Discovering Douglas-Fir Woodlands in the Historical Forests of Umatilla National Forest, Eastern Oregon and Washington


Abstract
1. Introduction
2. Materials and Methods
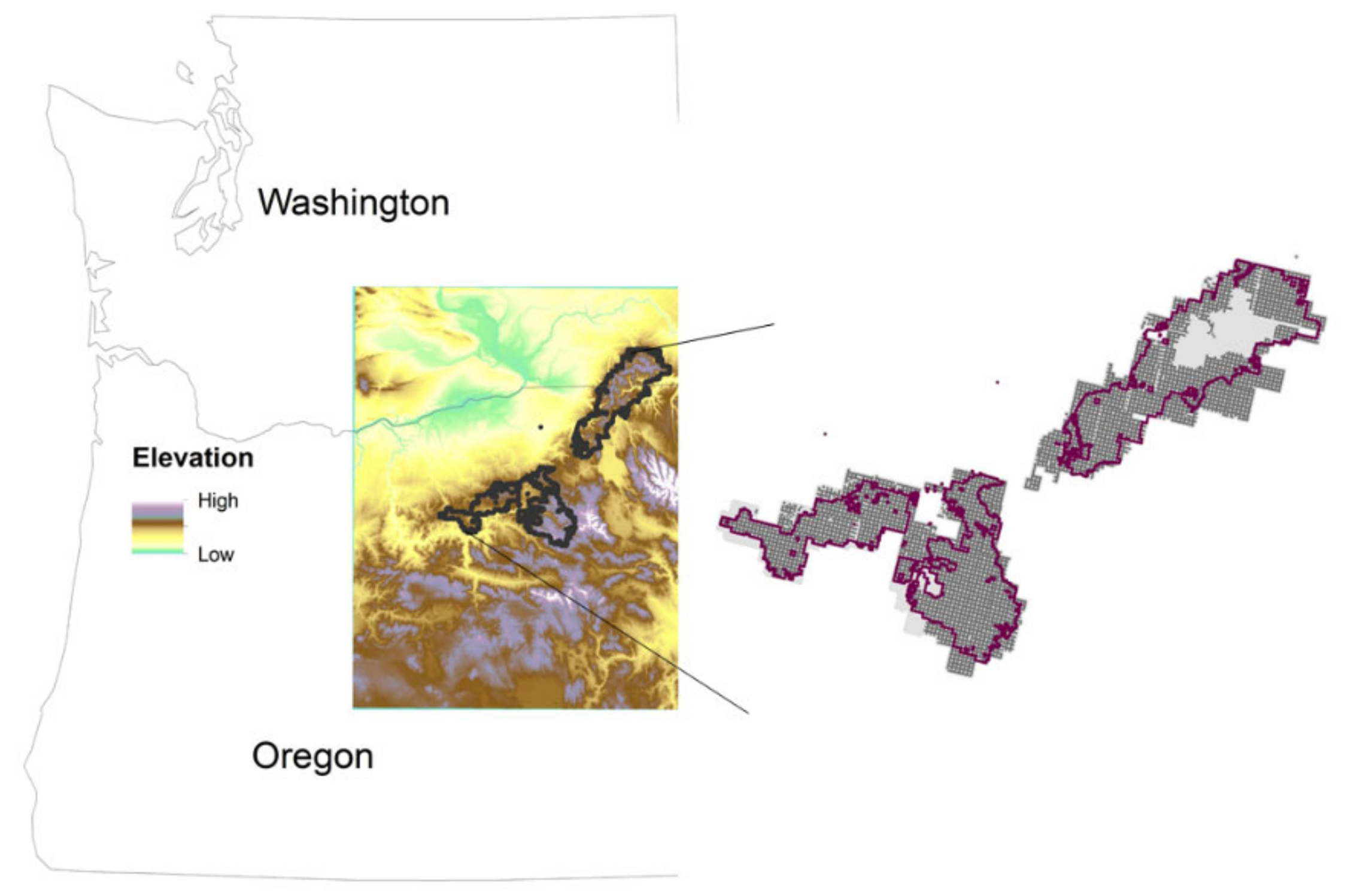
2.1. Study Area
2.2. Historical Surveys and Composition
2.3. Models of Identified and Unidentified Firs and Pines
2.4. Current Surveys and Composition
2.5. Similarity Metric
2.6. Historical and Current Tree Densities
3. Results
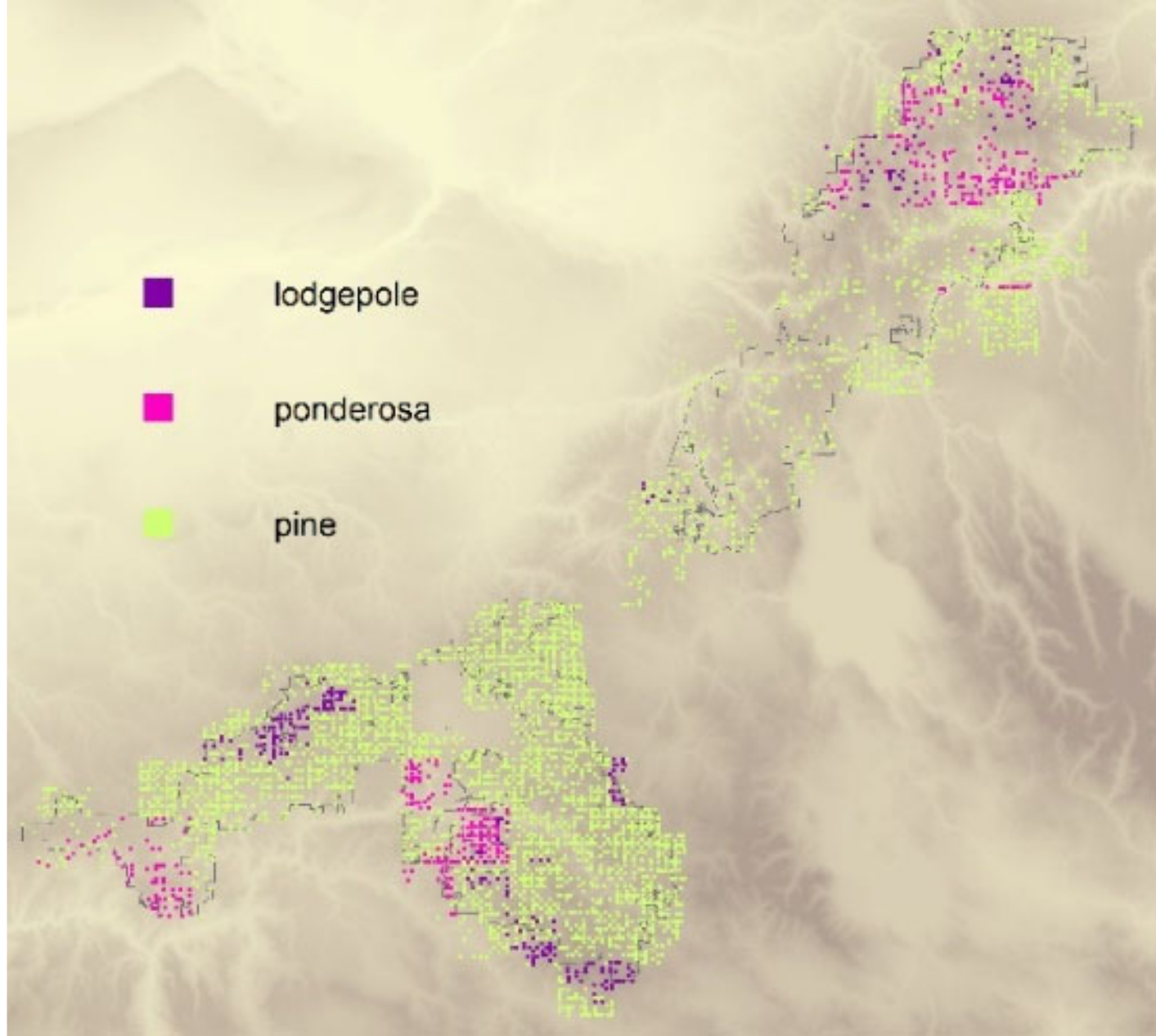
3.1. Historical Composition and Similarity
3.2. Models of Identified and Unidentified Firs and Pines
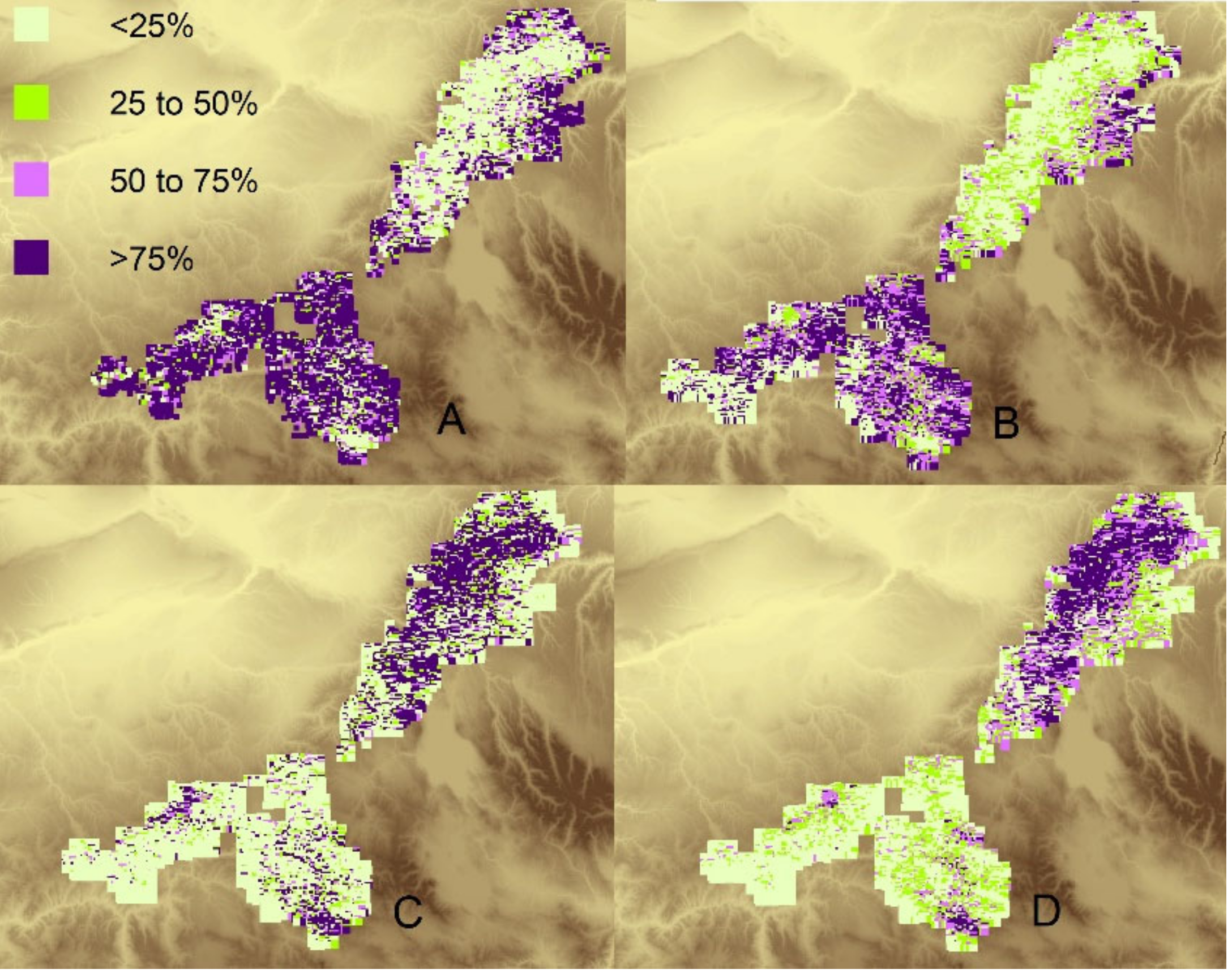
3.3. Comparison of Historical and Current Composition and Similarity
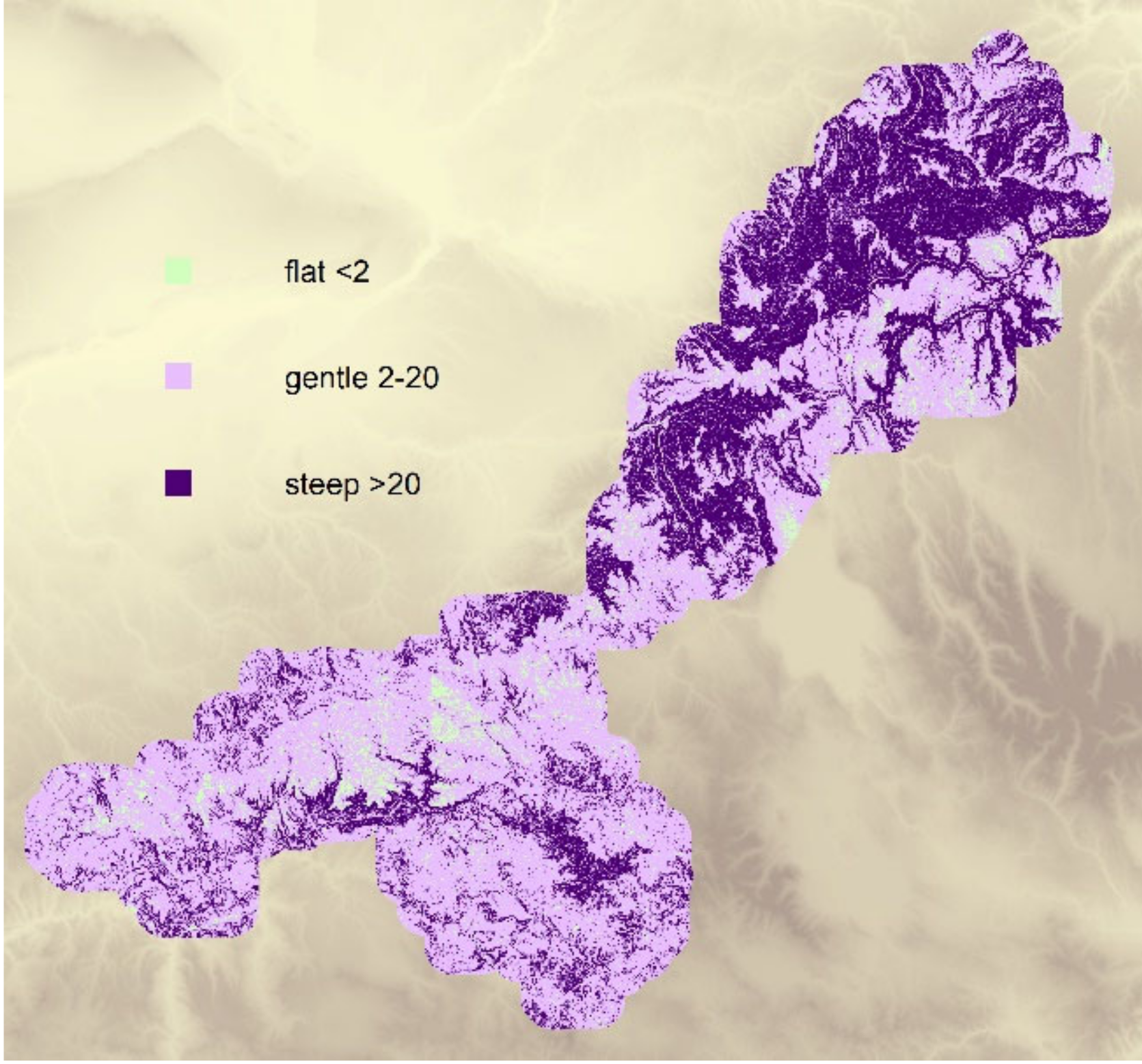
3.4. Comparison of Historical and Current Density
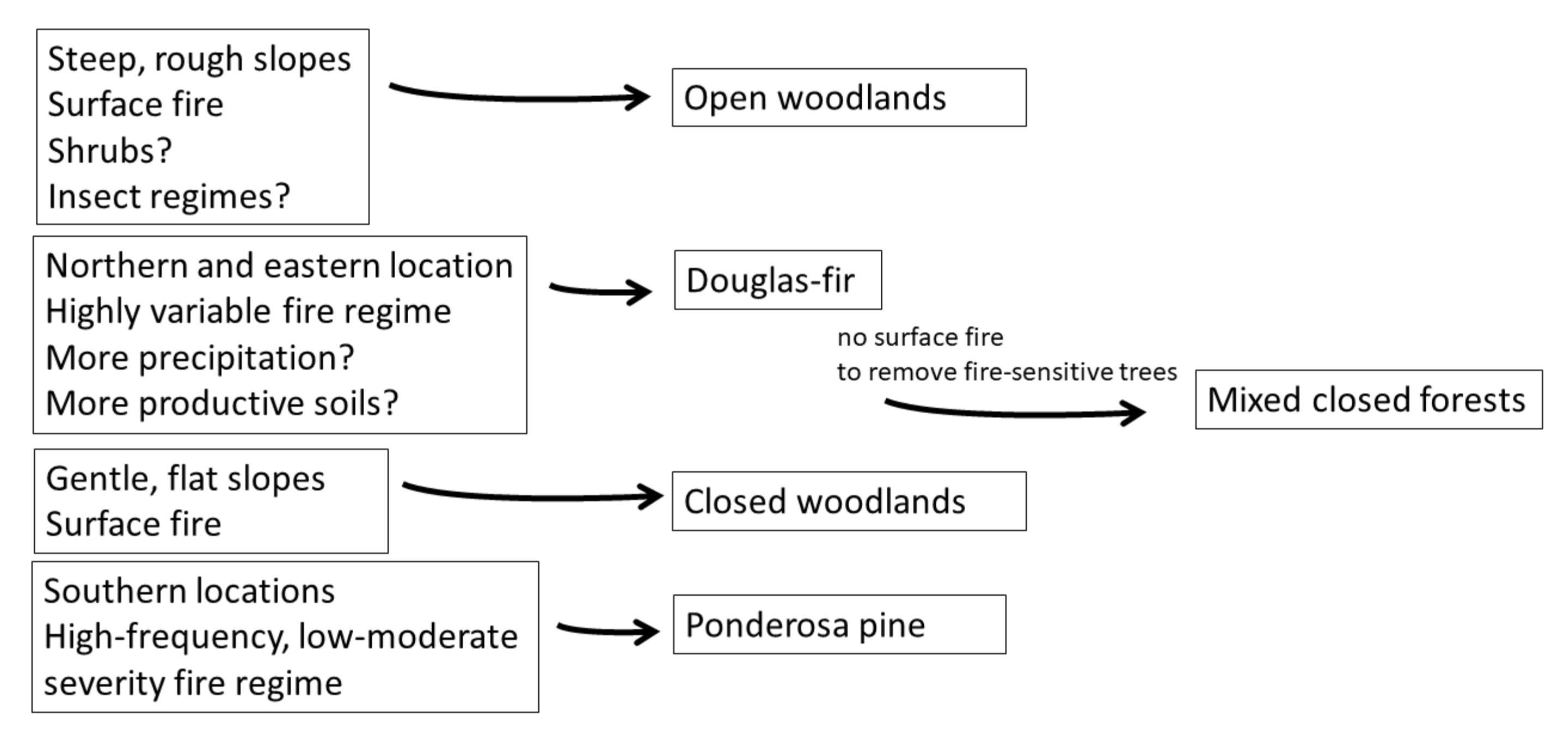
4. Discussion
The Information Deficit and Historical Tree Surveys
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weaver, H. Fire as an ecological and silvicultural factor in the ponderosa-pine region of the Pacific slope. J. For. 1943, 41, 7–14. [Google Scholar] [CrossRef]
- Kilgore, B.M. The ecological role of fire in Sierran conifer forests: Its application to national park management. Quat. Res. 1973, 3, 496–513. [Google Scholar] [CrossRef]
- Dickman, A. Reduced fire frequency changes species composition of a ponderosa pine stand. J. For. 1978, 76, 24–25. [Google Scholar]
- Parsons, D.J.; DeBenedetti, S.H. Impact of fire suppression on a mixed-conifer forest. For. Ecol. Manag. 1979, 2, 21–33. [Google Scholar] [CrossRef]
- Steele, R.; Arno, S.F.; Geier-Hayes, K. Wildfire patterns change in central Idaho’s ponderosa pine–Douglas-fir forest. West. J. Appl. For. 1986, 1, 16–18. [Google Scholar] [CrossRef]
- Mutch, R.W.; Arno, S.F.; Brown, J.K.; Carlson, C.E.; Ottmar, R.D.; Peterson, J.L. Forest Health in the Blue Mountains: A Management Strategy for Fire-Adapted Ecosystems; USDA Forest Service, Pacific Northwest Research Station: Corvallis, OR, USA, 1993. [Google Scholar]
