of the northwestern Carpathians: A 40-years perspective. For. Ecol. Manag. 2012, 264, 125–133. [Google Scholar] [CrossRef]
- Drößler, L.; Meyer, P. Waldentwicklungsphasen in zwei Buchen-Urwaldreservaten in der Slowakei. Forstarchiv 2006, 77, 155–161. [Google Scholar]
- Feldmann, E.; Glatthorn, J.; Hauck, M.; Leuschner, C. A novel empirical approach for determining the extension of forest development stages in temperate old-growth forests. Eur. J. For. Res. 2018, 137, 321–335. [Google Scholar] [CrossRef]
- Brokaw, N.V.L. The definition of treefall gap and its effect on measures of forest dynamics. Biotropica 1982, 14, 158–160. [Google Scholar] [CrossRef]
- Glatthorn, J.; Feldmann, E.; Pichler, V.; Hauck, M.; Leuschner, C. Biomass stock and productivity of primeval and production beech forests: Greater canopy structural diversity promotes productivity. Ecosystems 2018, 21, 704–722. [Google Scholar] [CrossRef]
- Annighöfer, P.; Ameztegui, A.; Ammer, C.; Balandier, P.; Bartsch, N.; Bolte, A.; Coll, L.; Collet, C.; Ewald, J.; Frischbier, N.; et al. Species-specific and generic biomass equations for seedlings and saplings of European tree species. Eur. J. For. Res. 2016, 135, 313–329. [Google Scholar] [CrossRef]
- Čater, M.; Diaci, J.; Roženbergar, D. Gap size and position influence variable response of Fagus sylvatica L. and Abies alba Mill. For. Ecol. Manag. 2014, 325, 128–135. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 25 March 2020).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Turner, I.M. The ecology of Trees in the Tropical Rain Forest; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Canham, C.D. An index for understory light levels in and around canopy gaps. Ecology 1988, 69, 1634–1638. [Google Scholar] [CrossRef]
- Lieberman, M.; Lieberman, D.; Peralta, R. Forests are not just Swiss cheese—Canopy stereogeometry of non-gaps in tropical forests. Ecology 1989, 70, 550–552. [Google Scholar] [CrossRef]
- Ammer, C. Konkurrenz um Licht—Zur Entwicklung der Naturverjüngung im Bergmischwald. Forstliche Forschungsberichte München 1996, 158, 198. [Google Scholar]
- Ammer, C.; Mosandl, R.; El Kateb, H. Direct seeding of beech (Fagus sylvatica L.) in Norway spruce (Picea abies [L.] Karst.) stands—Effects of canopy density and fine root biomass on seed germination. For. Ecol. Manag. 2002, 159, 59–72. [Google Scholar] [CrossRef]
- Minotta, G.; Pinzauti, S. Effects of light and soil fertility on growth, leaf chlorophyll content and nutrient use efficiency of beech (Fagus sylvatica L.) seedlings. For. Ecol. Manag. 1996, 86, 61–71. [Google Scholar] [CrossRef]
- Collet, C.; Lanter, O.; Pardos, M. Effects of canopy opening on height and diameter growth in naturally regenerated beech seedlings. Ann. For. Sci. 2001, 58, 127–134. [Google Scholar] [CrossRef]
- Wada, N.; Ribbens, E. Japanese maple (Acer palmatum var. matsumurae, Aceraceae) recruitment patterns: Seeds, seedlings, and saplings in relation to conspecific adult neighbors. Am. J. Bot. 1997, 84, 1294–1300. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M. Natural Regeneration of Broadleaved Tree Species in Southern Sweden; Acta Universitatis Agriculturae Sueciae; Silvestria: Alnarp, Sweden, 2001; Volume 196. [Google Scholar]
- Mihók, B.; Gálhidy, L.; Kelemen, K.; Standovár, T. Study of gap-phase regeneration in a managed beech forest: Relations between tree regeneration and light, substrate features and cover of ground vegetation. Acta Silv. Lign. Hung 2005, 1, 25–38. [Google Scholar]
- Mountford, E.P.; Savill, P.S.; Bebber, D.P. Patterns of regeneration and ground vegetation associated with canopy gaps in a managed beechwood in southern England. Forestry 2006, 79, 389–408. [Google Scholar] [CrossRef]
- Ammer, C. Response of Fagus sylvatica (L.) seedlings to root competition by overstorey Picea abies (L.) Karst. Scand. J. For. Res. 2002, 17, 408–416. [Google Scholar] [CrossRef]
- Bauhus, J. Rooting patterns of old-growth forests: Is aboveground structural and functional diversity mirrored belowground? In Old-Growth Forests; Springer: Berlin/Heidelberg, Germany, 2009; pp. 211–229. [Google Scholar]
