Simple Is Best: Pine Twigs Are Better Than Artificial Lures for Trapping of Pine Weevils in Pitfall Traps



Abstract
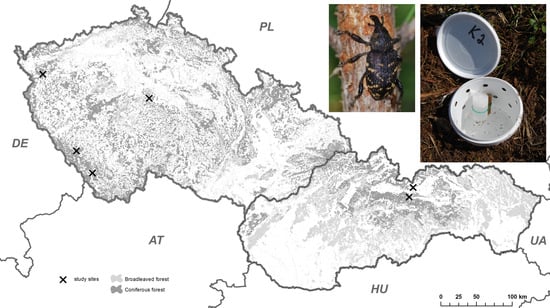
:
1. Introduction
2. Materials and Methods
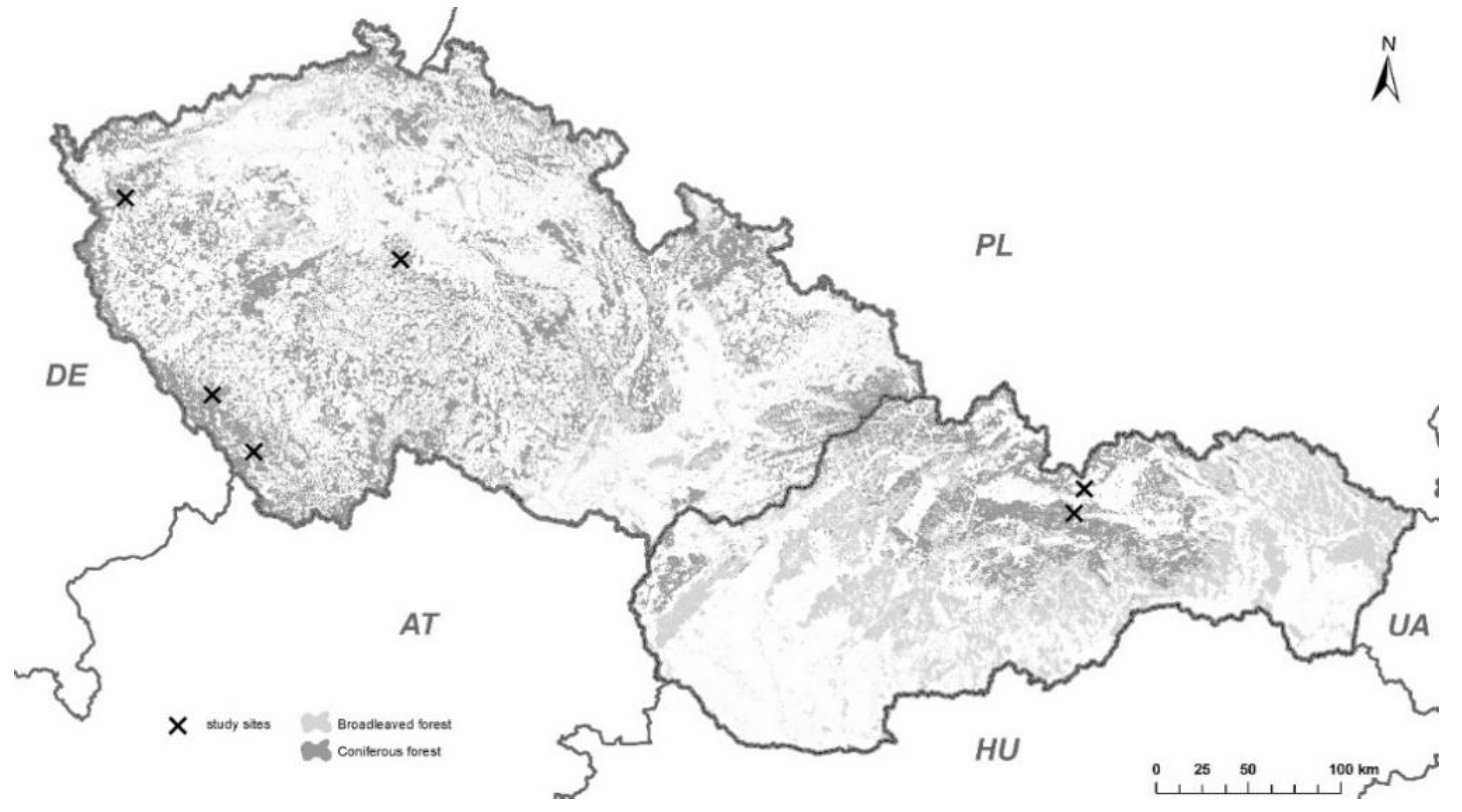
2.1. Localities
2.2. Attractants and Evaporators
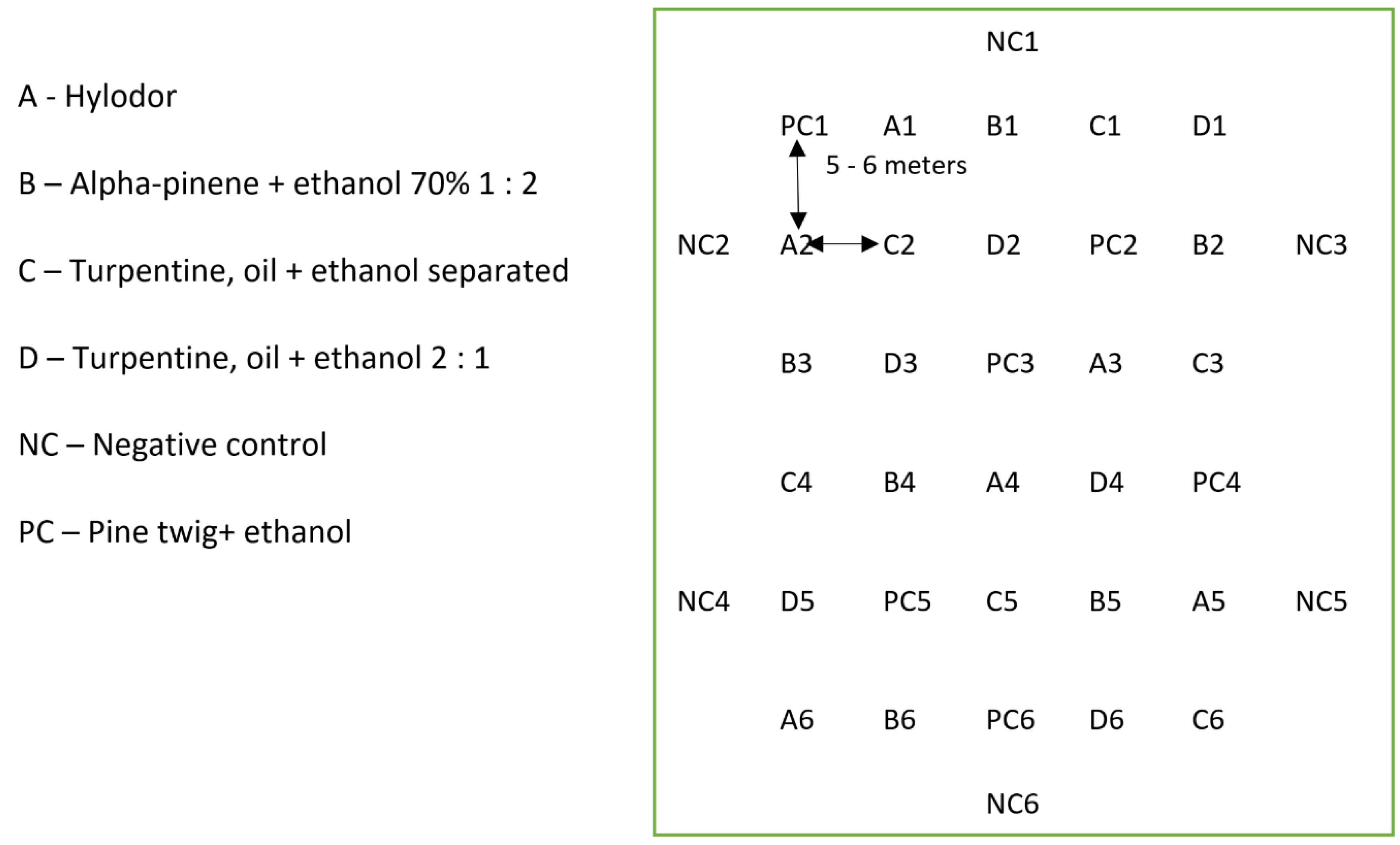
2.3. Pitfall Traps and Experimental Design
2.4. Statistical Analyses
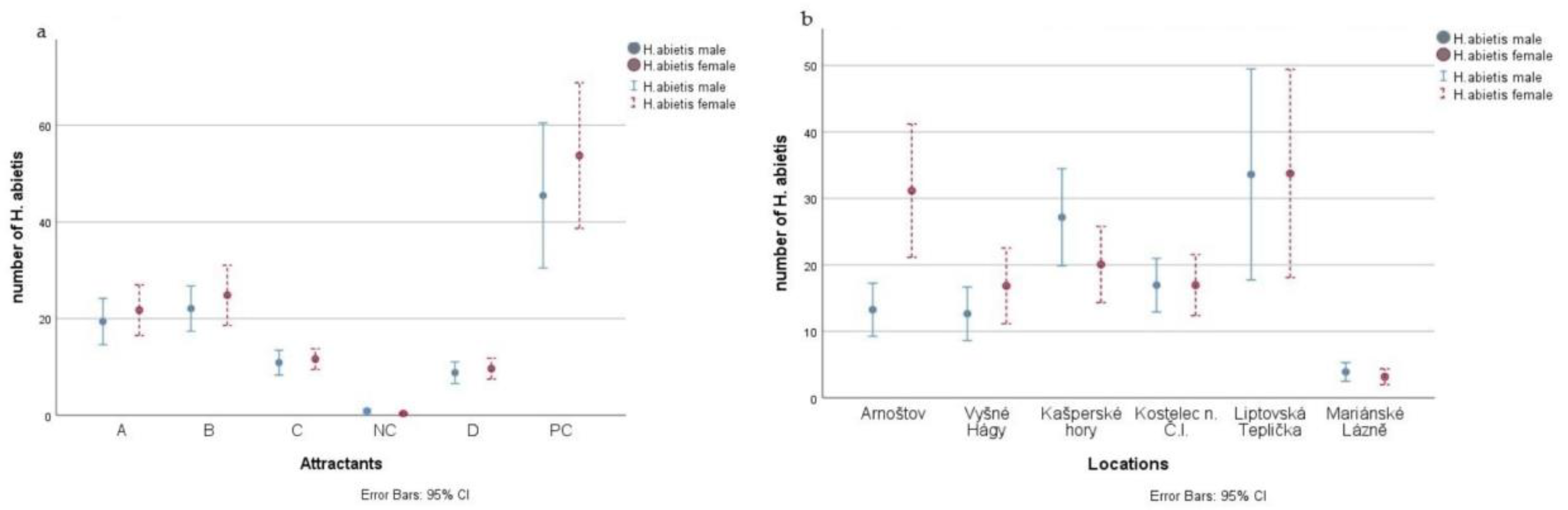
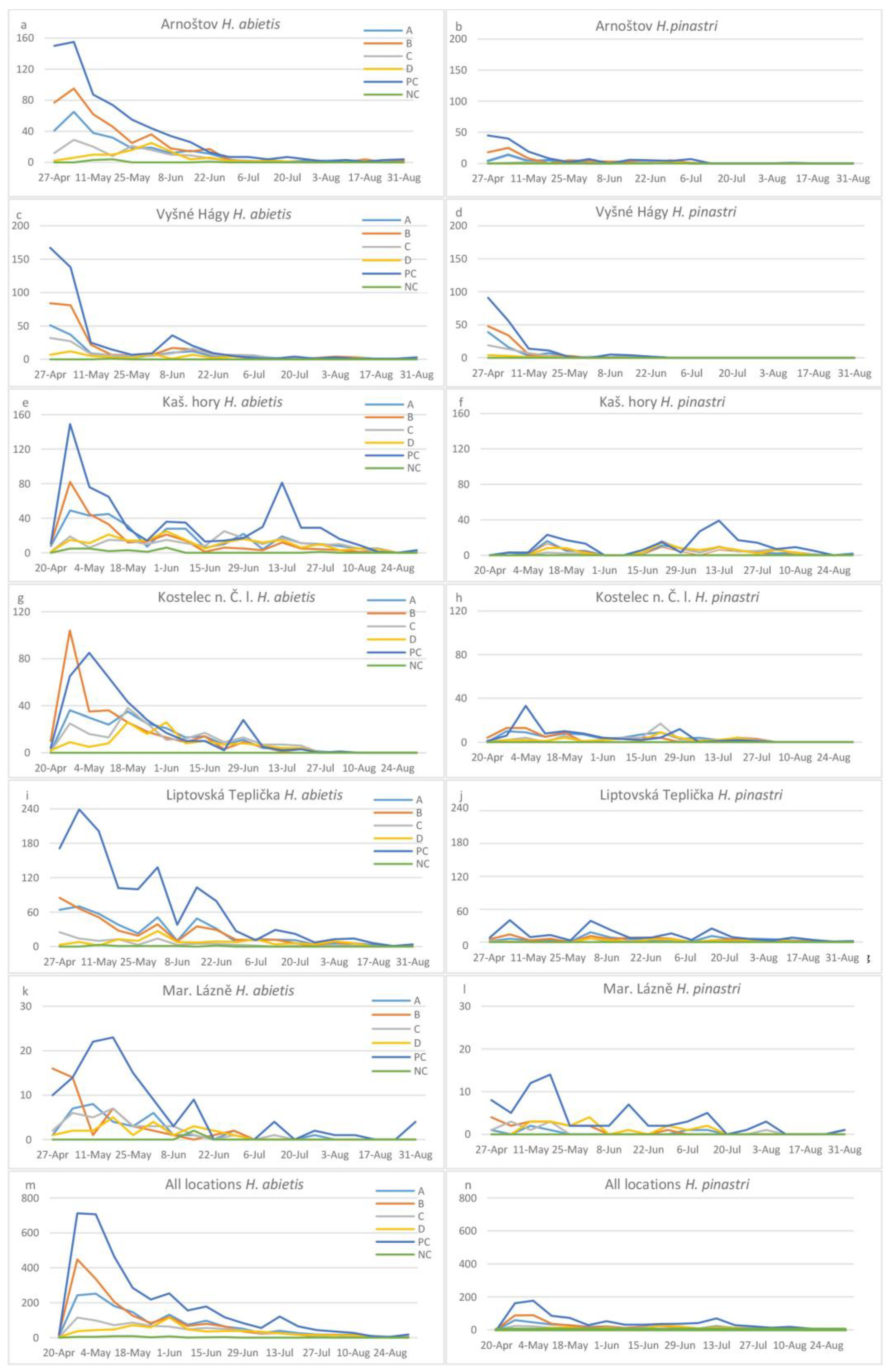
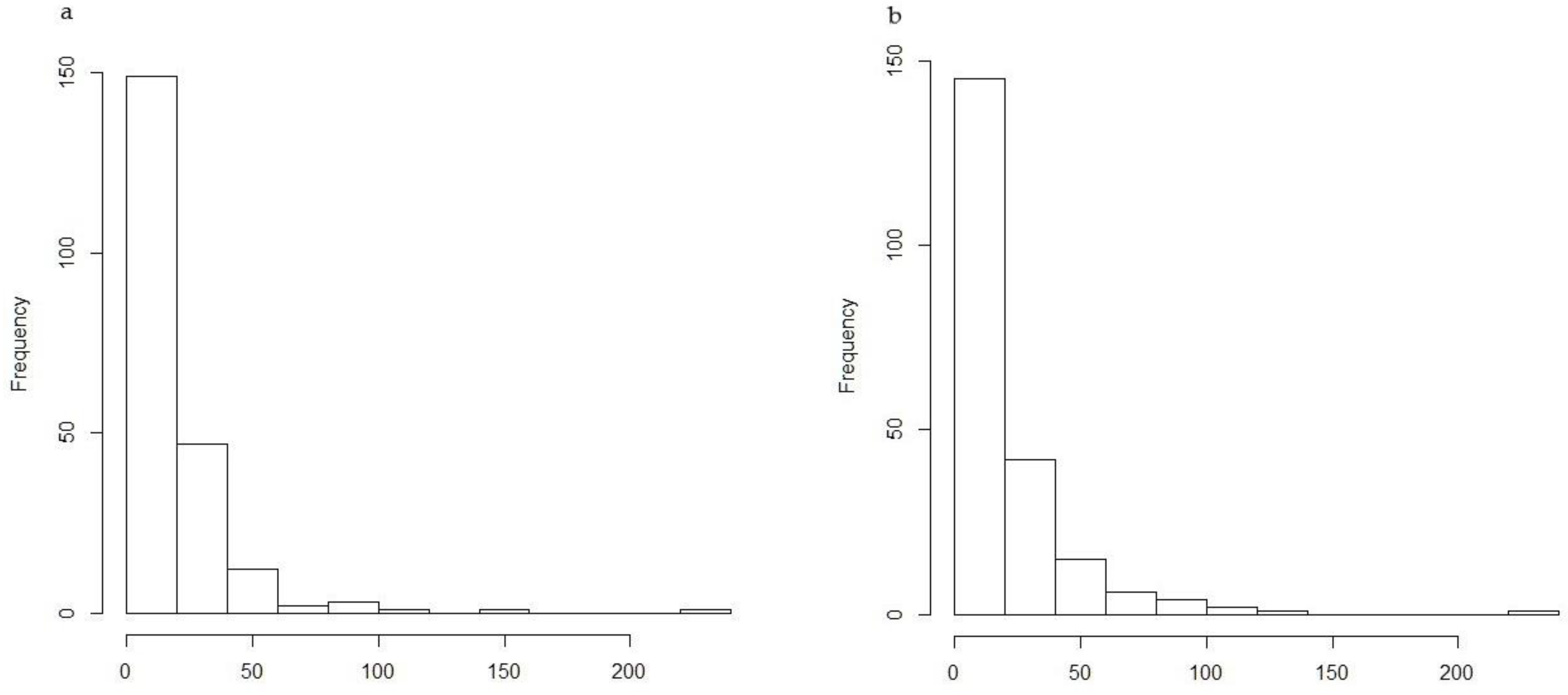
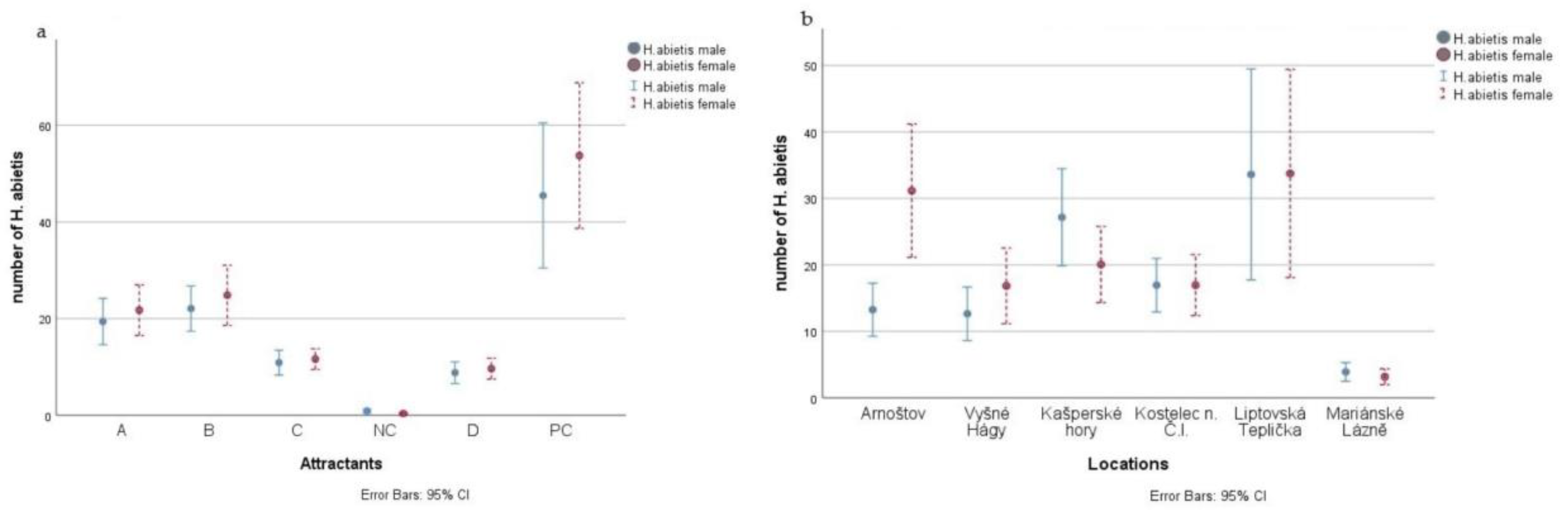
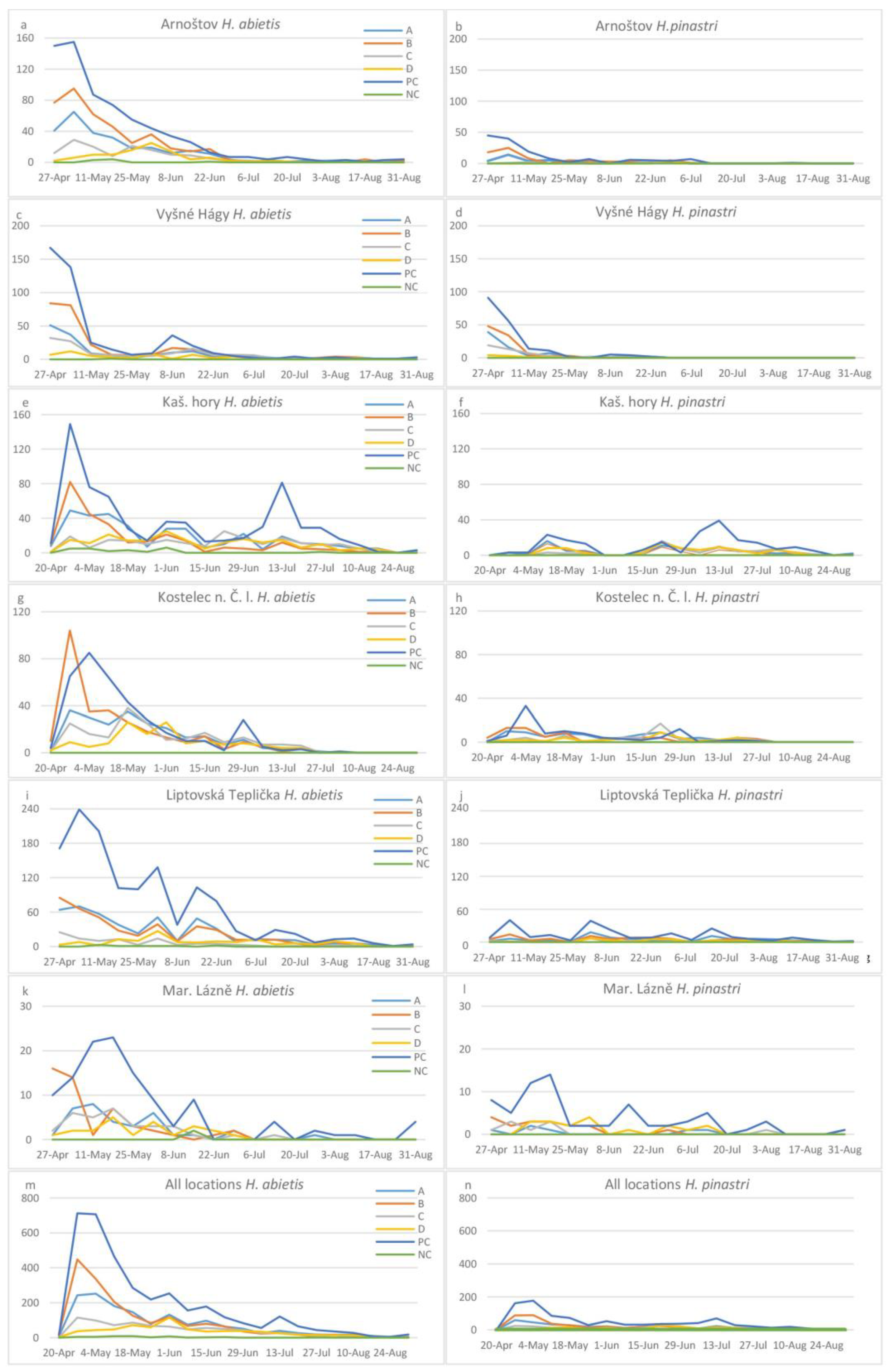
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
| Min | 1Q | Median | 3Q | Max |
| −10.2299 | −1.1360 | −0.1412 | 0.8858 | 10.0012 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimate | Std. | Error | z value Pr(>|z|) | |
|---|---|---|---|---|
| (Intercept) | 2.539 × 100 | 1.147 × 10−1 | 2.2134 × 101 | < 2 × 10−16 |
| atraktantB | 4.400 × 10−1 | 1.471 × 10−1 | 2.991 × 100 | 2.778 × 10−3 |
| atraktantC | −4.595 × 10−1 | 1.844 × 10−1 | −2.492 × 100 | 1.2685 × 10−2 |
