Multidecadal Growth of Western White Pine and Interior Douglas-Fir Following Site Preparation


Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Design
2.2. Data Collection
2.3. Laboratory Processing and Analysis
2.4. Statistical Analysis
3. Results
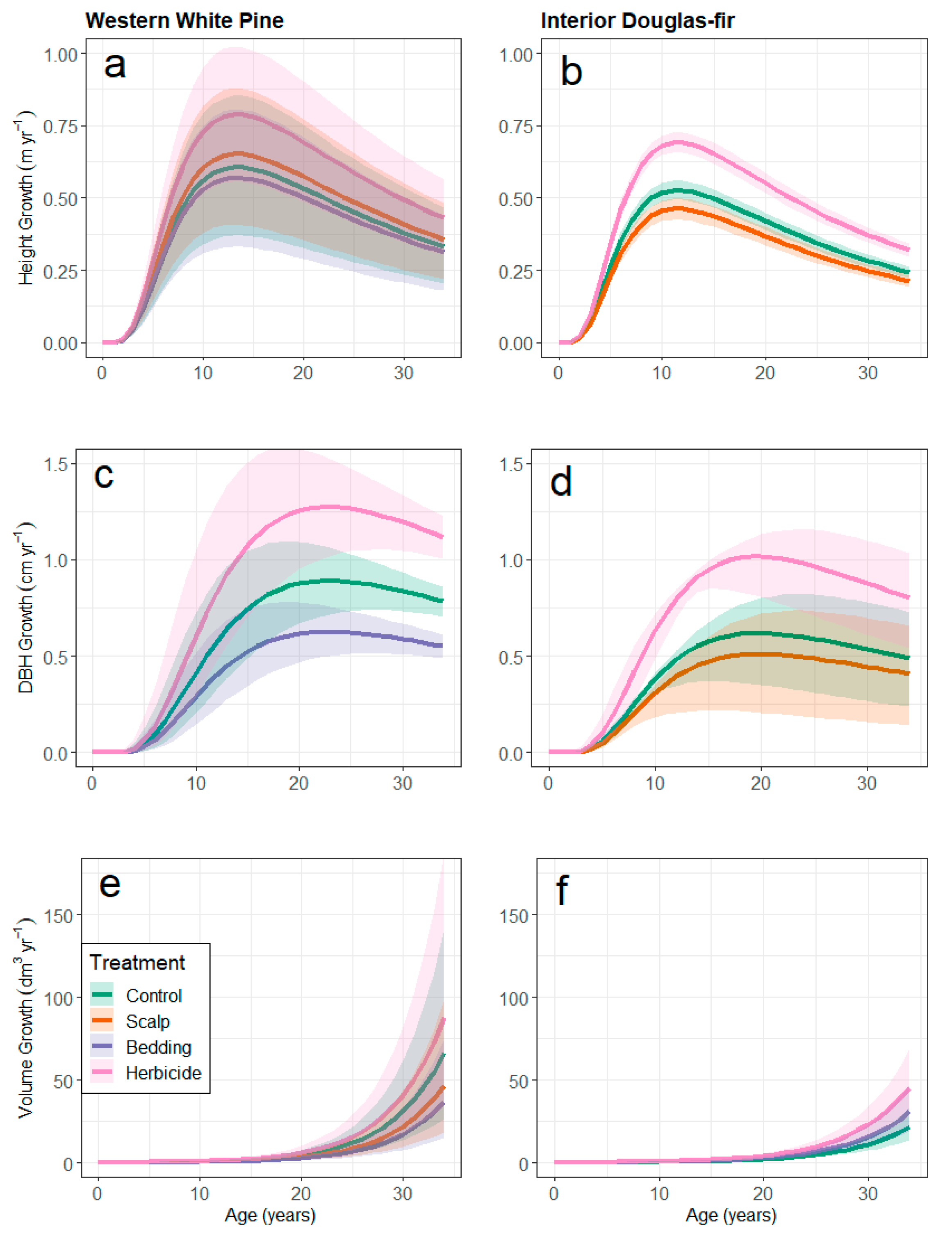
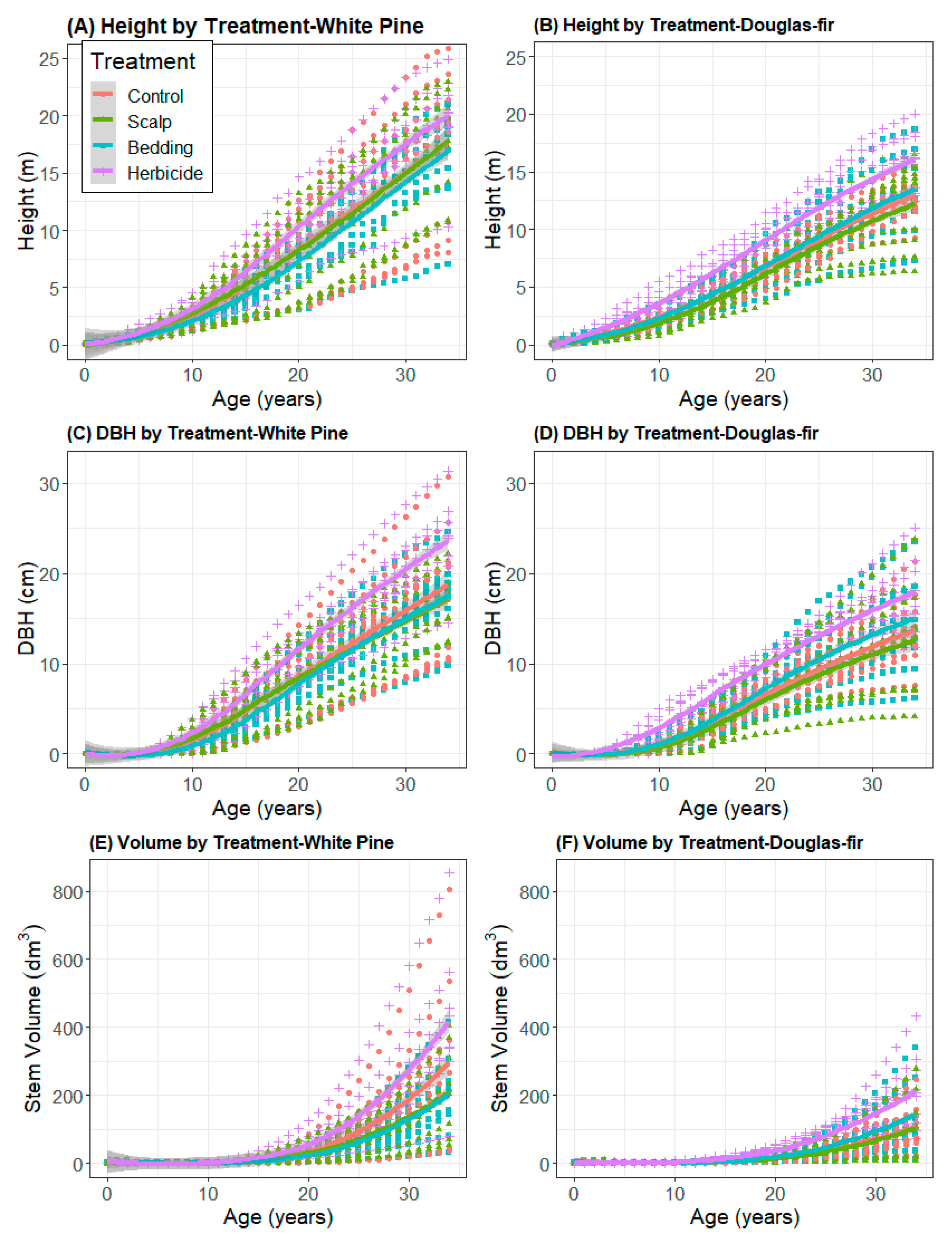
3.1. Cumulative Size and Growth by Treatment
3.2. Growth and Yield Models
3.3. Herbicide-Related Age-Shifts
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Ashton, M.S.; Kelty, M.J. The Practice of Silviculture: Applied Forest Ecology, 10th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2018; p. 758. [Google Scholar]
- Wagner, R.G.; Little, K.M.; Richardson, B.; McNabb, K. The role of vegetation management for enhancing productivity of the world’s forests. Forestry 2006, 79, 57–79. [Google Scholar] [CrossRef]
- Wiensczyk, A.; Swift, K.; Morneault, A.; Thiffault, N.; Szuba, K.; Bell, F.W. An overview of the efficacy of vegetation management alternatives for conifer regeneration in boreal forests. For. Chron. 2011, 87, 175–200. [Google Scholar] [CrossRef]
- Binkley, D.; Fisher, R.F. Ecology and Management of Forest Soils, 4th ed.; Wiley-Blackwell: Oxford, UK, 2013; p. 362. [Google Scholar]
- Morris, L.A.; Lowery, R.F. Influence of site preparation on soil conditions affecting stand establishment and tree growth. South. J. Appl. For. 1988, 12, 170–178. [Google Scholar] [CrossRef]
- South, D.B.; Miller, J.H.; Kimberley, M.O.; Vanderschaaf, C.L. Determining productivity gains from herbaceous vegetation management with ‘age-shift’calculations. Forestry 2006, 79, 43–56. [Google Scholar] [CrossRef]