- Petritan, I.C.; von Lüpke, B.; Petritan, A.M. Effects of root trenching of overstorey Norway spruce (Piceaabies) on growth and biomass of underplanted beech (Fagus sylvatica) and Douglas fir (Pseudotsuga menziesii) saplings. Eur. J. For. Res. 2011, 130, 813–828. [Google Scholar] [CrossRef]
- Bartsch, N.; Röhrig, E. Waldökologie; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Anderson, M.C. Studies of the woodland light climate: I. The photographic computation of light conditions. J. Ecol. 1964, 52, 27–41. [Google Scholar] [CrossRef]
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK, 1977. [Google Scholar]
- Scharenbroch, B.C.; Bockheim, J.G. The effects of gap disturbance on nitrogen cycling and retention in late-successional northern hardwood–hemlock forests. Biogeochemistry 2008, 87, 231–245. [Google Scholar] [CrossRef]
- Röhrig, E.; Bartsch, N.; von Lüpke, B. Waldbau auf ökologischer Grundlage; Ulmer: Stuttgart, Germany, 2006. [Google Scholar]
- Annighöfer, P. Stress relief through gap creation? Growth response of a shade tolerant species (Fagus sylvatica L.) to a changed light environment. For. Ecol. Manag. 2018, 415, 139–147. [Google Scholar] [CrossRef]
- Bílek, L.; Remes, J.; Podrazsky, V.; Rozenbergar, D.; Diaci, J.; Zahradník, D. Gap regeneration in near-natural European beech forest stands in Central Bohemia-the role of heterogeneity and micro-habitat factors. Dendrobiology 2014, 71, 59–71. [Google Scholar] [CrossRef]
- Stiers, M.; Willim, K.; Seidel, D.; Ammer, C.; Kabal, M.; Stillhard, J.; Annighöfer, P. Analyzing spatial distribution patterns of european beech (Fagus sylvatica L.) Regeneration in dependence of canopy openings. Forests 2019, 10, 637. [Google Scholar] [CrossRef]
- Knapp, S.P.; Webster, C.R.; Kern, C.C. The composition and height of saplings capturing silvicultural gaps at two long-term experiments in managed northern hardwood forests. Forests 2019, 10, 855. [Google Scholar] [CrossRef]
- Einhorn, K.S.; Rosenqvist, E.; Leverenz, J.W. Photoinhibition in seedlings of Fraxinus and Fagus under natural light conditions: Implications for forest regeneration. Oecologia 2004, 140, 241–251. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Pearcy, R.W. The importance of sunflecks for forest understory plants. Bioscience 1991, 41, 760–766. [Google Scholar] [CrossRef]
- Rees, U. Lichtflecken und Ihre Bedeutung Für Die CO2-Aufnahme Junger Buchen (Fagus sylvatica L.) im Unterwuchs Eines Buchenwaldes. Ph.D. Thesis, University Goettingen, Göttingen, Germany, 1999; 155 p. [Google Scholar]
- Rozenbergar, D.; Diaci, J. Architecture of Fagus sylvatica regeneration improves over time in mixed old-growth and managed forests. For. Ecol. Manag. 2014, 318, 334–340. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gap Position | Direct Radiation | Diffuse Radiation | Symbol |
|---|---|---|---|
| A | low | high | (LH) |
| B | high | high | (HH) |
| C | low | low | (LL) |
| D | high | low | (HL) |
| Response Variables | Influencing Factors | |||||
|---|---|---|---|---|---|---|
| High Dir. | High Diff. | Gap Size | Gap Age | Sapling Height | Comp. Intensity | |
| Sapling presence | 0.001 | 0.605 | 0.212 | 0.889 | — | — |
| Sapling density | 0.031 | 0.382 | 0.077 | 0.860 | — | — |
| Shoot length growth | 0.971 | <0.001 | <0.001 | — | <0.001 | 0.472 |
| Sapling biomass | 0.069 | 0.634 | <0.001 | 0.004 | — | — |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feldmann, E.; Glatthorn, J.; Ammer, C.; Leuschner, C. Regeneration Dynamics Following the Formation of Understory Gaps in a Slovakian Beech Virgin Forest. Forests 2020, 11, 585. https://doi.org/10.3390/f11050585
Feldmann E, Glatthorn J, Ammer C, Leuschner C. Regeneration Dynamics Following the Formation of Understory Gaps in a Slovakian Beech Virgin Forest. Forests. 2020; 11(5):585. https://doi.org/10.3390/f11050585
Chicago/Turabian StyleFeldmann, Eike, Jonas Glatthorn, Christian Ammer, and Christoph Leuschner. 2020. "Regeneration Dynamics Following the Formation of Understory Gaps in a Slovakian Beech Virgin Forest" Forests 11, no. 5: 585. https://doi.org/10.3390/f11050585
APA StyleFeldmann, E., Glatthorn, J., Ammer, C., & Leuschner, C. (2020). Regeneration Dynamics Following the Formation of Understory Gaps in a Slovakian Beech Virgin Forest. Forests, 11(5), 585. https://doi.org/10.3390/f11050585