| atraktantCon. | −2.539 × 100 | 4.241 × 10−1 | −5.987 × 100 | 2.13 × 10−9 |
| atraktantD | −7.198 × 10−1 | 2.005 × 10−1 | −3.591 × 100 | 3.30 × 10−4 |
| atraktantPC | 9.268 × 10−1 | 1.355 × 10−1 | 6.838 × 100 | 8.00 × 10−12 |
| lokalitaVyšné Hágy | −2.532 × 10−1 | 1.735 × 10−1 | −1.459 × 100 | 1.44503 × 10−1 |
| lokalitaKašperské hory | 9.775 × 10−1 | 1.346 × 10−1 | 7.264 × 100 | 3.75 × 10−13 |
| lokalitaKostelec n.C.l. | 5.213 × 10−1 | 1.448 × 10−1 | 3.600 × 100 | 3.18 × 10−4 |
| lokalitaLiptovská Teplička | 1.035 × 100 | 1.335 × 10−1 | 7.753 × 100 | 8.99 × 10−15 |
| lokalitaMarianske lázne | −1.386 × 100 | 2.565 × 10−1 | −5.405 × 100 | 6.49 × 10−8 |
| atraktantB:lokalitaVyšné Hágy | 2.616 × 10−1 | 2.168 × 10−1 | 1.207 × 100 | 2.27423 × 10−1 |
| atraktantC:lokalitaVyšné Hágy | 5.409 × 10−1 | 2.580 × 10−1 | 2.096 × 100 | 3.6046 × 10−2 |
| atraktantCon.:lokalitaVyšné Hágy | −1.539 × 100 | 1.094 × 100 | −1.406 × 100 | 1.59605 × 10−1 |
| atraktantD:lokalitaVyšné Hágy | −6.189 × 10−2 | 3.069 × 10−1 | −2.02 × 10−1 | 8.40180 × 10−1 |
| atraktantPC:lokalitaVyšné Hágy | 2.161 × 10−1 | 2.018 × 10−1 | 1.071 × 100 | 2.84314 × 10−1 |