- Kershaw, J.A.; Ducey, M.J.; Beers, T.W.; Husch, B. Forest Mensuration, 5th ed.; John Wiley and Sons, Inc.: Chichester, West Sussex, UK, 2017; p. 613. [Google Scholar]
- Duff, G.H.; Nolan, N.J. Growth and morphogenesis in the Canadian forest species: I. The controls of cambial and apical activity in Pinus resinosa. Can. J. Bot. 1953, 31, 471–513. [Google Scholar] [CrossRef]
- Wilhite, L.P.; Jones, E.P., Jr. Bedding effects in maturing slash pine stands. South. J. Appl. For. 1981, 5, 24–27. [Google Scholar] [CrossRef]
- Heitzman, E.; Pregitzer, K.S.; Miller, R.O. Origin and early development of northern white-cedar stands in northern Michigan. Can. J. For. Res. 1997, 27, 1953–1961. [Google Scholar] [CrossRef]
- Weiskittel, A.R.; Hann, D.W.; Kershaw, J.A.; Vanclay, J.K. Forest Growth and Yield Modeling; John Wiley & Sons, Ltd.: West Sussex, UK, 2011. [Google Scholar]
- Fontes, L.; Tomé, M.; Coelho, M.B.; Wright, H.; Luis, J.S.; Savill, P. Modelling dominant height growth of Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) in Portugal. Forestry 2003, 76, 509–523. [Google Scholar] [CrossRef]
- McDill, M.E.; Amateis, R.L. Measuring forest site quality using the parameters of a dimensionally compatible height growth function. For. Sci. 1992, 38, 409–429. [Google Scholar] [CrossRef]
- Finklin, A.I. Climate of Priest River Experimental Forest, Northern Idaho; Gen. Tech. Rep. INT-159. 53p; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1983.
