Growth, Nutrient Assimilation, and Carbohydrate Metabolism in Korean Pine (Pinus koraiensis) Seedlings in Response to Light Spectra




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Pre-Culture
2.2. Seedling Transplant
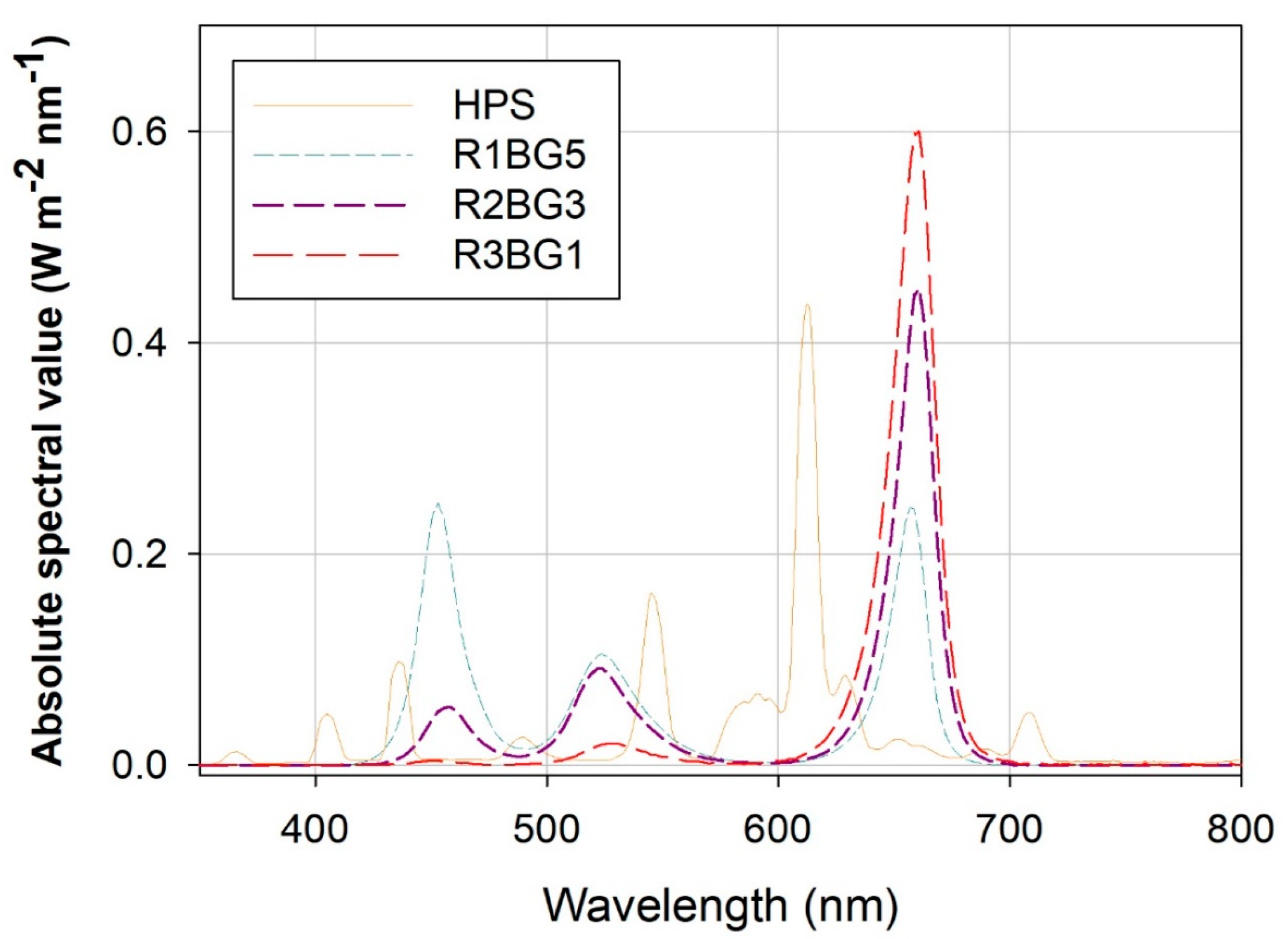
2.3. Illumination Treatment
2.4. Seedling Culture
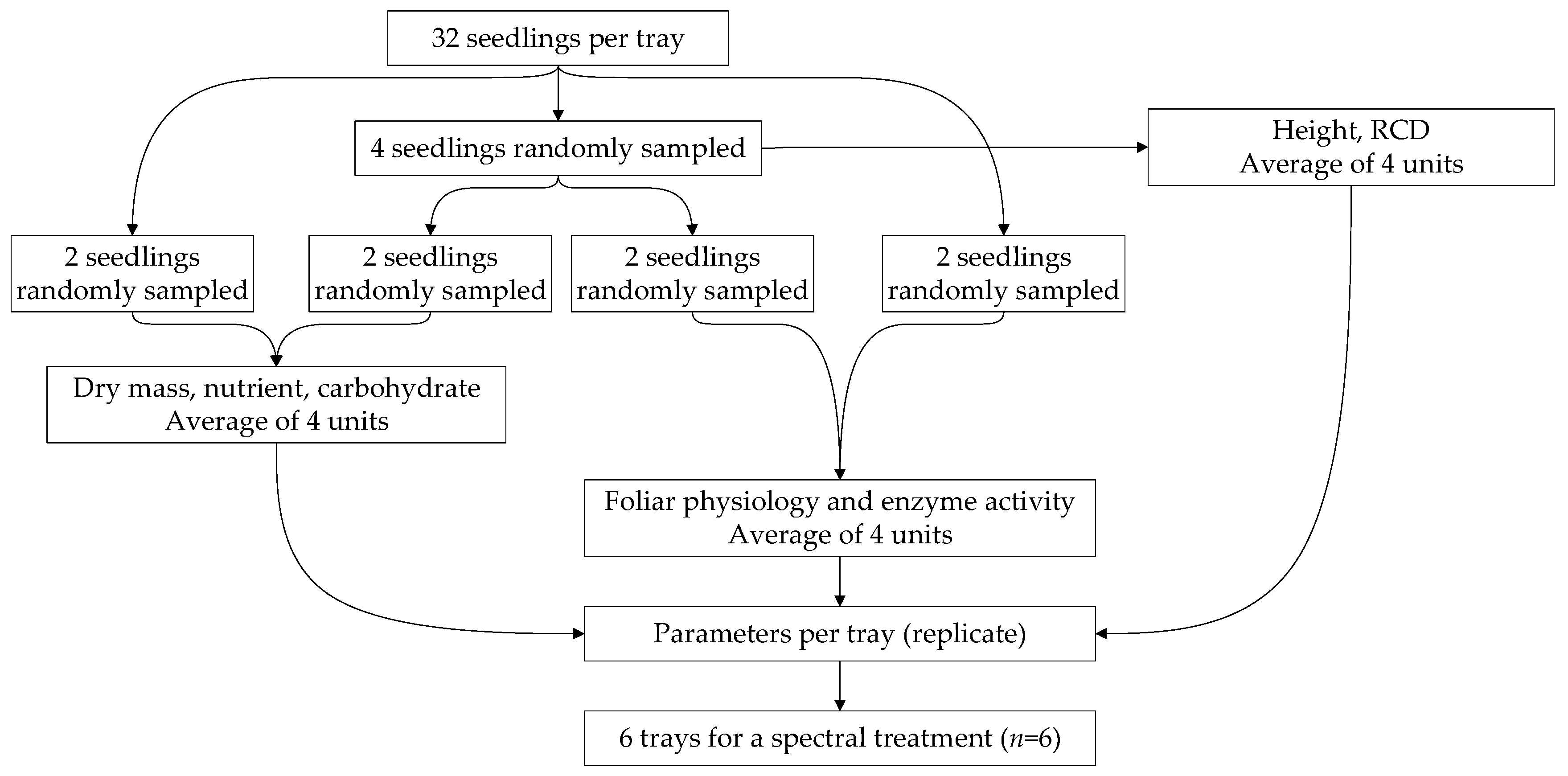
2.5. Seedling Sampling
2.6. Chemical Analysis
