Modeling of Forest Communities’ Spatial Structure at the Regional Level through Remote Sensing and Field Sampling: Constraints and Solutions



Abstract
1. Introduction
2. Materials and Methods
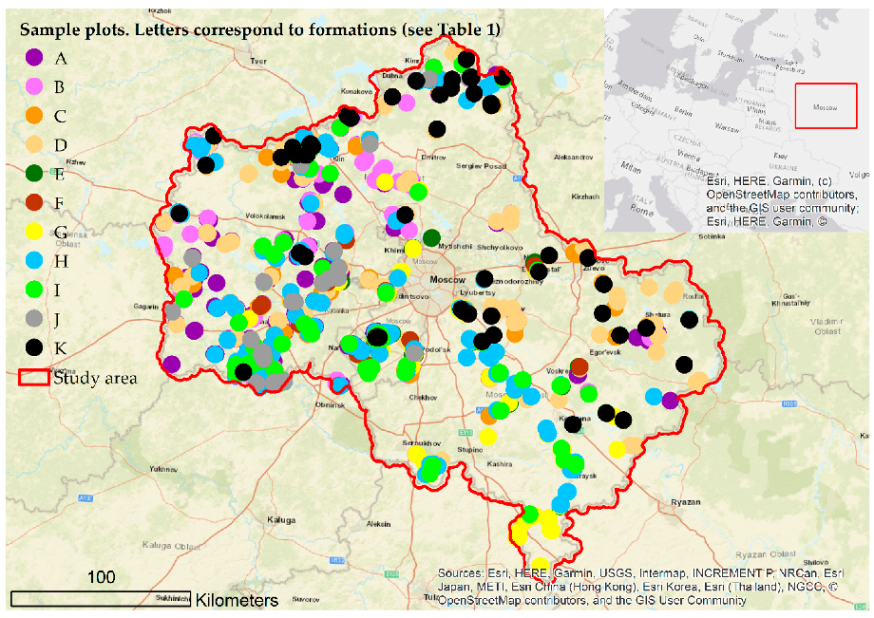
2.1. Study Area
2.2. Design of the Study
- Composition and structure of the tree layer (projective crown cover, average height of mature trees and undergrowth).
- Complete species composition of shrub, grass-dwarf shrub and moss layers, with an estimate of the cover in percent.
- Species saturation of the plants of the ground layers, estimated as the average number of species per unit area (to assess the species diversity).
3. Results
3.1. Pre-Processing of Samples
- Dwarf shrubs-small herb-green moss (DShG),
- Small herb (Sh),
- Small herb-broad herb (ShBh),
- Broad herb (Bh),
- Moist herb-broad herb (MhBh),
- Grass-marsh (Gm),
- Herb (H),
- Dwarf shrubs-herb-sphagnum (DHS).
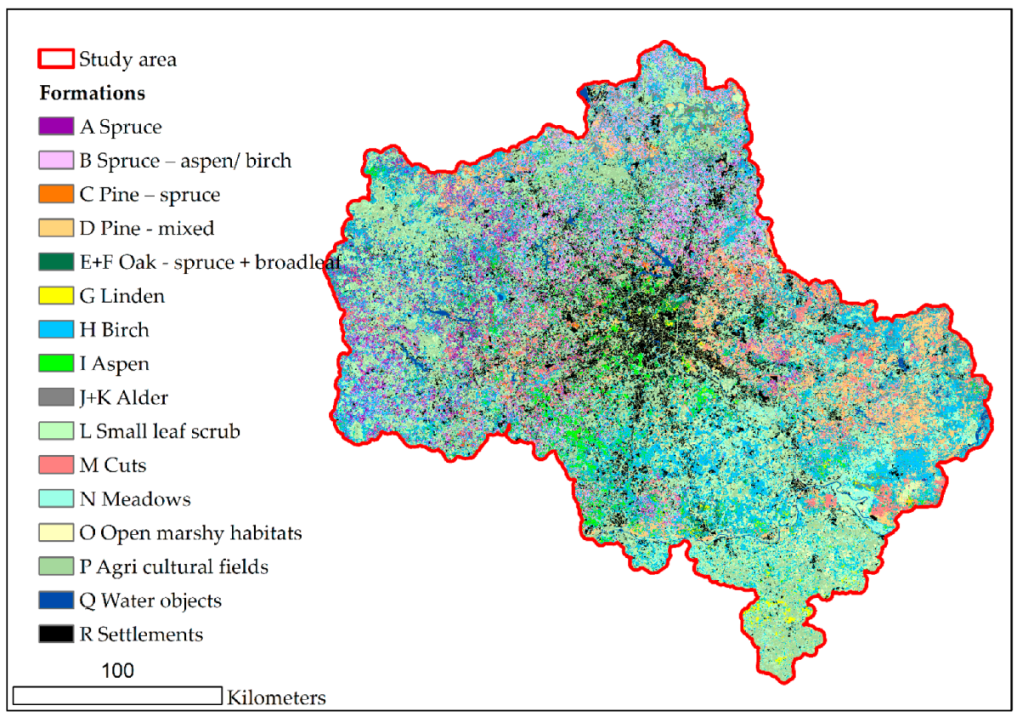
3.2. Modeling of Formations
3.3. Modeling of Association Groups
4. Discussion
- Creation of the set of field descriptions, evenly distributed in space and taking into account rare and remote habitats.