| atraktantB:lokalitaKašperské hory | −5.949 × 10−1 | 1.799 × 10−1 | −3.307 × 100 | 9.43 × 10−4 |
| atraktantC:lokalitaKašperské hory | −9.515 × 10−2 | 2.181 × 10−1 | −4.36 × 10−1 | 6.62641 × 10−1 |
| atraktantCon.:lokalitaKašperské hory | −1.302 × 10−1 | 5.062 × 10−1 | −2.57 × 10−1 | 7.96947 × 10−1 |
| atraktantD:lokalitaKašperské hory | 1.300 × 10−1 | 2.325 × 10−1 | 5.59 × 10−1 | 5.75957 × 10−1 |
| atraktantPC:lokalitaKašperské hory | −3.462 × 10−1 | 1.615 × 10−1 | −2.143 × 100 | 3.2100 × 10−2 |
| atraktantB:lokalitaKostelec n.C.l. | −3.867 × 10−1 | 1.920 × 10−1 | −2.014 × 100 | 4.3966 × 10−2 |
| atraktantC:lokalitaKostelec n.C.l. | 2.709 × 10−1 | 2.264 × 10−1 | 1.197 × 100 | 2.31322 × 10−1 |
| atraktantCon.:lokalitaKostelec n.C.l. | −1.582 × 101 | 5.208 × 102 | −3.0 × 10−2 | 9.75762 × 10−1 |
| atraktantD:lokalitaKostelec n.C.l. | 2.667 × 10−2 | 2.522 × 10−1 | 1.06 × 10−1 | 9.15798 × 10−1 |
| atraktantPC:lokalitaKostelec n.C.l. | −6.026 × 10−1 | 1.784 × 10−1 | −3.378 × 100 | 7.30 × 10−4 |
| atraktantB:lokalitaLiptovská Teplička | −3.988 × 10−1 | 1.755 × 10−1 | −2.273 × 100 | 2.3056 × 10−2 |