- Page, D.S. The Effect of Bedding on Soil Physical and Chemical Properties in Northern Idaho. Masters‘ Thesis, Michigan Technological University, Houghton, MI, USA, 1985. [Google Scholar]
- Harvey, A.E.; Page-Dumroese, D.S.; Jurgensen, M.F.; Graham, R.T.; Tonn, J.R. Site preparation alters biomass, root and ectomycorrhizal development of outplanted western white pine and Douglas-fir. New For. 1996, 11, 255–270. [Google Scholar] [CrossRef]
- Soil Survey Staff. Natural Resources Conservation Service, United States Department of Agriculture. Web Soil Survey. Available online: https://websoilsurvey.sc.egov.usda.gov (accessed on 26 March 2019).
- Graham, R.T.; Harvey, A.E.; Jurgensen, M.F. Effect of site preparation on survival and growth of Douglas-fir (Pseudotsuga menziessi Mirb. Franco.) seedlings. New For. 1989, 3, 89–98. [Google Scholar] [CrossRef]
- Tinkham, W.T.; Denner, R.; Graham, R.T. Climate, Snowpack, and Streamflow of Priest River Experimental Forest, Revisted; Gen. Tech. Rep. RMRS-331. 49p; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2015.
- Cooper, S.V.; Neiman, K.E.; Roberts, D.W. Forest Habitat Types of Northern Idaho: A Second Approximation; Gen. Tech. Rep. INT-236. 143p; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1991.
- Page-Dumroese, D.S.; Jurgensen, M.F.; Harvey, A.E.; Graham, R.T.; Tonn, J.R. Soil changes and tree seedling response associated with site preparation in northern Idaho. West. J. Appl. For. 1997, 12, 81–88. [Google Scholar] [CrossRef]
- Carmean, W.H. Site index curves for upland oaks in the Central States. For. Sci. 1972, 18, 109–120. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017.