- Bringing the minimum number of descriptions of association groups to at least 50 (additional 494 descriptions), and in the long term, to 80 (1240 additional descriptions).
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| # | Sensor | Mosaic Date | Index | Removed Due to Autocorrelation > 95% |
|---|---|---|---|---|
| 1 | SRTM | 2009 | Elevation (meters) | |
| 2 | SRTM | 2009 | Slope (degrees) | |
| 3 | SRTM | 2009 | Aspect | |
| 4 | SRTM | 2009 | Shaded relief | |
| 5 | SRTM | 2009 | Profile Curvature | |
| 6 | SRTM | 2009 | Plan Convexity | |
| 7 | SRTM | 2009 | Longitude Convexity | yes |
| 8 | SRTM | 2009 | Cross Sectional Convexity | |
| 9 | SRTM | 2009 | Minimum Curvature | |
| 10 | SRTM | 2009 | Maximum Curvature | |
| 11 | SRTM | 2009 | Elevation Root Mean Square Error | |
| 12 | SRTM | 2009 | Slope (percent) | yes |
| 13 | SRTM | 2009 | Laplacian | |
| 14 | Landsat 8 | March 2019 | Band 1 | |
| 15 | Landsat 8 | March 2019 | Band 2 | yes |
| 16 | Landsat 8 | March 2019 | Band 3 | yes |
| 17 | Landsat 8 | March 2019 | Band 4 | yes |
| 18 | Landsat 8 | March 2019 | Band 5 | |
| 19 | Landsat 8 | March 2019 | Band 6 | |
| 20 | Landsat 8 | March 2019 | Band 7 | yes |
| 21 | Landsat 8 | March 2019 | EVI | |
| 22 | Landsat 8 | March 2019 | MSAVI | |
| 23 | Landsat 8 | March 2019 | NBR | |
| 24 | Landsat 8 | March 2019 | NBR2 | yes |
| 25 | Landsat 8 | March 2019 | NDMI | yes |
| 26 | Landsat 8 | March 2019 | NDVI | yes |
| 27 | Landsat 8 | March 2019 | SAVI | |
| 28 | Landsat 8 | May 2019 | Band 1 | |
| 29 | Landsat 8 | May 2019 | Band 2 | yes |
| 30 | Landsat 8 | May 2019 | Band 3 | yes |
| 31 | Landsat 8 | May 2019 | Band 4 | yes |
| 32 | Landsat 8 | May 2019 | Band 5 | |
| 33 | Landsat 8 | May 2019 | Band 6 | |
| 34 | Landsat 8 | May 2019 | Band 7 | yes |
| 35 | Landsat 8 | May 2019 | EVI | |
| 36 | Landsat 8 | May 2019 | MSAVI | |
| 37 | Landsat 8 | May 2019 | NBR | |
| 38 | Landsat 8 | May 2019 | NBR2 | yes |
| 39 | Landsat 8 | May 2019 | NDMI | yes |
| 40 | Landsat 8 | May 2019 | NDVI | |
| 41 | Landsat 8 | May 2019 | SAVI | |
| 42 | Landsat 8 | July 2019 | Band 1 | |
| 43 | Landsat 8 | July 2019 | Band 2 | |
| 44 | Landsat 8 | July 2019 | Band 3 | |
| 45 | Landsat 8 | July 2019 | Band 4 | |
| 46 | Landsat 8 | July 2019 | Band 5 | |
| 47 | Landsat 8 | July 2019 | Band 6 | |
| 48 | Landsat 8 | July 2019 | Band 7 | |
| 49 | Landsat 8 | July 2019 | EVI | |
| 50 | Landsat 8 | July 2019 | MSAVI | |
| 51 | Landsat 8 | July 2019 | NBR | |
| 52 | Landsat 8 | July 2019 | NBR2 | yes |