2.7. Indices Calculation
2.8. Statistical Analysis
3. Results
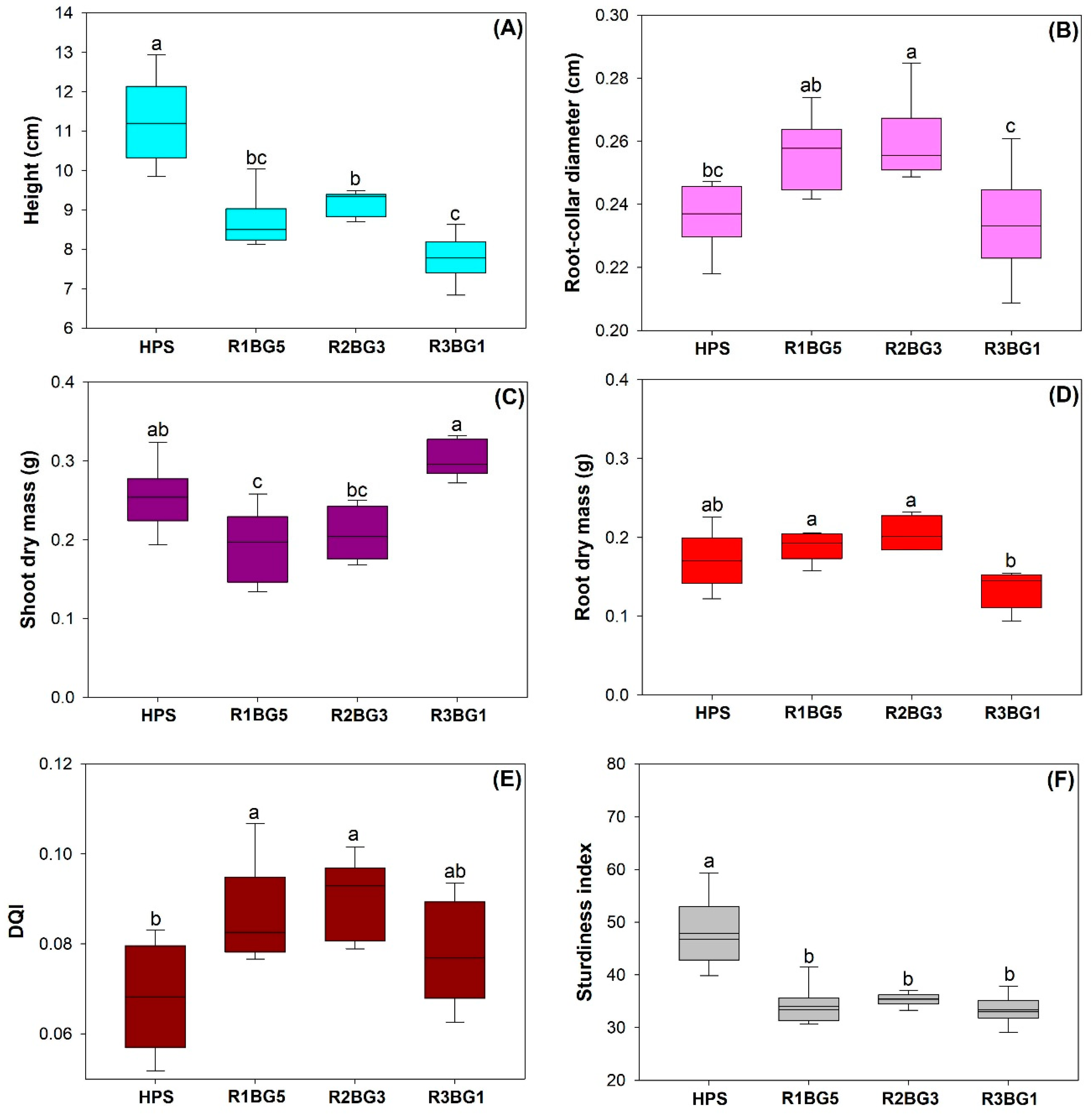
3.1. Seedling Growth, Biomass, and Quality Estimation
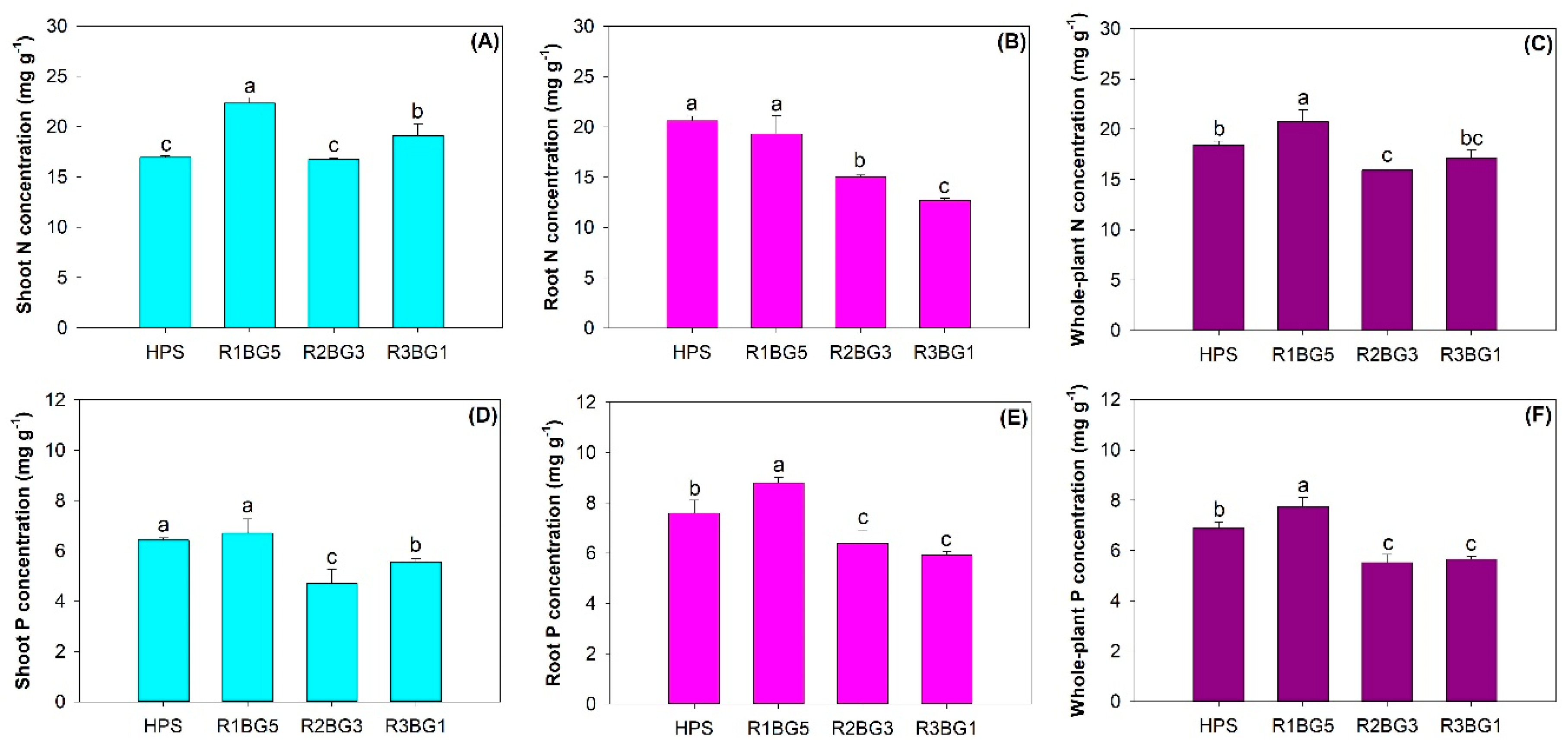
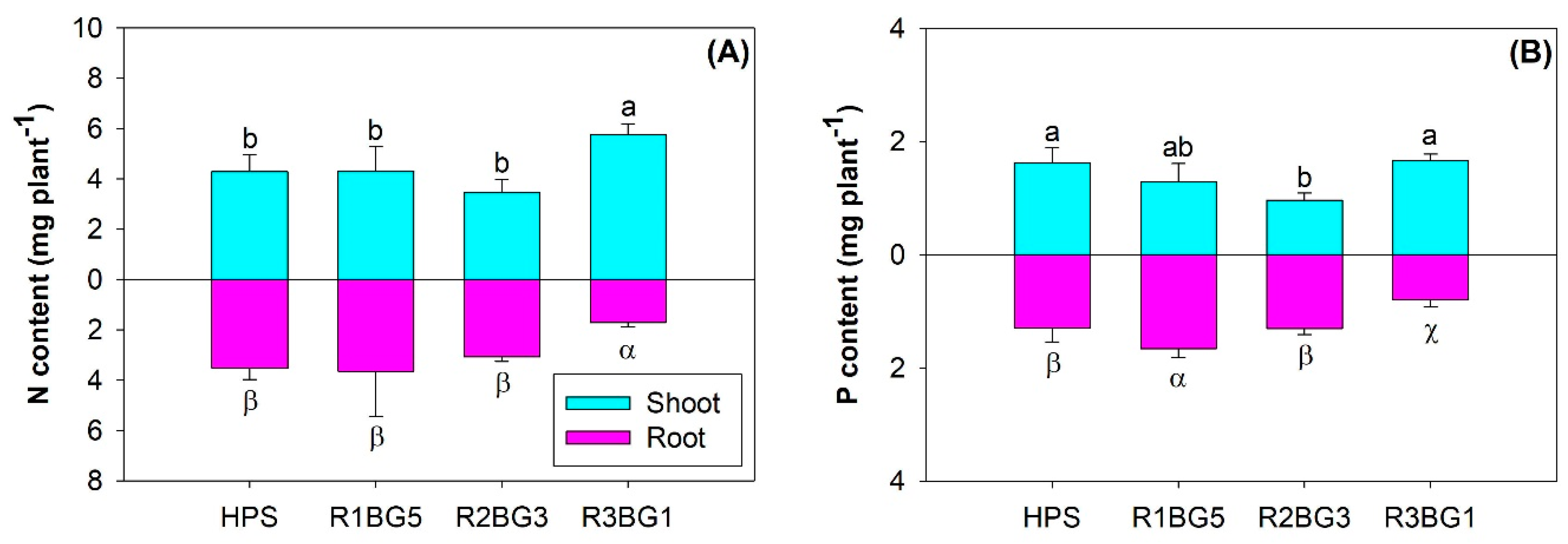
3.2. Nutrient Concentration and Content
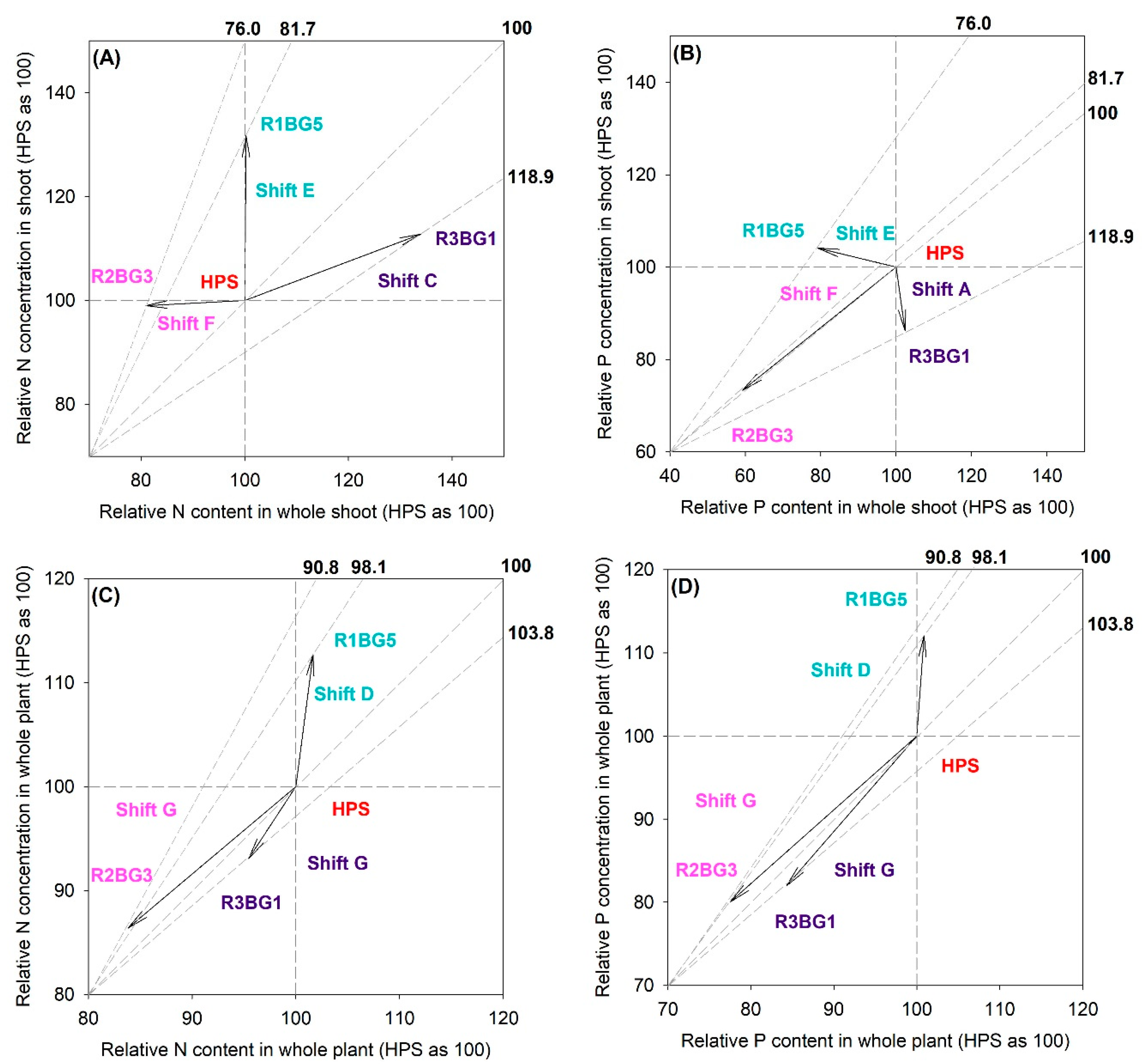
3.3. Vector Analysis on Nutritional Status
3.4. Physiological Responses
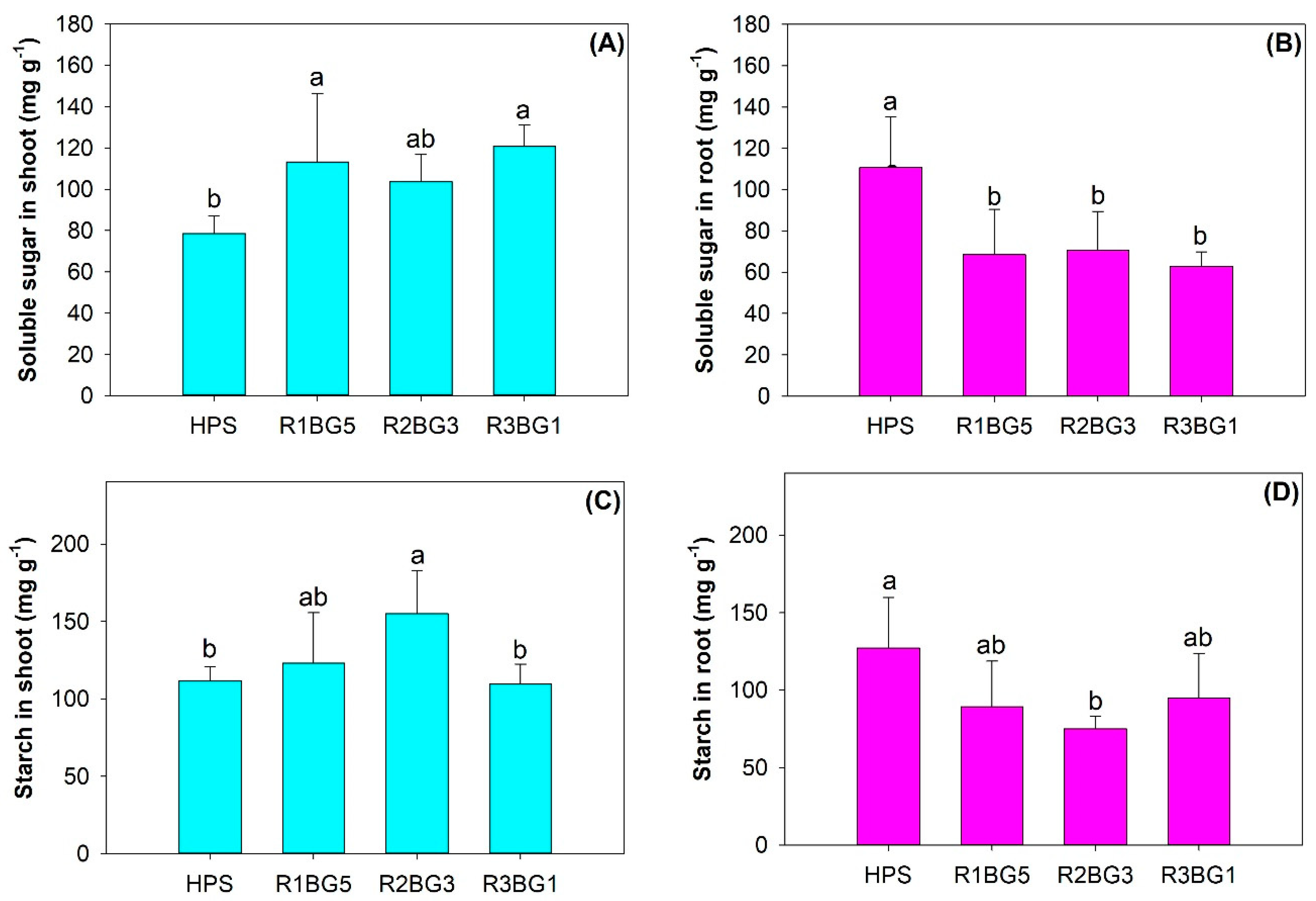
