Applying LiDAR to Quantify the Plant Area Index Along a Successional Gradient in a Tropical Forest of Thailand



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Datasets
2.3. Forest Age Datasets
2.4. LiDAR Data Acquisition
2.5. Overall Methodology
2.5.1. LiDAR Data and Topographic Factors Extraction
2.5.2. Estimation of LiDAR-Based Plant Area Index
2.5.3. Statistical Analyses
3. Results
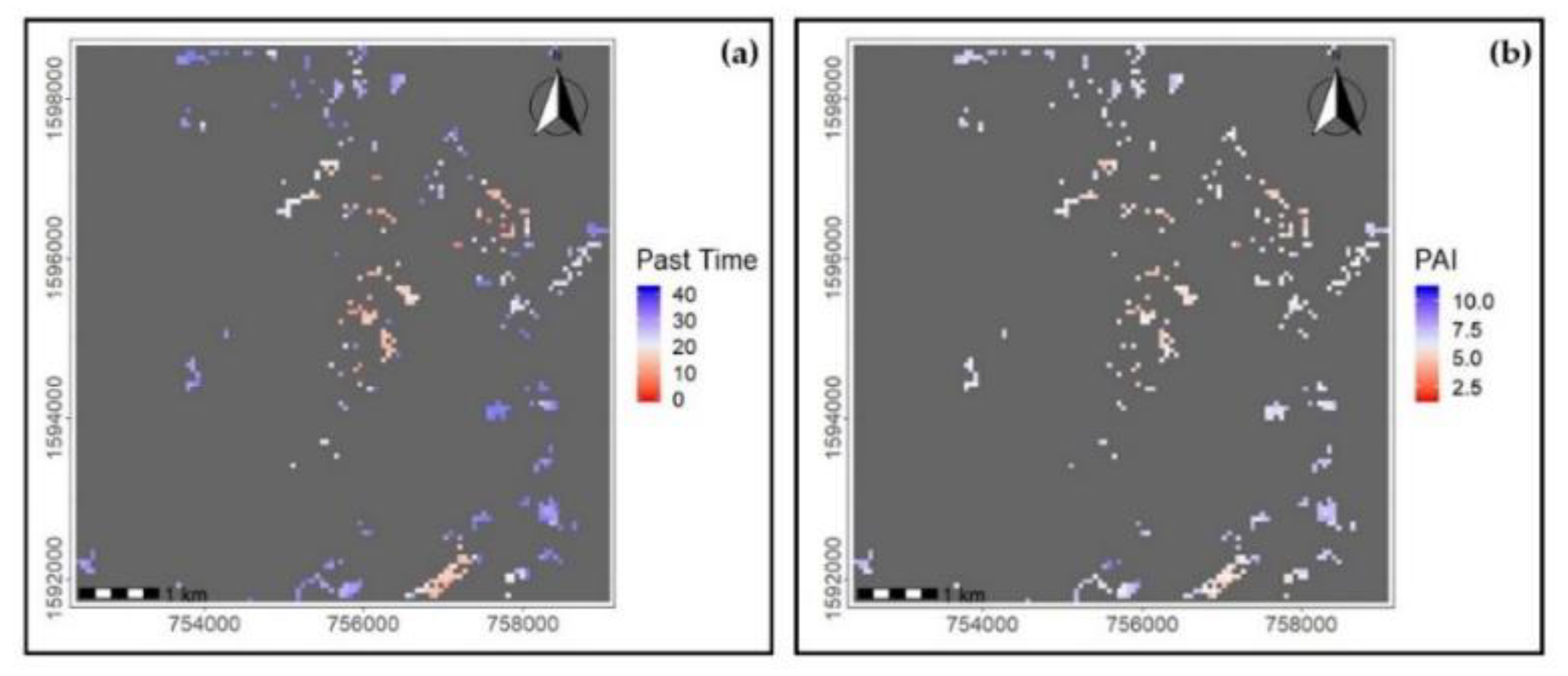
3.1. Forest Successional Structure and Spatial PAI Variability
3.2. PAI Recovery Analysis
4. Discussion
4.1. Spatial PAI Variability
4.2. Long-Term PAI Accumulation Through Succession
4.3. Topographic Factors Influence the Variation in the PAI Increase
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Estoque, R.C.; Ooba, M.; Avitabile, V.; Hijioka, Y.; Dasgupta, R.; Togawa, T.; Murayama, Y. The future of Southeast Asia’s forests. Nat. Commun. 2019, 10, 1829. [Google Scholar] [CrossRef] [PubMed]
- Stibig, H.-J.; Achard, F.; Carboni, S.; Raši, R.; Miettinen, J. Change in tropical forest cover of Southeast Asia from 1990 to 2010. Biogeosciences 2014, 11, 247–258. [Google Scholar] [CrossRef]
- Chazdon, R.L. Change and determinism in tropical forest succession. In Tropical Forest Community Ecology; Wiley-Blackwell: Oxford, UK, 2008; pp. 384–408. [Google Scholar]
- Lu, D.; Mausel, P.; Brondízio, E.; Moran, E.F. Classification of successional forest stages in the Brazilian Amazon basin. For. Ecol. Manag. 2003, 181, 301–312. [Google Scholar] [CrossRef]
- Pena-Claros, M. Changes in Forest Structure and Species Composition during Secondary Forest Succession in the Bolivian Amazon. BIOTROPICA 2003, 35, 450–461. [Google Scholar] [CrossRef]
- Brown, S.; Lugo, A.E. Tropical secondary forests. J. Trop. Ecol. 1990, 6, 1–32. [Google Scholar] [CrossRef]
- Chanthorn, W.; Ratanapongsai, Y.; Brockelman, W.Y.; Allen, M.A.; Favier, C.; Dubois, M.A. Viewing tropical forest succession as a three-dimensional dynamical system. Theor. Ecol. 2015, 9, 163–172. [Google Scholar] [CrossRef]
- Asner, G.P.; Scurlock, J.M.O.; Hicke, J.A. Global synthesis of leaf area index observations: Implications for ecological and remote sensing studies. Glob. Ecol. Biogeogr. 2003, 12, 191–205. [Google Scholar] [CrossRef]
- Chen, J.M.; Cihlar, J. Retrieving Leaf Area Index of boreal conifer forests using Landsat TM images. Remote Sens. Environ. 1996, 55, 153–162. [Google Scholar] [CrossRef]
- Zheng, G.; Moskal, L.M. Retrieving Leaf Area Index (LAI) Using Remote Sensing: Theories, Methods and Sensors. Sensors 2009, 9, 2719–2745. [Google Scholar] [CrossRef]
- Gupta, R.; Sharma, L.K. The process-based forest growth model 3-PG for use in forest management: A review. Ecol. Model. 2019, 397, 55–73. [Google Scholar] [CrossRef]
- Zheng, D.; Wang, Y.; Shao, Y.; Wang, L. The Vegetation Dynamics and Climate Change Responses by Leaf Area Index in the Mu Us Desert. Sustainability 2019, 11, 3151. [Google Scholar] [CrossRef]
