Ten Years of Provenance Trials and Application of Multivariate Random Forests Predicted the Most Preferable Seed Source for Silviculture of Abies sachalinensis in Hokkaido, Japan


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Regional Groups and Seed Provenances
2.3. Range-Wide Provenance Tests
2.4. Climatic Data
2.5. Statistical Model for the Height Growth of A. sachalinensis
3. Results
3.1. Measured Height Growth Performance among Testing Sites
3.2. Model Accuracy and Climatic Conditions Controlling Height Growth of A. sachalinensis
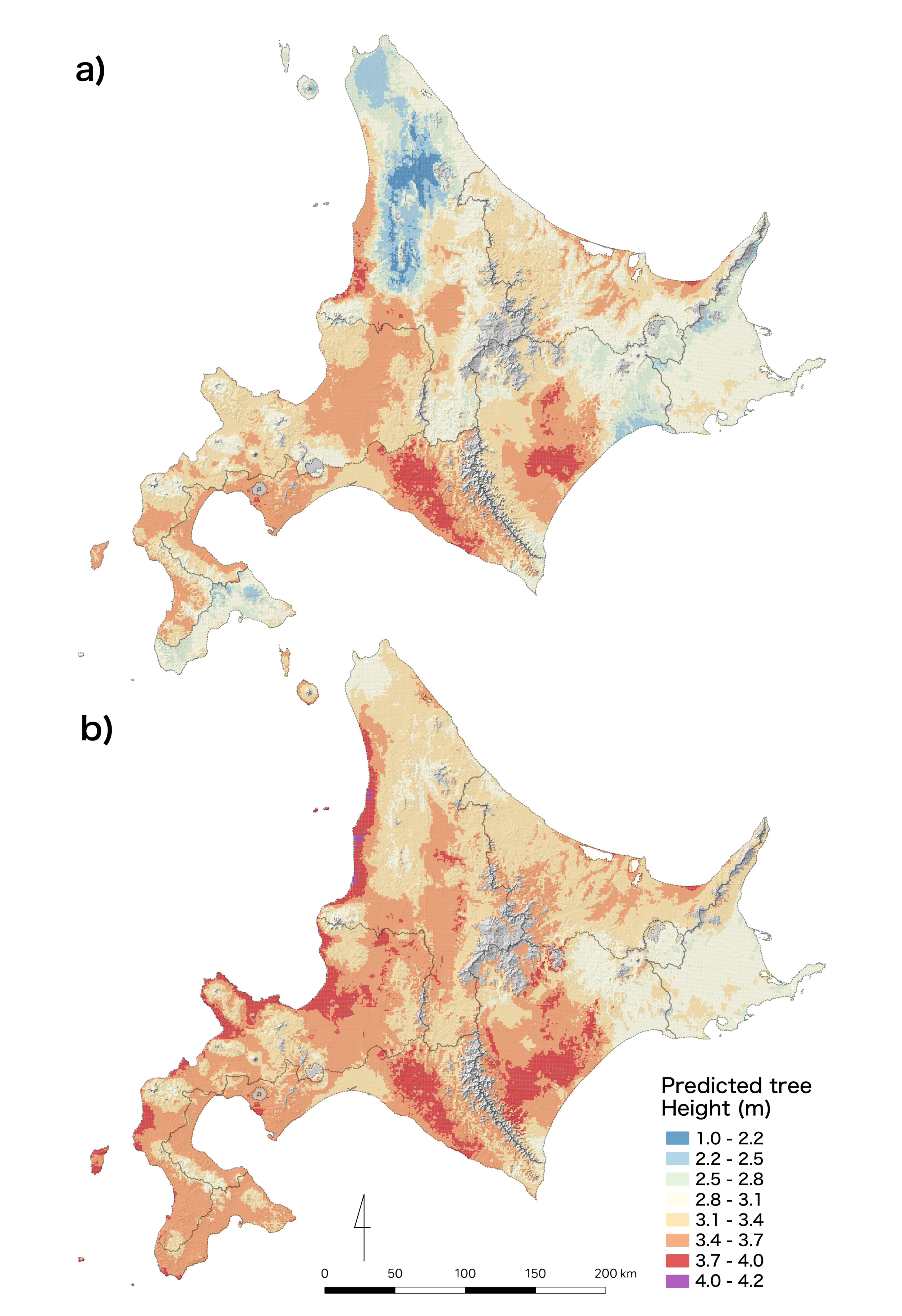
3.3. Prediction Maps for Tree Height
4. Discussion
4.1. Important Climatic Factors Affecting the Height Growth of A. sachalinensis
4.2. Optimum Regions and Seed Sources for Height Growth of A. sachalinensis
4.3. Implications for Improving the Height Growth of A. sachalinensis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HRO | Hokkaido Research Organization |
| WI | Warmth Index |
| TMC | Monthly mean daily minimum temperature |
| PRS | Precipitation during summer |
| MSD | Maximum snow depth |
| WinSR | Solar radiation in winter |
| SprSR | Solar radiation in spring |
| SumSR | Solar radiation in summer |
| AutSR | Solar radiation in autumn |
| MRF | Multivariate random forests |
| RF | Random forests |
| QGIS | Quantum Geographic Information System |
References
- Hufford, K.M.; Mazer, S.J. Plant ecotypes: Genetic differentiation in the age of ecological restoration. Trends Ecol. Evol. 2003, 18, 147–155. [Google Scholar] [CrossRef]
- Montalvo, A.M.; Ellstrand, N.C. Transplantation of the subshrub Lotus scoparius: Testing the home-site advantage hypothesis. Conserv. Biol. 2000, 14, 1034–1045. [Google Scholar] [CrossRef]
- Savolainen, O.; Pyhäjärvi, T.; Knürr, T. Gene flow and local adaptation in trees. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- Turesson, G. The genotypical response of the plant species to the habitat. Hereditas 1922, 3, 211–350. [Google Scholar] [CrossRef]
- Morgenstern, E.K. Geographic Variation in Forest Trees: Genetic Basis and Application of Knowledge in Silviculture; UBC Press: Vancouver, BC, Canada, 1996. [Google Scholar]
- Isik, F.; Keskin, S.; McKeand, S.E. Provenance variation and provenance-site interaction in Pinus brutia Ten.: Consequences of defining breeding zones. Silvae Genet. 2000, 49, 213–223. [Google Scholar]
- White, T.; Adams, W.; Neale, D. Forest Genetics; CAB International: Wallingford, UK, 2007. [Google Scholar]
- Reich, P.B.; Oleksyn, J. Climate warming will reduce growth and survival of Scots pine except in the far north. Ecol. Lett. 2008, 11, 588–597. [Google Scholar] [CrossRef]
