Success Factors for Experimental Partial Harvesting in Unmanaged Boreal Forest: 10-Year Stand Yield Results


 ,
,
Abstract
:1. Introduction
2. Methods
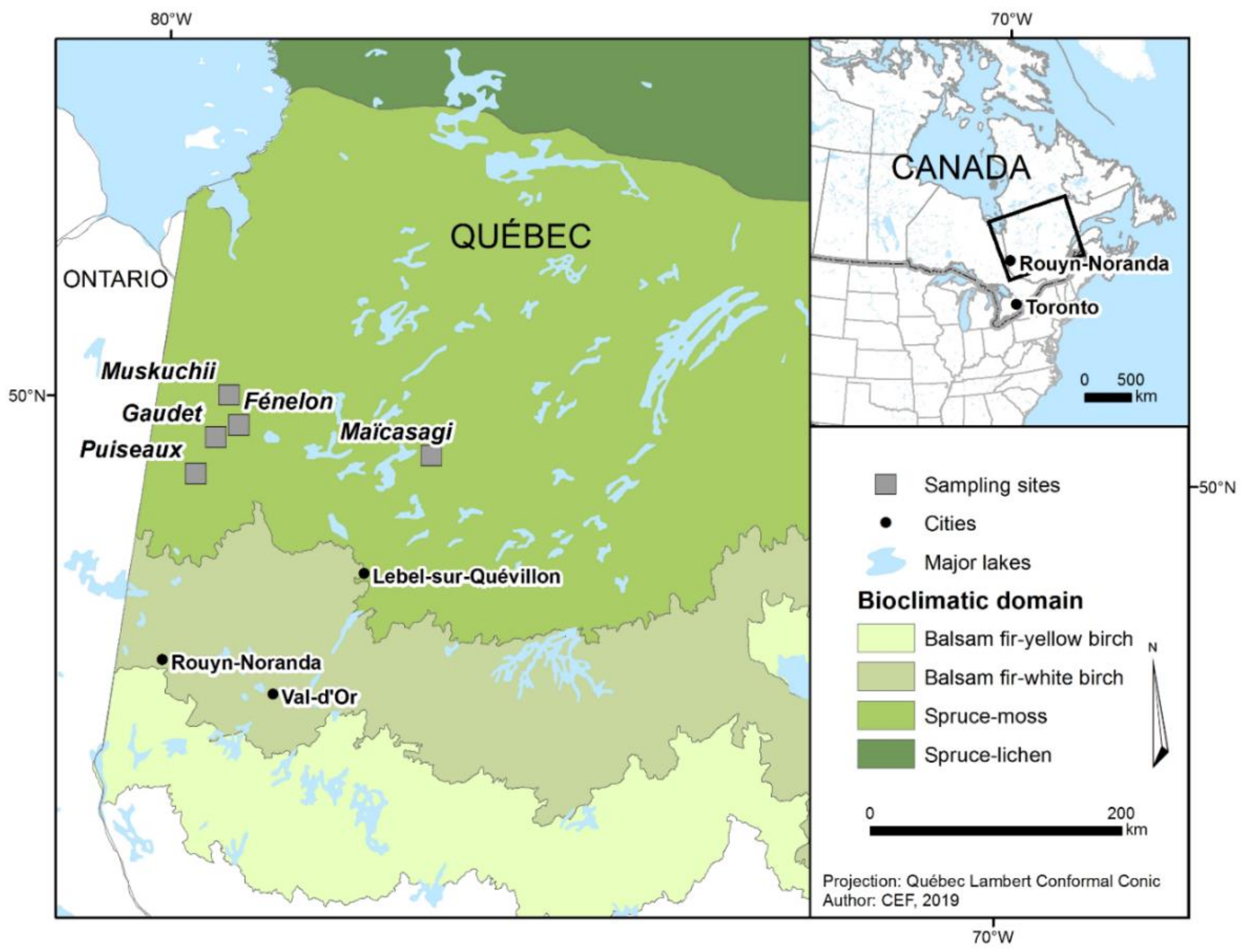
2.1. Study Area
2.2. Experimental Design
2.3. Forest Inventory Data
2.4. Data Analyses
3. Results
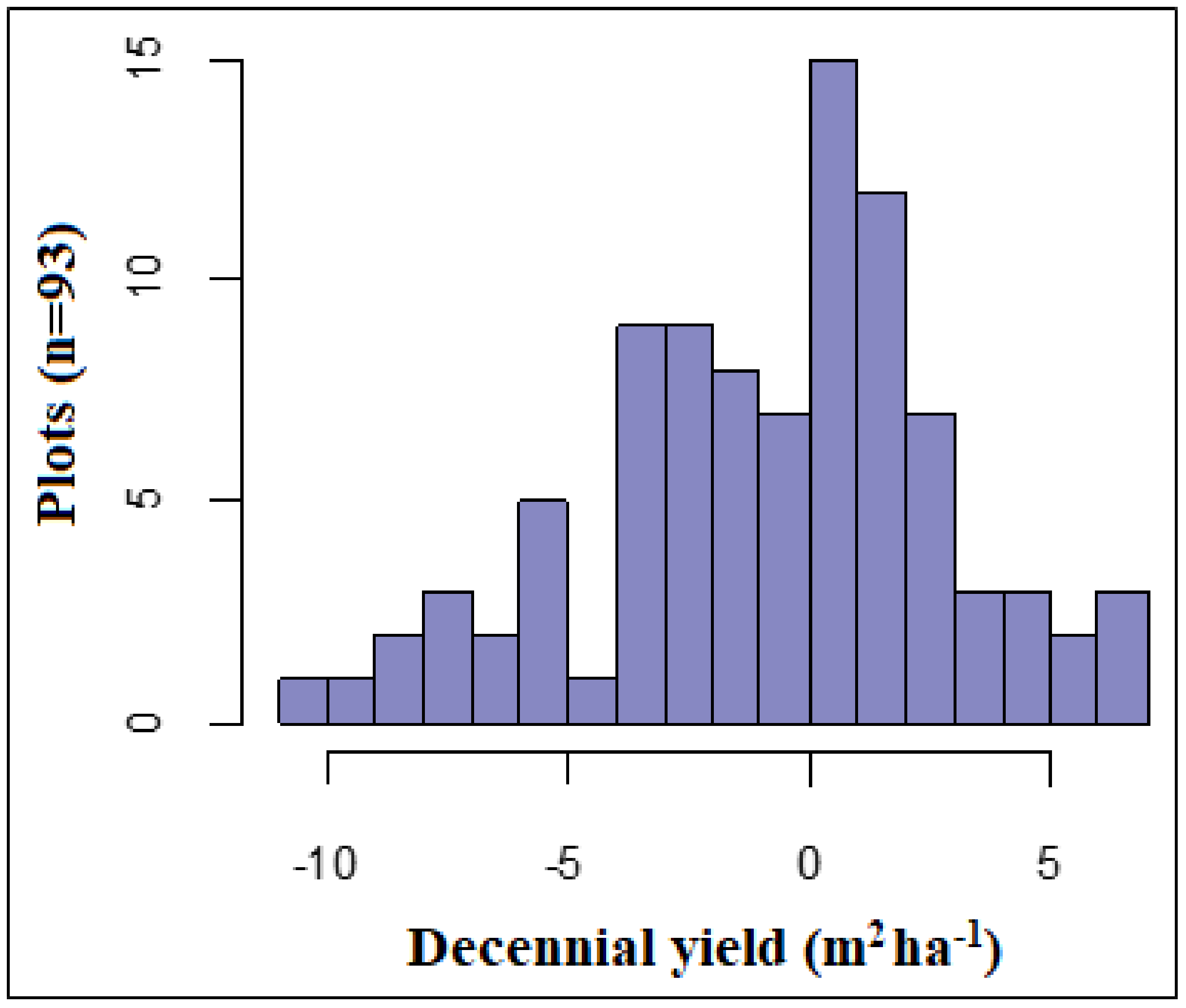
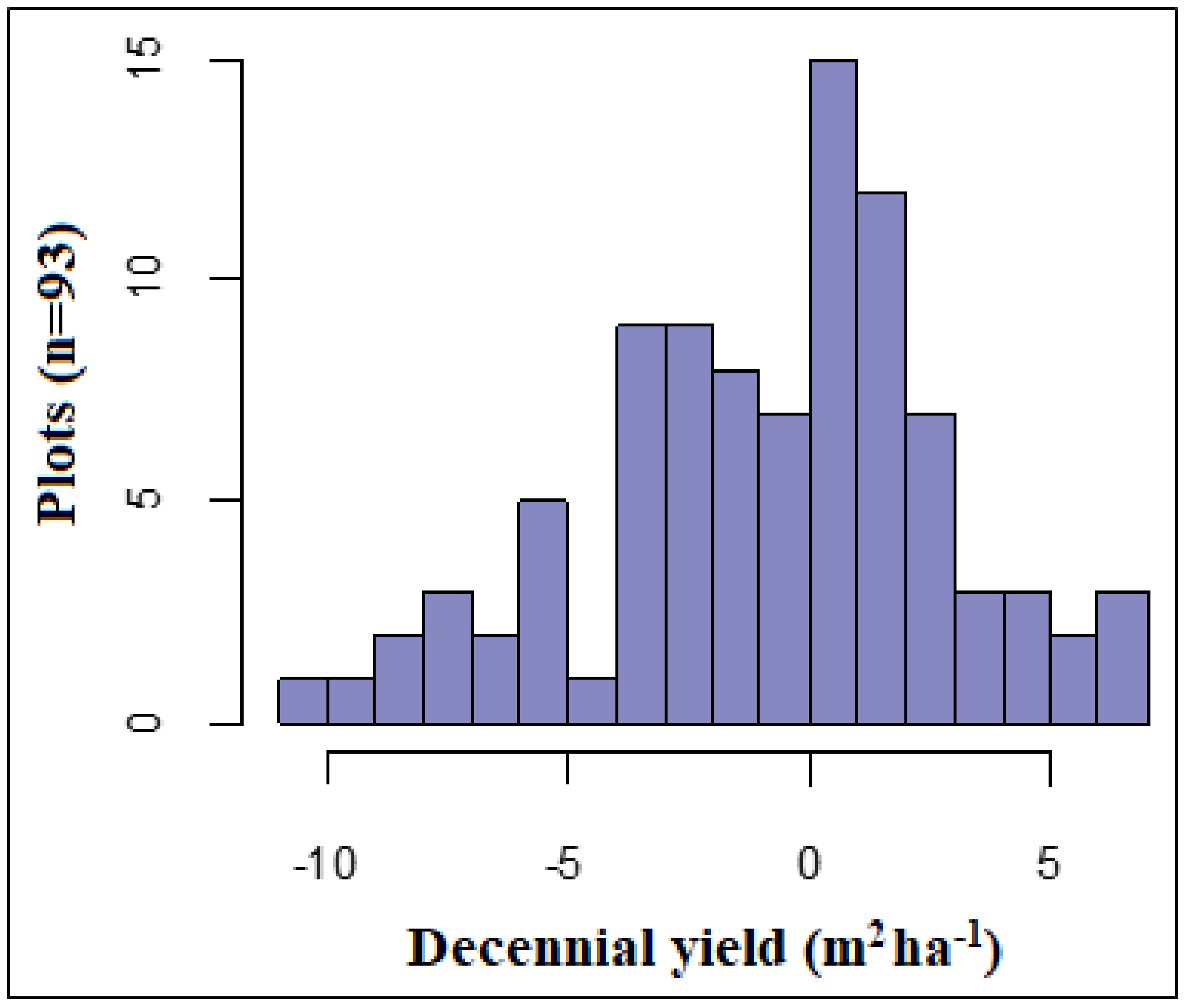
3.1. Decennial Yield and Partial Harvest Success
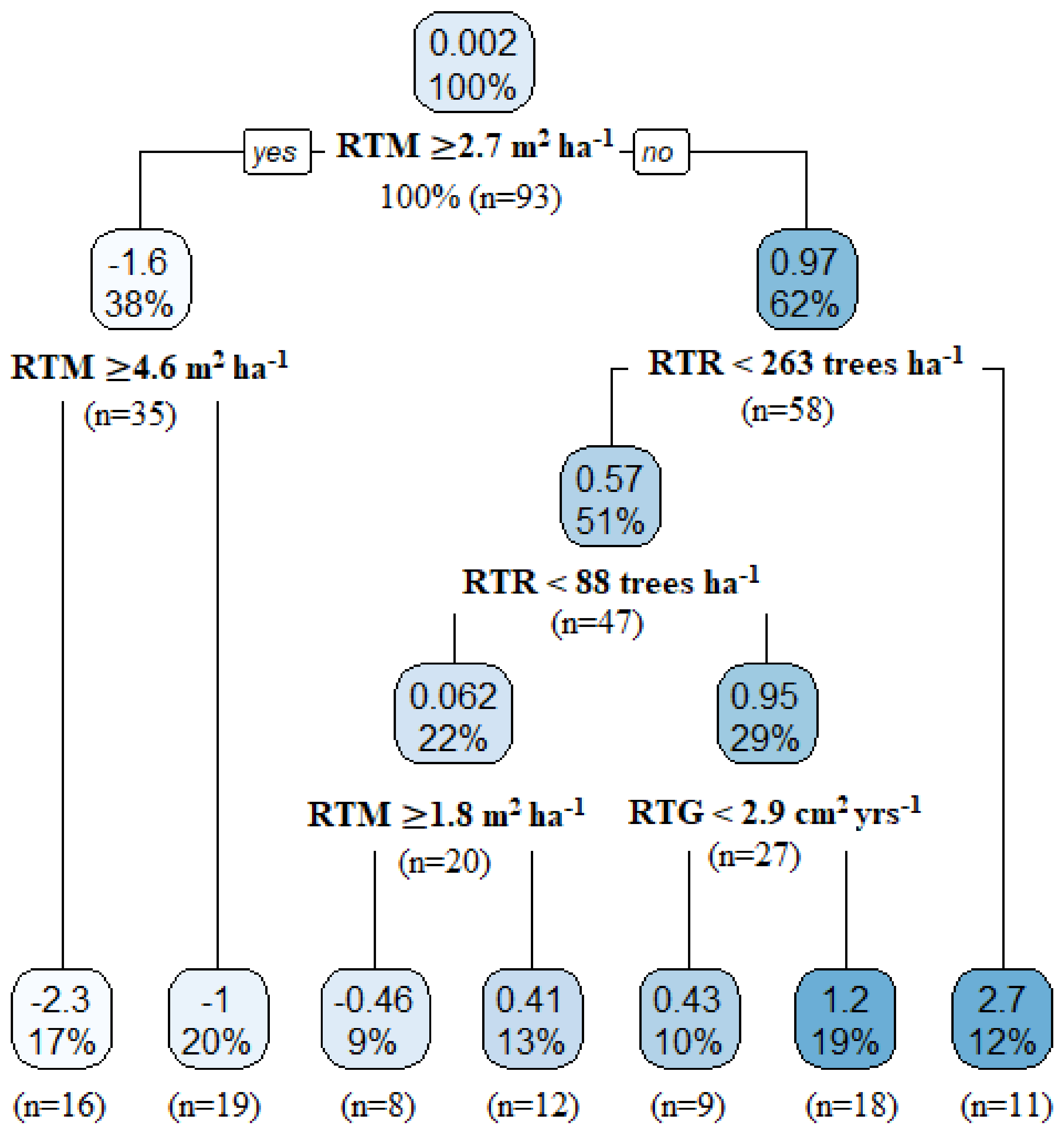
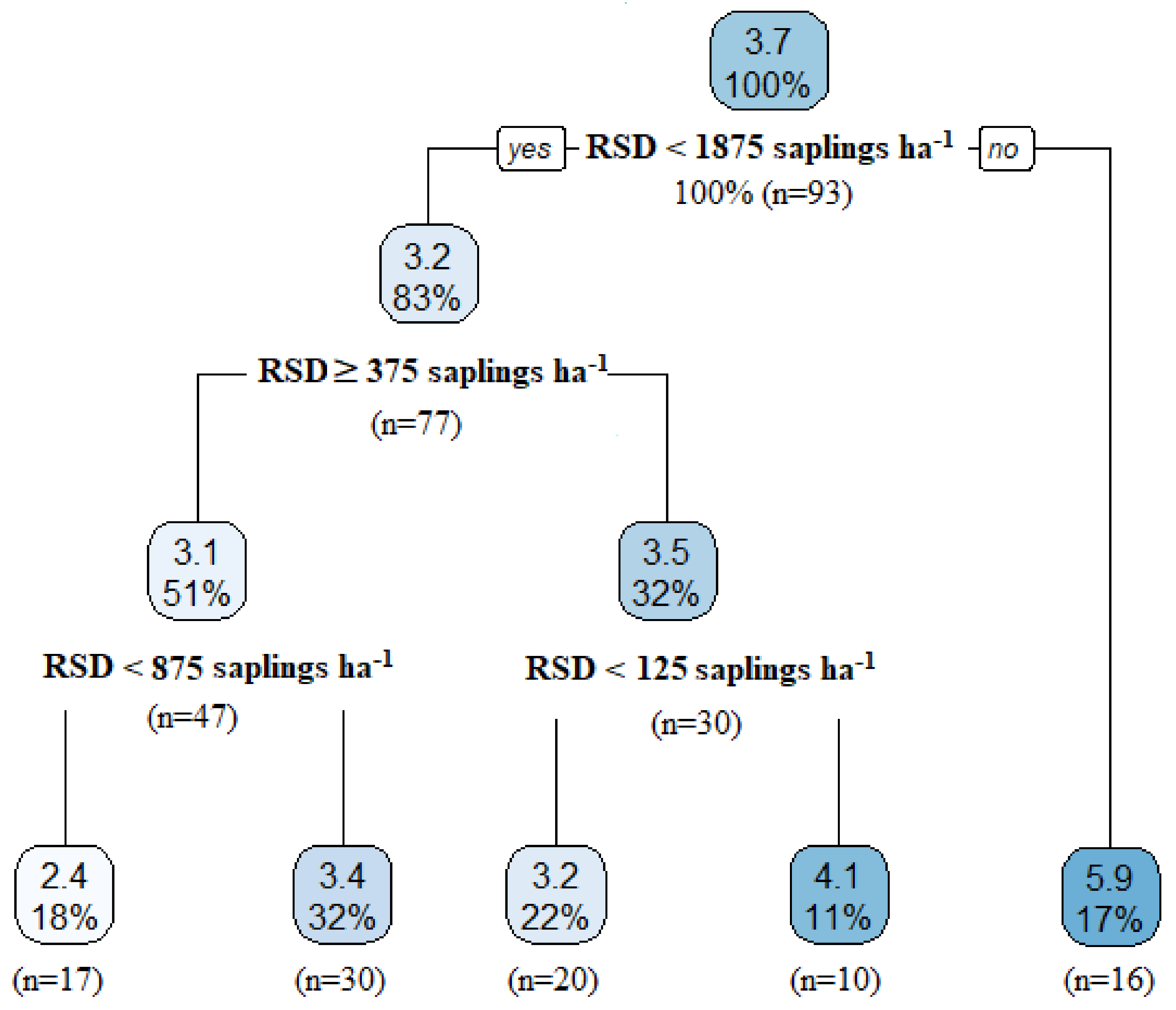
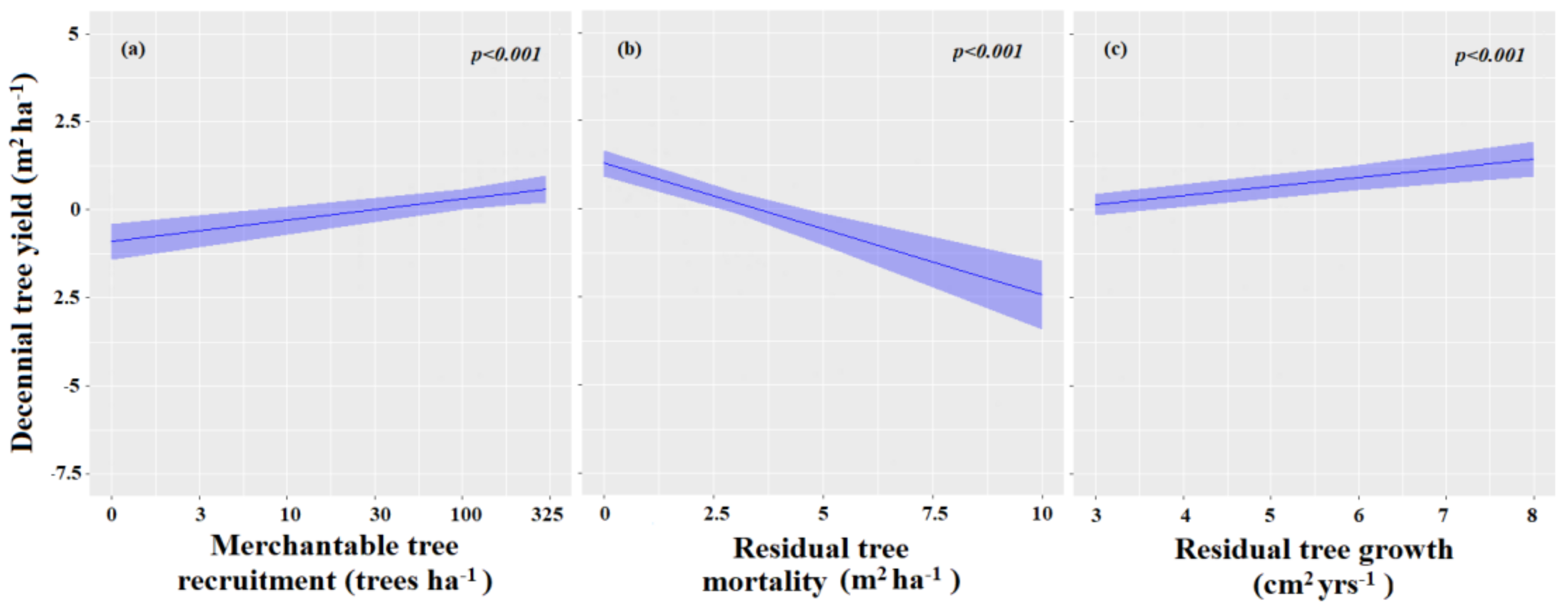
3.2. Importance of Stand Components on Decennial Stand Yield Following Partial Harvesting
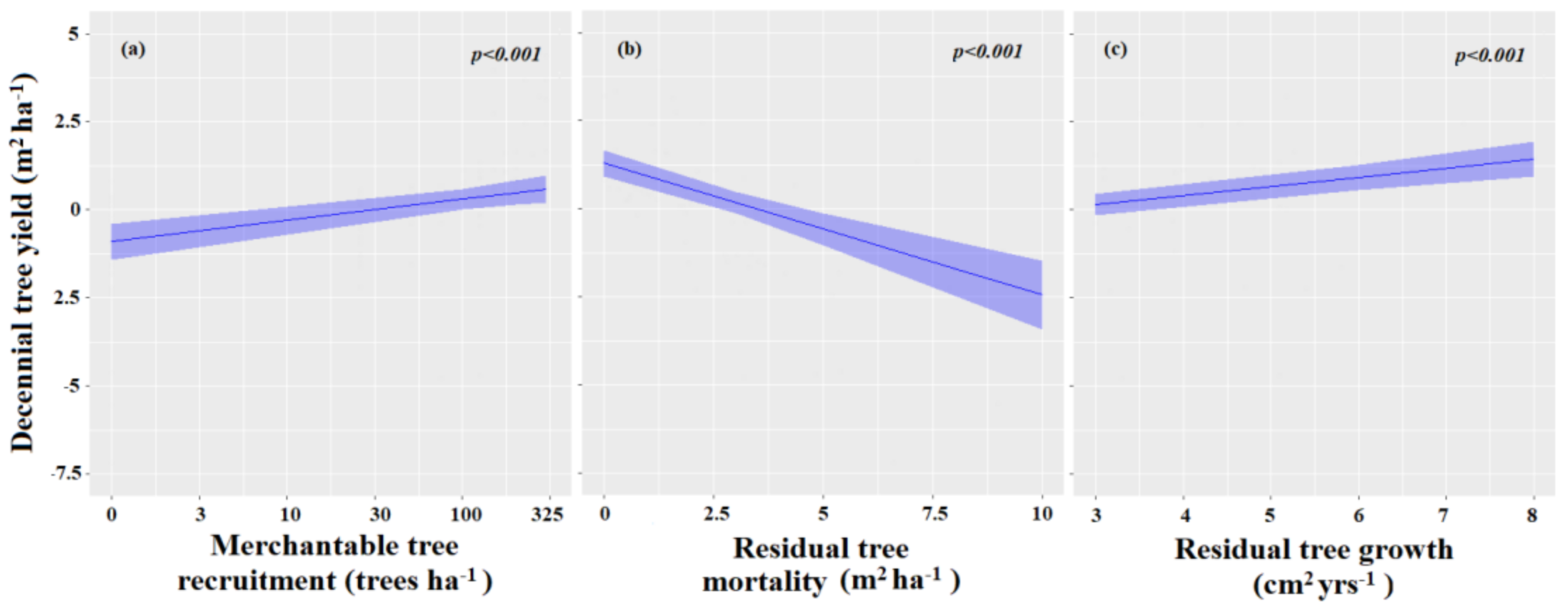
3.3. Factors Influencing Components of Stand Yield Ten Years Post-Harvest
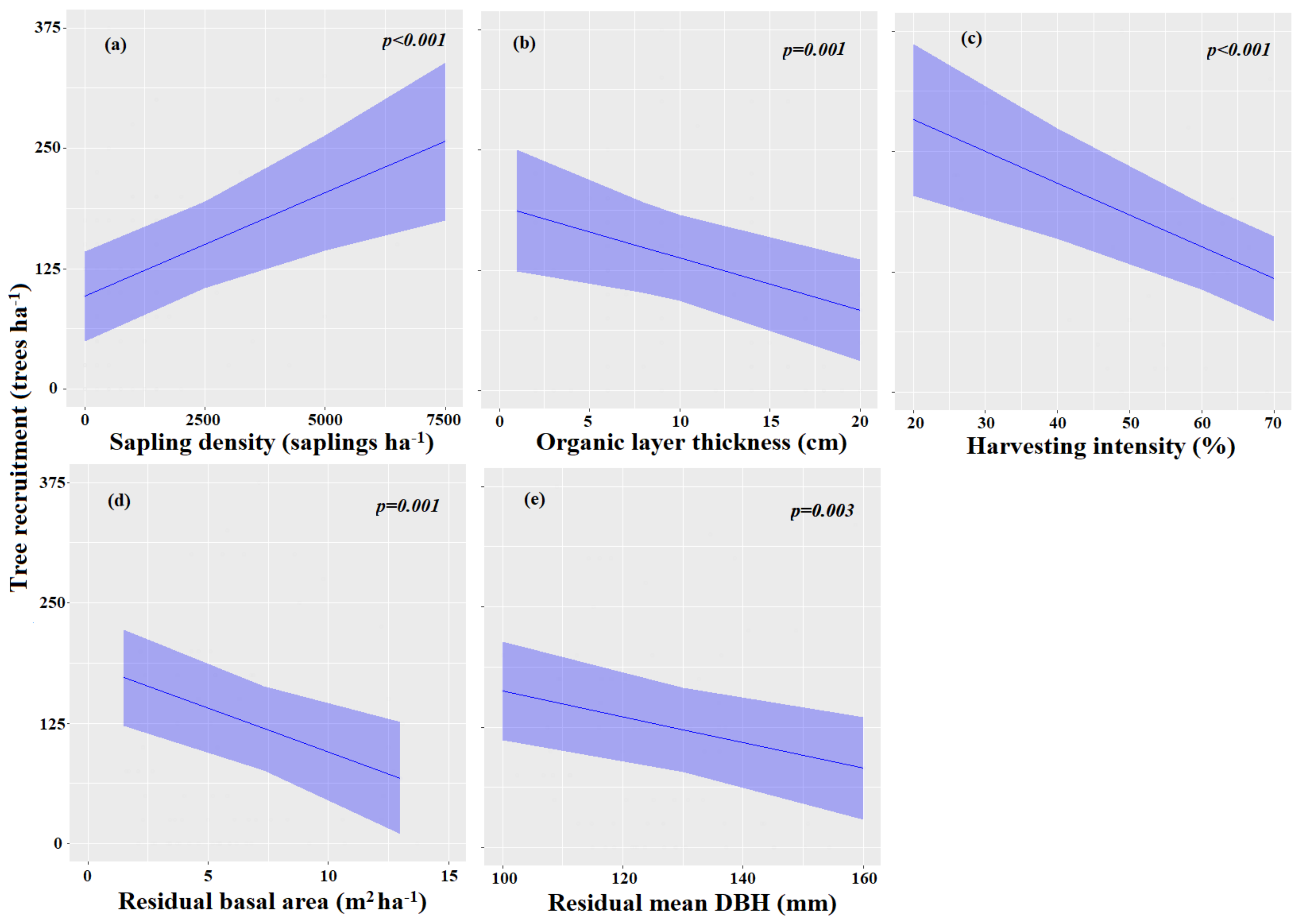
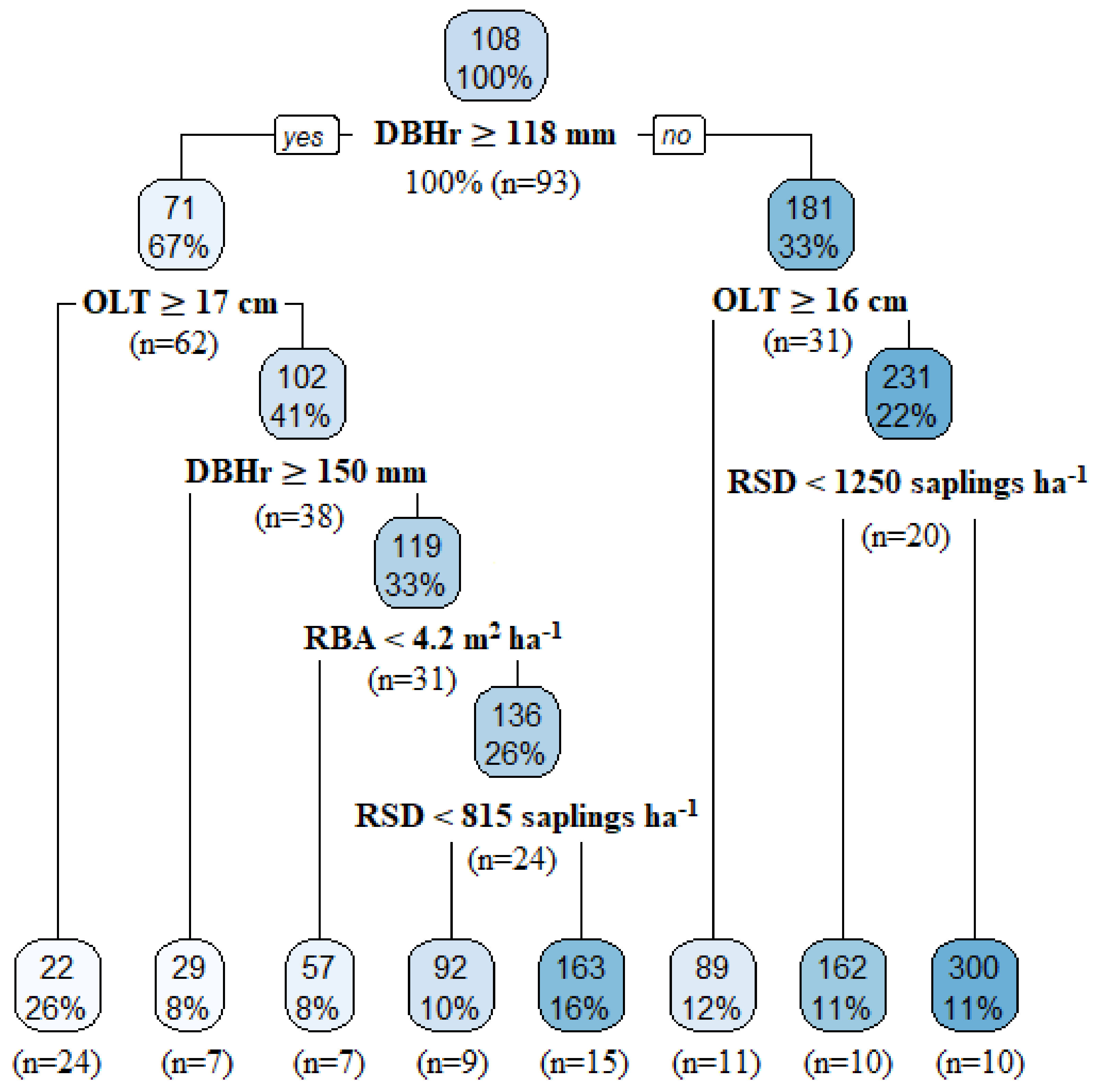
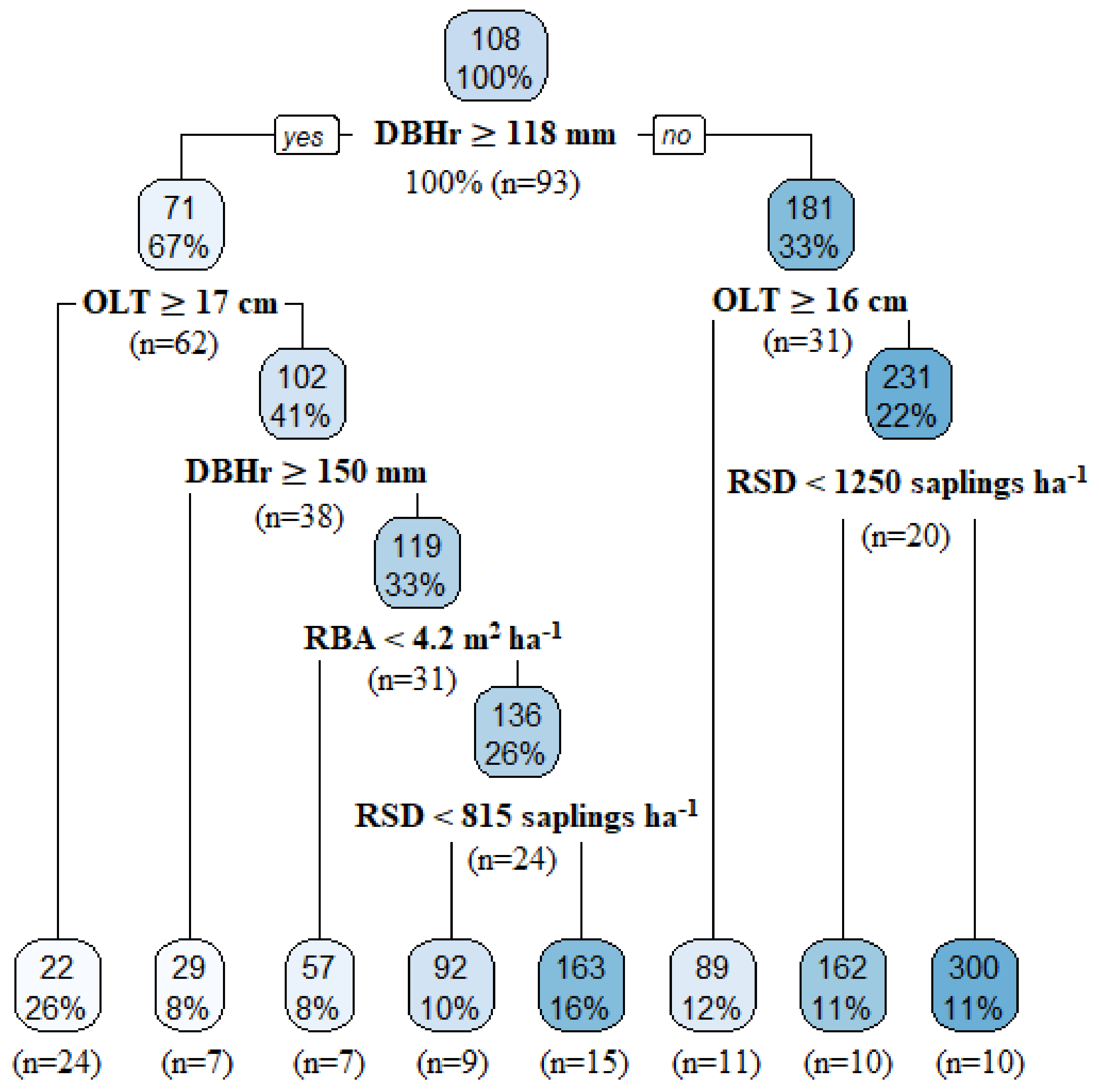
3.3.1. Factors Influencing Merchantable Tree Recruitment
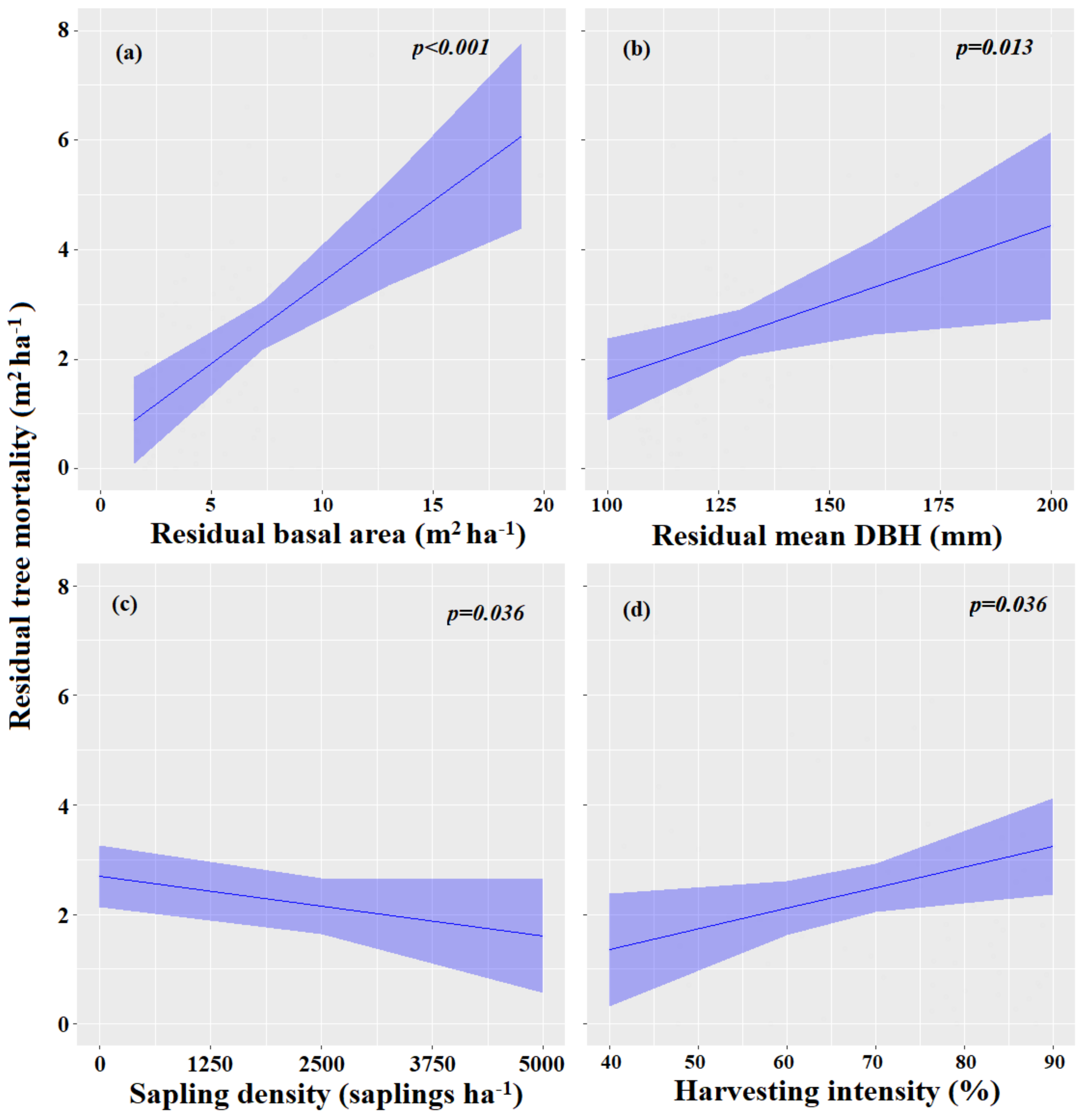
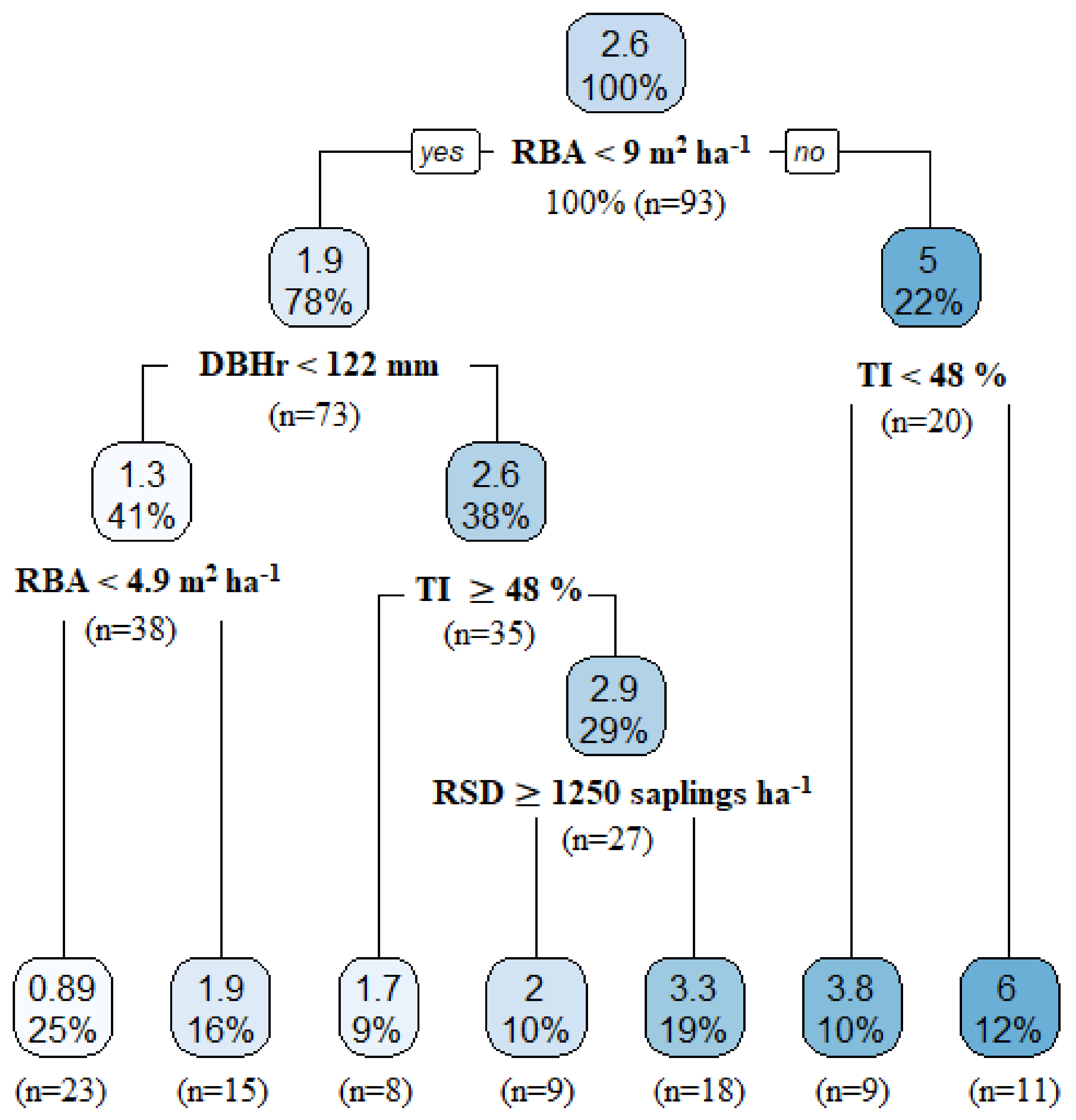
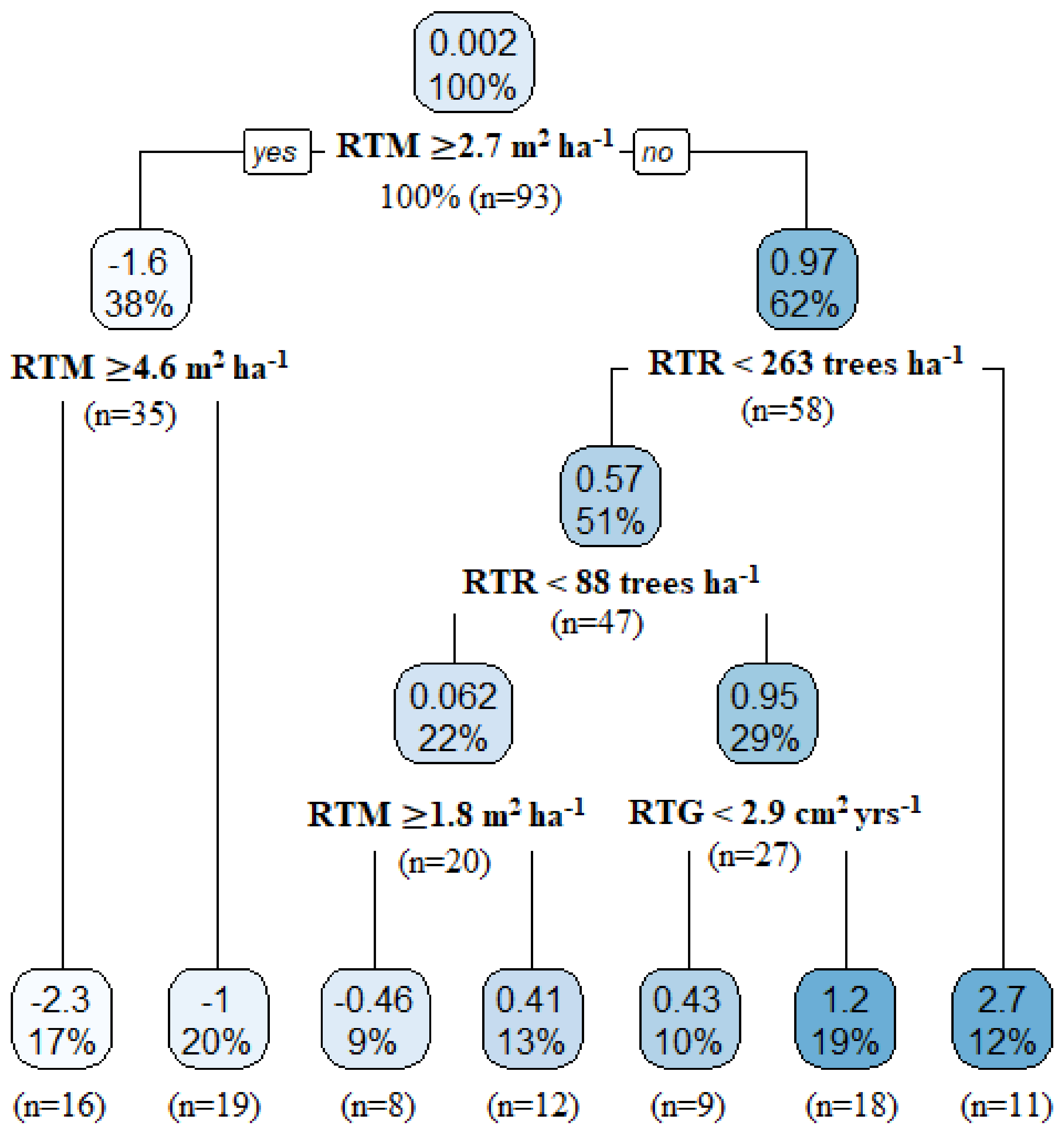
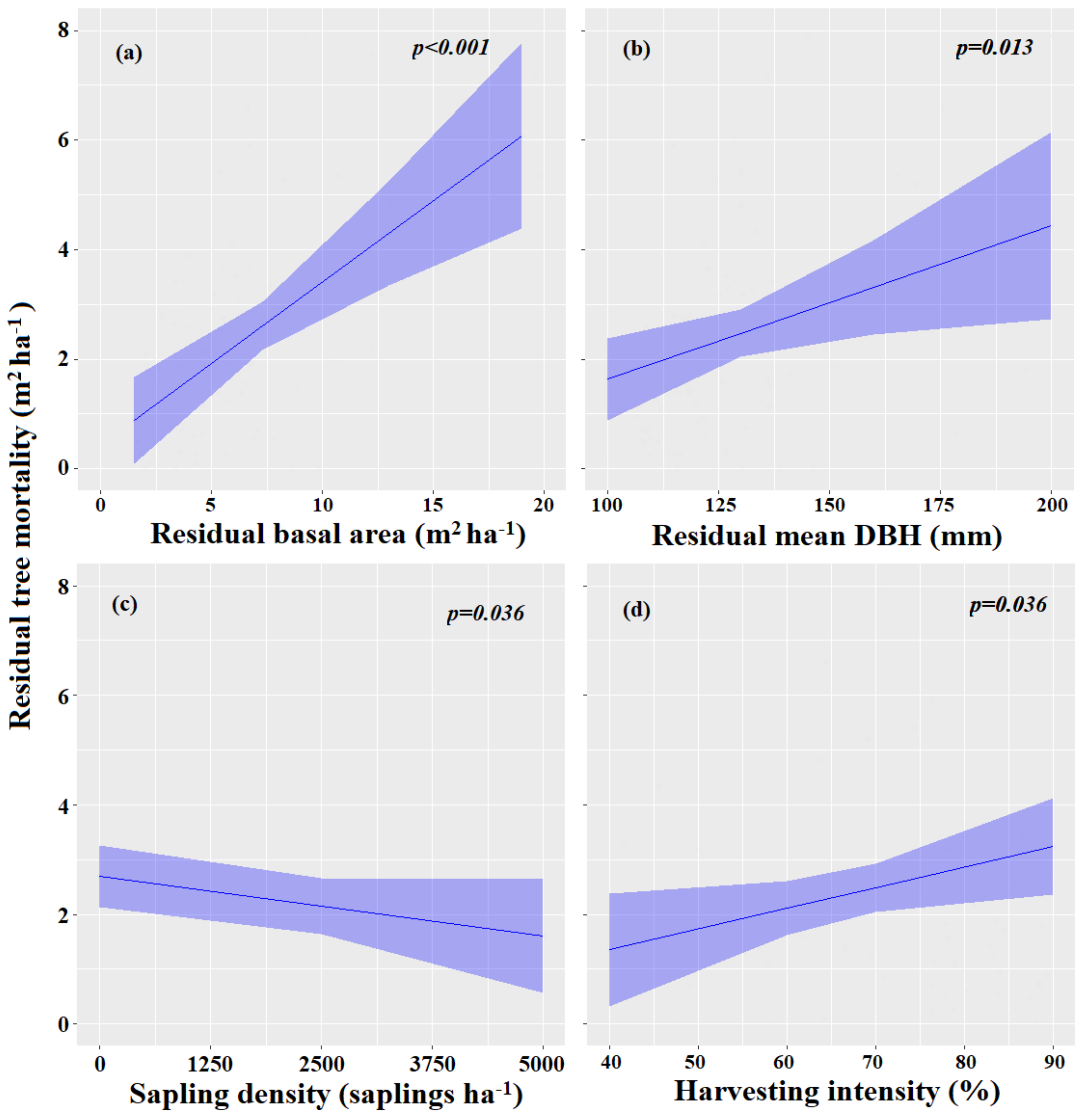
3.3.2. Factors Influencing Tree Mortality
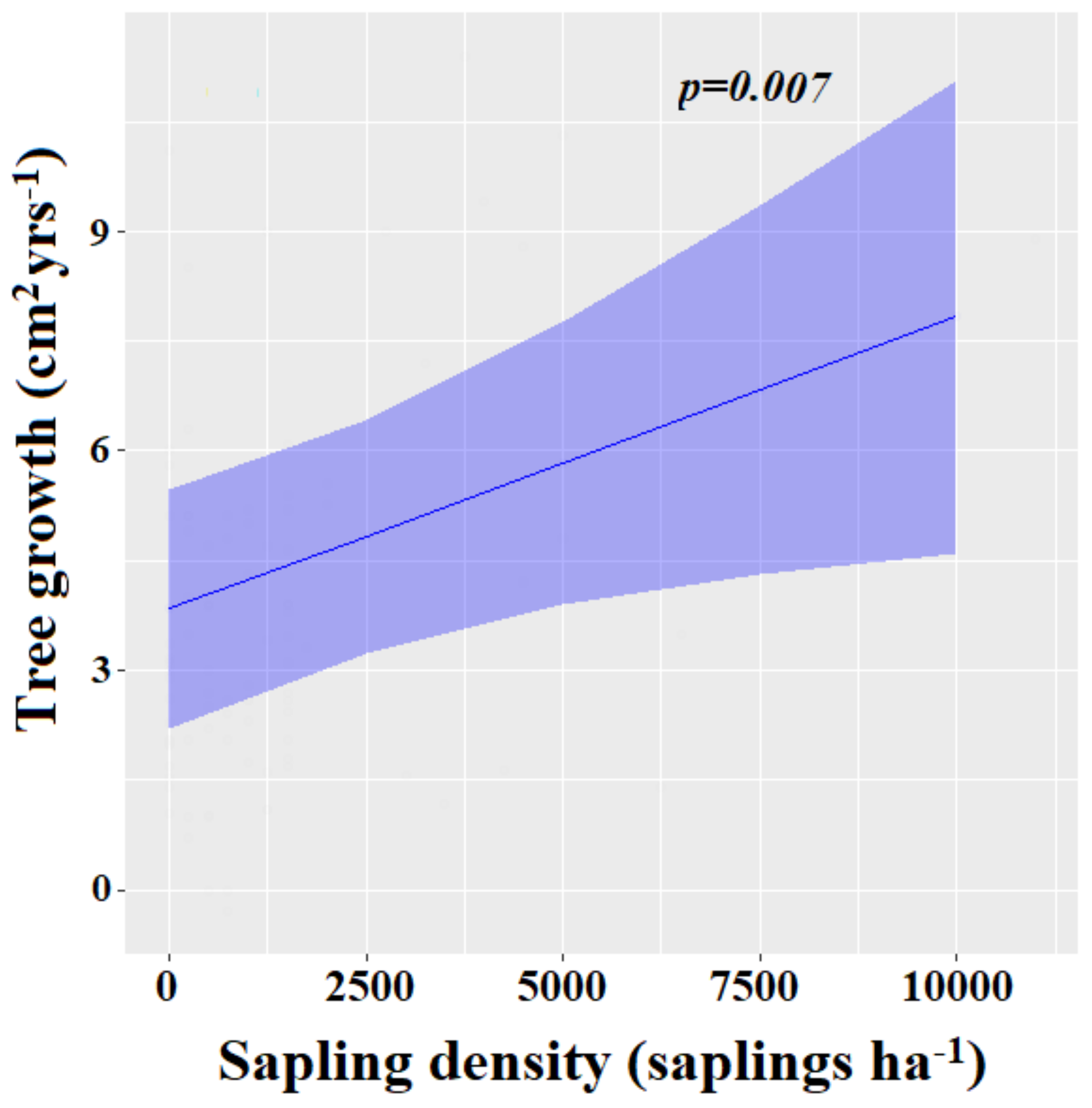
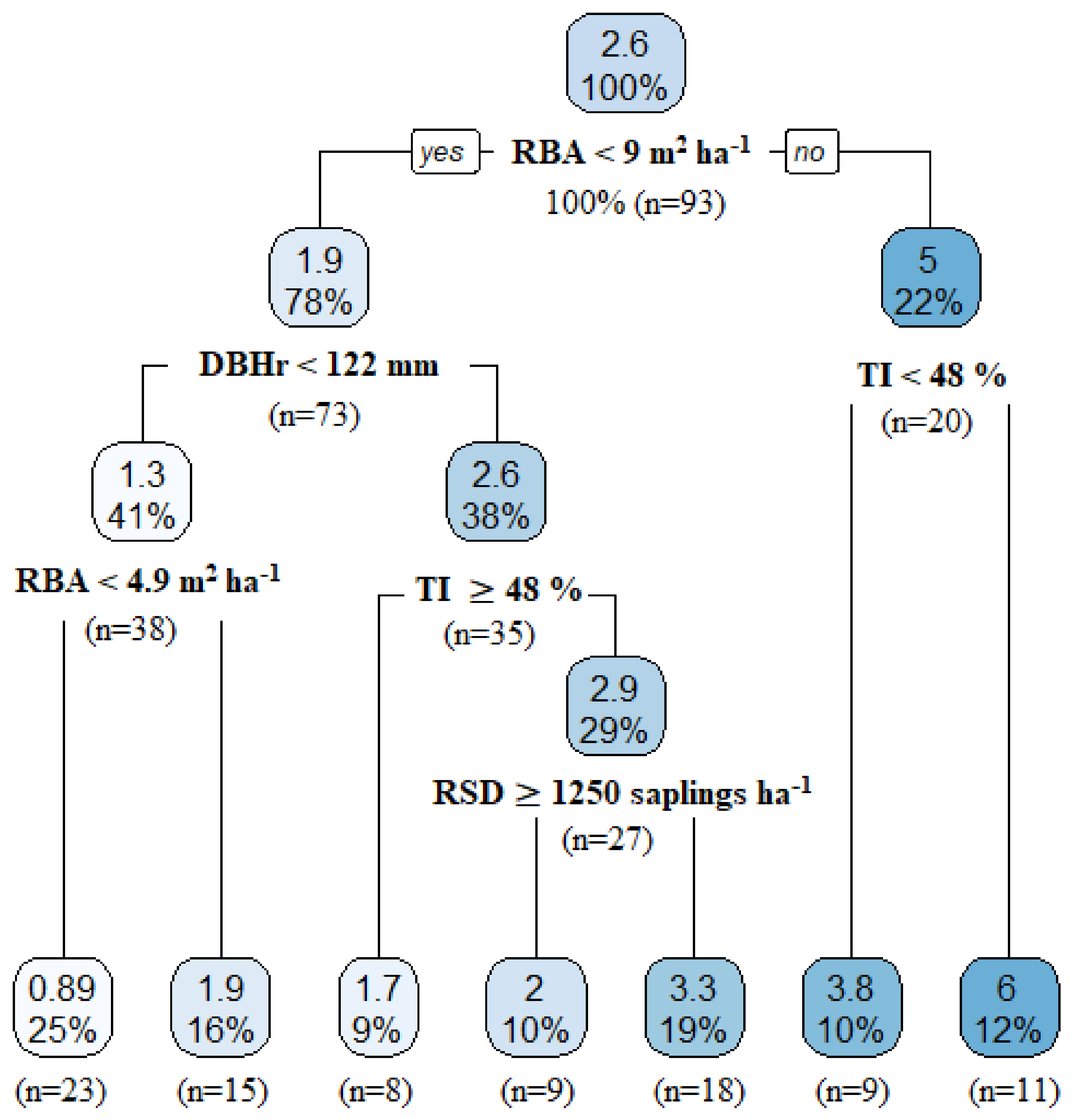
3.3.3. Factors Influencing the Residual Tree Growth
4. Discussion
5. Conclusions and Implications for Forest Management
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fixed Effect | Model AIC | Estimate | SE | t Value | Pr (>|t|) |
|---|---|---|---|---|---|
| One factor models | |||||
| Residual tree mortality | 338.4 | −0.5 | 0.06 | −7.7 | 1.81 × 10−11 *** |
| Merchantable tree recruitment | 351.5 | 1.2 | 0.2 | 6.4 | 2.1 × 10−8 *** |
| Residual tree growth | 368.3 | 0.3 | 0.07 | 4.9 | 1.11 × 10−5 *** |
| Two factor models | |||||
| Residual tree mortality + | 324.5 | −0.4 | 0.06 | −5.8 | 2.86 × 10−5 *** |
| Merchantable tree recruitement | 0.8 | 0.2 | 4.6 | 8.12 × 10−8 *** | |
| Residual tree mortality + | 317.2 | −0.5 | 0.05 | −8.5 | 3.51 × 10−13 *** |
| Residual tree growth | 0.3 | 0.05 | 5.7 | 3.04 × 10−7 *** | |
| Three factor model | |||||
| Residual tree mortality + | 304.2 | −0.4 | 0.06 | −6.3 | 8.54 × 10−9 *** |
| Residual tree growth+ | 0.2 | 0.05 | 4.9 | 3.95 × 10−6 *** | |
| Merchantable tree recruitment | 0.6 | 0.1 | 3.9 | 0.000149 *** |
References
- Franklin, J.F.; Berg, D.; Thornburg, D.; Tappeiner, J.C. Alternative Silvicultural Approaches to Timber Harvesting: Variable Retention Harvest Systems. In Creating a Forestry for the 21st Century: The Science of Ecosystem Management; Island Press: Washington, DC, USA, 1997. [Google Scholar]
