Characterization and Justification of Trees on an Inner-City Golf Course in Halifax, Canada: An Investigation into the Ecological Integrity of Institutional Greenspace



Abstract
1. Introduction
2. Literature Review
The Acadian Forest Context
3. Materials and Methods
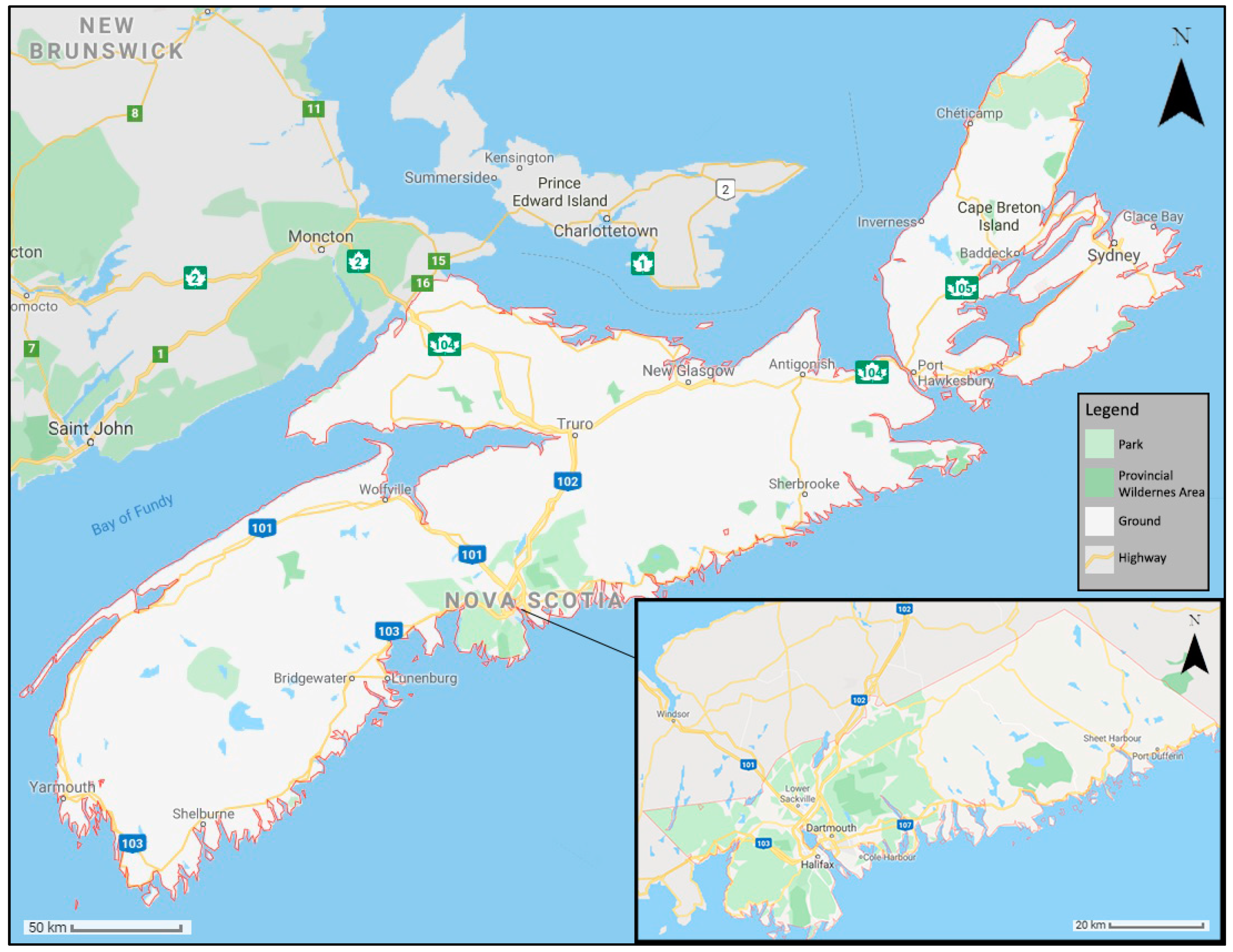
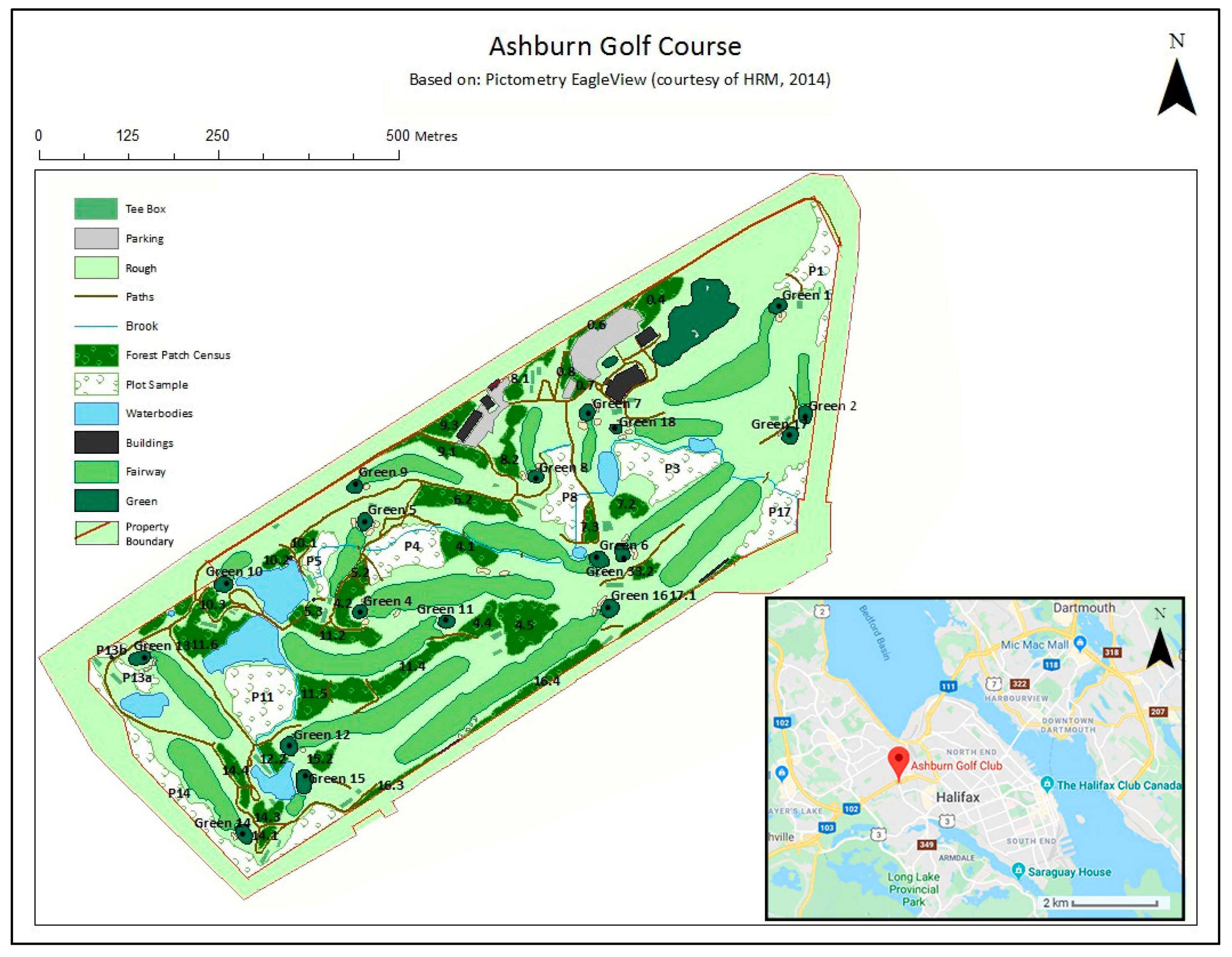
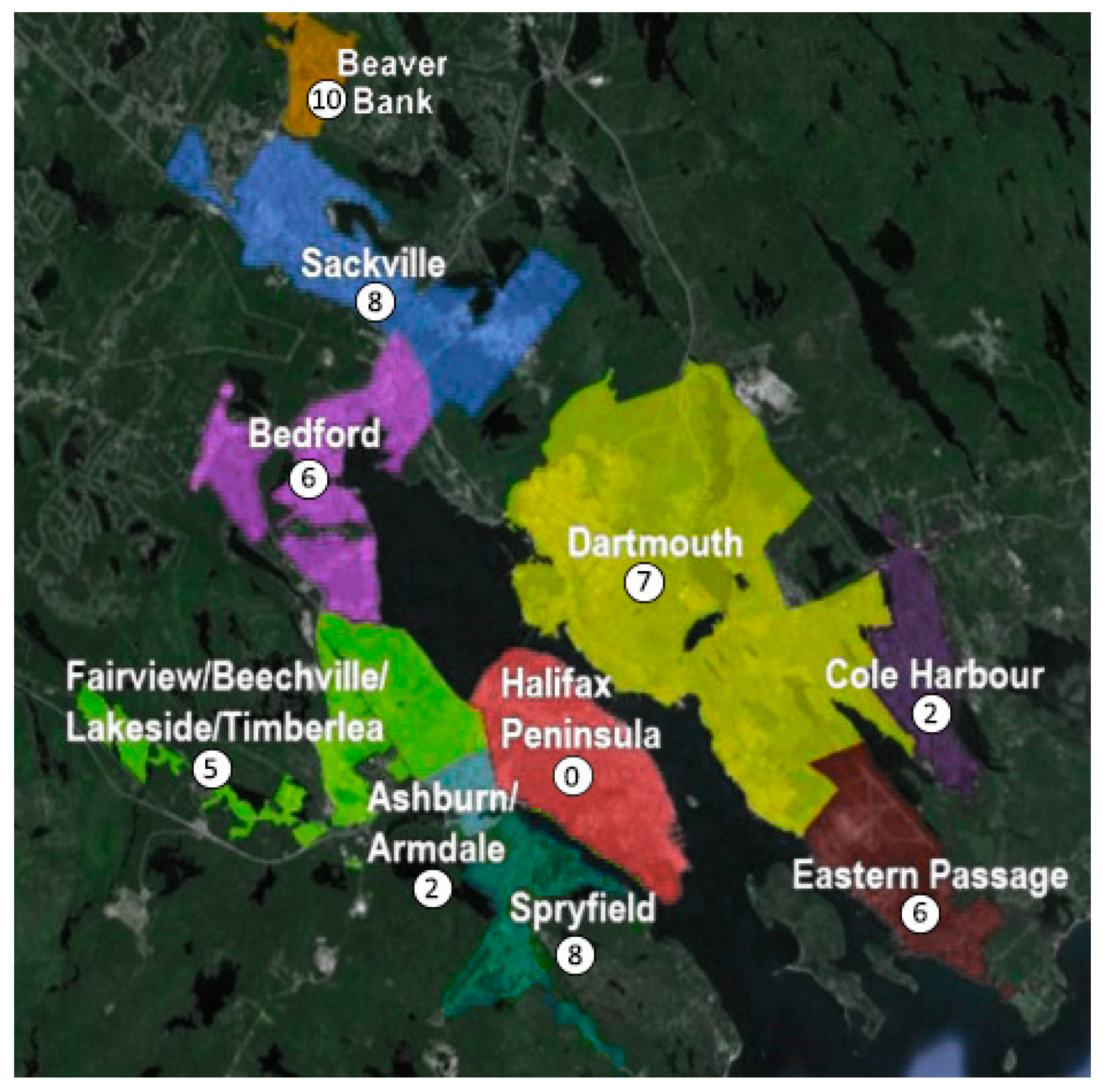
3.1. Study Area
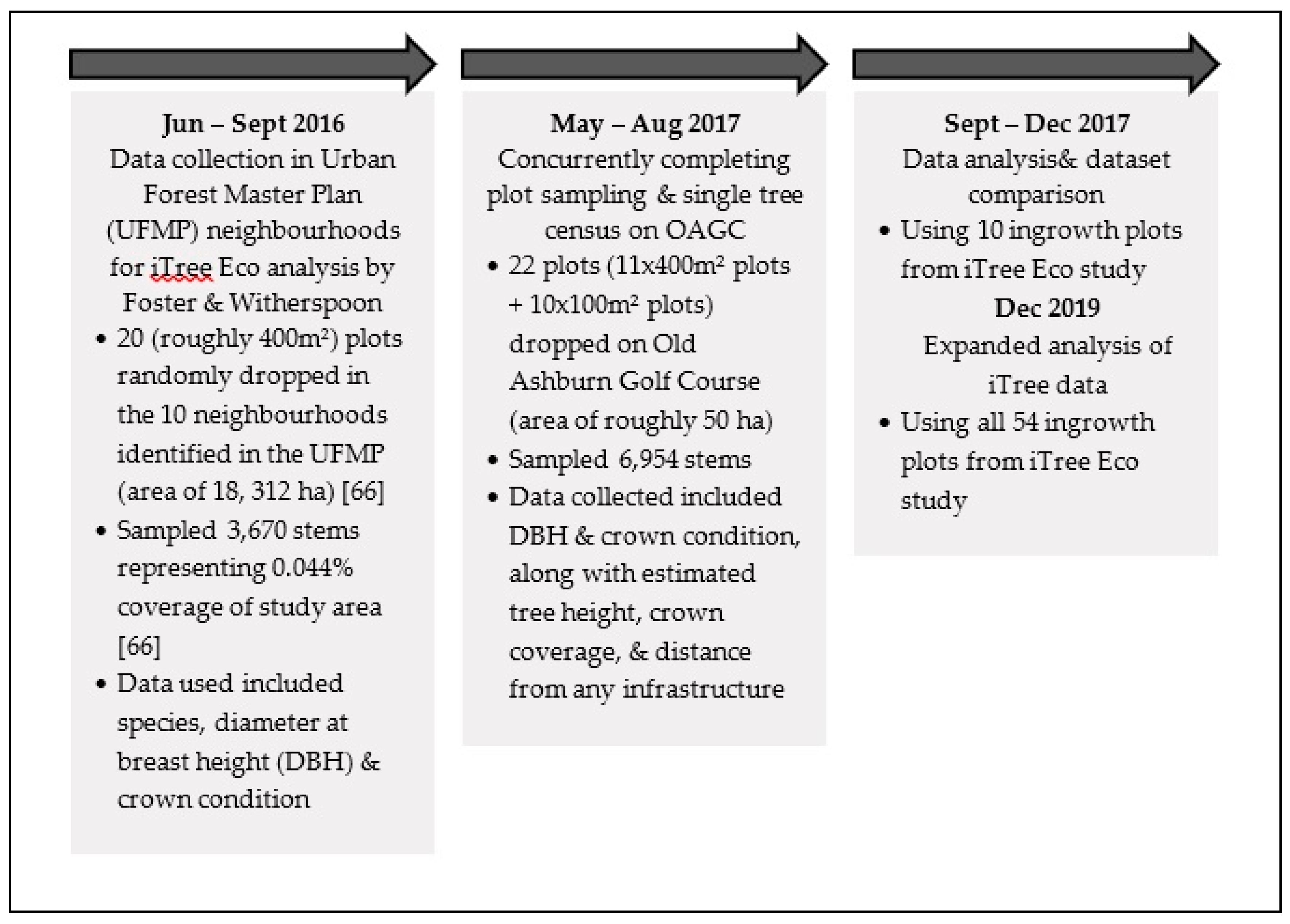
3.2. Inventory Overview
3.3. Comparative Analysis: Data Selection
4. Results and Discussion
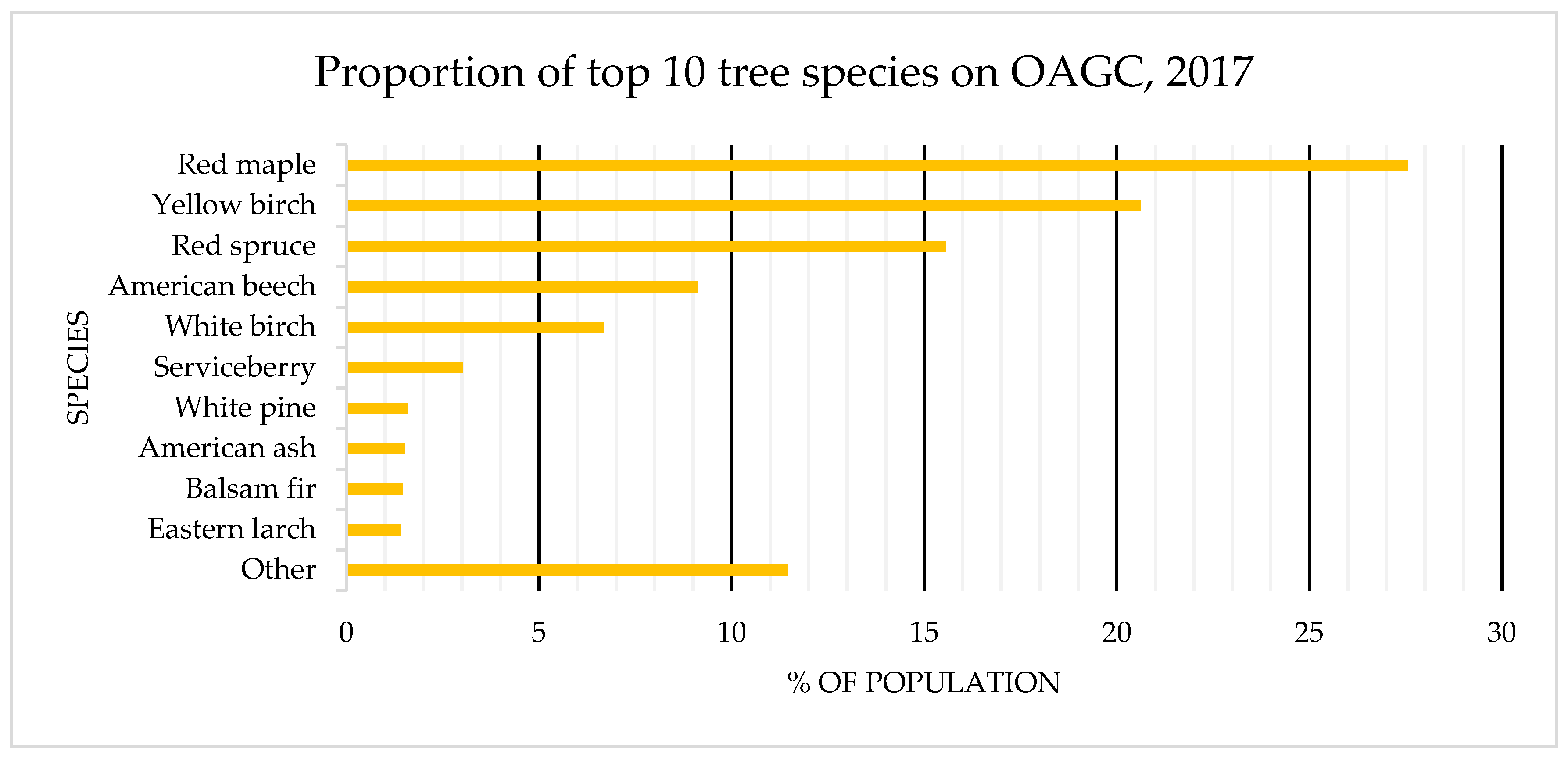
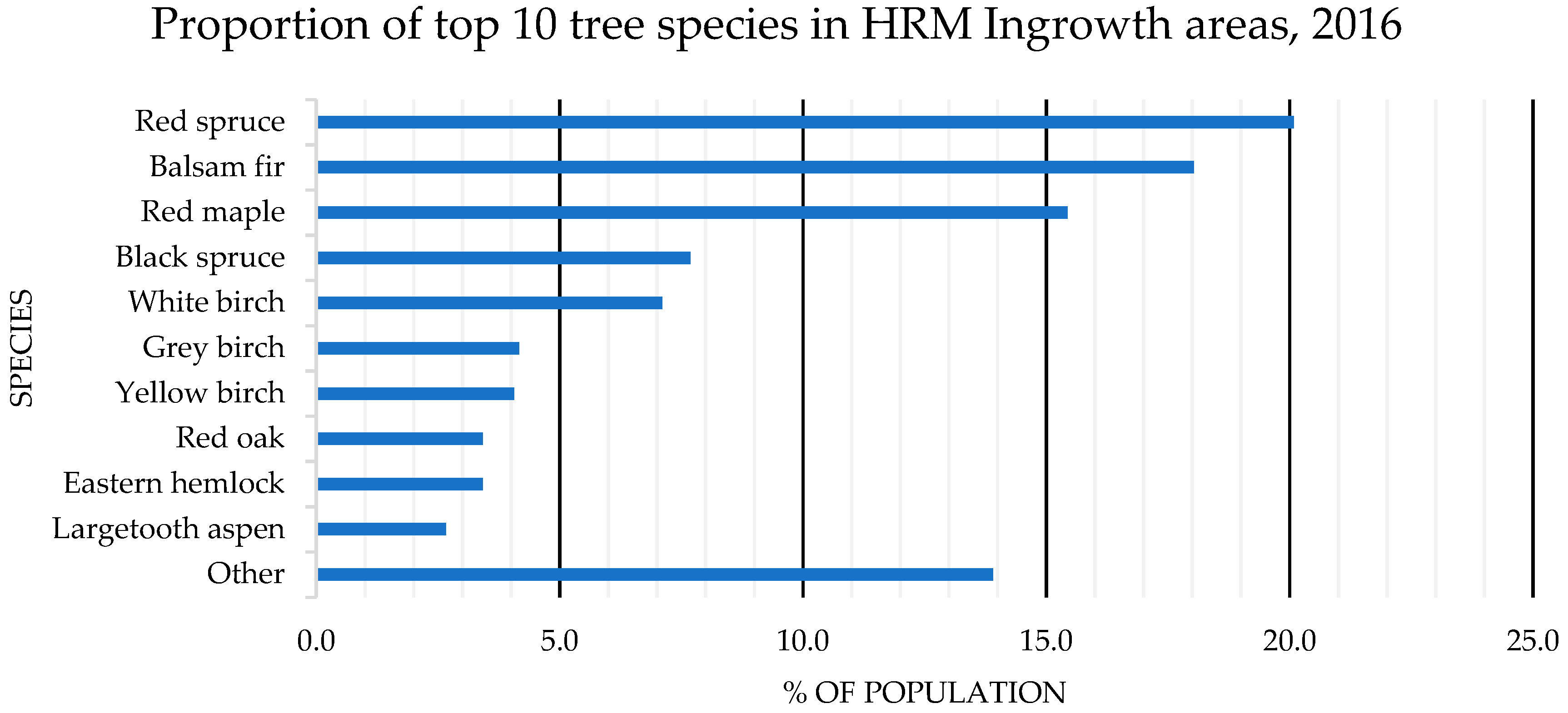
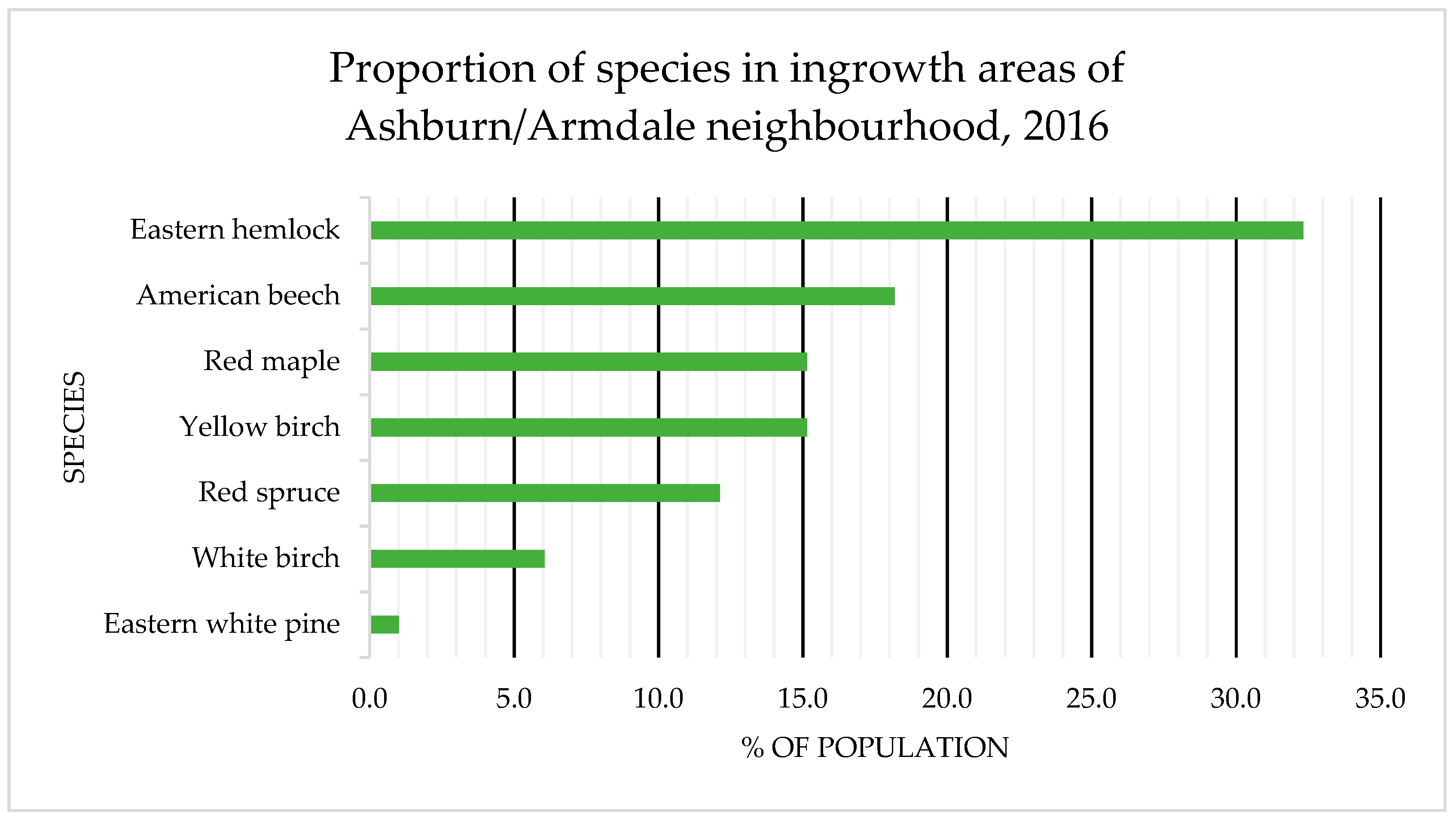
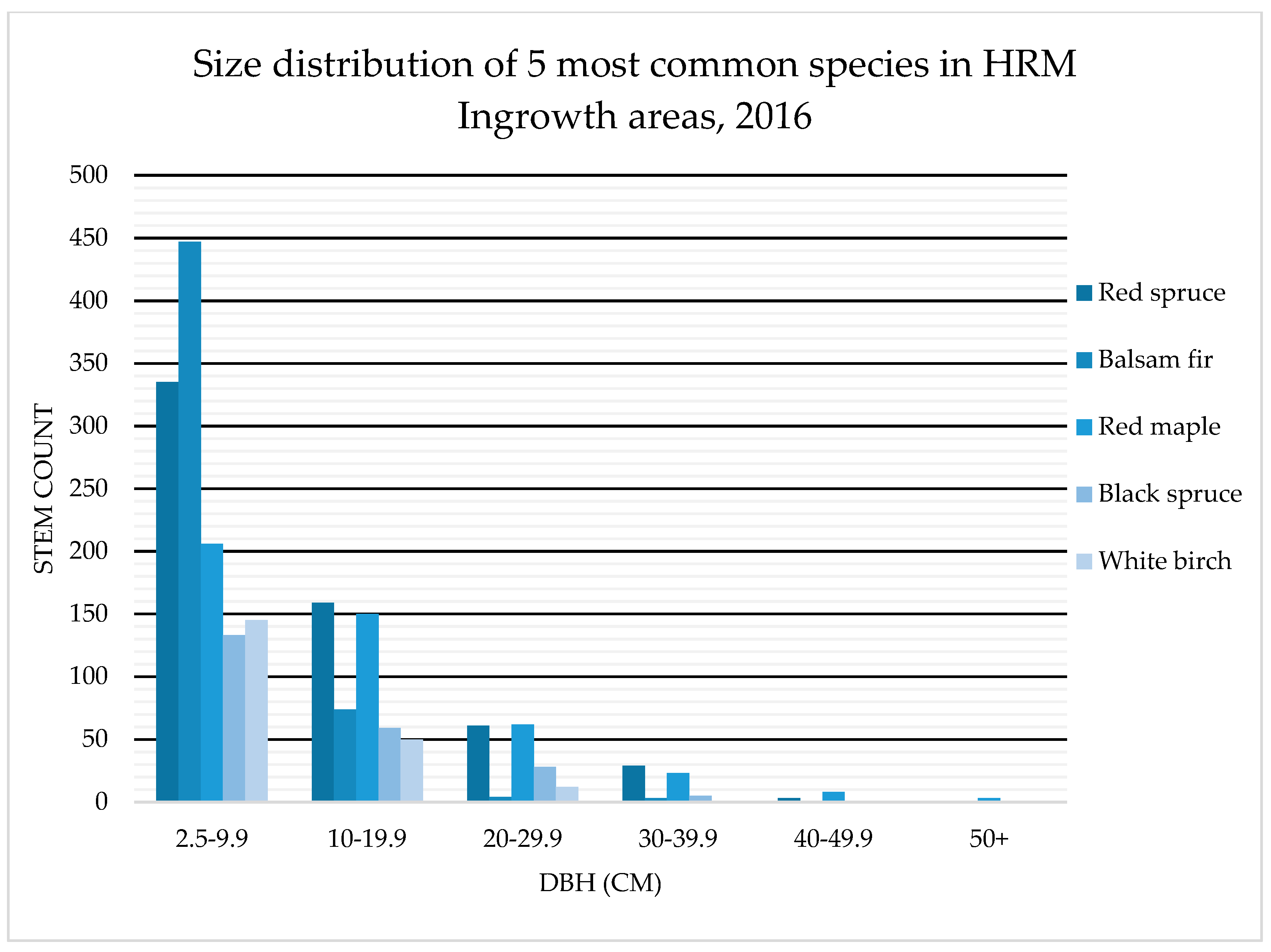
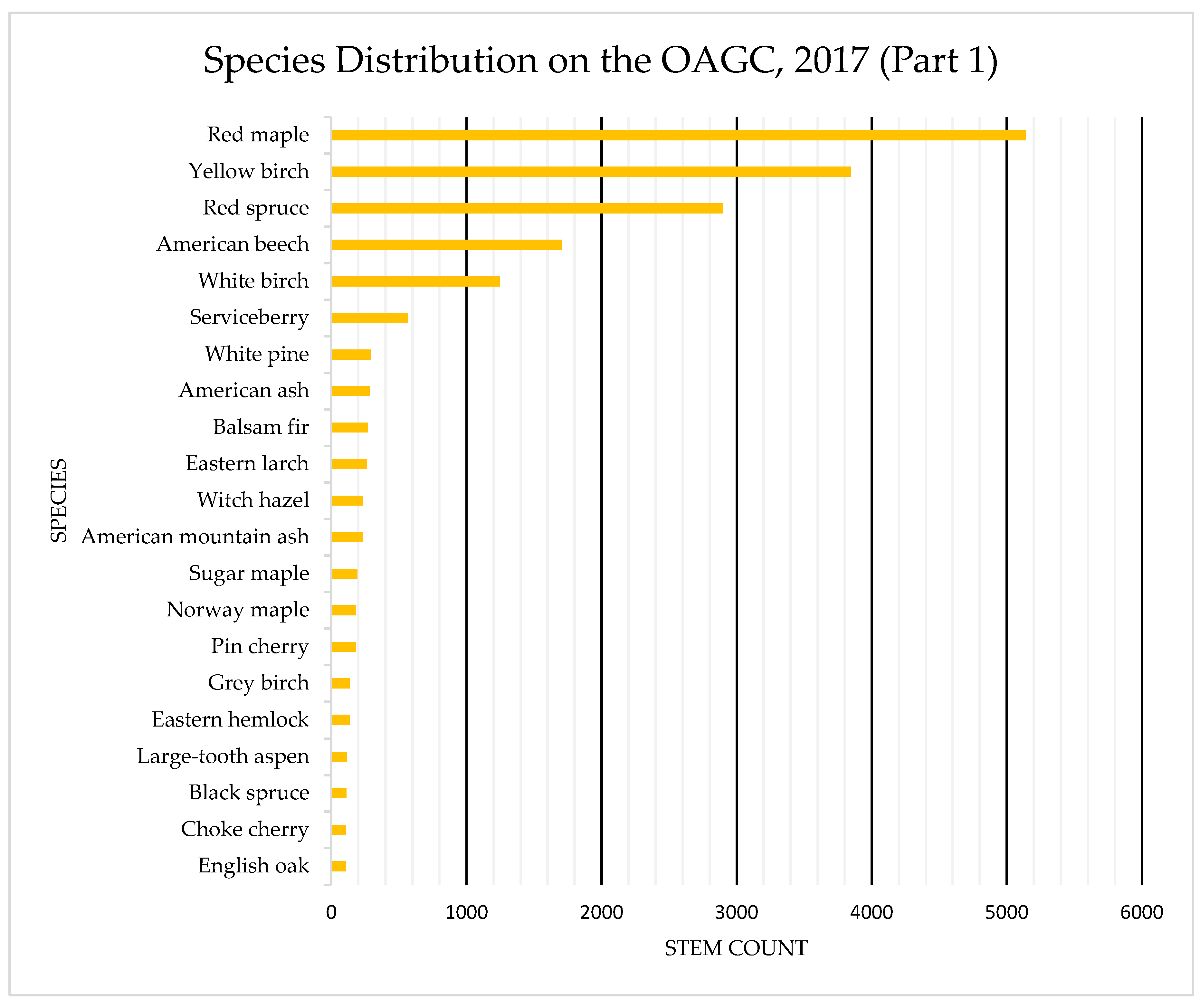
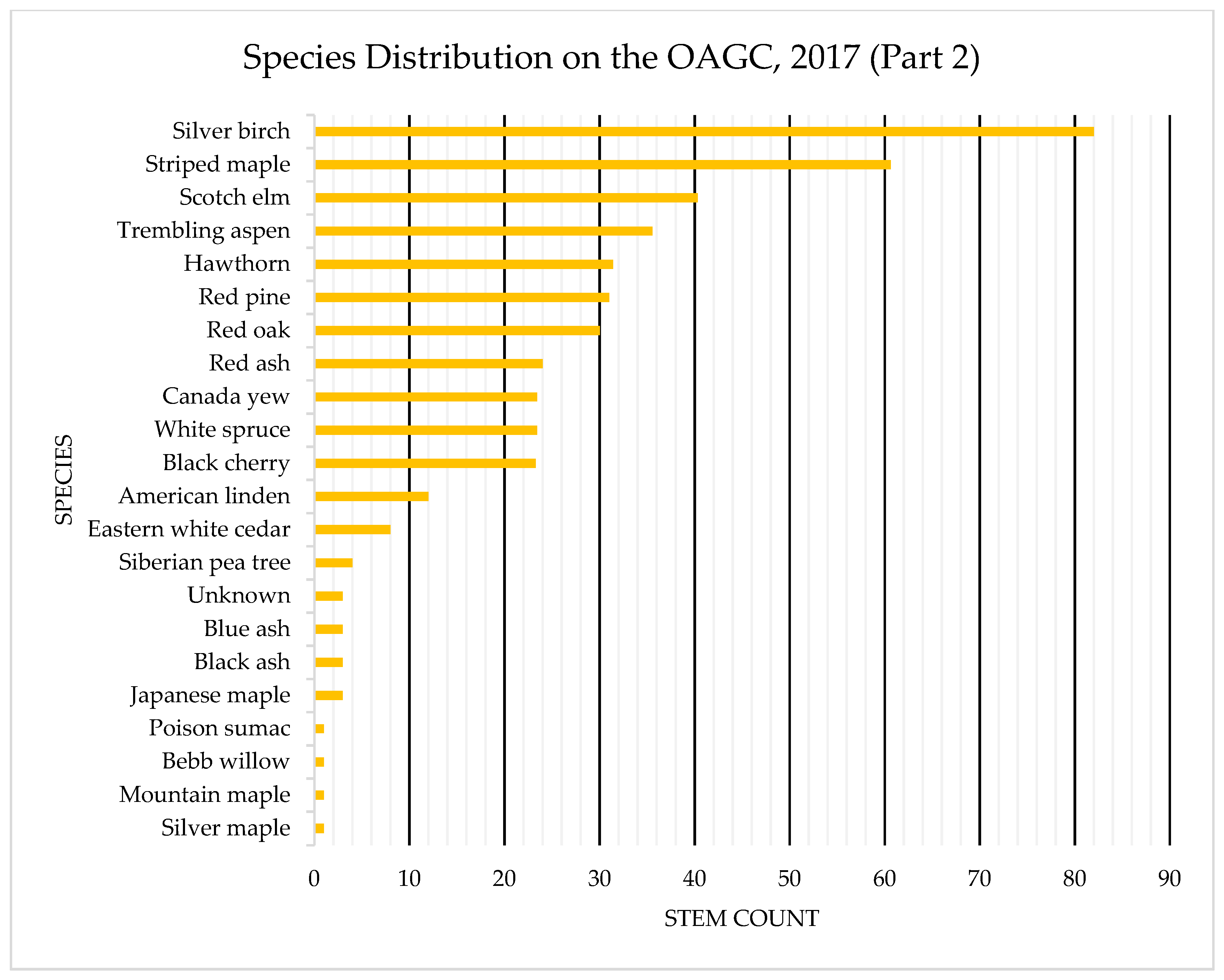
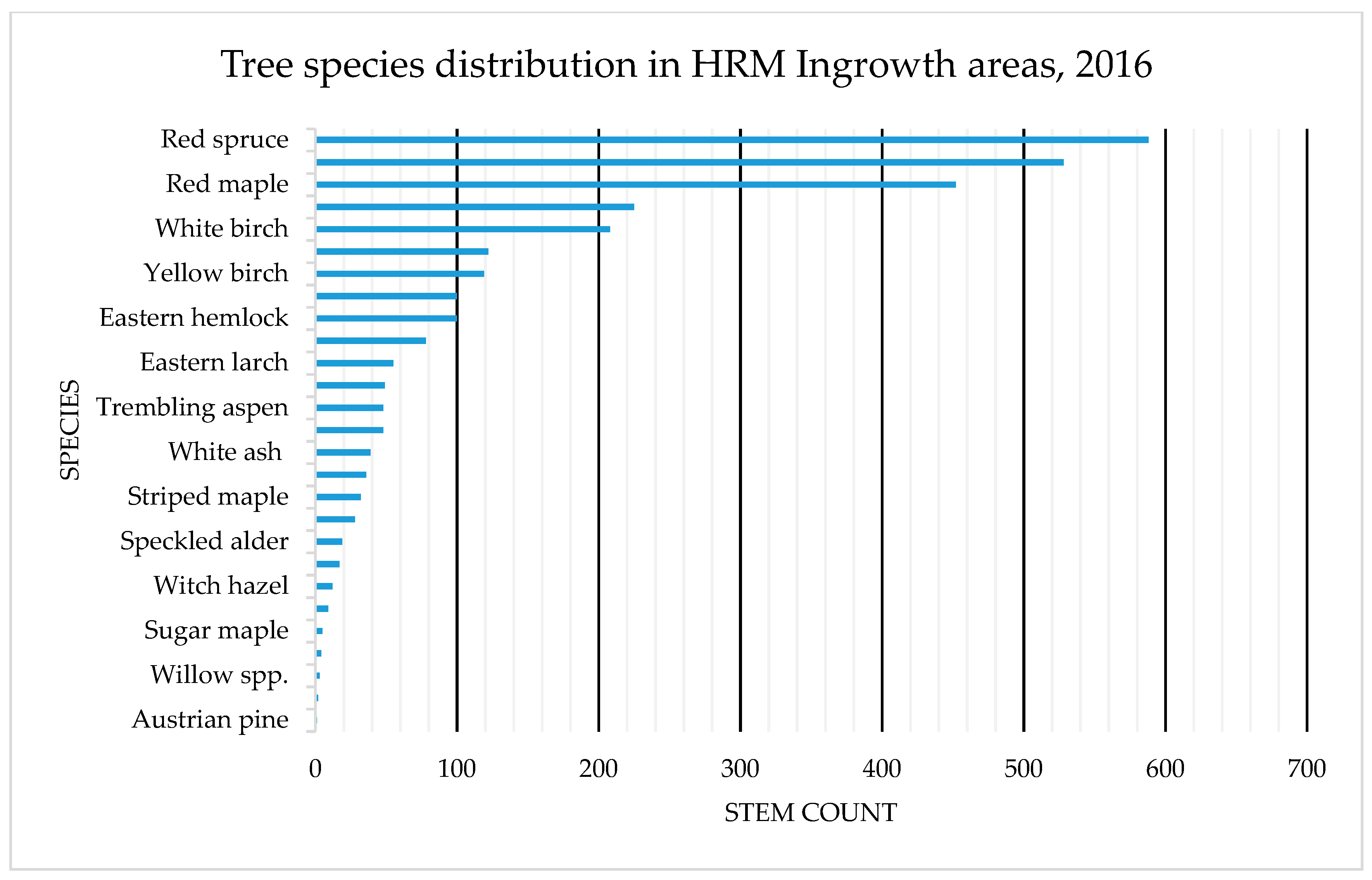
4.1. Species Composition: OAGC versus Ingrowth