- Kitzmiller, J.H. Provenance trials of ponderosa pine in northern California. For. Sci. 2005, 51, 595–607. [Google Scholar]
- Ying, C.C.; Yanchuk, A.D. The development of British Columbia’s tree seed transfer guidelines: Purpose, concept, methodology, and implementation. For. Ecol. Manag. 2006, 227, 1–13. [Google Scholar] [CrossRef]
- Gray, L.K.; Hamann, A.; John, S.; Rweyongeza, D.; Barnhardt, L.; Thomas, B.R. Climate change risk management in tree improvement programs: Selection and movement of genotypes. Tree Genet. Genomes 2016, 12, 23. [Google Scholar] [CrossRef]
- Lindgren, D.; Ying, C.C. A model integrating seed source adaptation and seed use. New For. 2000, 20, 87–104. [Google Scholar] [CrossRef]
- Schmidtling, R.C.; Carroll, E.; LaFarge, T. Allozyme diversity of selected and natural loblolly pine populations. Silvae Genet. 1999, 48, 35–45. [Google Scholar]
- Hamann, A.; Gylander, T.; Chen, P.Y. Developing seed zones and transfer guidelines with multivariate regression trees. Tree Genet. Genomes 2011, 7, 399–408. [Google Scholar] [CrossRef]
- Nagamitsu, T.; Shimada, K.i.; Kanazashi, A. A reciprocal transplant trial suggests a disadvantage of northward seed transfer in survival and growth of Japanese red pine (Pinus densiflora) trees. Tree Genet. Genomes 2015, 11, 813. [Google Scholar] [CrossRef]
- Potter, K.M.; Hargrove, W.W. Determining suitable locations for seed transfer under climate change: A global quantitative method. New For. 2012, 43, 581–599. [Google Scholar] [CrossRef][Green Version]
- Isaac-Renton, M.G.; Roberts, D.R.; Hamann, A.; Spiecker, H. Douglas-fir plantations in Europe: A retrospective test of assisted migration to address climate change. Glob. Chang. Biol. 2014, 20, 2607–2617. [Google Scholar] [CrossRef]
- Matsui, T.; Yagihashi, T.; Nakaya, T.; Tanaka, N.; Taoda, H. Climatic controls on distribution of Fagus crenata forests in Japan. J. Veg. Sci. 2004, 15, 57–66. [Google Scholar] [CrossRef]
- Tsuyama, I.; Horikawa, M.; Nakao, K.; Matsui, T.; Kominami, Y.; Tanaka, N. Factors controlling the distribution of a keystone understory taxon, dwarf bamboo of the section Crassinodi, at a national scale: Application to impact assessment of climate change in Japan. J. For. Res. 2012, 17, 137–148. [Google Scholar] [CrossRef]
- Higa, M.; Tsuyama, I.; Nakao, K.; Nakazono, E.; Matsui, T.; Tanaka, N. Influence of nonclimatic factors on the habitat prediction of tree species and an assessment of the impact of climate change. Landsc. Ecol. Eng. 2013, 9, 111–120. [Google Scholar] [CrossRef]