- Drapeau, P.; Nappi, A.; Giroux, J.-F.; Leduc, A.; Savard, J.-P. Distribution patterns of birds associated with snags in natural and managed eastern boreal forests. In Proceedings of the Symposium on the Ecology and Management of Dead Wood in Western Forests’, Reno, NV; General Technical Report PSW-GTR-181; Laudenslayer, W.F., Shea, P.J., Valentine, B.E., Weatherspoon, C.P., Lisle, T.E., Eds.; USDA Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2002; pp. 193–205. [Google Scholar]
- Puettmann, K.J. Silvicultural challenges and options in the context of global change: “Simple” fixes and opportunities for new management approaches. J. For. 2011, 109, 321–331. [Google Scholar]
- Kuuluvainen, T.; Gauthier, S. Young and old forest in the boreal: Critical stages of ecosystem dynamics and management under global change. For. Ecosyst. 2018, 5, 26. [Google Scholar] [CrossRef]
- Sanghyun, K.; Axelsson, P.E.; Montoro Girona, M.; Senior, J.K. Continuous-cover forestry maintains soil fungal communities in Norway spruce dominated boreal forests. For. Ecol. Manag. 2020, 480, 118659. [Google Scholar]
- Gauthier, S.; Leduc, A.; Bergeron, Y. Forest dynamics modelling under natural fire cycles: A tool to define natural mosaic diversity for forest management. In Global to Local: Ecological Land Classification; Springer: Berlin/Heidelberg, Germany, 1996; pp. 417–434. [Google Scholar]
- Bergeron, Y.; Gauthier, S.; Kafka, V.; Lefort, P.; Lesieur, D. Natural fire frequency for the eastern Canadian boreal forest: Consequences for sustainable forestry. Can. J. For. Res. 2001, 31, 384–391. [Google Scholar] [CrossRef]
- Cyr, D.; Gauthier, S.; Bergeron, Y.; Carcaillet, C. Forest management is driving the eastern North American boreal forest outside its natural range of variability. Front. Ecol. Environ. 2009, 7, 519–524. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Bergeron, Y.; Coates, K.D. Restoration and ecosystem-based management in the circumboreal forest: Background, challenges, and opportunities. In Restoration of Boreal and Temperate Forests, 2nd ed.; CRC Lewis: Boca Raton, FL, USA, 2015; pp. 251–270. [Google Scholar]
- Gauthier, S. Ecosystem Management in the Boreal Forest; PUQ Les presses de l’université du Québec: Québec, QC, Canada, 2009. [Google Scholar]
- Gustafsson, L.; Kouki, J.; Sverdrup-Thygeson, A. Tree retention as a conservation measure in clear-cut forests of northern Europe: A review of ecological consequences. Scand. J. For. Res. 2010, 25, 295–308. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Grenfell, R. Natural disturbance emulation in boreal forest ecosystem management—Theories, strategies, and a comparison with conventional even-aged management. Can. J. For. Res. 2012, 42, 1185–1203. [Google Scholar] [CrossRef]