| 53 | Landsat 8 | July 2019 | NDMI | yes |
| 54 | Landsat 8 | July 2019 | NDVI | |
| 55 | Landsat 8 | July 2019 | SAVI | |
| 56 | Landsat 5 | July 2010 | Band 1 | |
| 57 | Landsat 5 | July 2010 | Band 2 | yes |
| 58 | Landsat 5 | July 2010 | Band 3 | yes |
| 59 | Landsat 5 | July 2010 | Band 4 | yes |
| 60 | Landsat 5 | July 2010 | Band 5 | |
| 61 | Landsat 5 | July 2010 | Band 6 | |
| 62 | Landsat 5 | July 2010 | Band 7 | yes |
| 63 | Landsat 5 | July 2010 | EVI | |
| 64 | Landsat 5 | July 2010 | MSAVI | |
| 65 | Landsat 5 | July 2010 | NBR | |
| 66 | Landsat 5 | July 2010 | NBR2 | yes |
| 67 | Landsat 5 | July 2010 | NDMI | yes |
| 68 | Landsat 5 | July 2010 | NDVI | yes |
| 69 | Landsat 5 | July 2010 | SAVI | |
| 70 | Landsat 5 | September 2019 | Band 1 | |
| 71 | Landsat 8 | September 2019 | Band 2 | yes |
| 72 | Landsat 8 | September 2019 | Band 3 | yes |
| 73 | Landsat 8 | September 2019 | Band 4 | yes |
| 74 | Landsat 8 | September 2019 | Band 5 | |
| 75 | Landsat 8 | September 2019 | Band 6 | |
| 76 | Landsat 8 | September 2019 | Band 7 | yes |
| 77 | Landsat 8 | September 2019 | EVI | |
| 78 | Landsat 8 | September 2019 | MSAVI | |
| 79 | Landsat 8 | September 2019 | NBR | |
| 80 | Landsat 8 | September 2019 | NBR2 | yes |
| 81 | Landsat 8 | September 2019 | NDMI | |
| 82 | Landsat 8 | September 2019 | NDVI | |
| 83 | Landsat 8 | September 2019 | SAVI | |
| 84 | Palsar-2 | 2019 | HH polarization | |
| 85 | Palsar-2 | 2019 | HV polarization |
References
- Olsson, H.; Nilsson, M.; Persson, A. Geoss possibilities and challenges related to nation wide forest monitoring. In Proceedings of the Proc. ISPRS Commission VII Mid Term Symposium, Vienna, Austria, 5–7 July 2010; p. 10. [Google Scholar]
- Tomppo, E. The Finnish national forest inventory. In Forest Inventory; Springer: Berlin/Heidelberg, Germany, 2006; pp. 179–194. [Google Scholar]
- Lisovsky, A.; Dudov, S. Advantages and limitations of application of the species distribution modeling methods. 2. Maxent. Biol. Bull. Rev. 2020, 81. [Google Scholar] [CrossRef]
- Edwards, T.C.; Cutler, D.R.; Zimmermann, N.E.; Geiser, L.; Moisen, G.G. Effects of sample survey design on the accuracy of classification tree models in species distribution models. Ecol. Model. 2006, 199, 132–141. [Google Scholar] [CrossRef]
- Cochran, W.G. Sampling Techniques, 3rd ed.; Wiley Series in Probability and Mathematical Statistics; Wiley: New York, NY, USA, 1977; ISBN 978-0-471-16240-7. [Google Scholar]