- Nakao, K.; Higa, M.; Tsuyama, I.; Lin, C.T.; Sun, S.T.; Lin, J.R.; Chiou, C.R.; Chen, T.Y.; Matsui, T.; Tanaka, N. Changes in the potential habitats of 10 dominant evergreen broad-leaved tree species in the Taiwan-Japan archipelago. Plant Ecol. 2014, 215, 639–650. [Google Scholar] [CrossRef]
- Nakao, K.; Higa, M.; Tsuyama, I.; Matsui, T.; Horikawa, M.; Tanaka, N. Spatial conservation planning under climate change: Using species distribution modeling to assess priority for adaptive management of Fagus crenata in Japan. J. Nat. Conserv. 2013, 21, 406–413. [Google Scholar] [CrossRef]
- Tsuyama, I.; Higa, M.; Nakao, K.; Matsui, T.; Horikawa, M.; Tanaka, N. How will subalpine conifer distributions be affected by climate change? Impact assessment for spatial conservation planning. Reg. Environ. Chang. 2015, 15, 393–404. [Google Scholar] [CrossRef]
- Serra-Diaz, J.M.; Franklin, J.; Ninyerola, M.; Davis, F.W.; Syphard, A.D.; Regan, H.M.; Ikegami, M. Bioclimatic velocity: The pace of species exposure to climate change. Divers. Distrib. 2014, 20, 169–180. [Google Scholar] [CrossRef]
- Carroll, C.; Lawler, J.J.; Roberts, D.R.; Hamann, A. Biotic and climatic velocity identify contrasting areas of vulnerability to climate change. PLoS ONE 2015, 10, e0140486. [Google Scholar] [CrossRef] [PubMed]
- Hamann, A.; Roberts, D.R.; Barber, Q.E.; Carroll, C.; Nielsen, S.E. Velocity of climate change algorithms for guiding conservation and management. Glob. Chang. Biol. 2015, 21, 997–1004. [Google Scholar] [CrossRef]
- Barber, Q.E.; Nielsen, S.E.; Hamann, A. Assessing the vulnerability of rare plants using climate change velocity, habitat connectivity, and dispersal ability: A case study in Alberta, Canada. Reg. Environ. Chang. 2016, 16, 1433–1441. [Google Scholar] [CrossRef]
- Yamazaki, T. Pinaceae. In Flora of Japan: Pteridophyta and Gymnospermae; Iwatsuki, K., Yamazaki, T., Boufford, D.E., Ohba, H., Eds.; Kodansha: Tokyo, Japan, 1995; pp. 266–277. [Google Scholar]
- Hatakeyama, S. Genetical and breeding studies on the regional differences of interprovenance variation in Abies sachalinensis Mast. Bull. Hokkaido For. Res. Inst. 1981, 19, 1–19, (In Japanese with English Summary). [Google Scholar]
- Kuromaru, M. Effect of seed zoning for breeding seed-lot of Abies sachalinensis. Kosyunai Kihou 1989, 76, 1–3. (In Japanese) [Google Scholar]
- Nakada, R.; Sakamoto, S.; Nishioka, N.; Hanaoka, S.; Kita, K.; Kon, H.; Ishizuka, W.; Kuromaru, M. Selection of superior genotypes from the population of plus trees in Abies sachalinensis by the results from progeny trials in Hokkaido. Bull. FFPRI 2018, 17, 155–174, (In Japaneses with English Summary). [Google Scholar]