3.5. Carbohydrate Metabolism
4. Discussion
4.1. Growth, Biomass, and Seedling Quality
4.2. Nutrient Uptake and Allocation
4.3. Physiology and Carbohydrates
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, F.; Liang, D.Y.; Pei, X.N.; Zhang, Q.H.; Zhang, P.; Zhang, J.Q.; Lu, Z.M.; Yang, Y.C.; Liu, G.F.; Zhao, X.Y. Study on the physiological indices of Pinus siberica and Pinus koraiensis seedlings under cold stress. J. Res. 2019, 30, 1255–1265. [Google Scholar] [CrossRef]
- Zhang, M.; Yan, Q.L.; Zhu, J.J. Optimum light transmittance for seed germination and early seedling recruitment of Pinus koraiensis: Implications for natural regeneration. iForest 2015, 8, 853–859. [Google Scholar] [CrossRef]
- Zhao, G.Y.; Yang, C.T. Discussion of introducing Pinus koraiensis to the Greater Xing’an Mountains. Sci. Technol. 1991, 27, 149–153. (In Chinese) [Google Scholar]
- Zhu, J.J. A review on fundamental studies of secondary forest management. J. Appl. Ecol. 2002, 13, 1689–1694. (In Chinese) [Google Scholar]
- State Forestry Administration of China. Tree Seedling Quality Grading of Major Species for Afforestation; GB 6000-1999; State Bureau of Quality Technical Supervision: Beijing, China, 1999; p. 193.