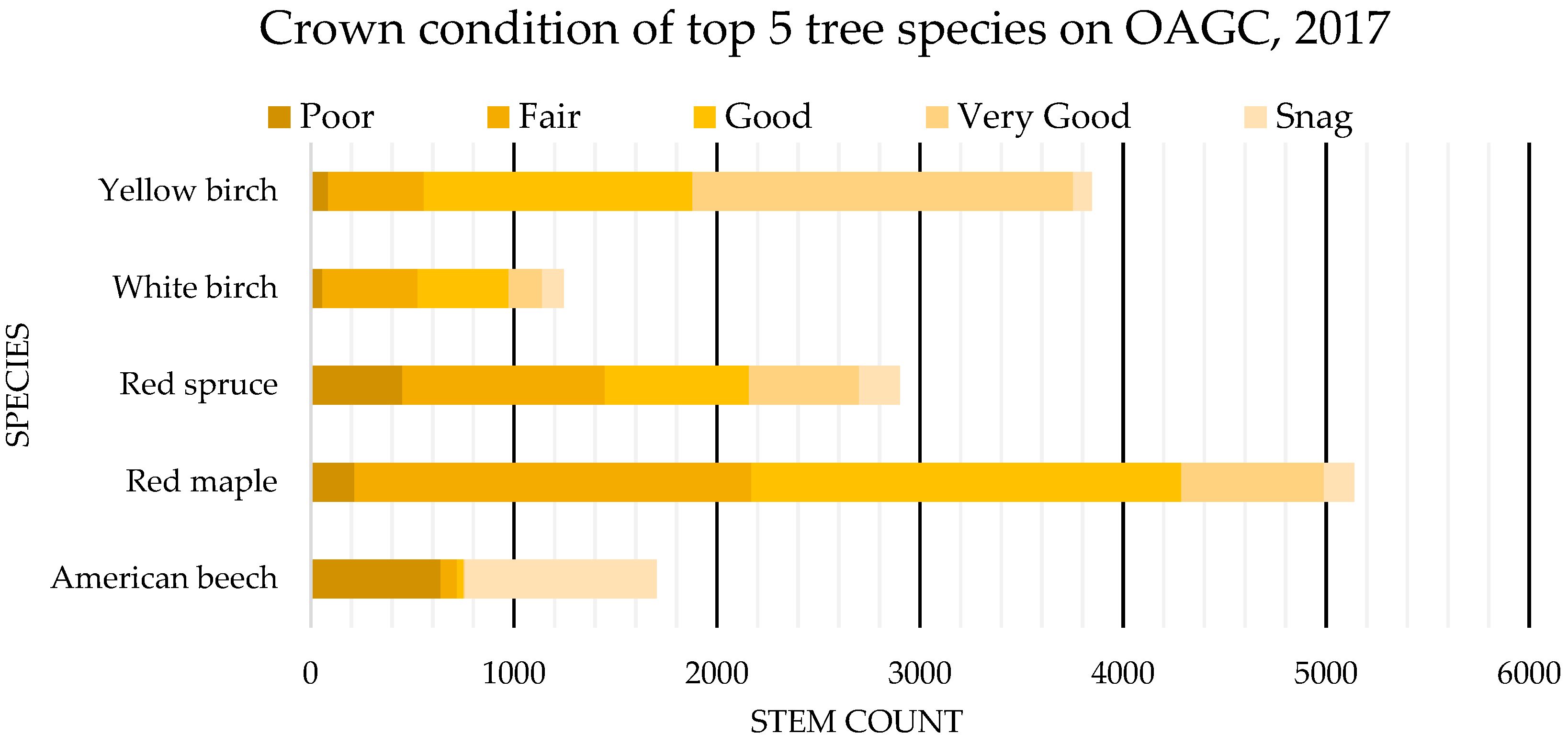
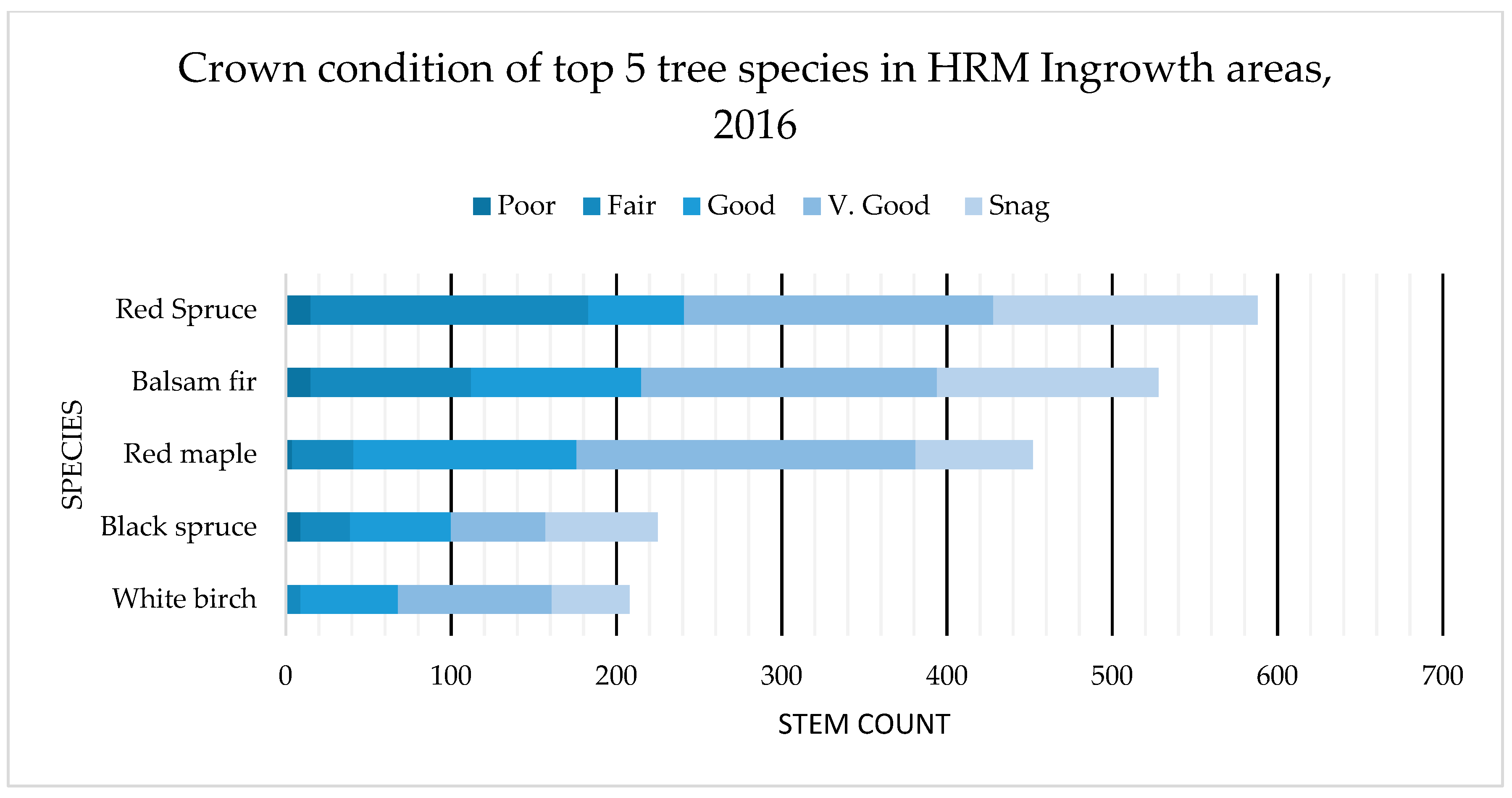
4.2. Tree Health: OAGC Versus Ingrowth
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
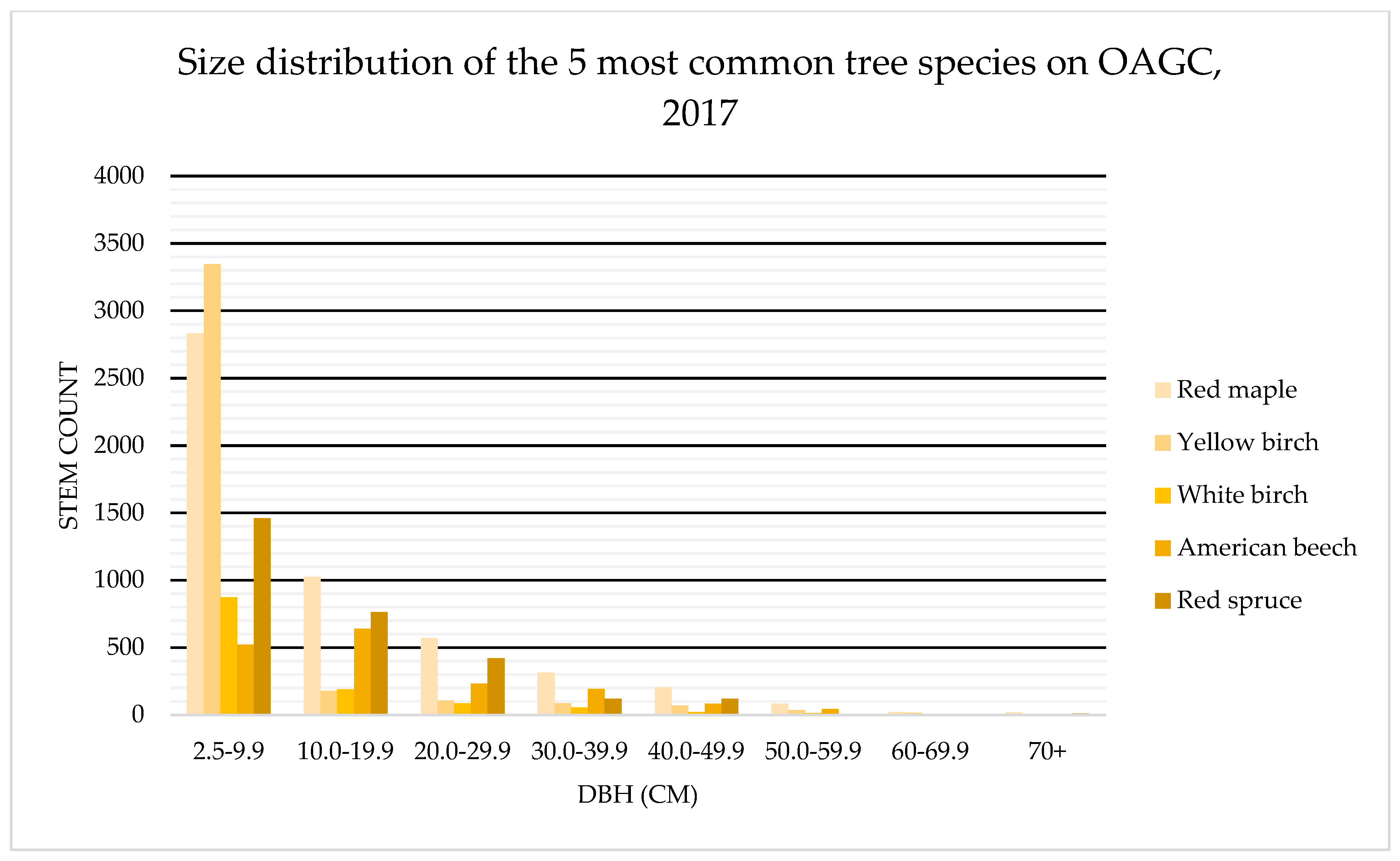{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Broadleaf Species | |
|---|---|
| Latin Name | English Name |
| Acer pensylvanicum | Striped maple |
| Acer rubrum | Red maple |
| Acer saccharinum | Silver maple |
| Acer saccharum | Sugar maple |
| Acer spicatum | Mountain maple |
| Amelanchier spp. | Serviceberry |
| Alnus incana | Speckled alder |
| Betula alleghaniensis | Yellow birch |
| Betula papyrifera | White birch |
| Betula populifolia | Grey birch |
| Fagus grandifolia | American beech |
| Fraxinus americana | White ash |
| Fraxinus nigra | Black ash |
| Populus grandidentata | Largetooth aspen |