- Eiga, S.; Sakai, A. Regional Variation in Cold Hardiness of Sakhalin fir (Abies sachalinensis Mast.) in Hokkaido, Japan; Li, P.H., Ed.; Plant Cold Hardiness; Alan R. Liss, Inc.: New York, NY, USA, 1987; pp. 169–182. [Google Scholar]
- Tatewaki, M. Forest ecology of the islands of the North Pacific Ocean. J. Fac. Agric. Hokkaido Univ. 1958, 50, 371–486. [Google Scholar]
- Kitamura, K.; Uchiyama, K.; Ueno, S.; Ishizuka, W.; Tsuyama, I.; Goto, S. Geographical gradients of genetic diversity and differentiation among the southernmost marginal populations of Abies sachalinensis revealed by EST-SSR polymorphism. Forests 2020, 11, 233. [Google Scholar] [CrossRef]
- Hirosawa, T.; Sasaki, K.; Watanabe, K.; Imoto, M.; Kuromaru, M. Inter-provenance differences in growth conditions of Abies sachalinensis plantations in heavy snow zone. For. Tree Breed. Hokkaido 2000, 43, 9–11. (In Japanese) [Google Scholar]
- Ishizuka, W.; Kon, H.; Kita, K. Selection for 2nd generation plus tree in Abies sachalinensis in eastern Hokkaido, Japan. Kosyunai Kihou 2015, 176, 9–14. (In Japanese) [Google Scholar]
- Kuromaru, M. The difference of adaptability to heavy snow zone among provenances and families of Abies sachalinensis. For. Tree Breed. 1988, 146, 6–9. (In Japanese) [Google Scholar]
- Kuromaru, M. Inter-provenance variation in resistance to Scleroderris canker of using progeny of plus trees in Abies sachalinensis. For. Tree Breed. Hokkaido 1994, 37, 20–23. (In Japanese) [Google Scholar]
- Japan Meteorological Agency. Mesh Climate Data of Japan; Japan Meteorological Business Support Center: Tokyo, Japan, 2002. (In Japanese) [Google Scholar]
- Kira, T. Forest ecosystems of east and southeast Asia in a global perspective. Ecol. Res. 1991, 6, 185–200. [Google Scholar] [CrossRef]
- Segal, M.; Xiao, Y. Multivariate random forests. Wiley Interdiscip. Rev. Data Min. Knowl. Discov. 2011, 1, 80–87. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Ishwaran, H.; Kogalur, U.; Blackstone, E.; Lauer, M. Random survival forests. Ann. Appl. Stat. 2008, 2, 841–860. [Google Scholar] [CrossRef]
- Ishwaran, H.; Kogalur, U. Fast Unified Random Forests for Survival, Regression, and Classification (RF-SRC). R Package Version 2.9.3. 2020. Available online: https://cran.r-project.org/web/packages/randomForestSRC/randomForestSRC.pdf (accessed on 30 July 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- QGIS Geographic Information System. QGIS.org. Open Source Geospatial Foundation Project. 2020. Available online: https://qgis.org/en/site/forusers/download.html (accessed on 30 July 2020).