- Versluijs, M.; Hekkala, A.-M.; Lindberg, E.; Lämås, T.; Hjältén, J. Comparing the effects of even-aged thinning and selective felling on boreal forest birds. For. Ecol. Manag. 2020, 475, 118404. [Google Scholar] [CrossRef]
- Fenton, N.J.; Imbeau, L.; Work, T.; Jacobs, J.; Bescond, H.; Drapeau, P.; Bergeron, Y. Lessons learned from 12 years of ecological research on partial cuts in black spruce forests of northwestern Québec. For. Chron. 2013, 89, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Ruel, J.-C.; Fortin, D.; Pothier, D. Partial cutting in old-growth boreal stands: An integrated experiment. For. Chron. 2013, 89, 360–369. [Google Scholar] [CrossRef] [Green Version]
- Beese, W.J.; Dunsworth, B.; Zielke, K.; Bancroft, B. Maintaining attributes of old-growth forests in coastal BC through variable retention. For. Chron. 2003, 79, 570–578. [Google Scholar] [CrossRef]
- Rosenvald, R.; Lohmus, A. For what, when, and where is green-tree retention better than clear-cutting? A review of the biodiversity aspects. For. Ecol. Manag. 2008, 255, 1–15. [Google Scholar] [CrossRef]
- Siitonen, J. Forest management, coarse woody debris and saproxylic organisms: Fennoscandian boreal forests as an example. Ecol. Bull. 2001, 49, 11–41. [Google Scholar]
- Boudreault, C.; Bergeron, Y.; Gauthier, S.; Drapeau, P. Bryophyte and lichen communities in mature to old-growth stands in eastern boreal forests of Canada. Can. J. For. Res. 2002, 32, 1080–1093. [Google Scholar] [CrossRef]
- Drapeau, P.; Leduc, A.; Bergeron, Y.; Gauthier, S.; Savard, J.-P. Les communautés d’oiseaux des vieilles forêts de la pessière à mousses de la ceinture d’argile: Problèmes et solutions face à l’aménagement forestier. For. Chron. 2003, 79, 531–540. [Google Scholar] [CrossRef] [Green Version]
- Work, T.T.; Shorthouse, D.P.; Spence, J.R.; Volney, W.J.A.; Langor, D. Stand composition and structure of the boreal mixedwood and epigaeic arthropods of the Ecosystem Management Emulating Natural Disturbance (EMEND) landbase in northwestern Alberta. Can. J. For. Res. 2004, 34, 417–430. [Google Scholar] [CrossRef]
- Fenton, N.J.; Bergeron, Y. Facilitative succession in a boreal bryophyte community driven by changes in available moisture and light. J. Veg. Sci. 2006, 17, 65–76. [Google Scholar] [CrossRef]
- Thorpe, H.; Thomas, S.; Caspersen, J. Residual-tree growth responses to partial stand harvest in the black spruce (Picea mariana) boreal forest. Can. J. For. Res. 2007, 37, 1563–1571. [Google Scholar] [CrossRef]
- Thorpe, H.; Thomas, S.; Caspersen, J. Tree mortality following partial harvests is determined by skidding proximity. Ecol. Appl. 2008, 18, 1652–1663. [Google Scholar] [CrossRef]