- Sloan, J.P. Historical Density and Stand Structure of an Old-Growth Forest in the Boise Basin of Central Idaho; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 1998. [Google Scholar]
- Sloan, J.P. Interruption of the Natural Fire Cycle in a Grand Fir Forest of Central Idaho: Changes in Stand Structure and Composition; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 1998. [Google Scholar]
- Youngblood, A.; Max, T.; Coe, K. Stand structure in eastside old-growth ponderosa pine forests of Oregon and northern California. For. Ecol. Manag. 2004, 199, 191–217. [Google Scholar] [CrossRef]
- Keeling, E.G.; Sala, A.; DeLuca, T.H. Effects of fire exclusion on forest structure and composition in unlogged ponderosa pine/Douglas-fir forests. For. Ecol. Manag. 2006, 237, 418–428. [Google Scholar] [CrossRef]
- Knapp, E.E.; Skinner, C.N.; North, M.P.; Estes, B.L. Long-term overstory and understory change following logging and fire exclusion in a Sierra Nevada mixed-conifer forest. For. Ecol. Manag. 2013, 310, 903–914. [Google Scholar] [CrossRef]
- Reynolds, R.T.; Sanchez Meador, A.J.; Youtz, J.A.; Nicolet, T.; Matonis, M.S.; Jackson, P.L.; DeLorenzo, D.G.; Graves, A.D. Restoring Composition and Structure in Southwestern Frequent-Fire Forests: A Science-Based Framework for Improving Ecosystem Resiliency; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 2013. [Google Scholar]
- Powell, D.C. Active Management of Dry Forests in the Blue Mountains: Silvicultural Considerations; USDA Forest Service, Pacific Northwest Research Station: Corvallis, OR, USA, 2014. [Google Scholar]
- Stephens, S.L.; Lydersen, J.M.; Collins, B.M.; Fry, D.L.; Meyer, M.D. Historical and current landscape-scale ponderosa pine and mixed conifer forest structure in the Southern Sierra Nevada. Ecosphere 2015, 6, 79. [Google Scholar] [CrossRef]