| atraktantC:lokalitaLiptovská Teplička | −1.078 × 100 | 2.458 × 10−1 | −4.385 × 100 | 1.16 × 10−5 |
| atraktantCon.:lokalitaLiptovská Teplička | −7.476 × 10−1 | 5.563 × 10−1 | −1.344 × 100 | 1.79030 × 10−1 |
| atraktantD:lokalitaLiptovská Teplička | −5.353 × 10−1 | 2.475 × 10−1 | −2.163 × 100 | 3.0552 × 10−2 |
| atraktantPC:lokalitaLiptovská Teplička | 1.965 × 10−1 | 1.567 × 10−1 | 1.254 × 100 | 2.09960 × 10−1 |
| atraktantB:lokalitaMarianske lázne | 8.855 × 10−2 | 3.336 × 10−1 | −2.65 × 10−1 | 7.90676 × 10−1 |
| atraktantC:lokalitaMarianske lázne | −9.352 × 10−15 | 4.123 × 10−1 | 0 × 100 | 1.000000 × 100 |
| atraktantCon.:lokalitaMarianske lázne | 2.877 × 10−1 | 8.558 × 10−1 | 3.36 × 10−1 | 7.36764 × 10−1 |
| atraktantD:lokalitaMarianske lázne | 5.480 × 10−1 | 3.941 × 10−1 | 1.390 × 100 | 1.64403 × 10−1 |
| atraktantPC:lokalitaMarianske lázne | 2.877 × 10−1 | 2.943 × 10−1 | 9.77 × 10−1 | 3.28344 × 10−1 |
| (Intercept) | *** |
|---|---|
| atraktantB | ** |
| atraktantC | * |
| atraktantCon. | *** |
| atraktantD | *** |
| atraktantPC | *** |
| lokalitaVyšné Hágy | - |