- Okada, S.; Mukaide, H.; Sakai, A. Genetic variation in Saghalien fir from different areas of Hokkaido. Silvae Genet. 1973, 22, 1–2. [Google Scholar]
- Sakagami, Y.; Fujimura, Y. Seasonal changes in the net photosynthetic and respiratory rate of Abies sachalinensis and Picea glehnii seedlings. J. Jpn. For. Soc. 1981, 63, 194–200, (In Japanese with English Summary). [Google Scholar]
- Kitao, M.; Kitaoka, S.; Harayama, H.; Agathokleous, E.; Han, Q.; Uemura, A.; Furuya, N.; Ishibashi, S. Sustained growth suppression in forest-floor seedlings of Sakhalin fir associated with previous-year springtime photoinhibition after a winter cutting of canopy trees. Eur. J. For. Res. 2019, 138, 143–150. [Google Scholar] [CrossRef]
- Hirano, Y.; Saitoh, T.M.; Fukatsu, E.; Kobayashi, H.; Muraoka, H.; Shen, Y.; Yasue, K. Effects of climate factors on the tree-ring structure of Cryptomeria japonica in central Japan. J. Jpn. Wood Res. Soc. 2020, 66, 117–127, (In Japanese with English Summary). [Google Scholar] [CrossRef]
- Koyama, Y.; Takahashi, M.; Murauchi, Y.; Fukatsu, E.; Watanabe, A.; Tomaru, N. Japanese beech (Fagus crenata) plantations established from seedlings of non-native genetic lineages. J. For. Res. 2012, 17, 116–120. [Google Scholar] [CrossRef]
- Tsumura, Y.; Uchiyama, K.; Moriguchi, Y.; Ueno, S.; Ihara-Ujino, T. Genome scanning for detecting adaptive genes along environmental gradients in the Japanese conifer, Cryptomeria japonica. Heredity 2012, 109, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Farjon, A. Pinaceae. In Drawings and Descriptions of the Genera Abies, Cedrus, Pseudolarix, Keteleeria, Nothotsuga, Tsuga, Cathaya, Pseudotsuga, Larix and Picea; Koeltz Scientific Books: Königstein, Germany, 1990. [Google Scholar]
- Semerikova, S.; Semerikov, V. Molecular phylogenetic analysis of the genus Abies (Pinaceae) based on the nucleotide sequence of chloroplast DNA. Rus. J. Genet. 2014, 50, 7–19. [Google Scholar] [CrossRef]
- Etterson, J.R.; Cornett, M.W.; White, M.A.; Kavajecz, L.C. Assisted migration across fixed seed zones detects adaptation lags in two major North American tree species. Ecol. Appl. 2020, 30, e02092. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Tchebakova, N.M.; Parfenova, Y.I.; Wykoff, W.R.; Kuzmina, N.A.; Milyutin, L.I. Intraspecific responses to climate in Pinus sylvestris. Glob. Chang. Biol. 2002, 8, 912–929. [Google Scholar] [CrossRef]
- Kapeller, S.; Lexer, M.J.; Geburek, T.; Hiebl, J.; Schueler, S. Intraspecific variation in climate response of Norway spruce in the eastern Alpine range: Selecting appropriate provenances for future climate. For. Ecol. Manag. 2012, 271, 46–57. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Crookston, N.L.; Warwell, M.V.; Evans, J.S. Empirical analyses of plant-climate relationships for the western United States. Int. J. Plant Sci. 2006, 167, 1123–1150. [Google Scholar] [CrossRef]
- Bower, A.D.; Aitken, S.N. Ecological genetics and seed transfer guidelines for Pinus albicaulis (Pinaceae). Am. J. Bot. 2008, 95, 66–76. [Google Scholar] [CrossRef]
- Chakraborty, D.; Schueler, S.; Lexer, M.J.; Wang, T. Genetic trials improve the transfer of Douglas-fir distribution models across continents. Ecography 2019, 42, 88–101. [Google Scholar] [CrossRef]
- O’Neill, G.; Wang, T.; Ukrainetz, N.; Charleson, L.; McAuley, L.; Yanchuk, A.; Zedel, S. A Proposed Climate-Based Seed Transfer System for British Columbia. Technical Report, BC Tech. Rep. 099. 2017. Available online: www.for.gov.bc.ca/hfd/pubs/Docs/Tr/Tr099.htm (accessed on 30 July 2020).