| Populus tremuloides | Trembling aspen |
| Prunus pensylvanica | Pin cherry |
| Prunus serotine | Black cherry |
| Prunus virginiana | Choke cherry |
| Quercus alba | White oak |
| Quercus rubra | Red oak |
| Sorbus americana | American mountain ash |
| Tilia americana | Common linden (Basswood) |
| Ulmus americana | White (American) elm |
| Conifer Species | |
| Latin Name | English Name |
| Abies balsamea | Balsam fir |
| Larix laricina | Eastern larch |
| Picea glauca | White spruce |
| Picea mariana | Black spruce |
| Picea rubens | Red spruce |
| Pinus resinosa | Red pine |
| Pinus banksiana | Jack pine |
| Pinus strobus | Eastern white pine |
| Thuja occidentalis | Eastern white cedar |
| Tsuga canadensis | Eastern hemlock |
Appendix B. Tree Species Distribution on OAGC and HRM Ingrowth Areas



References
- Cilliers, S.S.; Seibert, S.J. Urban flora and vegetation: Patterns and processes. In Urban Ecology: Patterns, Processes, and Applications; Niemelä, J., Breuste, J.H., Elmqvist, T., Gutenspergen, G., James, P., McIntyre, N.E., Eds.; Oxford University Press: New York, NY, USA, 2011; pp. 148–158. [Google Scholar]
- Ordóñez, C.; Duinker, P.N. Ecological integrity in urban forests. Urban Ecosyst. 2012, 15, 863–877. [Google Scholar] [CrossRef]
- Endreny, T.; Santagata, R.; Perna, A.; De Stefano, C.; Rallo, R.F.; Ulgiati, S. Implementing and managing urban forests: A much needed conservation strategy to increase ecosystem services and urban wellbeing. Ecol. Model. 2017, 360, 328–335. [Google Scholar] [CrossRef]
- Toni, S.A.; Duinker, P.N. A framework for urban–woodland naturalization in Canada. Environ. Rev. 2015, 23, 321–336. [Google Scholar] [CrossRef]