- Schreuder, H.T.; Gregoire, T.G.; Wood, G.B. Sampling Methods for Multiresource Forest Inventory; Wiley: New York, NY, USA, 1993. [Google Scholar]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Vallejos, R.; Osorio, F. Effective sample size of spatial process models. Spat. Stat. 2014, 9, 66–92. [Google Scholar] [CrossRef]
- Lisovsky, A.; Dudov, S.; Obolenskaya, E. Advantages and limitations of application of the species distribution modeling methods. 1. A general approach. Biol. Bull. Rev. 2020, 81. [Google Scholar] [CrossRef]
- FAO. Knowledge Reference for National Forest Assessment; FAO: Rome, Italy, 2015; ISBN 978-92-5-108832-6. [Google Scholar]
- Chernenkova, T.V.; Puzachenko, M.Y.; Belyaeva, N.G.; Kotlov, I.P.; Morozova, O.V. Pine Forests in Moscow Oblast: History and Perspectives of Preservation. Russ. J. For. Sci. 2019, 5, 449–464. (In Russian) [Google Scholar] [CrossRef]
- Hasegawa, H.; Yoshimura, T. Estimation of GPS positional accuracy under different forest conditions using signal interruption probability. J. For. Res. 2007, 12, 1–7. [Google Scholar] [CrossRef]
- Thomson, G.; Newman, P. Urban fabrics and urban metabolism–from sustainable to regenerative cities. Resour. Conserv. Recycl. 2018, 132, 218–229. [Google Scholar] [CrossRef]
- Girardet, H. Creating Regenerative Cities; Routledge: London, UK, 2014; ISBN 1-317-65410-2. [Google Scholar]
- Pedersen Zari, M. Devising Urban Biodiversity Habitat Provision Goals: Ecosystem Services Analysis. Forests 2019, 10, 391. [Google Scholar] [CrossRef]
- Karpachevsky, M.L.; Yaroshenko, A.Y.; Zenkevich, Y.E.; Aksenov, D.E.; Egorov, A.V.; Zhuravleva, I.V.; Rogova, N.V.; Tikhomirova, O.M.; Antonova, T.A.; Kurakina, I.N.; et al. The Nature of the Moscow Region: Losses of the Last Two Decades; Publishing House of the Center for Wildlife Conservation In Russian: Moscow, Russia, 2009; ISBN 978-5-93699-073-1. [Google Scholar]
- Forest Plan of Moscow Region for 2019–2028 (Lesnoj Plan Moskovskoj Oblasti na 2019–2028 gody); Moscow Region Government; Forestry Committee of the Moscow Region: Moscow, Russia, 2018.
- Vegetation of Moscow Region (Rastitel’nost’ Moskovskoj Oblasti); Ogureeva, G.N., Miklyaeva, I.M., Suslova, E.G., Shvergunova, L.V., Eds.; EKOR: Moscow, Russia, 1996. [Google Scholar]
- Gavriliuk, E.A.; Ershov, D.V. Method for joint processing of multi-season Landsat-TM images and creation of a map of terrestrial ecosystems of the Moscow region on their basis. Curr. Probl. Remote Sens. Earth Space 2012, 9, 15–23. [Google Scholar]
- Forest Management Instruction; Ministry of Natural Resources and Environment of the Russian Federation: Moscow, Russia, 2018.