- Kolb, T.E.; Agee, J.K.; Fule, P.Z.; McDowell, N.G.; Pearson, K.; Sala, A.; Waring, R.H. Perpetuating old ponderosa pine. For. Ecol. Manag. 2007, 249, 141–157. [Google Scholar] [CrossRef]
- Martinson, E.J.; Omi, P.N. Fuel Treatments and Fire Severity: A Meta-Analysis; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 2013. [Google Scholar]
- Stine, P.; Hessburg, P.; Spies, T.; Kramer, M.; Fettig, C.J.; Hansen, A.; Lehmkuhl, J.; O’Hara, K.; Polivka, K.; Singleton, P.; et al. The Ecology and Management of Moist Mixed-Conifer Forests in Eastern Oregon and Washington: A Synthesis of the Relevant Biophysical Science and Implications for Future Land Management; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 2014. [Google Scholar]
- Davis, K.T.; Dobrowski, S.Z.; Higuera, P.E.; Holden, Z.A.; Veblen, T.T.; Rother, M.T.; Parks, S.A.; Sala, A.; Maneta, M.P. Wildfires and climate change push low-elevation forests across a critical climate threshold for tree regeneration. Proc. Natl. Acad. Sci. USA 2019, 116, 6193–6198. [Google Scholar] [CrossRef] [PubMed]
- Downing, W.M.; Krawchuk, M.A.; Meigs, G.W.; Haire, S.L.; Coop, J.D.; Walker, R.B.; Whitman, E.; Chong, G.; Miller, C. Influence of fire refugia spatial pattern on post-fire forest recovery in Oregon’s Blue Mountains. Landsc. Ecol. 2019, 34, 771–792. [Google Scholar] [CrossRef]
- Hankin, L.E.; Higuera, P.E.; Davis, K.T.; Dobrowski, S.Z. Impacts of growing-season climate on tree growth and post-fire regeneration in ponderosa pine and Douglas-fir forests. Ecosphere 2019, 10, e02679. [Google Scholar] [CrossRef]
- Korb, J.E.; Fornwalt, P.J.; Stevens-Rumann, C.S. What drives ponderosa pine regeneration following wildfire in the western United States? For. Ecol. Manag. 2019, 454, 117663. [Google Scholar] [CrossRef]