- Riopel, M.; Bégin, J.; Ruel, J.-C. Probabilités de pertes des tiges individuelles, cinq ans après des coupes avec protection des petites tiges marchandes, dans des forêts résineuses du Québec. Can. J. For. Res. 2010, 40, 1458–1472. [Google Scholar] [CrossRef]
- Etheridge, D.A.; Kayahara, G.J. Challenges and implications of incorporating multi-cohort management in northeastern Ontario, Canada: A case study. For. Chron. 2013, 89, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Bose, A.K.; Harvey, B.D.; Brais, S.; Beaudet, M.; Leduc, A. Constraints to partial cutting in the boreal forest of Canada in the context of natural disturbance-based management: A review. Forestry 2014, 87, 11–28. [Google Scholar] [CrossRef]
- Hautala, H.; Vanha-Majamaa, I. Immediate tree uprooting after retention-felling in a coniferous boreal forest in Fennoscandia. Can. J. For. Res. 2006, 36, 3167–3172. [Google Scholar] [CrossRef]
- Dragotescu, I.; Kneeshaw, D.D. A comparison of residual forest following fires and harvesting in boreal forests in Quebec, Canada. Silva Fenn. 2012, 46, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, S.; Ruel, J.-C.; Bergeron, Y.; Harvey, B.D. Windthrow after group and dispersed tree retention in eastern Canada. For. Ecol. Manag. 2012, 269, 158–167. [Google Scholar] [CrossRef]
- Urgenson, L.S.; Halpern, C.B.; Anderson, P.D. Level and pattern of overstory retention influence rates and forms of tree mortality in mature, coniferous forests of the Pacific Northwest, USA. For. Ecol. Manag. 2013, 308, 116–127. [Google Scholar] [CrossRef]
- Montoro Girona, M.; Morin, H.; Lussier, J.-M.; Ruel, J.-C. Post-cutting mortality following experimental silvicultural treatments in unmanaged boreal forest stands. Front. For. Glob. Chang. 2019, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Moussaoui, L.; Fenton, N.J.; Leduc, A.; Bergeron, Y. Deadwood abundance in post-harvest and post-fire residual patches: An evaluation of patch temporal dynamics in black spruce boreal forest. For. Ecol. Manag. 2016, 368, 17–27. [Google Scholar] [CrossRef]
- Moussaoui, L.; Fenton, N.J.; Leduc, A.; Bergeron, Y. Can retention harvest maintain natural structural complexity? A comparison of post-harvest and post-fire residual patches in boreal forest. Forests 2016, 7, 243. [Google Scholar] [CrossRef] [Green Version]
- Bergeron, Y.; Engelmark, O.; Harvey, B.; Morin, H.; Sirois, L. Key issues in disturbance dynamics in boreal forests: Introduction. J. Veg. Sci. 1998, 9, 464–468. [Google Scholar] [CrossRef]
- Labrecque-Foy, J.-P.; Morin, H.; Girona, M.M. Dynamics of territorial occupation by North American beavers in Canadian boreal forests: A novel dendroecological approach. Forests 2020, 11, 221. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.; Girona, M.M.; Morin, H. Driving factors of conifer regeneration dynamics in eastern Canadian boreal old-growth forests. bioRxiv 2020. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.; Carleton, T.; Adams, R. Understorey vegetation change in a Picea mariana chronosequence. Vegetatio 1988, 73, 63–72. [Google Scholar] [CrossRef]