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R-core; EISPACK. Nlme: Linear and Nonlinear Mixed Effects Models. Available online: https://rdrr.io/cran/nlme/ (accessed on 14 April 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer-Verlag: New York, NY, USA, 2016. [Google Scholar]
- Sit, V.; Poulin-Costello, M. Catalogue of Curves for Curve Fitting; Biometrics Information Handbook No. 4. 110p; Province of British Columbia, Ministry of Forests Research Program: Victoria, BC, Canada, 1994. [Google Scholar]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer Verlag New York, LLC: New York, NY, USA, 2000; p. 528. [Google Scholar]
- O’Neal, D.D.; Houston, A.E.; Buckner, E.R.; Meadows, J.S. An index of competition based on relative crown position and size. In General Technical Report SRS-1; Boyd, E.M., Ed.; U.S. Department of Agriculture Forest Service, Southern Research Station: Asheville, NC, USA, 1995; pp. 172–175. [Google Scholar]
- Ketchum, D.E.; Wellner, C.A.; Evans, S.S. Western white pine management programs realigned on northern Rocky Mountain National Forests. J. For. 1968, 66, 329–332. [Google Scholar] [CrossRef]
- Fins, L.; Byler, J.; Ferguson, D.; Harvey, A.; Mahalovich, M.F.; McDonald, G.; Miller, D.; Schwandt, J.; Zack, A. Return of the giants: Restoring western white pine to the Inland Northwest. J. For. 2002, 100, 20–26. [Google Scholar] [CrossRef]
- Zhang, J.; Powers, R.F.; Oliver, W.W.; Young, D.H. Response of ponderosa pine plantations to competing vegetation control in Northern California, USA: A meta-analysis. Forestry 2013, 86, 3–11. [Google Scholar] [CrossRef]
- Newton, M.; Cole, E.C. Twenty-six-year response of ponderosa pine and Douglas-fir plantations to woody competitor density in treated stands of madrone and whiteleaf manzanita. For. Ecol. Manag. 2008, 256, 410–420. [Google Scholar] [CrossRef]
- Outcalt, K.W. Influence of bed height on the growth of slash and loblolly pine on a Leon fine sand in Northeast Florida. South. J. Appl. For. 1984, 8, 29–31. [Google Scholar] [CrossRef]
- Page-Dumroese, D.S.; Jurgensen, M.F.; Graham, R.T.; Harvey, A.E. Soil Chemical Properties of Raised Planting Beds in a Northern Idaho Forest; Research Paper INT-419. 7p; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1989.
- McKee, W.H.J.; Shoulders, E. Slash pine biomass response to site preparation soil properties. Proc. Soil Sci. Soc. Am. 1974, 38, 144–148. [Google Scholar] [CrossRef]
- Shoulders, E.; Terry, T.A. Dealing with site disturbance from harvesting and site preparation in the Lower Coastal Plain. In Principles of Maintaining Productivity on Prepared Sites: Proceedings of the Symposium, New Orleans, LA, USA, 21–22 March 1978; Mississippi State University: Mississippi State, MS, USA, 1978; pp. 85–89. [Google Scholar]
- Schultz, R.P. Environmental Change after Site Preparation and Slash Pine Planting on a Floodwood Site; Research Paper SE-156. 20p; U.S. Department of Agriculture, Forest Service, Southeastern Forest Experiment Station: Asheville, NC, USA, 1976. [Google Scholar]
- Kyle, K.H.; Andrews, L.J.; Fox, T.R.; Aust, W.M.; Burger, J.A.; Hansen, G.H. Long-term effects of drainage, bedding, and fertilization on growth of loblolly pine (Pinus taeda L.) in the Coastal Plain of Virginia. South. J. Appl. For. 2005, 29, 205–214. [Google Scholar] [CrossRef]