| lokalitaKašperské hory | *** |
| lokalitaKostelec n. C.l. | *** |
| lokalitaLiptovská Teplička | *** |
| lokalitaMarianske lázne | *** |
| atraktantB:lokalitaVyšné Hágy | - |
| atraktantC:lokalitaVyšné Hágy | * |
| atraktantCon.:lokalitaVyšné Hágy | - |
| atraktantD:lokalitaVyšné Hágy | - |
| atraktantPC:lokalitaVyšné Hágy | - |
| atraktantB:lokalitaKašperské hory | *** |
| atraktantC:lokalitaKašperské hory | - |
| atraktantCon.:lokalitaKašperské hory | - |
| atraktantD:lokalitaKašperské hory | - |
| atraktantPC:lokalitaKašperské hory | * |
| atraktantB:lokalitaKostelec n. C.l. | * |
| atraktantC:lokalitaKostelec n. C.l. | - |
| atraktantCon.:lokalitaKostelec n. C.l. | - |
| atraktantD:lokalitaKostelec n. C.l. | - |
| atraktantPC:lokalitaKostelec n. C.l. | *** |
| atraktantB:lokalitaLiptovská Teplička | * |
| atraktantC:lokalitaLiptovská Teplička | *** |
| atraktantCon.:lokalitaLiptovská Teplička | - |
| atraktantD:lokalitaLiptovská Teplička | * |
| atraktantPC:lokalitaLiptovská Teplička | - |
| atraktantB:lokalitaMarianske lázne | - |
| atraktantC:lokalitaMarianske lázne | - |