- Gange, A.; Lindsay, D.; Schofield, M. The ecology of golf courses. Biologist 2003, 50, 63–68. [Google Scholar]
- Petrosillo, I.; Valente, D.; Pasimeni, M.R.; Aretano, R.; Smeraro, T.; Zurlini, G. Can a golf course support biodiversity and ecosystem services? The landscape context matter. Landsc. Ecol. 2019, 34, 2213–2228. [Google Scholar] [CrossRef]
- Yang, Y.; He, Z.; Wang, Y.; Fan, J.; Liang, Z.; Stoffella, P.J. Dissolved organic matter in relation to nutrients (N and P) and heavy metals in surface runoff water as affected by temporal variation and land uses—A case study from Indian River Area, south Florida, USA. Agric. Water Manag. 2013, 118, 38–49. [Google Scholar] [CrossRef]
- Nakayama, T.; Watanabe, M.; Tanji, K.; Morioka, T. Effect of underground urban structures on eutrophic coastal environment. Sci. Total Environ. 2007, 373, 270–288. [Google Scholar] [CrossRef]
- Udeigwe, T.K.; Young, J.; Kandakji, T.; Weindorf, D.C.; Mahmoud, M.A.; Stietiya, M.H. Elemental quantification, chemistry, and source apportionment in golf course facilities in a semi-arid urban landscape using a portable X-ray fluorescence spectrometer. Solid Earth 2015, 6, 415–424. [Google Scholar] [CrossRef][Green Version]
- Streeter, M.T.; Schilling, K.E. Effects of golf course management on subsurface soil properties in Iowa. Soil 2018, 4, 93–100. [Google Scholar] [CrossRef]
- Foster, D.; Witherspoon, K. i-Tree Eco Analysis Database. Unpublished raw data. 2016. [Google Scholar]
- Rowe, J.S. Forest Regions of Canada; Department of the Environment, Canadian Forestry Service Publisher No. 1300: Ottawa, ON, Canada, 1972. Available online: http://www.cfs.nrcan.gc.ca/publications/?id=24040 (accessed on 10 September 2017).