- Smith, D.M.; Larson, B.C.; Kelty, M.J.; Ashton, P.M.S. The Practice of Silviculture: Applied Forest Ecology; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 1997. [Google Scholar]
- Moussaoui, L.; Leduc, A.; Fenton, N.J.; Lafleur, B.; Bergeron, Y. Changes in forest structure along a chronosequence in the black spruce boreal forest: Identifying structures to be reproduced through silvicultural practices. Ecol. Indic. 2019, 97, 89–99. [Google Scholar] [CrossRef]
- Harper, K.A.; Bergeron, Y.; Drapeau, P.; Gauthier, S.; De Grandpré, L. Structural development following fire in black spruce boreal forest. For. Ecol. Manag. 2005, 206, 293–306. [Google Scholar] [CrossRef]
- Lecomte, N.; Simard, M.; Bergeron, Y.; Larouche, A.; Asnong, H.; Richard, P.J. Effects of fire severity and initial tree composition on understorey vegetation dynamics in a boreal landscape inferred from chronosequence and paleoecological data. J. Veg. Sci. 2005, 16, 665–674. [Google Scholar] [CrossRef]
- Smyth, C.; Schieck, J.; Boutin, S.; Wasel, S. Influence of stand size on pattern of live trees in mixedwood landscapes following wildfire. For. Chron. 2005, 81, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Brassard, B.W.; Chen, H.Y.; Wang, J.R.; Duinker, P.N. Effects of time since stand-replacing fire and overstory composition on live-tree structural diversity in the boreal forest of central Canada. Can. J. For. Res. 2008, 38, 52–62. [Google Scholar] [CrossRef]
- Martin, M.; Fenton, N.; Morin, H. Structural diversity and dynamics of boreal old-growth forests case study in Eastern Canada. For. Ecol. Manag. 2018, 422, 125–136. [Google Scholar] [CrossRef]
- Bergeron, Y.; Drapeau, P.; Gauthier, S.; Lecomte, N. Using knowledge of natural disturbances to support sustainable forest management in the northern Clay Belt. For. Chron. 2007, 83, 326–337. [Google Scholar] [CrossRef] [Green Version]
- Anderson, B.D.; Windmuller-Campione, M.A.; Russell, M.B.; Palik, B.J.; Kastendick, D.N. Short-and Long-Term Results of Alternative Silviculture in Peatland Black Spruce in Minnesota, USA. For. Sci. 2020, 66, 256–265. [Google Scholar] [CrossRef]
- Riopel, M.; Bégin, J.; Ruel, J.-C. Coefficients de distribution de la régénération, cinq ans après des coupes avec protection des petites tiges marchandes appliquées dans des sapinières et des pessières noires du Québec. For. Chron. 2011, 87, 669–683. [Google Scholar] [CrossRef] [Green Version]