- Boag, A.E.; Ducey, M.J.; Palace, M.W.; Hartter, J. Topography and fire legacies drive variable post-fire juvenile conifer regeneration in eastern Oregon, USA. For. Ecol. Manag. 2020, 474, 118312. [Google Scholar] [CrossRef]
- Hanula, J.L.; Ulyshen, M.D.; Horn, S. Conserving pollinators in North American forests: A review. Nat. Areas J. 2016, 36, 427–439. [Google Scholar] [CrossRef]
- Quigley, T.M.; Arbelbide, S.J. An Assessment of Ecosystem Components in the Interior Columbia Basin and Portions of the Klamath and Great Basins; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 1997. [Google Scholar]
- Wisdom, M.J.; Holthausen, R.S.; Wales, B.C.; Hargis, C.D.; Saab, V.A.; Lee, D.C.; Hann, W.J.; Rich, T.D.; Rowland, M.M.; Murphy, W.J.; et al. Source Habitats for Terrestrial Vertebrates of Focus in the Interior Columbia Basin: Broadscale Trends and Management Implications; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 2000. [Google Scholar]
- Bunnell, F.L. Sustaining cavity-using species: Patterns of cavity use and implications to forest management. ISRN For. 2013, 457698. [Google Scholar] [CrossRef]
- Irwin, L.L.; Riggs, R.A.; Verschuyl, J.P. Reconciling wildlife conservation to forest restoration in moist mixed-conifer forests of the inland northwest: A synthesis. For. Ecol. Manag. 2018, 424, 288–311. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Dumroese, R.K. Biodiversity and representative species in dry pine forests. In Northeastern California Plateaus Bioregion Science Synthesis to Support Forest Plan Development by the Lassen National Forest and the Modoc National Forest; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 2020. [Google Scholar]
- Hall, F.C. Fire History—Blue Mountains, Oregon; USDA Forest Service, Pacific Northwest Research Station: Corvallis, OR, USA, 1980; pp. 75–81. Available online: https://www.fs.usda.gov/treesearch/pubs/41408 (accessed on 20 October 2020).