- Loo, J.; Ives, N. The Acadian forest: Historical condition and human impacts. For. Chron. 2003, 79, 462–474. [Google Scholar] [CrossRef]
- Quinton, J.M.; Duinker, P.N. Beyond burial: Researching and managing cemeteries as urban green spaces, with examples from Canada. Environ. Rev. 2019, 27, 252–262. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Ossola, A.; Hahs, A.K.; Williams, N.S.G.; Wilson, L.; Livesley, S.J. Variation in vegetation structure and composition across urban green space types. Front. Ecol. Evol. 2016, 4, 1–12. [Google Scholar] [CrossRef]
- Jarrett, M.; Shackleton, C.M. Integrating biodiversity considerations into urban golf courses: managers’ perceptions and woody plant diversity in the Eastern Cape, South Africa. J. Land Use Sci. 2017, 12, 292–311. [Google Scholar] [CrossRef]
- Nooten, S.S.; Schultheiss, P.; Wright, J.; Macdonald, C.; Singh, B.K.; Cook, J.M.; Power, S.A. What shapes plant and animal diversity on urban golf courses? Urban Ecosyst. 2018, 21, 565–576. [Google Scholar] [CrossRef]
- Guzmán, C.A.P.; Fernández, D.J.M. Environmental impacts by golf courses and strategies to minimize them: State of the art. Int. J. Arts Sci. 2014, 7, 403–417. [Google Scholar]
- Golf Course Superintendents Association of America. Golf Course Environmental Profile; GCSAA Press: Lawrence, KS, USA, 2007; Available online: https://www.golf coursesaa.org/uploadedfiles/Environment/Environmental-Profile/Nutrient/Golf-Course-Environmental-Profile--Nutrient-Management-Report.pdf (accessed on 11 October 2017).
- Golf Canada & The PGA of Canada. Golf Facilities in Canada: The Definitive Report of Golf Facilities and Development in Canada. Available online: http://golfcanada.ca/app/uploads/2015/07/NGF-GOLF-CANADA_ENGLISH-Final-July8.pdf (accessed on 20 November 2017).
- Baris, R.D.; Cohen, S.Z.; Barnes, N.L.; Lam, J.; Ma, Q. Quantitative analysis of over 20 years of golf course monitoring studies. Environ. Toxicol. Chem. 2010, 29, 1224–1236. [Google Scholar] [CrossRef]
- Larson, E.K.; Perrings, C. The value of water-related amenities in an arid city: The case of the Phoenix metropolitan area. Landsc. Urban Plan. 2013, 109, 45–55. [Google Scholar] [CrossRef]
- Golf Course Information. Types of Golf Courses. Available online: http://www.golf-information.info/types-of-golf-courses.html (accessed on 21 November 2017).
- Terman, M.R. Natural links: Naturalistic golf courses as wildlife habitat. Landsc. Urban Plan. 1997, 38, 183–197. [Google Scholar] [CrossRef]
- Yasuda, M.; Koike, F. Do golf courses provide a refuge for flora and fauna in Japanese urban landscapes? Landsc. Urban Plan. 2006, 75, 58–68. [Google Scholar] [CrossRef]
- Jim, C.; Chen, W. Legacy effect of trees in the heritage landscape of a peri-urban golf course. Urban Ecosyst. 2016, 19, 1717–1734. [Google Scholar] [CrossRef]
- Hodgkison, S.C.; Hero, J.M.; Warnken, J. The conservation value of suburban golf courses in a rapidly urbanising region of Australia. Landsc. Urban Plan. 2007, 79, 323–337. [Google Scholar] [CrossRef]
- Guerena, K.B.; Castelli, P.M.; Nichols, T.C.; Williams, C.K. Spatially-explicit land use effects on nesting of Atlantic Flyway resident Canada geese in New Jersey. Wildl. Biol. 2016, 20, 115–121. [Google Scholar] [CrossRef]
- Burgin, S.; Wutherspoon, D. The potential for golf courses to support restoration of biodiversity for BioBanking offsets. Urban Ecosyst. 2008, 12, 145–155. [Google Scholar] [CrossRef]
- Colding, J.; Folke, C. The role of golf courses in biodiversity conservation and ecosystem management. Ecosystems 2009, 12, 191–206. [Google Scholar] [CrossRef]
- Colding, J.; Lundberg, J.; Lundberg, S.; Andersson, E. Golf courses and wetland fauna. Ecol. Appl. 2009, 19, 1481–1491. [Google Scholar] [CrossRef]
- Berrill, M.; Bertram, S.; Wilson, A.; Louis, S.; Brigham, D.; Stromberg, C. Lethal and sublethal impacts of pyrethroid insecticides on amphibian embryos and tadpoles. Environ. Toxicol. Chem. 1993, 12, 525–539. [Google Scholar] [CrossRef]
- Rice, P.J.; Horgan, B.P.; Hamlin, J.L. Evaluation of individual and combined management practices to reduce the off-site transport of pesticides from golf course turf. Sci. Total Environ. 2017, 583, 72–80. [Google Scholar] [CrossRef]
- Hunter, A.; Kealy, E.; Forrest, M. The current use of indigenous trees on Irish golf courses. Acta Hortic. 2010, 885, 153–160. [Google Scholar] [CrossRef]
- Ashburn Golf Club. Club History. Available online: http://www.ashburngolfclub.com/clubhistory.html (accessed on 1 November 2017).
- Jiggens, M. Turf must trump trees on golf courses. Moeller Turf Rec. 8 March 2013. Available online: https://www.turfandrec.com/agronomy/turf-must-trump-trees-on-golf-courses-moeller-3074 (accessed on 20 September 2017).
- Gross, P. The Seven Dirty Words of Golf Course Maintenance. Available online: http://www.wctaturf.com/topics/golf/goct99b.html (accessed on 21 November 2017).
- Boving, T.B.; Klement, J.; Rowell, R.; Xing, B. Effectiveness of wood and bark in removing organic and inorganic contaminates from aqueous solution. Mol. Cryst. Liq. Cryst. 2008, 483, 339–347. [Google Scholar] [CrossRef]
- Bartens, J.; Day, S.D.; Harris, J.R.; Wynn, T.M.; Dove, J.E. Transpiration and root development of urban trees in structural soil stormwater reservoirs. Environ. Manag. 2009, 44, 646–657. [Google Scholar] [CrossRef]
- Selhorst, A.L.; Lal, R. Carbon sequestration in golf course turfgrass systems and recommendations for the enhancement of climate change mitigation potential. In Carbon Sequestration in Urban Ecosystems; Lal, R., Augustin, B., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 249–263. ISBN 978-94-007-2366-5. [Google Scholar]
- Bartlett, M.D.; James, I.T. A model of greenhouse gas emissions from the management of turf on two golf courses. Sci. Total Environ. 2011, 409, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Snow, J.T. Trees—The Biggest Problem of Golf Course Turf. Available online: http://archive.lib.msu.edu/tic/mitgolf course/article/199385.pdf (accessed on 23 November 2017).
- Nicholson, G. The Case against Trees. Available online: http://scoregolf.com/feature/the-case-against-trees/ (accessed on 24 November 2017).
- Evergreen. Keeping It Green: A Citizen’s Guide to Urban Land Protection in Canada. Available online: https://www.evergreen.ca/downloads/pdfs/Keeping-It-Green.pdf (accessed on 10 April 2019).
- Baur, J.W.R.; Tynon, J.F. Small-scale urban nature parks: Why should we care? Leis. Sci. 2010, 32, 195–200. [Google Scholar] [CrossRef]
- Carmichael, C.E.; McDonough, M.H. Community stories: Explaining resistance to street tree-planting programs in Detroit, Michigan, USA. Soc. Nat. Resour. 2019, 32, 588–605. [Google Scholar] [CrossRef]
- Lorentz, D.; Getherall, E.; Bein, E.; Beditz, J. Golf and the Millennial Generation. Available online: https://golf2020.com/wp-content/uploads/2017/11/millennial_report_ngf.pdf (accessed on 27 March 2019).