- Li, G.; Zhu, Y.; Li, Q.; Liu, Y.; Zou, S.; Huang, Y. Effect of seedling age on the seedling quality and field performance of Pinus koraiensis. Sci. Silvae Sin. 2012, 48, 35–41, (In Chinese with English Abstract). [Google Scholar]
- Riikonen, J.; Kettunen, N.; Gritsevich, M.; Hakala, T.; Sarkka, L.; Tahvonen, R. Growth and development of Norway spruce and Scots pine seedlings under different light spectra. Environ. Exp. Bot. 2016, 121, 112–120. [Google Scholar] [CrossRef]
- Alakarppa, E.; Taulavuori, E.; Valledor, L.; Marttila, T.; Jokipii-Lukkari, S.; Karppinen, K.; Nguyen, N.; Taulavuori, K.; Haggman, H. Early growth of Scots pine seedlings is affected by seed origin and light quality. J. Plant Physiol. 2019, 237, 120–128. [Google Scholar] [CrossRef]
- Li, X.W.; Chen, Q.X.; Lei, H.Q.; Wang, J.W.; Yang, S.; Wei, H.X. Nutrient uptake and utilization by Fragrant rosewood (Dalbergia odorifera) seedlings cultured with oligosaccharide addition under different lighting spectra. Forests 2018, 9, 29. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, X.; Wei, H.X.; Lv, J.; Chen, C.; Liu, X.Y.; Wen, Q.; Jia, L.M. Nutrient uptake and utilization in Prince Rupprecht’s larch (Larix principis-rupprechtii Mayr.) seedlings exposed to a combination of light-emitting diode spectra and exponential fertilization. Soil Sci. Plant Nutr. 2019, 65, 358–368. [Google Scholar] [CrossRef]
- Riikonen, J. Pre-cultivation of Scots pine and Norway spruce transplant seedlings under four different light spectra did not affect their field performance. New For. 2016, 47, 607–619. [Google Scholar] [CrossRef]
- Gu, Y.Y.; Zhang, F.H.; Zeng, Y.L.; Zhang, L.; Tan, X.F.; Cao, H.P.; Li, Z. Physiological responses of Tung Tree (Vernicia fordii) saplings to different red, white and blue light-emitting diodes. Int. J. Agric. Biol. 2019, 22, 569–577. [Google Scholar]
- Smirnakou, S.; Ouzounis, T.; Radoglou, K.M. Continuous spectrum LEDs promote seedling quality traits and performance of Quercus ithaburensis var. macrolepis. Front. Plant Sci. 2017, 8, 16. [Google Scholar] [CrossRef]
- Velasco, M.H.; Mattsson, A. Light quality and intensity of light-emitting diodes during pre-cultivation of Picea abies (L.) Karst. and Pinus sylvestris L. Seedlings—Impact on growth performance, seedling quality and energy consumption. Scand. J. For. Res. 2019, 34, 159–177. [Google Scholar] [CrossRef]
- Sun, Y.R.; Zhu, J.J.; Sun, O.J.; Yan, Q.L. Photosynthetic and growth responses of Pinus koraiensis seedlings to canopy openness: Implications for the restoration of mixed-broadleaved Korean pine forests. Environ. Exp. Bot. 2016, 129, 118–126. [Google Scholar] [CrossRef]
- Zhang, M.; Zhu, J.; Yan, Q. Seed germination of Pinus koraiensis Siebold & Zucc. in response to light regimes caused by shading and seed positions. For. Syst. 2012, 21, 426–438. [Google Scholar]
- Wei, H.X.; Ren, J.; Zhou, J.H. Effect of exponential fertilization on growth and nutritional status in Buddhist pine (Podocarpus macrophyllus Thunb. D. Don) seedlings cultured in natural and prolonged photoperiods. Soil Sci. Plant Nutr. 2013, 59, 933–941. [Google Scholar] [CrossRef]
- Apostol, K.G.; Dumroese, R.K.; Pinto, J.R.; Davis, A.S. Response of conifer species from three latitudinal populations to light spectra generated by light-emitting diodes and high-pressure sodium lamps. Can. J. For. Res. 2015, 45, 1711–1719. [Google Scholar] [CrossRef]
- Wei, H.X.; Xu, C.Y.; Hawkins, B.J.; Ma, L.Y.; Jiang, L.N. Organic amendment and inorganic fertilization affect soil properties and quality of Larix olgensis bareroot stock. New For. 2012, 43, 155–168. [Google Scholar] [CrossRef]
- Li, X.W.; Gao, Y.; Wei, H.X.; Xia, H.T.; Chen, Q.X. Growth, biomass accumulation and foliar nutrient status in fragrant rosewood (Dalbergia odorifera TC Chen) seedlings cultured with conventional and exponential fertilizations under different photoperiod regimes. Soil Sci. Plant Nutr. 2017, 63, 153–162. [Google Scholar] [CrossRef]