- Bergeron, Y.; Fenton, N.J. Boreal forests of eastern Canada revisited: Old growth, nonfire disturbances, forest succession, and biodiversity. Botany 2012, 90, 509–523. [Google Scholar] [CrossRef]
- Lafleur, B.; Renard, S.; Leroy, C.; Fenton, N.J.; Simard, M.; Gauthier, S.; Paré, D.; Leduc, A.; Thiffault, N.; Bergeron, Y. Silviculture to sustain productivity in black spruce paludified forests. For. Ecol. Manag. 2016, 375, 172–181. [Google Scholar] [CrossRef]
- Payette, S.; Rochefort, L. Écologie des Tourbières du Québec-Labrador; Presses Université Laval: Québec, QC, Canada, 2001. [Google Scholar]
- Fenton, N.J.; Simard, M.; Bergeron, Y. Emulating natural disturbances: The role of silviculture in creating even-aged and complex structures in the black spruce boreal forest of eastern North America. J. For. Res. 2009, 14, 258–267. [Google Scholar] [CrossRef]
- Fenton, N.; Bescond, H.; Imbeau, L.; Boudreault, C.; Drapeau, P.; Bergeron, Y. Évaluation sylvicole et écologique de la coupe partielle dans la forêt boréale de la ceinture d’argile. In Aménagement Écosystémique en Forêt Boréale; Presses de l’Université du Québec: Quebec, QC, Canada, 2008; pp. 393–416. [Google Scholar]
- Pamerleau-Couture, E.; Krause, C.; Pothier, D.; Weiskittel, A. Effect of three partial cutting practices on stand structure and growth of residual black spruce trees in north-eastern Quebec. For. Int. J. For. Res. 2015, 88, 471–483. [Google Scholar] [CrossRef] [Green Version]
- Montoro Girona, M.; Morin, H.; Lussier, J.-M.; Walsh, D. Radial growth response of black spruce stands ten years after experimental shelterwoods and seed-tree cuttings in boreal forest. Forests 2016, 7, 240. [Google Scholar] [CrossRef] [Green Version]
- Montoro Girona, M.; Rossi, S.; Lussier, J.-M.; Walsh, D.; Morin, H. Understanding tree growth responses after partial cuttings: A new approach. PLoS ONE 2017, 12, e0172653. [Google Scholar] [CrossRef]
- Bescond, H.; Fenton, N.J.; Bergeron, Y. Partial harvests in the boreal forest: Response of the understory vegetation five years after harvest. For. Chron. 2011, 87, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Robitaille, A.; Saucier, J. Paysages régionaux du Québec méridional. Direction de la Gestion des Stocks Forestiers et Direction des Relations Publiques, Ministère des Ressources Naturelles du Québec; Les publications du Québec: Quebec, QC, Canada, 1998. [Google Scholar]
- Vincent, J.-S.; Hardy, L. L’évolution et l’extension des lacs glaciaires Barlow et Ojibway en territoire québécois. Géographie Phys. Quat. 1977, 31, 357–372. [Google Scholar] [CrossRef] [Green Version]
- Environment Canada. 1981–2010 Climate Normals and Averages; Government of Canada: Ottawa, ON, Canada, 2015.