- Youde, K. Rough times for golf as Britain’s amateurs turn their backs on the sport. Independent. 23 August 2014. Available online: http://www.independent.co.uk/sport/golf/rough-times-for-golf-as-britains-amateurs-turn-their-backs-on-the-sport-9687900.html (accessed on 10 April 2019).
- Harwell, D. Why America fell out of love with golf. Washington Post. 5 March 2015. Available online: https://www.washingtonpost.com/news/wonk/wp/2015/03/05/why-america-fell-out-of-love-with-golf/?utm_term=.73f8cc970faa (accessed on 10 April 2019).
- Robinson, P. Golf in Canada: Not a cheap pursuit. CBC Sports. 24 May 2012. Available online: http://www.cbc.ca/sports-content/golf/opinion/2012/05/golf-in-canada-not-a-cheap-pursuit.html (accessed on 10 April 2019).
- Cain, P. Golfers are Disappearing from Public City Courses. Should Some be Made into Parks? Global News. 10 April 2016. Available online: https://globalnews.ca/news/2612990/as-fewer-golfers-use-public-courses-should-some-be-made-into-parks/ (accessed on 21 November 2017).
- Javed, N. GTA housing developers targeting golf courses. Toronto Star. 27 March 2016. Available online: https://www.thestar.com/news/gta/2016/03/27/gta-housing-developers-targeting-golf-courses.html (accessed on 10 April 2019).
- Mercer. Mercer’s Annual Cost of Living Survey Finds Asian, European, and African Cities Most Expensive Locations for Employees. Available online: https://www.mercer.com/newsroom/cost-of-living-2018.html (accessed on 30 November 2017).
- Halliday, W.E.D. A Forest Classification for Canada; Forest Service, Department of Mines and Resources: Ottawa, ON, Canada, 1937. Available online: http://cfs.nrcan.gc.ca/pubwarehouse/pdfs/26268.pdf (accessed on 19 February 2019).
- Farrar, J.L. Trees in Canada, 2nd ed.; Fitzhenry and Whiteside: Markham, ON, Canada, 2017; ISBN 978-1554554065. [Google Scholar]
- Natural Resources Canada. Forest Classification. Available online: http://www.nrcan.golf course.ca/forests/measuring-reporting/classification/13179 (accessed on 25 November 2017).
- Davis, M.; Gratton, L.; Adams, J.; Goltz, J.; Stewart, C.; Buttrick, S.; Zinger, N.; Kavanagh, K.; Sims, M.; Mann, G. New England—Acadian Forests. Available online: https://www.worldwildlife.org/ecoregions/na0410 (accessed on 25 March 2019).
- Duinker, P.N.; Ordóñez, C.; Steenberg, J.W.N.; Miller, K.H.; Toni, S.A.; Nitoslawski, S. Trees in Canadian cities: Indispensable life form for urban sustainability. Sustainability 2015, 7, 7379–7396. [Google Scholar] [CrossRef]
- Halifax Regional Municipality. Pictometry. Unpublished raw data. 2014. [Google Scholar]
- Avery, T.E.; Burkhart, H.E. Forest Measurements, 5th ed.; McGraw Hill: Toronto, ON, Canada, 2015; ISBN 978-1478629085. [Google Scholar]
- Neighborwoods. Quick Reference Guide. Available online: https://harbordvillage.com/wp-content/uploads/2016/06/TreesNeighbourwoodsQuickReferenceGuide.pdf (accessed on 8 December 2019).
- Van Laar, A.; Akça, A. Sampling for forest inventories. In Forest Mensuration: Managing Forest Ecosystems, 2nd ed.; Springer: Dordrecht, The Netherlands, 1997; pp. 229–315. ISBN 978-1-4020-5991-9. [Google Scholar]
- Nowak, D.J.; Walton, J.T.; Stevens, J.C.; Crane, D.E.; Hoehn, R.E. Effect of plot and sample size on timing and precision of urban forest assessments. Arboric. Urban For. 2008, 34, 386–390. [Google Scholar]
- USDA Forest Service. i-Tree Eco User’s Manual. USDA Forest Service. 2 July 2019. Available online: https://www.itreetools.org/resources/manuals/Ecov6_ManualsGuides/Ecov6_UsersManual.pdf (accessed on 30 May 2017).
- Halifax Regional Municipality. Halifax Regional Municipality Urban Forest Master Plan. Available online: https://www.halifax.ca/sites/default/files/documents/transportation/streetssidewalks/HALREG%201246%20UrbanForestReport_HighRes_SINGLEPAGE_Mon20_Combined.pdf (accessed on 8 December 2019).
- Foster, D.; Duinker, P.N. The HRM Urban Forest in 2016; School for Resource and Environmental Studies, Dalhousie University: Halifax, NS, Canada, 2017; Available online: https://www.itreetools.org/resources/reports/FosterDuinker_2017_iTreeEcoForHalifax_Feb2017.pdf (accessed on 15 September 2017).
- Neily, P.; Keys, K.; Quigley, E.; Basquill, S.; Stewart, B. Forest Ecosystem Classification for Nova Scotia; Renewable Resources Branch (Report 2013-1); Nova Scotia Department of Natural Resources: Truro, NS, Canada, 2010. Available online: https://novascotia.ca/natr/forestry/veg-types/ (accessed on 30 May 2017).
- Snyder, M. What is a climax forest? Northern Woodlands Magazine. 1 September 2006. Available online: http://northernwoodlands.org/articles/article/what_is_a_climax_forest (accessed on 19 August 2017).
- Mosseler, A.; Lynds, J.A.; Major, J.E. Old-growth forests of the Acadian Forest Region. Environ. Rev. 2003, 11, S47–S77. [Google Scholar] [CrossRef]
- Canadian Council of Forest Ministers. Forest Pest Monitoring in Canada: Current Situation, Compatibilities, Gaps, and Proposed Enhanced Monitoring Program (Cat. No.: Fo79-8/2012E-PDF); Forest Service Publications: Ottawa, ON, Canada, 2012; ISBN 978-1-100-20920-3.
- Stephanson, C.; Ribarik Coe, N. Impacts of Beech Bark Disease and Climate Change on American Beech. Forests 2017, 8, 155. [Google Scholar] [CrossRef]
- Natural Resources Canada. Beech Leaf-Mining Weevil. Canadian Food Inspection Agency, Impact Note No. 061, 2016. Available online: https://cfs.nrcan.gc.ca/publications?id=37778 (accessed on 25 November 2017).
- Government of Canada. Hemlock Woolly Adelgid Confirmed in Nova Scotia. Canadian Food Inspection Agency, News Release, 2017. Available online: https://www.canada.ca/en/food-inspection-agency/news/2017/08/hemlock_woolly_adelgidconfirmedinnovascotia.html (accessed on 12 December 2019).
- Zurita, G.; Pe’er, G.; Bellocq, M.I.; Hansbauer, M.M. Edge effects and their influence on habitat suitability calculations: A continuous approach applied to birds of the Atlantic forest. J. Appl. Ecol. 2012, 49, 503–512. [Google Scholar] [CrossRef]
- Bolund, P.; Hunhammar, S. Ecosystem services in urban areas. Ecol. Econ. 1999, 29, 293–301. [Google Scholar] [CrossRef]
- Nowak, D.J. Institutionalizing urban forestry as a “biotechnology” to improve environmental quality. Urban For. Urban Green. 2006, 5, 93–100. [Google Scholar] [CrossRef]
- Hesler, G.M. Health impacts of ultraviolet radiation in urban ecosystems: A review. Urban Ecosyst. 2000, 4, 193–229. [Google Scholar] [CrossRef]
- Grinde, B.; Patil, G.G. Biophilia: Does visual contact with nature impact on health and well-being? Int. J. Environ. Res. Public Health 2009, 6, 2332–2343. [Google Scholar] [CrossRef] [PubMed]
- Natural Resources Canada. Assisted Migration. Available online: https://www.nrcan.gc.ca/climate-change/impacts-adaptations/climate-change-impacts-forests/adaptation/assisted-migration/13121 (accessed on 24 December 2019).












| Crown Condition | |
|---|---|
| Snag (0) | Dead Tree |
| Poor (1) | Significant defoliation (0%–50% remaining canopy), weak/discolored foliage |
| Fair (2) | Some defoliation (50%–74% remaining canopy), some weak/discoloring foliage |
| Good (3) | Minor defoliation (75%–84% remaining canopy), mostly healthy foliage |
| Very Good (4) | No defoliation (85%–100%), healthy foliage |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doll, S.; Duinker, P.N. Characterization and Justification of Trees on an Inner-City Golf Course in Halifax, Canada: An Investigation into the Ecological Integrity of Institutional Greenspace. Forests 2020, 11, 96. https://doi.org/10.3390/f11010096
Doll S, Duinker PN. Characterization and Justification of Trees on an Inner-City Golf Course in Halifax, Canada: An Investigation into the Ecological Integrity of Institutional Greenspace. Forests. 2020; 11(1):96. https://doi.org/10.3390/f11010096
Chicago/Turabian StyleDoll, Shauna, and Peter N. Duinker. 2020. "Characterization and Justification of Trees on an Inner-City Golf Course in Halifax, Canada: An Investigation into the Ecological Integrity of Institutional Greenspace" Forests 11, no. 1: 96. https://doi.org/10.3390/f11010096
APA StyleDoll, S., & Duinker, P. N. (2020). Characterization and Justification of Trees on an Inner-City Golf Course in Halifax, Canada: An Investigation into the Ecological Integrity of Institutional Greenspace. Forests, 11(1), 96. https://doi.org/10.3390/f11010096