| atraktantCon.:lokalitaMarianske lázne | - |
| atraktantD:lokalitaMarianske lázne | - |
| atraktantPC:lokalitaMarianske lázne | - |
References
- Escherich, K. Die Forstinsekten Mitteleuropas, 2nd ed.; Paul Parey: Berlin, Germany, 1923; p. 663. [Google Scholar]
- Day, K.R.; Leather, S.R. Threats to forestry by insect pests in Europe. In Forests and Insects; Watt, A.D., Stork, N.E., Hunter, M.D., Eds.; Chapman & Hall: London, UK, 1997; pp. 177–205. [Google Scholar]
- Långstöm, B.; Day, K.R. Damage, control and management of weevil pests, especially Hylobius abietis. In Bark and Wood Boring Insects in Living Trees in Europe: A Synthesis; Lieutier, F., Day, K.R., Battisti, A., Grégoire, J.-C., Evans, H.F., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 415–444. [Google Scholar]
- Eidmann, H.H. Hylobius Schönh. In Die Forstschädlinge Europas, 2nd ed.; Schwenke, W.K., Ed.; Paul Parey: Hamburg/Berlin, Germany, 1974; pp. 275–293. [Google Scholar]
- Lindström, A.; Hellqvist, C.; Gyldberg, B.; Långström, B.; Mattsson, A. Field performance of a protective collar against damage by Hylobius abietis. Scand. J. For. Res. 1986, 1, 3–15. [Google Scholar] [CrossRef]