- Bergeron, Y.; Gauthier, S.; Flannigan, M.; Kafka, V. Fire regimes at the transition between mixedwood and coniferous boreal forest in northwestern Quebec. Ecology 2004, 85, 1916–1932. [Google Scholar] [CrossRef]
- Imbeau, L.; St-Laurent, M.-H.; Marzell, L.; Brodeur, V. Current capacity to conduct ecologically sustainable forest management in northeastern Canada reveals challenges for conservation of biodiversity. Can. J. For. Res. 2015, 45, 567–578. [Google Scholar] [CrossRef]
- MFFP. Cartographie du 5e Inventaire Écoforestier du Québec Méridional: Méthodes et Données Associées, Ministère des Forêts, de la Faune et Des Parcs, Secteur des Forêts, Direction des Inventaires Forestiers; Gouvernement du Québec (Qué): Quebec City, QC, Canada, 2018; p. 11.
- Fenton, N.J.; Bergeron, Y. Sphagnum community change after partial harvest in black spruce boreal forests. For. Ecol. Manag. 2007, 242, 24–33. [Google Scholar] [CrossRef]
- Bergeron, Y.; Harvey, B. Basing silviculture on natural ecosystem dynamics: An approach applied to the southern boreal mixedwood forest of Quebec. For. Ecol. Manag. 1997, 92, 235–242. [Google Scholar] [CrossRef]
- Deans, A.M.; Malcolm, J.R.; Smith, S.M.; Carleton, T.J. A comparison of forest structure among old-growth, variable retention harvested, and clearcut peatland black spruce (Picea mariana) forests in boreal northeastern Ontario. For. Chron. 2003, 79, 579–589. [Google Scholar] [CrossRef] [Green Version]
- Pouliot, B.; Leblanc, M.; Méthot, S. Coupe Avec Protection des Petites Tiges Marchandes et Coupe Avec Protection des Tiges à Diamètre Variable-Méthode de Suivi des Traitements; Gouvernement du Québec, Ministère des Ressources naturelles et de la Faune, Direction de la protection des forêts et Direction de l’aménagement et de l’environnement forestiers: Quebec, QC, Canada, 2011; p. 11.
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Therneau, T.M.; Atkinson, E.J. An Introduction to Recursive Partitioning using the RPART Routines; Technical Report no. 61; Mayo Foundation: Rochester (MM), MN, USA, 1997; p. 52.1. [Google Scholar]
- De’Ath, G. Multivariate regression trees: A new technique for modeling species–environment relationships. Ecology 2002, 83, 1105–1117. [Google Scholar]
- Larsen, D.R.; Speckman, P.L. Multivariate regression trees for analysis of abundance data. Biometrics 2004, 60, 543–549. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Development Core Team, R: Vienna, Austria, 2013. [Google Scholar]
- Ruel, J.-C. Understanding windthrow: Silvicultural implications. For. Chron. 1995, 71, 434–445. [Google Scholar] [CrossRef]
- Simard, M.; Lecomte, N.; Bergeron, Y.; Bernier, P.Y.; Paré, D. Forest productivity decline caused by successional paludification of boreal soils. Ecol. Appl. 2007, 17, 1619–1637. [Google Scholar] [CrossRef] [PubMed]
- Simard, M.; Bernier, P.Y.; Bergeron, Y.; Pare, D.; Guérine, L. Paludification dynamics in the boreal forest of the James Bay Lowlands: Effect of time since fire and topography. Can. J. For. Res. 2009, 39, 546–552. [Google Scholar] [CrossRef]
- Fenton, N.; Lecomte, N.; Légaré, S.; Bergeron, Y. Paludification in black spruce (Picea mariana) forests of eastern Canada: Potential factors and management implications. For. Ecol. Manag. 2005, 213, 151–159. [Google Scholar] [CrossRef]
- Lecomte, N.; Simard, M.; Bergeron, Y. Effects of fire severity and initial tree composition on stand structural development in the coniferous boreal forest of northwestern Québec, Canada. Ecoscience 2006, 13, 152–163. [Google Scholar] [CrossRef]
- Lafleur, B.; Fenton, N.; Paré, D.; Simard, M.; Bergeron, Y. Contrasting effects of season and method of harvest on soil properties and the growth of black spruce regeneration in the boreal forested peatlands of eastern Canada. Silva Fenn. 2010, 44, 799–813. [Google Scholar] [CrossRef] [Green Version]
- Lafleur, B.; Cazal, A.; Leduc, A.; Bergeron, Y. Soil organic layer thickness influences the establishment and growth of trembling aspen (Populus tremuloides) in boreal forests. For. Ecol. Manag. 2015, 347, 209–216. [Google Scholar] [CrossRef]
- Gewehr, S.; Drobyshev, I.; Berninger, F.; Bergeron, Y. Soil characteristics mediate the distribution and response of boreal trees to climatic variability. Can. J. For. Res. 2014, 44, 487–498. [Google Scholar] [CrossRef] [Green Version]
- Greene, D.F.; Macdonald, S.E.; Cumming, S.; Swift, L. Seedbed variation from the interior through the edge of a large wildfire in Alberta. Can. J. For. Res. 2005, 35, 1640–1647. [Google Scholar] [CrossRef]
- Laamrani, A.; Valeria, O.; Bergeron, Y.; Fenton, N.; Cheng, L.Z.; Anyomi, K. Effects of topography and thickness of organic layer on productivity of black spruce boreal forests of the Canadian Clay Belt region. For. Ecol. Manag. 2014, 330, 144–157. [Google Scholar] [CrossRef]
- Messier, C.; Doucet, R.; Ruel, J.-C.; Claveau, Y.; Kelly, C.; Lechowicz, M.J. Functional ecology of advance regeneration in relation to light in boreal forests. Can. J. For. Res. 1999, 29, 812–823. [Google Scholar] [CrossRef]
- Groot, A.; Hökkä, H. Persistence of suppression effects on peatland black spruce advance regeneration after overstory removal. Can. J. For. Res. 2000, 30, 753–760. [Google Scholar] [CrossRef]
- Kneeshaw, D.D.; Williams, H.; Nikinmaa, E.; Messier, C. Patterns of above-and below-ground response of understory conifer release 6 years after partial cutting. Can. J. For. Res. 2002, 32, 255–265. [Google Scholar] [CrossRef] [Green Version]
- MacIsaac, D.; Krygier, R. Development and long-term evaluation of harvesting patterns to reduce windthrow risk of understorey spruce in aspen–white spruce mixedwood stands in Alberta, Canada. Forestry 2009, 82, 323–342. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, B.; Stacey, G.; Belcher, R.; Wood, C. Field and wind tunnel assessments of the implications of respacing and thinning for tree stability. For. Int. J. For. Res. 1997, 70, 233–252. [Google Scholar] [CrossRef]
- Rich, R.L.; Frelich, L.E.; Reich, P.B. Wind-throw mortality in the southern boreal forest: Effects of species, diameter and stand age. J. Ecol. 2007, 95, 1261–1273. [Google Scholar] [CrossRef]
- Savill, P.; PS, S. Silviculture in Windy Climates. For. Abstr. 1983, 44, 443–488. [Google Scholar]
- Gardiner, B.; Marshall, B.; Achim, A.; Belcher, R.; Wood, C. The stability of different silvicultural systems: A wind-tunnel investigation. Forestry 2005, 78, 471–484. [Google Scholar] [CrossRef]
- Jönsson, M.T.; Fraver, S.; Jonsson, B.G.; Dynesius, M.; Rydgård, M.; Esseen, P.-A. Eighteen years of tree mortality and structural change in an experimentally fragmented Norway spruce forest. For. Ecol. Manag. 2007, 242, 306–313. [Google Scholar] [CrossRef]
- Steventon, D. Retention Patches: Windthrow and Recruitment of Habitat Structure 12–16 Years Post-Harvest. J. Ecosyst. Manag. 2011, 11, 3. [Google Scholar]
- Anyomi, K.A.; Ruel, J.-C. A multiscale analysis of the effects of alternative silvicultural treatments on windthrow within balsam fir dominated stands. Can. J. For. Res. 2015, 45, 1739–1747. [Google Scholar] [CrossRef] [Green Version]
- Roy Proulx, S.; Jutras, S.; Leduc, A.; Mazerolle, M.J.; Fenton, N.J.; Bergeron, Y. Partial harvest in paludified black spruce: Effects on water table and variation in stem diameter. Can. J. For. Res. in review.