- Timmer, V.R.; Armstrong, G. Growth and nutrition of containerized Pinus resinosa at exponentially increasing nutrient additions. Can. J. For. Res. 1987, 17, 644–647. [Google Scholar] [CrossRef]
- Wallin, E.; Grans, D.; Jacobs, D.F.; Lindstrom, A.; Verhoef, N. Short-day photoperiods affect expression of genes related to dormancy and freezing tolerance in Norway spruce seedlings. Ann. For. Sci. 2017, 74, 14. [Google Scholar] [CrossRef]
- Wei, H.X.; Zhao, H.T.; Chen, X. Foliar N:P stoichiometry in Aralia elata distributed on different slope degrees. Not. Bot. Hort. Agrobo. Cluj-Napoca 2019, 47, 887–895. [Google Scholar] [CrossRef]
- Dickson, A.; Leaf, A.L.; Hosner, J.F. Quality appraisal of white spruce and white pine seedling stock in nurseries. For. Chron. 1960, 36, 10–13. [Google Scholar] [CrossRef]
- Salifu, K.F.; Timmer, V.R. Optimizing nitrogen loading of Picea mariana seedlings during nursery culture. Can. J. For. Res. 2003, 33, 1287–1294. [Google Scholar] [CrossRef]
- Zhu, K.Y.; Liu, H.C.; Wei, H.X.; Zhou, J.H.; Zou, Q.C.; Ma, G.Y.; Zhang, J.Q. Prediction of nutrient leaching from culture of containerized Buddhist pine and Japanese maple seedlings exposed to extended photoperiod. Int. J. Agric. Biol. 2016, 18, 425–434. [Google Scholar] [CrossRef]
- Kim, H.H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Green-light supplementation for enhanced lettuce growth under red- and blue-light-emitting diodes. Hortscience 2004, 39, 1617–1622. [Google Scholar] [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hashida, S.; Yoshihara, T. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. Hortscience 2010, 45, 1809–1814. [Google Scholar] [CrossRef]
- Bayala, J.; Dianda, M.; Wilson, J.; Ouedraogo, S.J.; Sanon, K. Predicting field performance of five irrigated tree species using seedling quality assessment in Burkina Faso, West Africa. New Fors. 2009, 38, 309–322. [Google Scholar] [CrossRef]
- Bian, Z.H.; Cheng, R.F.; Wang, Y.; Yang, Q.C.; Lu, C.G. Effect of green light on nitrate reduction and edible quality of hydroponically grown lettuce (Lactuca sativa L.) under short-term continuous light from red and blue light-emitting diodes. Environ. Exp. Bot. 2018, 153, 63–71. [Google Scholar] [CrossRef]
- Astolfi, S.; Marianello, C.; Grego, S.; Bellarosa, R. Preliminary investigation of LED lighting as growth light for seedlings from different tree species in growth chambers. Not. Bot. Hort. Agrobo. Cluj-Napoca 2012, 40, 31–38. [Google Scholar] [CrossRef]
- Zhang, T.; Shi, Y.Y.; Piao, F.Z.; Sun, Z.Q. Effects of different LED sources on the growth and nitrogen metabolism of lettuce. Plant Cell Tissue Organ Cult. 2018, 134, 231–240. [Google Scholar] [CrossRef]
- Elmlinger, M.W.; Mohr, H. Glutamine synthetase in Scots pine seedlings and its control by blue light and light absorbed by phytochrome. Planta 1992, 188, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Meya, G.; Kowallik, W. Blue and red light-dependent alterations in the ratio of two forms of glutamine synthetase in Chlorella kessleri. Plant Biol. 1995, 108, 247–254. [Google Scholar]
- Naureen, Z.; Sham, A.; Al Ashram, H.; Gilani, S.A.; Al Gheilani, S.; Mabood, F.; Hussain, J.; Al Harrasi, A.; AbuQamar, S.F. Effect of phosphate nutrition on growth, physiology and phosphate transporter expression of cucumber seedlings. Plant Physiol. Biochem. 2018, 127, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Hong, K.; Yi, Y. Effect of short-term phosphate starvation on acid phosphatase activity of Carpinus pubescens and Eurycorymbus cavalerei. Russ. J. Plant Physiol. 2015, 62, 57–64. [Google Scholar] [CrossRef]
- Lasceve, G.; Leymarie, J.; Vavasseur, A. Alterations in light-induced stomatal opening in a starch-deficient mutant of Arabidopsis thaliana L deficient in chloroplast phosphoglucomutase activity. Plant Cell Environ. 1997, 20, 350–358. [Google Scholar] [CrossRef]
- Olsen, R.L.; Pratt, R.B.; Gump, P.; Kemper, A.; Tallman, G. Red light activates a chloroplast-dependent ion uptake mechanism for stomatal opening under reduced CO2 concentrations in Vicia spp. New Phytol. 2002, 153, 497–508. [Google Scholar] [CrossRef]