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping Species Distributions with MAXENT Using a Geographically Biased Sample of Presence Data: A Performance Assessment of Methods for Correcting Sampling Bias. PLoS ONE 2014, 9, e97122. [Google Scholar] [CrossRef]
- Puzachenko, M.Y.; Chernenkova, T.V. Definition of factors of spatial variation in vegetation using RSD, DEM and field data by example of the central part of Murmansk Region. Curr. Probl. Remote Sens. Earth Space 2016, 13, 167–191. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [PubMed]
- Marzialetti, P.; Giovanni, L.; Santilli, G.; Huang, W.; Zappacosta, D. Maxent Model Application For Tree Pests Monitoring. In Proceedings of the IGARSS 2019–2019 IEEE International Geoscience and Remote Sensing Symposium, Yokohama, Japan, 28 July–2 August 2019; p. 6667. [Google Scholar]
- Chaiyos, J.; Suwannatrai, K.; Thinkhamrop, K.; Pratumchart, K.; Sereewong, C.; Tesana, S.; Kaewkes, S.; Sripa, B.; Wongsaroj, T.; Suwannatrai, A. MaxEnt modeling of soil-transmitted helminth infection distributions in Thailand. Parasitol. Res. 2018, 117. [Google Scholar] [CrossRef]
- Park, H.C.; Lim, J.C.; Lee, J.H.; Lee, G.G. Predicting the Potential Distributions of Invasive Species Using the Landsat Imagery and Maxent: Focused on “Ambrosia trifida L. var. trifida” in Korean Demilitarized Zone. J. Korean Soc. Environ. Restor. Technol. 2017, 20. [Google Scholar] [CrossRef][Green Version]
- Lahoz-Monfort, J.J.; Guillera-Arroita, G.; Milner-Gulland, E.J.; Young, R.P.; Nicholson, E. Satellite imagery as a single source of predictor variables for habitat suitability modelling: How Landsat can inform the conservation of a critically endangered lemur. J. Appl. Ecol. 2010, 47, 1094–1102. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, K.; Xia, Q. A MaxEnt Model for Mineral Prospectivity Mapping. Nat. Resour. Res. 2018. [Google Scholar] [CrossRef]
- Convertino, M.; Troccoli, A.; Catani, F. Detecting Fingerprints of Landslide Drivers: A MaxEnt Model. J. Geophys. Res. 2013, 118. [Google Scholar] [CrossRef]
- Casalegno, S.; Amatulli, G.; Bastrup-Birk, A.; Durrant, T.; Pekkarinen, A. Modelling and mapping the suitability of European forest formations at 1-km resolution. Eur. J. For. Res. 2011, 130, 971–981. [Google Scholar] [CrossRef]
- Attorre, F.; Francesconi, F.; Alfo, M.; Martella, F.; Valenti, R.; Vitale, M.; Sanctis, M. Classifying and Mapping Potential Distribution of Forest Types Using a Finite Mixture Model. Folia Geobot. 2012, 49, 313–335. [Google Scholar] [CrossRef]
- Chust, G.; Chave, J.; Condit, R.; Aguilar, S.; Lao, S.; Pérez, R. Determinants and spatial modeling of tree β-diversity in a tropical forest landscape in Panama. J. Veg. Sci. 2006, 17, 83–92. [Google Scholar] [CrossRef]
- Bergamin, R.; Müller, S.; Mello, R. Indicator species and floristic patterns in different forest formations in southern Atlantic rainforests of Brazil. Community Ecol. 2012, 13, 162–170. [Google Scholar] [CrossRef]
- Amici, V.; Marcantonio, M.; La Porta, N.; Rocchini, D. A multi-temporal approach in MaxEnt modelling: A new frontier for land use/land cover change detection. Ecol. Inform. 2017, 40, 40–49. [Google Scholar] [CrossRef]