- Maruoka, K.R. Fire History of Pseudotsuga menziesii and Abies grandis Stands in the Blue Mountains of Oregon and Washington. Master’s Thesis, University of Washington, Seattle, WA, USA, 1994. [Google Scholar]
- Heyerdahl, E.K.; Brubaker, L.B.; Agee, J.K. Spatial controls of historical fire regimes: A multiscale example from the interior west, USA. Ecology 2001, 82, 660–678. [Google Scholar] [CrossRef]
- Johnston, J.D.; Bailey, J.D.; Dunn, C.J. Influence of fire disturbance and biophysical heterogeneity on pre-settlement ponderosa pine and mixed conifer forests. Ecosphere 2016, 7, e01581. [Google Scholar] [CrossRef]
- Arno, S.F. Fire in western forest ecosystems. In Wildland Fire in Ecosystems: Effects of Fire on Flora; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 2000. [Google Scholar]
- Hessburg, P.F.; Spies, T.A.; Perry, D.A.; Skinner, C.N.; Taylor, A.H.; Brown, P.M.; Stephens, S.L.; Larson, A.J.; Churchill, D.J.; Povak, N.A. Tamm review: Management of mixed-severity fire regime forests in Oregon, Washington, and Northern California. For. Ecol. Manag. 2016, 366, 221–250. [Google Scholar] [CrossRef]
- Beckham, S.D. The Grande Ronde Valley and Blue Mountains: Impressions and Experiences of Travelers and Emigrants, the Oregon Trail, 1812–1880; Beckham and Associates: Lake Oswego, OR, USA, 1991. Available online: https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fseprd482690.pdf (accessed on 20 October 2020).
- Evans, J.W. Powerful Rockey: The Blue Mountains and the Oregon Trail, 1811–1883, 1st ed.; Eastern Oregon State College: La Grande, OR, USA, 1991. [Google Scholar]
- Fremont, J.C. Report of the Exploring Expedition to the Rocky Mountains in the Year 1842, and to Oregon and North California in the Years 1843–1844; Gales and Seaton, Printers: Washington, DC, USA, 1845; Available online: https://archive.org/download/mobot31753002784947/mobot31753002784947.pdf (accessed on 20 October 2020).