- Mäkinen, H.; Nöjd, P.; Isomäki, A. Radial, height and volume increment variation in Picea abies (L.) Karst. stands with varying thinning intensities. Scand. J. For. Res. 2002, 17, 304–316. [Google Scholar] [CrossRef]
- Vincent, M.; Krause, C.; Zhang, S. Radial growth response of black spruce roots and stems to commercial thinning in the boreal forest. Forestry 2009, 82, 557–571. [Google Scholar] [CrossRef] [Green Version]
- Goudiaby, V.; Brais, S.; Berninger, F.; Schneider, R. Vertical patterns in specific volume increment along stems of dominant jack pine (Pinus banksiana) and black spruce (Picea mariana) after thinning. Can. J. For. Res. 2012, 42, 733–748. [Google Scholar] [CrossRef]
- Soucy, M.; Lussier, J.-M.; Lavoie, L. Long-term effects of thinning on growth and yield of an upland black spruce stand. Can. J. For. Res. 2012, 42, 1669–1677. [Google Scholar] [CrossRef]


| Site Name | Muskuchii | Maïcasagi * | Gaudet | Fénélon | Puiseaux |
|---|---|---|---|---|---|
| Site level | |||||
| Surficial deposit | Glaciofluvial | Glaciolacustrine—Cochrane till | Cochrane till | Cochrane till | Cochrane till |
| Drainage | dry–mesic | mesic-sub-hydric | sub-hydric | sub-hydric | sub-hydric |
| Soil texture | Sand | clay-thin organic | Clay | Clay | Clay |
| Silvicultural level | |||||
| Treatment type (n) | HARP (42) | CCCC (16) | CCCC (12) | CCCC (13) | CCCC (10) |
| Harvesting intensity, i.e., percentage of harvested basal area (%, TI) | 62.2 (3.3) | ~45 | 74.6 (4.8) | 73.5 (5.7) | 68.8 (2.9) |
| Initial measurements at stand level | |||||
| Organic layer thickness (cm, OLT) | 22.1 (1.3) | 40.5 (2.5) | 54.4 (3.5) | 30.5 (3.1) | 50.4 (6.8) |
| Initial basal area (m2ha−1, IBA) | 21.7 (1.3) | - | 14.1 (1.6) | 20.6 (2.6) | 21.0 (3.0) |
| Initial tree density (trees ha−1, ITD) | 1147.6 (64.4) | - | 960.4 (74.4) | 922.5 (128.8) | 1141.7 (186.0) |
| Initial mean diameter at breast height (mm, DBHi) | 150.9 (4.7) | - | 132.1 (2.4) | 159.6 (5.1) | 154.3 (21.4) |
| Post-harvest measurements at stand level in spring of the year following harvest | |||||
| Residual basal area (m2ha−1, RBA) | 7.8 (1.3) | 8.1 (1.2) | 3.2 (0.4) | 5.5 (1.3) | 5.8 (0.6) |
| Residual tree density (trees ha−1, RTD) | 714.9 (86.9) | 420.3 (52.5) | 302.1 (36.2) | 360 (65.4) | 448.1 (62.2) |
| Mean diameter at breast height (mm, DBHr) | 127.8 (3.2) | 152.8 (5.7) | 114.8 (2.0) | 130.3 (8.0) | 130.1 (11.4) |
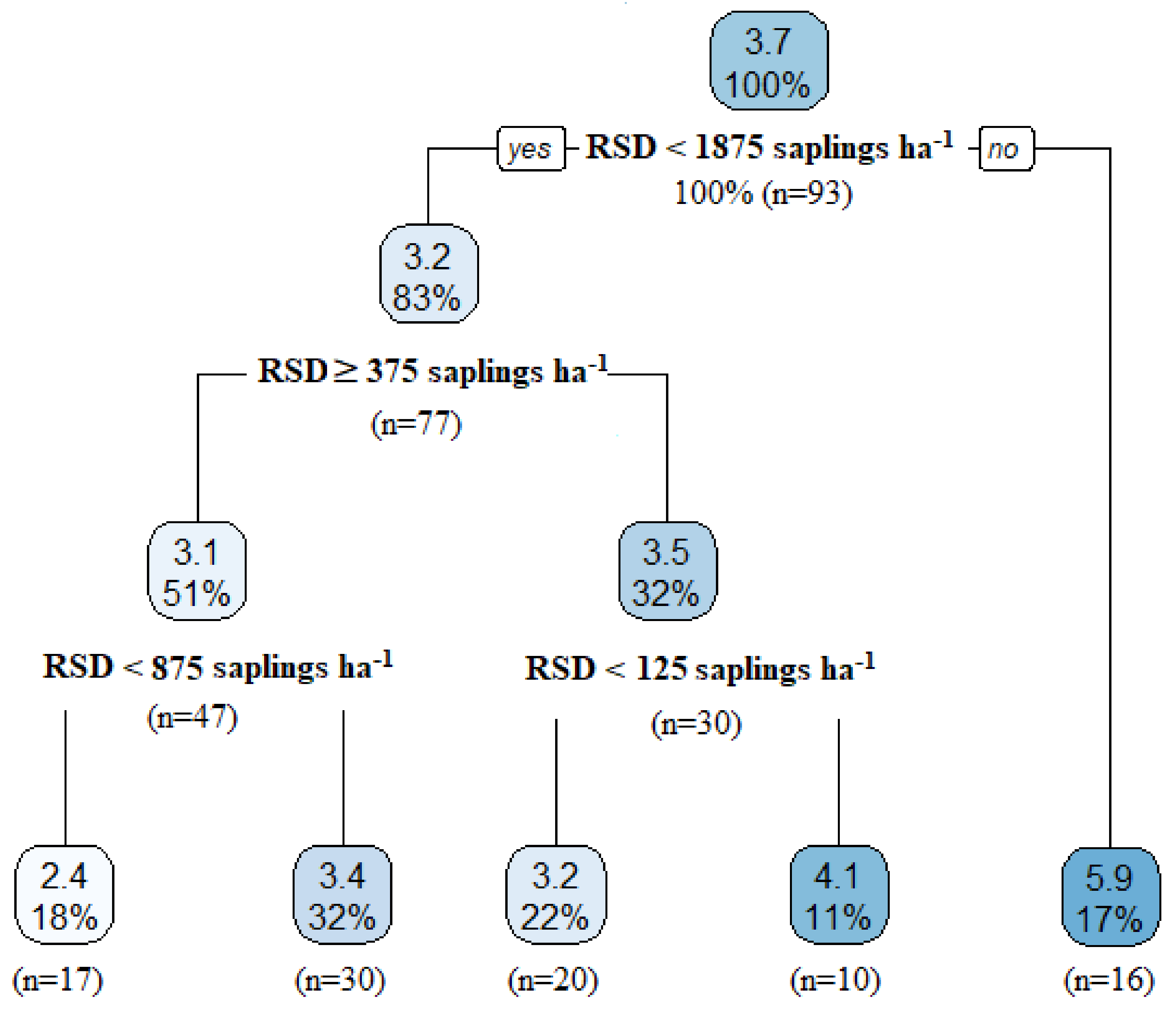
| Sapling density (saplings ha−1, RSD) | 18,182.7 (192.8) | 1265.6 (465.7) | 625 (158.4) | 3700 (1028.6) | 711.5 (246.5) |
| Variable (Abbreviation, Unit) | Description | Study Sites | ||||
|---|---|---|---|---|---|---|
| Muskuchii | Maïcasagi | Gaudet | Fénélon | Puiseaux | ||
| Residual tree growth (RTG, cm2 yrs−1 decade−1) | Average annual radial growth of trees >9 cm DBH ten years after harvest | 3.8 (0.33) | 2.2 (0.4) | 4.2 (0.9) | 7.2 (3.1) | 2.1 (0.4) |
| Residual tree mortality (RTM, m2 ha−1 decade−1) | Basal area of dead residual trees ten years after harvest | 2.7 (0.4) | 3.3 (0.6) | 1.2 (0.2) | 1.5 (0.4) | 3.5 (0.5) |
| Merchantable tree recruitment (RTR, trees ha−1 decade−1) | Density of living stems that reached 9 cm DBH in the ten years following the harvest | 129.2 (16.5) | 51.6 (21.9) | 116.7 (22.5) | 220 (51.1) | 11.5 (4.6) |
| Decennial stand yield (DTY, m2 h−1) | Net gain in basal area ten years after harvesting compared to residual basal area at the remeasurement before the first growing season of the year following harvest | +0.25 (0.22) | −2.65 (0.95) | +0.67 (0.17) | +1.64 (1.33) | −1.60 (0.61) |
| Predictors | Estimates | CI | p |
|---|---|---|---|
| (Intercept) | −0.90 | −1.62–−0.18 | 0.014 |
| Merchantable tree recruitment | 0.61 | 0.31–0.91 | <0.001 |
| Residual tree mortality | −0.37 | −0.49–0.26 | <0.001 |
| Residual tree growth | 0.26 | 0.16–0.16 | <0.001 |
| Predictors | Estimates | CI | p |
|---|---|---|---|
| (Intercept) | 24.53 | 18.10–10.96 | <0.001 |
| Organic layer thickness | −0.20 | −0.33–−0.08 | 0.001 |
| Residual tree basal area | −0.35 | −0.56–−0.14 | 0.001 |
| Sapling density | 0.02 | 0.01–0.03 | <0.001 |
| Residual diameter at breast height | −0.05 | −0.09–−0.02 | 0.003 |
| Harvesting intensity | −0.13 | −0.18–−0.08 | <0.001 |
| Predictors | Estimates | CI | p |
|---|---|---|---|
| (Intercept) | −5.40 | −9.04–−1.76 | 0.004 |
| Organic layer thickness | 0.02 | −0.03–0.08 | 0.374 |
| Residual tree basal area | 0.30 | 0.17–0.43 | <0.001 |
| Sapling density | −0.01 | −0.01–−0.00 | 0.036 |
| Residual diameter at breast height | 0.03 | 0.01–0.05 | 0.013 |
| Harvesting intensity | 0.04 | 0.00–0.07 | 0.036 |
| Predictors | Estimates | CI | p |
|---|---|---|---|
| (Intercept) | 0.45 | −4.35–−5.25 | 0.854 |
| Organic layer thickness | −0.03 | −0.12–0.07 | 0.571 |
| Residual tree basal area | 0.08 | −0.07–0.23 | 0.301 |
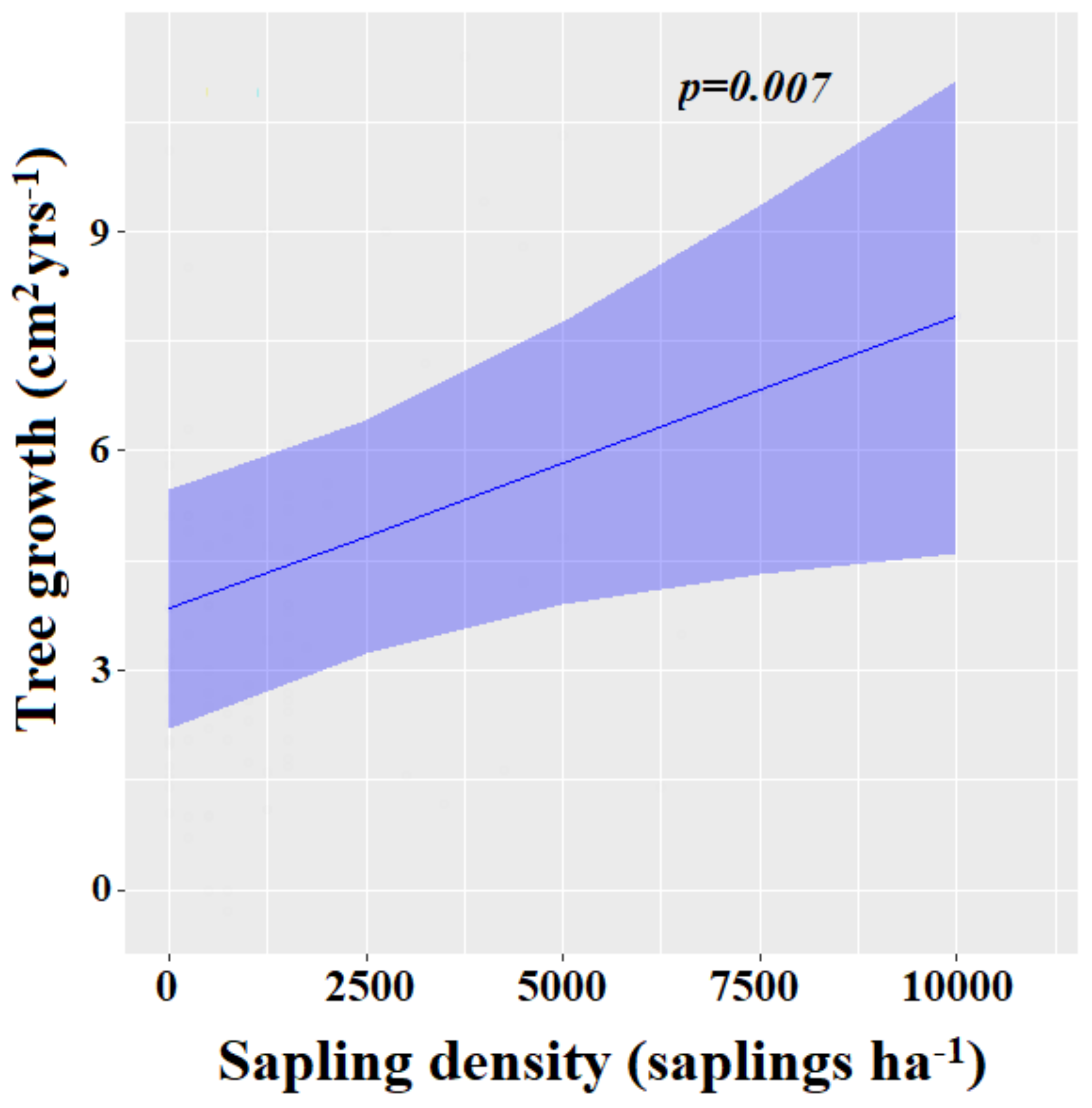
| Sapling density | 0.01 | 0.00–0.02 | 0.007 |
| Residual diameter at breast height | 0.01 | −0.02–0.03 | 0.695 |
| Harvesting intensity | 0.03 | −0.00 – 0.07 | 0.075 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moussaoui, L.; Leduc, A.; Girona, M.M.; Bélisle, A.C.; Lafleur, B.; Fenton, N.J.; Bergeron, Y. Success Factors for Experimental Partial Harvesting in Unmanaged Boreal Forest: 10-Year Stand Yield Results. Forests 2020, 11, 1199. https://doi.org/10.3390/f11111199
Moussaoui L, Leduc A, Girona MM, Bélisle AC, Lafleur B, Fenton NJ, Bergeron Y. Success Factors for Experimental Partial Harvesting in Unmanaged Boreal Forest: 10-Year Stand Yield Results. Forests. 2020; 11(11):1199. https://doi.org/10.3390/f11111199
Chicago/Turabian StyleMoussaoui, Louiza, Alain Leduc, Miguel Montoro Girona, Annie Claude Bélisle, Benoit Lafleur, Nicole J. Fenton, and Yves Bergeron. 2020. "Success Factors for Experimental Partial Harvesting in Unmanaged Boreal Forest: 10-Year Stand Yield Results" Forests 11, no. 11: 1199. https://doi.org/10.3390/f11111199
APA StyleMoussaoui, L., Leduc, A., Girona, M. M., Bélisle, A. C., Lafleur, B., Fenton, N. J., & Bergeron, Y. (2020). Success Factors for Experimental Partial Harvesting in Unmanaged Boreal Forest: 10-Year Stand Yield Results. Forests, 11(11), 1199. https://doi.org/10.3390/f11111199