- Vergel, K.; Zinicovscaia, I.; Yushin, N.; Frontasyeva, M.V. Heavy Metal Atmospheric Deposition Study in Moscow Region, Russia. Bull. Environ. Contam. Toxicol. 2019, 103, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Nefedova, T.G.; Mkrtchan, N.V. Migration of rural population and dynamics of agricultural employment in the regions of Russia. Vestn. Mosk. Univ. 2017, 58–67. [Google Scholar]
- Lurie, I.K.; Baldina, E.A.; Prasolova, A.I.; Prokhorova, E.A.; Semin, V.N.; Chistov, S.V. A series of maps of the environmental-geographical assessment of land resources of the New Moscow territory. Vestn. Mosk. Univ. 2015, 50–59. [Google Scholar]
- Chernenkova, T.V.; Morozova, O.V. Classification and Mapping of Coenotic Diversity of Forests. Contemp. Probl. Ecol. 2017, 10, 738–747. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J.; Gordon, A.; Kujala, H.; Lentini, P.E.; McCarthy, M.A.; Tingley, R.; Wintle, B.A. Is my species distribution model fit for purpose? Matching data and models to applications. Glob. Ecol. Biogeogr. 2015, 24, 276–292. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Model. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- Chernenkova, T.V.; Kotlov, I.P.; Belyaeva, N.G.; Morozova, O.V.; Suslova, E.G.; Puzachenko, M.Y.; Krenke, A.N. Sustainable Forest Management Tools for the Moscow Region. Sustain. For. Manag. Tools Mosc. Reg. 2019, 12, 35–56. [Google Scholar] [CrossRef]
- Chernenkova, T.V.; Kotlov, I.P.; Belyaeva, N.G.; Suslova, E.G.; Morozova, O.V.; Pesterova, O.; Arkhipova, M.V. Role of Silviculture in the Formation of Norway Spruce Forests along the Southern Edge of Their Range in the Central Russian Plain. Forests 2020, 11, 778. [Google Scholar] [CrossRef]
- Shimada, M.; Itoh, T.; Motooka, T.; Watanabe, M.; Shiraishi, T.; Thapa, R.; Lucas, R. New global forest/non-forest maps from ALOS PALSAR data (2007–2010). Remote Sens. Environ. 2014, 155, 13–31. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R. High-resolution global maps of 21st-century forest cover change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Nitsenko, A.A. Typology of Small-Leaved Forests in the European Part of the USSR; Leningrad University Publishing House: Leningrad, Russian, 1972. [Google Scholar]
- Tømmerås, B. Skogens Naturlige Dynamikk. Elementer og Prosesser i Naturlig Skogutvikling; Direktoratet for Naturforvaltning: Trondheim, Norway, 1994; Volume 5, pp. 1–47. [Google Scholar]
- Kurnaev, S.F. Main Forest Types of Russian Plain Middle Part (Osnovnye Tipy Lesa Srednej Chasti Russkoj Ravniny); Nauka: Moscow, Russian, 1968. [Google Scholar]
- Suslova, E.G. Forests of Moscow Region. Ecosyst. Ecol. Dyn. 2019, 3, 119–190. [Google Scholar]
- Yurkevich, I.D.; Geltman, V.S.; Parfenov, V.I. Gray Alder Forests and Their Economic Use; Publishing House of the Byelorussian Academy of Sciences: Minsk, Belarus, 1963. [Google Scholar]
- Liksakova, N.S. Small-leaved forests of Chudovsky district, Novgorod region. Bot. J. 2004, 89, 1319–1342. [Google Scholar]
- Haklay, M.; Weber, P. Openstreetmap: User-generated street maps. IEEE Pervasive Comput. 2008, 7, 12–18. [Google Scholar] [CrossRef]
- Chernenkova, T.V.; Puzachenko, M.Y.; Belyaeva, N.G.; Morozova, O.V. Evaluation of the structure and composition of forests in Moscow region based on field and remote sensing data. Izv. Ross. Akad. Nauk. Seriya Geogr. 2019, 112–124. [Google Scholar] [CrossRef]
- Belyaeva, N.G.; Chernenkova, T.V.; Morozova, O.V.; Sandlerskii, R.B.; Arkhipova, M.V. Comparing Eco-Phytocoenotic and Eco-Floristic Methods of Classification to Estimate Coenotic Diversity and to Map Forest Vegetation. Contemp. Probl. Ecol. 2018, 11, 729–742. [Google Scholar] [CrossRef]