- Robbins, W.G.; Wolf, D.W. Landscape and the Intermontane Northwest: An Environmental History; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 1994. [Google Scholar]
- Munger, T.T. Western Yellow Pine in Oregon; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 1917. [Google Scholar]
- Shinn, D.A. Historical perspectives on range burning in the inland Pacific Northwest. Rangel. Ecol. Manag. J. Range Manag. Arch. 1980, 33, 415–423. [Google Scholar] [CrossRef]
- Kay, C.E. Aboriginal overkill. Hum. Nat. 1994, 5, 359–398. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.W. Early fire use in Oregon. Fire Manag. 2000, 60, 13–20. [Google Scholar]
- Scharf, E.A. A statistical evaluation of the relative influences of climate, vegetation, and prehistoric human population on the charcoal record of Five Lakes, Washington (USA). Quat. Int. 2010, 215, 4–86. [Google Scholar] [CrossRef]
- Show, S.B.; Kotok, E.I. The Role of Fire in the California Pine Forests; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 1924. [Google Scholar]
- Office of Policy Analysis. Wildland Fire Management Program Benefit-Cost Analysis: A Review of Relevant Literature. Available online: https://edit.doi.gov/sites/doi.gov/files/migrated/ppa/upload/Wildland_fire_literature_review_060812FINAL.pdf (accessed on 2 February 2020).
- Long, J.W.; Tarnay, L.W.; North, M.P. Aligning smoke management with ecological and public health goals. J. For. 2018, 116, 76–86. [Google Scholar] [CrossRef]
- Johnston, J.D.; Dunn, C.J.; Vernon, M.J.; Bailey, J.D.; Morrissette, B.A.; Morici, K.E. Restoring historical forest conditions in a diverse inland Pacific Northwest landscape. Ecosphere 2018, 9, e02400. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Jones-Farrand, D.T.; Kabrick, J.M. Historical open forest ecosystems in the Missouri Ozarks: Reconstruction and restoration targets. Ecol. Restor. 2014, 32, 407–416. [Google Scholar] [CrossRef]
- Williams, M.A.; Baker, W.L. Spatially extensive reconstructions show variable-severity fire and heterogeneous structure in historical western United States dry forests. Glob. Ecol. Biogeogr. 2012, 21, 1042–1052. [Google Scholar] [CrossRef]
- Baker, W.L. Implications of spatially extensive historical data from surveys for restoring dry forests of Oregon’s eastern Cascades. Ecosphere 2012, 3, 1–39. [Google Scholar] [CrossRef]
- Levine, C.R.; Cogbill, C.V.; Collins, B.M.; Larson, A.J.; Lutz, J.A.; North, M.P.; Restaino, C.M.; Safford, H.D.; Stephens, S.L.; Battles, J.J. Evaluating a new method for reconstructing forest conditions from General Land Office survey records. Ecol. Appl. 2017, 27, 1498–1513. [Google Scholar] [CrossRef] [PubMed]
- Hagmann, R.K.; Franklin, J.F.; Johnson, K.N. Historical conditions in mixed-conifer forests on the eastern slopes of the northern Oregon Cascade Range, USA. For. Ecol. Manag. 2014, 330, 158–170. [Google Scholar] [CrossRef]
- Heyerdahl, E.K.; Loehman, R.A.; Falk, D.A. A multi-century history of fire regimes along a transect of mixed-conifer forests in central Oregon, USA. Can. J. For. Res. 2019, 49, 76–86. [Google Scholar] [CrossRef]
- Foster, H.D. Report on the Silvics of the Wenaha Forest Reserve Washington and Oregon. Available online: https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fsbdev7_015540.pdf (accessed on 26 September 2020).
- Foster, H.D. Report on the Silvics of the Blue Mountains (E) National Forest, Oregon. Available online: https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fsbdev7_015620.pdf (accessed on 26 September 2020).
- Camp, A.; Oliver, C.; Hessburg, P.; Everett, R. Predicting late-successional fire refugia pre-dating European settlement in the Wenatchee Mountains. For. Ecol. Manag. 1997, 95, 63–77. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Fraver, S.; He, H.S.; Yang, J.; Dey, D.C.; Palik, B.J. Spatial pattern corrections and sample sizes for forest density estimates of historical tree surveys. Landsc. Ecol. 2011, 26, 59–68. [Google Scholar] [CrossRef]
- Cogbill, C.V.; Thurman, A.L.; Williams, J.W.; Zhu, J.; Mladenoff, D.J.; Goring, S.J. A retrospective on the accuracy and precision of plotless forest density estimators in ecological studies. Ecosphere 2018, 9, e02187. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Yang, J.; Kabrick, J.M.; He, H.S. Adjusting forest density estimates for surveyor bias in historical tree surveys. Am. Midl. Nat. 2012, 167, 285–306. [Google Scholar] [CrossRef]
- Powell, D.C. Using General Land Office Survey Notes to Characterize Historical Vegetation Conditions for the Umatilla National Forest; USDA Forest Service, Pacific Northwest Research Station: Corvallis, OR, USA, 2008. [Google Scholar]
- Hanberry, B.B.; Coursey, K.; Kush, J.S. Structure and composition of historical longleaf pine ecosystems in Mississippi, USA. Hum. Ecol. 2018, 46, 241–248. [Google Scholar] [CrossRef]
- Evans, J.S.; Oakleaf, J.; Cushman, S.A.; Theobald, D. An ArcGIS Toolbox for Surface Gradient and Geomorphometric Modeling, Version 2.0-0. Available online: http://evansmurphy.wix.com/evansspatial (accessed on 19 September 2020).
- PRISM Climate Group. 30-Year Normals. Available online: http://prism.oregonstate.edu (accessed on 20 September 2020).