- Zhang, J.; Busse, M.D.; Young, D.H.; Fiddler, G.O.; Sherlock, J.W.; TenPas, J.D. Aboveground biomass responses to organic matter removal, soil compaction, and competing vegetation control on 20-year mixed conifer plantations in California. For. Ecol. Manag. 2017, 401, 341–353. [Google Scholar] [CrossRef]
- Lorenzetti, F.; Delagrange, S.; Bouffard, D.; Nolet, P. Establishment, survivorship, and growth of yellow birch seedlings after site preparation treatments in large gaps. For. Ecol. Manag. 2008, 254, 350–361. [Google Scholar] [CrossRef]
- Macadam, A.; Kabzems, R. Vegetation management improves early growth of white spruce more than mechanical site preparation treatments. North. J. Appl. For. 2006, 23, 35–46. [Google Scholar] [CrossRef]
- Mason, E.G.; Cullen, A.W.J.; Rijkse, W.C. Growth of two Pinus radiata stock types on ripped and ripped/bedded plots at Karioi Forest. N. Z. J. For. Sci. 1988, 18, 287–296. [Google Scholar]
- Nelson, A.S.; Bragg, D.C. Multidecadal response of naturally regenerated southern pine to early competition control and commerical thinning. For. Sci. 2016, 62, 115–124. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Treatment | n | DBH (cm) | Height (m) | Volume (dm3) | Competition Index |
|---|---|---|---|---|---|
| White pine | |||||
| Control | 9 | 18.8 (10.3–30.7) | 17.4 (8.1–25.9) | 296.9 (34.9–805.5) | 22.8 (9.3–30.9) |
| Scalp | 9 | 17.1 (12.3–22.2) | 17.4 (10.6–22.9) | 210.8 (52.2–405.3) | 26.6 (15.8–33.3) |
| Bedding | 10 | 17.4 (9.8–24.6) | 16.4 (7.0–21.0) | 202.2 (31.6–416.9) | 32.2 (14.8–82.7) |
| Herbicide | 8 | 23.6 (14.5–31.4) | 19.8 (10.3–24.9) | 421.2 (80.2–855.9) | 21.4 (12.9–33.3) |
| Douglas-fir | |||||
| Control | 10 | 13.5 (7.6–21.3) | 12.7 (9.2–15.5) | 105.2 (23.8–247.9) | 34.4 (24.4–41.5) |
| Scalp | 9 | 12.5 (4.1–23.8) | 12.0 (6.4–16.5) | 101.8 (6.6–277.9) | 45.4 (8.7–101.2) |
| Bedding | 10 | 14.8 (6.1–23.6) | 13.2 (7.3–18.7) | 139.7 (12.4–341.3) | 34.9 (12.1–60.9) |
| Herbicide | 10 | 17.7 (11.8–25.1) | 15.8 (12.4–20.0) | 206.5 (71.8–432.3) | 27.7 (15.5–42.8) |
| Treatment | Age 5 | Age 10 | Age 15 | Age 20 | Age 25 | Age 30 | Age 34 | |
|---|---|---|---|---|---|---|---|---|
| White pine | ||||||||
| Control | 0.75 ab | 2.41 a | 4.97 a | 8.23 ab | 11.53 ab | 14.90 a | 17.17 a | |
| Scalp | 0.69 a | 2.71 ab | 5.13 ab | 8.10 a | 11.19 a | 14.94 a | 17.22 a | |
| Bedding | 0.71 a | 1.97 a | 4.28 a | 7.12 a | 10.65 a | 14.55 a | 16.55 a | |
| Herbicide | 1.05 b | 3.18 b | 6.50 b | 10.34 b | 14.25 b | 17.92 a | 20.10 a | |
| Std. Error | 0.13 | 0.70 | 1.75 | 2.54 | 2.93 | 3.26 | 3.33 | |
| Douglas-fir | ||||||||
| Control | 0.94 ab | 2.45 a | 4.42 a | 6.82 a | 9.36 a | 12.01 a | 13.24 a | |
| Scalp | 0.63 a | 1.87 a | 3.86 a | 6.16 a | 8.87 a | 11.08 a | 12.41 a | |
| Bedding | 0.78 a | 2.20 a | 4.39 a | 6.79 a | 9.41 a | 12.05 a | 13.52 ab | |
| Herbicide | 1.34 b | 3.58 b | 6.16 b | 9.26 b | 12.11 b | 14.99 b | 16.37 b | |
| Std. Error | 0.13 | 0.30 | 0.37 | 0.50 | 0.64 | 0.93 | 1.35 | |
| Treatment | Age 5 | Age 10 | Age 15 | Age 20 | Age 25 | Age 30 | Age 34 | |
|---|---|---|---|---|---|---|---|---|