- Porfiriev, V.S. On the application of the concepts of series and cycle in the study of coniferous-deciduous forests. Bull. Mosc. Soc. Nat. Biol. Dep. 1960, 93–99. [Google Scholar]
- Chernenkova, T.V.; Morozova, O.V.; Belyaeva, N.G.; Puzachenko, M.Y. Actual organization of forest communities with broad-leaved trees in broad-leaved-coniferous zone (with Moscow region as an example). Veg. Russ. 2018, 33, 107–130. [Google Scholar] [CrossRef]
- Varentsov, M.I.; Samsonov, T.E.; Kislov, A.V.; Konstantinov, P.I. Simulations of Moscow agglomeration heat island within the framework of the regional climate model Cosmo-CLM. Vestn. Mosk. Univ. 2017, 25–37. [Google Scholar]
- Rysin, L.P.; Savelieva, L.I. Cadastres of Forest Types and Types of Forest Biodeosenoses (Kadastry Tipov lesa i Tipov Lesnyh Biogeocenozov); Tovarishhestvo nauchnyh izdanij KMK: Moscow, Russia, 2007; ISBN 978-5-87317-397-6. [Google Scholar]
- Rysin, L.P.; Abaturov, A.V.; Savelieva, L.I. Dynamic of Coniferous Forests of Moscow Region (Dinamika Hvojnyh Lesov Podmoskov’ja); Nauka: Moscow, Russian, 2000. [Google Scholar]
- Irish, R.R. Landsat 7 science data users handbook. Natl. Aeronaut. Space Adm. Rep. 2000, 2000, 415–430. [Google Scholar]
- Rocchini, D. Effects of spatial and spectral resolution in estimating ecosystem alpha-diversity by satellite imagery. Remote Sens. Environ. 2007, 111, 423–434. [Google Scholar] [CrossRef]
- Toll, D.L. Landsat-4 Thematic Mapper scene characteristics of a suburban and rural area. Photogramm. Eng. Remote Sens. 1985, 51, 1471–1482. [Google Scholar]


| Formations 1 | Spruce | Spruce-Aspen/Birch | Pine-Spruce | Pine | Oak-Spruce | Broad Leaf-Spruce | Linden | Birch | Aspen | Grey Alder | Black Alder |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Association groups | A | B | C | D | E | F | G | H | I | J | K |
| DShG | 37 | 30 | 32 | 46 | |||||||
| Sh | 39 | 22 | 16 | 23 | 9 | ||||||
| ShBh | 146 | 78 | 44 | 35 | 29 | ||||||
| Bh | 147 | 102 | 41 | 64 | 57 | 38 | 112 | 154 | 84 | ||
| MhBh | 18 | 16 | 30 | 24 | |||||||
| Gm | 17 | 31 | |||||||||
| H | 15 | 24 | |||||||||
| DHS | 46 | 10 | |||||||||
| Total number of sample plots | 369 | 232 | 133 | 229 | 57 | 38 | 112 | 261 | 100 | 30 | 55 |
| Spatial rarefication, km | 10 | 10 | 1 | 5 | - | - | - | 10 | - | - | - |
| Number of sample plots after rarefication | 97 | 87 | 93 | 82 | 95 2 | 112 | 95 | 100 | 85 3 | ||
| Habitat Type | Small Leaf Scrub | Cuts | Meadows | Open Marshy Habitats | Agri Cultural Fields | Water Objects | Settlements |
|---|---|---|---|---|---|---|---|
| L | M | N | O | P | Q | R | |
| Number of points/source | 53 | Global forest watch (loss year) | 53 | 27 | 78 | Global forest watch (data mask) | Openstreetmap (OSM) |
| Forest Plan Data | Formations | Method of Modeling Proportion of Formation (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 8 1 | |||
| Spruce | 24.4 | A | 3.2 | 3.3 | 7.0 | 7.7 | 5.25 | 5.0 | 12.6 | 7.6 | 6.6 | 7.0 |
| B | 4.8 | 13.7 | 11.2 | 15.0 | 9.30 | 19.2 | 13.2 | 16.0 | 15.1 | 18.1 | ||
| Pine | 20.7 | C | 5.4 | 4.6 | 7.5 | 4.5 | 2.84 | 5.8 | 4.4 | 2.5 | 2.5 | 2.5 |
| D | 9.7 | 9.8 | 15.4 | 10.0 | 17.39 | 11.6 | 8.9 | 15.5 | 14.4 | 16.0 | ||
| Oak | 1.7 | E | 14.0 | 9.7 | 11.0 | 10.3 | 11.72 | 10.5 | 9.9 | 4.1 | 9.0 | 2.9 |