- Olivas, P.C.; Oberbauer, S.F.; Clark, D.B.; Clark, D.; Ryan, M.G.; O’Brien, J.J.; Ordoñez, H. Comparison of direct and indirect methods for assessing leaf area index across a tropical rain forest landscape. Agric. For. Meteorol. 2013, 177, 110–116. [Google Scholar] [CrossRef]
- Bréda, N. Ground-based measurements of leaf area index: A review of methods, instruments and current controversies. J. Exp. Bot. 2003, 54, 2403–2417. [Google Scholar] [CrossRef]
- Jonckheere, I.; Fleck, S.; Nackaerts, K.; Muys, B.; Coppin, P.; Weiss, M. Review of methods for in situ leaf area index determination Part I. Theories, sensors and hemispherical photography. Agric. For. Meteorol. 2004, 121, 19–35. [Google Scholar] [CrossRef]
- Zhu, X.; Skidmore, A.K.; Wang, T.; Liu, J.; Darvishzadeh, R.; Shi, Y.; Premier, J.; Heurich, M. Improving leaf area index (LAI) estimation by correcting for clumping and woody effects using terrestrial laser scanning. Agric. For. Meteorol. 2018, 263, 276–286. [Google Scholar] [CrossRef]
- Blinn, C.E.; House, M.N.; Wynne, R.; Thomas, V.; Fox, T.R.; Sumnall, M.J. Landsat 8 Based Leaf Area Index Estimation in Loblolly Pine Plantations. Forests 2019, 10, 222. [Google Scholar] [CrossRef]
- Davi, H.; Soudani, K.; Deckx, T.; Dufrene, E.; Le Dantec, V.; François, C. Estimation of forest leaf area index from SPOT imagery using NDVI distribution over forest stands. Int. J. Remote Sens. 2006, 27, 885–902. [Google Scholar] [CrossRef]
- She-Zhou, L.; Wang, C.; Gui-Bin, Z.; Xiao-Huan, X.; Gui-Cai, L. Forest Leaf Area Index (LAI) Estimation Using Airborne Discrete-Return Lidar Data. Chin. J. Geophys. 2013, 56, 233–242. [Google Scholar] [CrossRef]
- Van Ewijk, K.Y.; Treitz, P.; Scott, N.A. Characterizing Forest Succession in Central Ontario using Lidar-derived Indices. Photogramm. Eng. Remote Sens. 2011, 77, 261–269. [Google Scholar] [CrossRef]
- Falkowski, M.J.; Evans, J.; Martinuzzi, S.; Gessler, P.E.; Hudak, A.T. Characterizing forest succession with lidar data: An evaluation for the Inland Northwest, USA. Remote Sens. Environ. 2009, 113, 946–956. [Google Scholar] [CrossRef]
- Tang, H.; Dubayah, R.; Swantaran, A.; Hofton, M.A.; Sheldon, S.; Clark, D.B.; Blair, B. Retrieval of vertical LAI profiles over tropical rain forests using waveform lidar at La Selva, Costa Rica. Remote Sens. Environ. 2012, 124, 242–250. [Google Scholar] [CrossRef]
- Jensen, J.L.; Humes, K.S.; Vierling, L.A.; Hudak, A.T. Discrete return lidar-based prediction of leaf area index in two conifer forests. Remote Sens. Environ. 2008, 112, 3947–3957. [Google Scholar] [CrossRef]
- Morsdorf, F.; Kötz, B.; Meier, E.; Itten, K.; Allgöwer, B. Estimation of LAI and fractional cover from small footprint airborne laser scanning data based on gap fraction. Remote Sens. Environ. 2006, 104, 50–61. [Google Scholar] [CrossRef]