- Eidmann, H.H.; von Sydow, F. Stockings for protection of containerised seedlings against pine weevil (Hylobius abietis L.) damage. Scand. J. For. Res. 1989, 4, 537–547. [Google Scholar] [CrossRef]
- Hagner, M.; Jonsson, C. Survival after planting without soil preparation for pine and spruce seedlings protected from Hylobius abietis by physical and chemical shelters. Scand. J. For. Res. 1995, 10, 225–234. [Google Scholar] [CrossRef]
- Eidmann, H.H.; Nordenhem, H.; Weslien, J. Physical protection of conifer seedlings against pine weevil feeding. Scand. J. For. Res. 1996, 11, 68–75. [Google Scholar] [CrossRef]
- Zumr, V.; Stary, P. LATEX paint as an antifeedent against Hylobius abietis (L.) (Col., Curculionidae) on conifer seedlings. Anz. Schädl. Pflanzenschutz Umweltschutz 1995, 21, 42–43. [Google Scholar] [CrossRef]
- Nordlander, G.; Nordenhem, H.; Hellqvist, C. A flexible sand coating (Conniflex) for the protection of conifer seedlings against damage by the pine weevil Hylobius abietis. Agric. For. Entomol. 2009, 11, 91–100. [Google Scholar] [CrossRef]