| White pine | ||||||||
| Control | - | 1.0 a | 4.7 a | 8.5 a | 11.9 a | 15.5 a | 18.3 a | |
| Scalp | - | 1.4 ab | 5.0 a | 8.3 a | 11.4 a | 14.7 a | 17.1 a | |
| Bedding | - | 0.4 a | 3.7 a | 7.9 a | 11.9 a | 15.5 a | 18.0 a | |
| Herbicide | - | 2.1 b | 6.9 b | 11.8 b | 16.3 b | 20.7 b | 24.0 b | |
| Std. Error | 0.7 | 2.1 | 2.7 | 2.7 | 2.8 | 2.6 | ||
| Douglas-fir | ||||||||
| Control | - | 0.8 a | 4.0 a | 7.0 a | 10.0 ab | 13.2 ab | 15.1 ab | |
| Scalp | - | 0.4 a | 3.0 a | 6.1 a | 8.9 a | 11.4 a | 13.0 a | |
| Bedding | - | 0.7 a | 3.8 a | 7.4 ab | 10.9 ab | 14.0 ab | 15.8 ab | |
| Herbicide | - | 2.6 b | 6.4 b | 9.8 b | 13.2 b | 16.5 b | 18.3 b | |
| Std. Error | 0.4 | 0.5 | 0.7 | 1.2 | 2.0 | 3.0 | ||
| Variable | Cumulative Height | Cumulative DBH | Cumulative Volume | |||
|---|---|---|---|---|---|---|
| White Pine | Douglas-Fir | White Pine | Douglas-Fir | White Pine | Douglas-Fir | |
| a0 | 32.09774 (6.49013) | 24.32248 (0.93438) | 79.36525 (3.02039) | 48.16674 (13.61810) | 0.48165 (0.05717) | 0.33330 (0.03168) |
| a1 | 2.38997 (1.09853) | −3.02248 (0.54826) | − | −8.45039 (1.31893) | −0.13564 (0.02973) | − |
| a2 | −1.99569 (0.97916) | − | −23.67744 (2.38052) | − | −0.21425 (0.03371) | 0.13970 (0.02360) |
| a3 | 9.64942 (1.04102) | 7.60409 (0.53928) | 33.34426 (2.59648) | 29.42092 (1.46571) | 0.15329 (0.03037) | 0.36238 (0.03774) |
| b | −28.64136 (0.66678) | −24.97974 (0.47124) | −48.01134 (4.70142) | −40.69068 (4.19755) | 0.18634 (0.01117) | 0.16950 (0.00628) |
| CAR(1) Φ | 0.46404 | 0.24057 | 0.30560 | 0.01932 | 0.30208 | 0.02913 |
| Power variance parameter | 1.19325 | 0.87609 | 2.7609 | 2.3825 | 1.4950 | 1.2303 |
| R2 (fixed) | 0.781 | 0.885 | 0.795 | 0.775 | 0.589 | 0.645 |
| R2 (fixed +random) | 0.904 | 0.890 | 0.854 | 0.822 | 0.757 | 0.725 |
| RMSE | 3.64 | 2.26 | 4.31 | 3.70 | 88.08 | 44.15 |
| Age-Shift (Years) | Gain | ||||
|---|---|---|---|---|---|
| Species | Age 5 | Age 15 | Age 25 | Age 34 | Age 34 |
| Height | |||||
| White pine | 0.3 | 2.1 | 5.0 | 8.5 | 30% |
| Douglas-fir | 0.4 | 2.4 | 5.7 | 9.7 | 19% |
| DBH | |||||
| White pine | − | 1.7 | 4.2 | 7.1 | 43% |
| Douglas-fir | − | 2.5 | 6.1 | 10.2 | 63% |
| Stem Volume | |||||
| White pine | − | − | 25.2 | 33.8 | 31% |
| Douglas-fir | − | − | 28.0 | 36.7 | 109% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cherico, J.R.; Nelson, A.S.; Jain, T.B.; Graham, R.T. Multidecadal Growth of Western White Pine and Interior Douglas-Fir Following Site Preparation. Forests 2020, 11, 509. https://doi.org/10.3390/f11050509
Cherico JR, Nelson AS, Jain TB, Graham RT. Multidecadal Growth of Western White Pine and Interior Douglas-Fir Following Site Preparation. Forests. 2020; 11(5):509. https://doi.org/10.3390/f11050509
Chicago/Turabian StyleCherico, Jonathan R., Andrew S. Nelson, Teresa B. Jain, and Russell T. Graham. 2020. "Multidecadal Growth of Western White Pine and Interior Douglas-Fir Following Site Preparation" Forests 11, no. 5: 509. https://doi.org/10.3390/f11050509
APA StyleCherico, J. R., Nelson, A. S., Jain, T. B., & Graham, R. T. (2020). Multidecadal Growth of Western White Pine and Interior Douglas-Fir Following Site Preparation. Forests, 11(5), 509. https://doi.org/10.3390/f11050509