- Solberg, S.; Brunner, A.; Hanssen, K.H.; Lange, H.; Næsset, E.; Rautiainen, M.; Stenberg, P. Mapping LAI in a Norway spruce forest using airborne laser scanning. Remote Sens. Environ. 2009, 113, 2317–2327. [Google Scholar] [CrossRef]
- Kucharik, C.J.; Norman, J.M.; Gower, S.T. Measurements of branch area and adjusting leaf area index indirect measurements. Agric. For. Meteorol. 1998, 91, 69–88. [Google Scholar] [CrossRef]
- Vincent, G.; Antin, C.; Laurans, M.; Heurtebize, J.; Durrieu, S.; LaValley, C.; Dauzat, J. Mapping plant area index of tropical evergreen forest by airborne laser scanning. A cross-validation study using LAI2200 optical sensor. Remote Sens. Environ. 2017, 198, 254–266. [Google Scholar] [CrossRef]
- Badreldin, N.; Sánchez-Azofeifa, G.A. Estimating Forest Biomass Dynamics by Integrating Multi-Temporal Landsat Satellite Images with Ground and Airborne LiDAR Data in the Coal Valley Mine, Alberta, Canada. Remote Sens. 2015, 7, 2832–2849. [Google Scholar] [CrossRef]
- Deo, R.K.; Russell, M.B.; Domke, G.M.; Woodall, C.; Falkowski, M.J.; Cohen, W.B. Using Landsat Time-series and LiDAR to Inform Aboveground Forest Biomass Baselines in Northern Minnesota, USA. Can. J. Remote Sens. 2016, 43, 28–47. [Google Scholar] [CrossRef]
- Jha, N.; Tripathi, N.K.; Chanthorn, W.; Brockelman, W.; Nathalang, A.; Pélissier, R.; Pimmasarn, S.; Ploton, P.; Sasaki, N.; Virdis, S.G.P.; et al. Forest aboveground biomass stock and resilience in a tropical landscape of Thailand. Biogeosciences 2020, 17, 121–134. [Google Scholar] [CrossRef]
- Brockelman, W.Y.; Nathalang, A.; Gale, G.A. The Mo Singto forest dynamics plot, Khao Yai National Park, Thailand. Nat. Hist. Bull. Siam Soc. 2011, 57, 35–55. [Google Scholar]
- Chanthorn, W.; Hartig, F.; Brockelman, W.Y. Structure and community composition in a tropical forest suggest a change of ecological processes during stand development. For. Ecol. Manag. 2017, 404, 100–107. [Google Scholar] [CrossRef]
- Tang, J.; Pilesjö, P. Estimating slope from raster data: A test of eight different algorithms in flat, undulating and steep terrain. River Basin Manag. VI 2011, 146, 143–154. [Google Scholar] [CrossRef]
- Mattivi, P.; Franci, F.; Lambertini, A.; Bitelli, G. TWI computation: A comparison of different open source GISs. Open Geospat. Data Softw. Stand. 2019, 4. [Google Scholar] [CrossRef]
- FAO. The State of Food and Agriculture 2012; FAO publications: Rome, Italy, 2012. [Google Scholar]
- Clark, D.B.; Olivas, P.C.; Oberbauer, S.F.; Clark, D.; Ryan, M.G. First direct landscape-scale measurement of tropical rain forest Leaf Area Index, a key driver of global primary productivity. Ecol. Lett. 2007, 11, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Aragão, L.E.O.; Shimabukuro, Y.E.; Espirito-Santo, F.; Williams, M. Landscape pattern and spatial variability of leaf area index in Eastern Amazonia. For. Ecol. Manag. 2005, 211, 240–256. [Google Scholar] [CrossRef]