- Fernández-Delgado, M.; Cernadas, E.; Barro, S.; Amorim, D. Do we need hundreds of classifiers to solve real world classification problems? J. Mach. Learn. Res. 2014, 15, 3133–3181. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Kuhn, M. Building Predictive Models in R Using the caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef]
- Hanberry, B.B.; He, H.S. Prevalence, statistical thresholds, and accuracy assessment for species distribution models. Web Ecol. 2013, 13, 13–19. [Google Scholar] [CrossRef]
- Christensen, G.A.; Dunham, P.; Powell, D.C.; Hiserote, B. Forest Resources of the Umatilla National Forest; US Department of Agriculture, Forest Service, Intermountain Research Station: Fort Collins, CO, USA, 2007. [Google Scholar]
- Overpeck, J.T.; Webb, T.I.; Prentice, I.C. Quantitative interpretation of fossil pollen spectra: Dissimilarity coefficients and the method of modern analogs. Quat. Res. 1985, 23, 87–108. [Google Scholar] [CrossRef]
- Hanberry, B.B. Compositional changes in selected forests of the western United States. Appl. Geogr. 2014, 52, 90–98. [Google Scholar] [CrossRef]
- Adams, D.L. The Northern Rocky Mountain Region. In Regional Silviculture of the United States; John Wiley: Hoboken, NJ, USA, 1980; pp. 341–389. [Google Scholar]
- Stout, D.L.; Sala, A. Xylem vulnerability to cavitation in Pseudotsuga menziesii and Pinus ponderosa from contrasting habitats. Tree Physiol. 2003, 23, 43–50. [Google Scholar] [CrossRef]
- Peter, D.; Shebitz, D. Historic anthropogenically maintained bear grass savannas of the southeastern Olympic Peninsula. Restor. Ecol. 2006, 14, 605–615. [Google Scholar] [CrossRef]
- Heyerdahl, E.K.; Miller, R.F.; Parsons, R.A. History of fire and Douglas-fir establishment in a savanna and sagebrush–grassland mosaic, southwestern Montana, USA. For. Ecol. Manag. 2006, 230, 107–118. [Google Scholar] [CrossRef]
- Heyerdahl, E.K.; Lertzman, K.; Wong, C.M. Mixed-severity fire regimes in dry forests of southern interior British Columbia, Canada. Can. J. For. Res. 2012, 42, 88–98. [Google Scholar] [CrossRef]
- NatureServe Explorer. Middle Rocky Mountain Montane Douglas-fir Forest and Woodland. Available online: https://explorer.natureserve.org/Search (accessed on 8 June 2020).
- Kent, W.H.B. The Proposed Wenaha Forest Reserve Washington and Oregon. Supervisor’s Office: U.S. Department of Agriculture, Bureau of Forestry. Available online: https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fsbdev7_015539.pdf (accessed on 20 October 2020).
- Stephens, S.L.; Martin, R.E.; Clinton, N.E. Prehistoric fire area and emissions from California’s forests, woodlands, shrublands, and grasslands. For. Ecol. Manag. 2007, 251, 205–216. [Google Scholar] [CrossRef]
- Odion, D.C.; Hanson, C.T.; Arsenault, A.; Baker, W.L.; DellaSala, D.A.; Hutto, R.L.; Klenner, W.; Moritz, M.A.; Sherriff, R.L.; Veblen, T.T.; et al. Examining historical and current mixed-severity fire regimes in ponderosa pine and mixed-conifer forests of western North America. PLoS ONE 2014, 9, e87852. [Google Scholar] [CrossRef]
- Hanson, C.T.; Sherriff, R.L.; Hutto, R.L.; DellaSala, D.A.; Veblen, T.T.; Baker, W.L. Setting the stage for mixed-and high-severity fire. In The Ecological Importance of Mixed-Severity Fires; DellaSala, D.A., Hanson, C.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 3–22. [Google Scholar]
- Fulé, P.Z.; Swetnam, T.W.; Brown, P.M.; Falk, D.A.; Peterson, D.L.; Allen, C.D.; Aplet, G.H.; Battaglia, M.A.; Binkley, D.; Farris, C.; et al. Unsupported inferences of high-severity fire in historical dry forests of the western United States: Response to Williams and Baker. Glob. Ecol. Biogeogr. 2014, 23, 825–830. [Google Scholar] [CrossRef]
- Hagmann, R.K.; Franklin, J.F.; Johnson, K.N. Historical structure and composition of ponderosa pine and mixed-conifer forests in south-central Oregon. For. Ecol. Manag. 2013, 304, 492–504. [Google Scholar] [CrossRef]
- Odion, D.C.; Hanson, C.T.; Baker, W.L.; DellaSala, D.A.; Williams, M.A. Areas of agreement and disagreement regarding ponderosa pine and mixed conifer forest fire regimes: A dialogue with Stevens et al. PLoS ONE 2016, 11, e0154579. [Google Scholar] [CrossRef] [PubMed]
- Hagmann, R.K.; Stevens, J.T.; Lydersen, J.M.; Collins, B.M.; Battles, J.J.; Hessburg, P.F.; Levine, C.R.; Merschel, A.G.; Stephens, S.L.; Taylor, A.H.; et al. Improving the use of early timber inventories in reconstructing historical dry forests and fire in the western United States: Comment. Ecosphere 2018, 9, e02232. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Palik, B.J.; He, H.S. Comparison of historical and current forest surveys for detection of homogenization and mesophication of Minnesota forests. Landsc. Ecol. 2012, 27, 1495–1512. [Google Scholar] [CrossRef]
- Hanberry, B.B. Trajectory from beech and oak forests to eastern broadleaf forests in Indiana, USA. Ecol. Process. 2019, 8, 3. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Bragg, D.C.; Hutchinson, T.F. A reconceptualization of open oak and pine ecosystems of eastern North America using a forest structure spectrum. Ecosphere 2018, 9, e02431. [Google Scholar] [CrossRef]