- Saebo, A.; Krekling, T.; Appelgren, M. Light quality affects photosynthesis and leaf anatomy of birch plantlets in vitro. Plant Cell Tissue Organ Cult. 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Chen, X.L.; Wang, L.C.; Li, T.; Yang, Q.C.; Guo, W.Z. Sugar accumulation and growth of lettuce exposed to different lighting modes of red and blue LED light. Sci. Rep. 2019, 9, 10. [Google Scholar] [CrossRef]
- Li, H.M.; Tang, C.M.; Xu, Z.G. Effects of different light quality on growth, photosynthetic characteristic and chloroplast ultrastructure of upland cotton (Gossypium hirsutum L.) seedlings. Emir. J. Food Agric. 2017, 29, 104–113. [Google Scholar] [CrossRef]
- Li, C.; Liu, D.; Li, L.L.; Hu, S.X.; Xu, Z.G.; Tang, C.M. Effects of light-emitting diodes on the growth of peanut plants. Agron. J. 2018, 110, 2369–2377. [Google Scholar] [CrossRef]
- Miyachi, S.; Miyachi, S. Ammonia induces starch degradation in Chlorella cells. Plant Cell Physiol. 1985, 26, 245–252. [Google Scholar]
- Miyachi, S.; Miyachi, S. Some biochemical changes related to starch breakdown induced by blue light illumination and by addition of ammonia to chlorella cells. Plant Cell Physiol. 1987, 28, 309–314. [Google Scholar]
- Hammond, J.P.; White, P.J. Sucrose transport in the phloem: Integrating root responses to phosphorus starvation. J. Exp. Bot. 2008, 59, 93–109. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Source | PPFD 1 (μmol m−2 s−1) | Intensity (Lx) | Red (%) | Green (%) | Blue (%) |
|---|---|---|---|---|---|
| HPS | 73.61 2 | 2193 | 43.9 | 54.7 | 1.5 |
| R1BG5 3 | 69.18 | 2678 | 13.9 | 77 | 9.2 |
| R2BG3 4 | 77.12 | 2392 | 26.2 | 70.2 | 3.5 |
| R3BG1 5 | 73.99 | 2499 | 42.3 | 57.3 | 0.4 |
| Organs | Nutrient Concentration | Nutrient Content | ||
|---|---|---|---|---|
| F Value | Pr > F | F value | Pr > F | |
| Nitrogen | ||||
| Shoot | 77.01 | <0.0001 | 9.94 | 0.0003 |
| Root | 76.90 | <0.0001 | 17.49 | <0.0001 |
| Whole-plant | 39.71 | <0.0001 | 3.07 | 0.0512 |
| Phosphorus | ||||
| Shoot | 24.78 | <0.0001 | 10.90 | 0.0002 |
| Root | 51.44 | <0.0001 | 22.40 | <0.0001 |
| Whole-plant | 69.56 | <0.0001 | 10.04 | 0.0003 |
| Parameter | HPS 1 | R1BG5 2 | R2BG3 3 | R3BG1 4 | F Value | Pr > F |
|---|---|---|---|---|---|---|
| Chlorophyll a (mg g−1 FW) | 1.00 ± 0.13ab | 1.06 ± 0.20ab | 1.09 ± 0.04a | 0.84 ± 0.08b | 3.72 | 0.0283 |
| Chlorophyll b (mg g−1 FW) | 0.96 ± 0.19b | 1.25 ± 0.30ab | 1.43 ± 0.13a | 1.25 ± 0.13ab | 4.68 | 0.0124 |
| Chlorophyll a+b (mg g−1 FW) | 1.97 ± 0.24b | 2.31 ± 0.40ab | 2.52 ± 0.10a | 2.09 ± 0.17ab | 4.68 | 0.0124 |
| Soluble protein (mg g−1 FW) | 0.61 ± 0.11b | 0.67 ± 0.08ab | 0.91 ± 0.20a | 0.57 ± 0.15b | 5.55 | 0.0061 |
| GS5 activity (A540nm mg−1 protein h−1) | 4.31 ± 1.61b | 7.52 ± 1.62a | 5.20 ± 1.62ab | 4.79 ± 1.21b | 4.36 | 0.0162 |
| AP6 activity (μgNPP g−1 FW min−1) | 2.12 ± 0.57b | 3.24 ± 0.70a | 2.46 ± 0.59ab | 2.64 ± 0.40ab | 3.31 | 0.0409 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, H.; Hauer, R.J.; Chen, G.; Chen, X.; He, X. Growth, Nutrient Assimilation, and Carbohydrate Metabolism in Korean Pine (Pinus koraiensis) Seedlings in Response to Light Spectra. Forests 2020, 11, 44. https://doi.org/10.3390/f11010044
Wei H, Hauer RJ, Chen G, Chen X, He X. Growth, Nutrient Assimilation, and Carbohydrate Metabolism in Korean Pine (Pinus koraiensis) Seedlings in Response to Light Spectra. Forests. 2020; 11(1):44. https://doi.org/10.3390/f11010044
Chicago/Turabian StyleWei, Hongxu, Richard J. Hauer, Guoshuang Chen, Xin Chen, and Xingyuan He. 2020. "Growth, Nutrient Assimilation, and Carbohydrate Metabolism in Korean Pine (Pinus koraiensis) Seedlings in Response to Light Spectra" Forests 11, no. 1: 44. https://doi.org/10.3390/f11010044
APA StyleWei, H., Hauer, R. J., Chen, G., Chen, X., & He, X. (2020). Growth, Nutrient Assimilation, and Carbohydrate Metabolism in Korean Pine (Pinus koraiensis) Seedlings in Response to Light Spectra. Forests, 11(1), 44. https://doi.org/10.3390/f11010044