- Moser, G.; Hertel, D.; Leuschner, C. Altitudinal Change in LAI and Stand Leaf Biomass in Tropical Montane Forests: A Transect Study in Ecuador and a Pan-Tropical Meta-Analysis. Ecosystems 2007, 10, 924–935. [Google Scholar] [CrossRef]
- Unger, M.; Homeier, J.; Leuschner, C. Relationships among leaf area index, below-canopy light availability and tree diversity along a transect from tropical lowland to montane forests in NE Ecuador. Trop. Ecol. 2013, 54, 33–45. [Google Scholar]
- Liu, L.; Zhang, R.; Zuo, Z. The Relationship between Soil Moisture and LAI in Different Types of Soil in Central Eastern China. J. Hydrometeorol. 2016, 17, 2733–2742. [Google Scholar] [CrossRef]
- Zhang, W.; Hu, B.; Woods, M.; Brown, G. Characterizing Forest Succession Stages for Wildlife Habitat Assessment Using Multispectral Airborne Imagery. Forests 2017, 8, 234. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MeanPAI (m2 m−2) | MeanCHM (m) | Age (Years) | Number of Sample Plots | |
|---|---|---|---|---|
| Stand initiation stage plots (SIS) | 4.19 ± 0.3 | 9.25 ± 0.3 | 8–20 | 3 |
| Stem exclusion stage plots (SES) | 6.63 ± 0.3 | 20.22 ± 1.8 | 35–41 | 3 |
| Old-growth stage plots (OGS) | 8.92 ± 0.65 | 21.87 ± 2.71 | >200 | 60 |
| Description | Coefficients | p-Value | S.E. | t Stat |
|---|---|---|---|---|
| Depedent PAI value | ||||
| Slope | 0.10156 ** | 0.000 | 0.02096 | 4.85 |
| Elevation | −0.30905 * | 0.017 | 0.12877 | −2.40 |
| Elevation2 | 0.00020 * | 0.020 | 0.00008 | 2.33 |
| Intercept | 126.02670 * | 0.010 | 48.82904 | 2.58 |
| Dependent PAI residual value | ||||
| Slope | −0.11043 * | 0.016 | 0.04588 | −2.41 |
| Elevation | 0.15834 | 0.070 | 0.08728 | 1.81 |
| Elevation2 | −0.00011 | 0.057 | 0.00006 | −1.91 |
| Intercept | −56.76872 | 0.088 | 33.20418 | −1.71 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pimmasarn, S.; Tripathi, N.K.; Ninsawat, S.; Sasaki, N. Applying LiDAR to Quantify the Plant Area Index Along a Successional Gradient in a Tropical Forest of Thailand. Forests 2020, 11, 520. https://doi.org/10.3390/f11050520
Pimmasarn S, Tripathi NK, Ninsawat S, Sasaki N. Applying LiDAR to Quantify the Plant Area Index Along a Successional Gradient in a Tropical Forest of Thailand. Forests. 2020; 11(5):520. https://doi.org/10.3390/f11050520
Chicago/Turabian StylePimmasarn, Siriruk, Nitin Kumar Tripathi, Sarawut Ninsawat, and Nophea Sasaki. 2020. "Applying LiDAR to Quantify the Plant Area Index Along a Successional Gradient in a Tropical Forest of Thailand" Forests 11, no. 5: 520. https://doi.org/10.3390/f11050520
APA StylePimmasarn, S., Tripathi, N. K., Ninsawat, S., & Sasaki, N. (2020). Applying LiDAR to Quantify the Plant Area Index Along a Successional Gradient in a Tropical Forest of Thailand. Forests, 11(5), 520. https://doi.org/10.3390/f11050520