- Downing, W.M.; Krawchuk, M.A.; Coop, J.D.; Meigs, G.W.; Haire, S.L.; Walker, R.B.; Whitman, E.; Chong, G.; Miller, C.; Tortorelli, C. How do plant communities differ between fire refugia and fire-generated early-seral vegetation? J. Veg. Sci. 2020, 31, 26–39. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Historical | Bearing | Line | Current | FSVeg | 1997 to 2005 | |||
|---|---|---|---|---|---|---|---|---|
| Species | Count | Percent | Count | Percent | Species | Percent | Percent | Scientific Name |
| ponderosa pine | 8677 | 43.01 | 390 | 53.79 | grand fir | 25.11 | 27.71 | Abies grandis |
| Douglas-fir | 7646 | 37.9 | 199 | 27.45 | Douglas-fir | 22.88 | 19.84 | Pseudotsuga menziesii |
| western larch | 2044 | 10.13 | 84 | 11.59 | ponderosa pine | 18.68 | 15.56 | Pinus ponderosa |
| Engelmann spruce | 852 | 4.22 | 24 | 3.31 | lodgepole pine | 16.07 | 17.07 | Pinus contorta |
| lodgepole pine | 519 | 2.57 | 2 | 0.28 | Engelmann spruce | 7.57 | 6.13 | Picea engelmannii |
| true fir | 130 | 0.64 | 16 | 2.21 | western larch | 5.03 | 6.74 | Larix occidentalis |
| subalpine fir | 2.62 | 5.02 | Abies lasiocarpa | |||||
| Full Model Fir, Pine | Reduced Model Fir, Pine | Fir | Pine | ||
|---|---|---|---|---|---|
| Accuracy 0.8097, Fir 0.849, Pine 0.760 | Accuracy 0.801, Fir 0.841, Pine 0.755 | Value | Value | ||
| precipitation | 100 | precipitation | 100 | 963 | 641 |
| roughness | 55 | roughness | 30 | 5295 | 2075 |
| slope | 51 | slope | 14 | 21.5 | 11.8 |
| temperature | 43 | elevation | 9 | 1386 | 1355 |
| elevation | 42 | dissection | 8 | 0.53 | 0.55 |
| integrated moisture | 40 | temperature | 6 | 6.6 | 6.9 |
| dissection | 38 | integrated moisture | 3 | 218 | 159 |
| compound moisture | 38 | compound moisture | 0 | 6.7 | 7.2 |
| available water | 28 | ||||
| soil depth | 27 | ||||
| sand | 26 | ||||
| organic | 26 | ||||
| cation exchange | 25 | ||||
| clay | 25 | ||||
| North Historical | North Current | FSVeg | 1997 to 2005 | ||
|---|---|---|---|---|---|
| Species | Count | Percent | Species | Percent | Percent |
| Douglas-fir | 5472 | 52.51 | grand fir | 38.90 | 38.42 |
| ponderosa pine | 2869 | 27.53 | Douglas-fir | 29.39 | 25.20 |
| western larch | 1055 | 10.12 | Engelmann spruce | 12.75 | 9.13 |
| Engelmann spruce | 617 | 5.92 | ponderosa pine | 7.96 | 8.38 |
| true fir | 126 | 1.21 | western larch | 4.07 | 6.33 |
| lodgepole pine | 102 | 0.98 | subalpine fir | 3.76 | 7.67 |
| lodgepole pine | 2.99 | 4.35 | |||
| South Historical | South Current | FSVeg | 1997 to 2005 | ||
| Species | Count | Percent | Species | Percent | Percent |
| ponderosa pine | 5808 | 59.54 | ponderosa pine | 27.48 | 21.93 |
| Douglas-fir | 2174 | 22.29 | lodgepole pine | 26.81 | 28.30 |
| western larch | 989 | 10.14 | Douglas-fir | 17.55 | 15.17 |
| lodgepole pine | 417 | 4.28 | grand fir | 13.80 | 18.29 |
| Engelmann spruce | 235 | 2.41 | western larch | 5.81 | 6.99 |
| western juniper | 62 | 0.64 | western juniper | 3.48 | 3.03 |
| true fir | 4 | 0.04 | Engelmann spruce | 3.31 | 3.43 |
| subalpine fir | 1.68 | 2.71 | |||
| Historical | 1997 to 2005 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Density (Unadjusted) | Density | Low | High | DBH | BA | Low | High | Stock | Low | High | Density | BA | |
| Umatilla | 130.9 | 164.5 | 112.3 | 184.4 | 39.9 | 27.8 | 18.9 | 31.1 | 38.9 | 26.5 | 43.6 | 388.9 | 29.8 |
| North | 97.3 | 122.2 | 83.4 | 137.0 | 42.3 | 22.3 | 15.2 | 25.0 | 31.5 | 21.5 | 35.3 | 365.6 | 34.8 |
| South | 167.9 | 211.0 | 144.0 | 236.5 | 37.4 | 32.5 | 22.2 | 36.5 | 45.8 | 31.3 | 51.4 | 411.9 | 24.8 |
| Density (Unadjusted) | Density | Low | High | Douglas-Fir | Ponderosa Pine | Grand Fir | Lodgepole Pine | |
|---|---|---|---|---|---|---|---|---|
| Flat | 125.2 | 157.3 | 107.4 | 176.4 | 22.5 | 58.3 | 0.5 | 2.6 |
| Gentle slopes | 152.7 | 191.9 | 131.0 | 215.1 | 29.2 | 48.3 | 0.6 | 3.5 |
| Steep slopes | 95.9 | 120.5 | 82.2 | 135.1 | 55.8 | 31.2 | 0.9 | 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanberry, B.B.; Justice, D.C.; Powell, D.C. Discovering Douglas-Fir Woodlands in the Historical Forests of Umatilla National Forest, Eastern Oregon and Washington. Forests 2020, 11, 1122. https://doi.org/10.3390/f11101122
Hanberry BB, Justice DC, Powell DC. Discovering Douglas-Fir Woodlands in the Historical Forests of Umatilla National Forest, Eastern Oregon and Washington. Forests. 2020; 11(10):1122. https://doi.org/10.3390/f11101122
Chicago/Turabian StyleHanberry, Brice B., Donald C. Justice, and David C. Powell. 2020. "Discovering Douglas-Fir Woodlands in the Historical Forests of Umatilla National Forest, Eastern Oregon and Washington" Forests 11, no. 10: 1122. https://doi.org/10.3390/f11101122
APA StyleHanberry, B. B., Justice, D. C., & Powell, D. C. (2020). Discovering Douglas-Fir Woodlands in the Historical Forests of Umatilla National Forest, Eastern Oregon and Washington. Forests, 11(10), 1122. https://doi.org/10.3390/f11101122