| Broad leaf 2 | 0.08 | F | 17.5 | 19.4 | ||||||||
| Linden | 0.64 | G | 0.7 | 1.7 | 2.1 | 2.0 | 4.4 | |||||
| Birch | 39.6 | H | 16.0 | 14.3 | 35.9 | 31.2 | 15.38 | 22.7 3 | 30.0 | 32.5 | 35.5 | 31.0 |
| Aspen | 8.4 | I | 2.6 | 9.3 | 4.9 | 6.6 | 21.23 | 12.6 | 6.4 | 6.2 | 6.1 | 5.1 |
| Grey alder | 2.3 | J | 15.3 | 4.4 | 7.1 | 14.8 | 16.88 | - | 14.7 | 13.5 | 9.0 | 13.0 |
| Black alder | 1.8 | K | 10.6 | 9.9 | 12.6 | |||||||
| Formation | A | B | C | D | E + F | G | H | I | J + K | Total | User Accuracy |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A | 30 | 8 | 15 | 8 | 10 | 4 | 4 | 0 | 3 | 82 | 0.37 |
| B | 25 | 39 | 8 | 5 | 11 | 4 | 11 | 15 | 7 | 125 | 0.31 |
| C | 13 | 1 | 36 | 7 | 0 | 0 | 2 | 0 | 0 | 59 | 0.61 |
| D | 4 | 9 | 24 | 51 | 0 | 2 | 6 | 4 | 3 | 103 | 0.50 |
| E + F | 2 | 4 | 1 | 0 | 29 | 8 | 4 | 12 | 2 | 62 | 0.47 |
| G | 0 | 1 | 0 | 1 | 2 | 46 | 5 | 5 | 3 | 63 | 0.73 |
| H | 13 | 18 | 4 | 9 | 21 | 17 | 51 | 23 | 4 | 160 | 0.32 |
| I | 3 | 6 | 0 | 1 | 8 | 16 | 7 | 28 | 3 | 72 | 0.39 |
| J + K | 2 | 0 | 0 | 0 | 5 | 5 | 4 | 4 | 54 | 74 | 0.73 |
| Total | 92 | 86 | 88 | 82 | 86 | 102 | 94 | 91 | 79 | 800 | 0.37 |
| P_Accuracy | 0.33 | 0.45 | 0.41 | 0.62 | 0.34 | 0.45 | 0.54 | 0.31 | 0.68 | 0.46 | |
| Kappa | 0.39 |
| Formations | Spruce | Spruce-Aspen/Birch | Pine-Spruce | Pine | Oak-Spruce | Broad-Leaf Spruce | Linden | Birch | Aspen | Grey Alder | Black Alder |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Association groups | A | B | C | D | E | F | G | H | I | J | K |
| DShG | 0.47 0.21 | 3.07 0.27 | 0.36 0.21 | 2.44 0.43 | |||||||
| Sh | 1.37 0.24 | 1.65 0.14 | 0.41 0.2 | 2.51 0.14 | 0.28 0.33 | ||||||
| ShBh | 0.62 0.11 | 1.58 0.14 | 0.38 0.33 | 0.19 0.03 | 1.18 0.08 | ||||||
| Bh | 1.31 0.2 | 1.71 0.18 | 0.08 0.19 | 1.12 0.38 | 1.10 0.38 | 1.01 0.19 | 1.73 0.35 | 2.85 0.25 | 1.73 0.28 | ||
| MhBh | 1.89 0.06 | 1.36 0.5 | 1.44 0.69 | 0.46 0.43 | |||||||
| Gm | 4.70 0.35 | 4.55 0.57 | |||||||||
| H | 0.10 0.23 | 3.72 0.56 | |||||||||
| DHS | 0.91 0.55 | 1.32 0.5 | |||||||||
| Mean % of point matching | 0.19 | 0.18 | 0.23 | 0.29 | 0.38 | 0.19 | 0.35 | 0.3 | 0.39 | 0.69 | 0.5 |
| Habitat Type | Small Leaf Scrub | Cuts | Meadows | Open Marshy Habitats | Agri Cultural Fields | Water Objects | Settlements |
|---|---|---|---|---|---|---|---|
| L | M | N | O | P | Q | R | |
| % total cover | 14.69 | 4.16 | 7.07 | 2.34 | 12.85 | 1.08 | 9.23 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotlov, I.; Chernenkova, T. Modeling of Forest Communities’ Spatial Structure at the Regional Level through Remote Sensing and Field Sampling: Constraints and Solutions. Forests 2020, 11, 1088. https://doi.org/10.3390/f11101088
Kotlov I, Chernenkova T. Modeling of Forest Communities’ Spatial Structure at the Regional Level through Remote Sensing and Field Sampling: Constraints and Solutions. Forests. 2020; 11(10):1088. https://doi.org/10.3390/f11101088
Chicago/Turabian StyleKotlov, Ivan, and Tatiana Chernenkova. 2020. "Modeling of Forest Communities’ Spatial Structure at the Regional Level through Remote Sensing and Field Sampling: Constraints and Solutions" Forests 11, no. 10: 1088. https://doi.org/10.3390/f11101088
APA StyleKotlov, I., & Chernenkova, T. (2020). Modeling of Forest Communities’ Spatial Structure at the Regional Level through Remote Sensing and Field Sampling: Constraints and Solutions. Forests, 11(10), 1088. https://doi.org/10.3390/f11101088