- Aitken, S.N.; Bemmels, J.B. Time to get moving: Assisted gene flow of forest trees. Evol. Appl. 2016, 9, 271–290. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, W.; Goto, S. Modeling intraspecific adaptation of Abies sachalinensis to local altitude and responses to global warming, based on a 36-year reciprocal transplant experiment. Evol. Appl. 2012, 5, 229–244. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Regional | Latitude | Longitude | No. of | Tree Height | Survival |
|---|---|---|---|---|---|---|
| Code | Group | (N) | (E) | Planted Families | (m) | Rate (%) |
| A30 | SS | 41.8488 | 141.1192 | 53 | 3.47 ± 0.70 | 90.5 |
| A31 | W | 42.7481 | 140.6308 | 41 | 2.79 ± 1.41 | 65.6 |
| A32 | S | 42.4586 | 142.4664 | 45 | 3.32 ± 0.96 | 63.7 |
| A33 | W | 43.2803 | 141.8785 | 82 | 3.50 ± 0.90 | 85.7 |
| A34 | N | 44.5550 | 142.2576 | 55 | 1.56 ± 0.75 | 76.8 |
| A35 | EN | 44.2273 | 142.9343 | 51 | 3.17 ± 0.91 | 77.1 |
| A36 | EN | 43.6542 | 143.7846 | 49 | 2.97 ± 0.98 | 59.4 |
| A37 | ES | 42.7309 | 143.4939 | 64 | 3.71 ± 0.84 | 93.7 |
| A38 | EE | 43.0415 | 144.9744 | 64 | 2.37 ±0.91 | 70.7 |
| Average | 56 | 3.14 ± 1.08 | 75.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsuyama, I.; Ishizuka, W.; Kitamura, K.; Taneda, H.; Goto, S. Ten Years of Provenance Trials and Application of Multivariate Random Forests Predicted the Most Preferable Seed Source for Silviculture of Abies sachalinensis in Hokkaido, Japan. Forests 2020, 11, 1058. https://doi.org/10.3390/f11101058
Tsuyama I, Ishizuka W, Kitamura K, Taneda H, Goto S. Ten Years of Provenance Trials and Application of Multivariate Random Forests Predicted the Most Preferable Seed Source for Silviculture of Abies sachalinensis in Hokkaido, Japan. Forests. 2020; 11(10):1058. https://doi.org/10.3390/f11101058
Chicago/Turabian StyleTsuyama, Ikutaro, Wataru Ishizuka, Keiko Kitamura, Haruhiko Taneda, and Susumu Goto. 2020. "Ten Years of Provenance Trials and Application of Multivariate Random Forests Predicted the Most Preferable Seed Source for Silviculture of Abies sachalinensis in Hokkaido, Japan" Forests 11, no. 10: 1058. https://doi.org/10.3390/f11101058
APA StyleTsuyama, I., Ishizuka, W., Kitamura, K., Taneda, H., & Goto, S. (2020). Ten Years of Provenance Trials and Application of Multivariate Random Forests Predicted the Most Preferable Seed Source for Silviculture of Abies sachalinensis in Hokkaido, Japan. Forests, 11(10), 1058. https://doi.org/10.3390/f11101058