- Kvaae.no. Available online: http://kvaae.no/what/ (accessed on 15 May 2019).
- Harte, J.; Holdren, C.; Schneider, R.; Shirley, C. Toxics A to Z, a Guide to Everyday Pollution Hazards; University of California: Oxford, UK, 1991; p. 478. ISBN 9780520072244. [Google Scholar]
- Rattner, B.A. History of wildlife toxicology. Ecotoxicology 2007, 18, 773–783. [Google Scholar] [CrossRef]
- Bradberry, S.M.; Cage, S.A.; Proudfoot, A.T.; Vale, J.A. Poisoning due to pyrethroids. Toxicol. Rev. 2005, 24, 93–106. [Google Scholar] [CrossRef]
- Pszczolkowski, M.A.; Dobrowolski, M. Circadian dynamics of locomotor activity and deltamethrin susceptibility in the pine Weevil Hylobius abietis. Phytoparasitica 1999, 27, 19–25. [Google Scholar] [CrossRef]
- Dobrowolski, M. The susceptibility of the large pine weevil (Hylobius abietis L.) to insecticides and the role of the oxidative metabolism in the developing of the pest resistance to DDT and pyrethroids. Folia For. Polon. 2000, 42, 83–94. [Google Scholar]
- Lempérière, G.; Julien, J. Protection against the pine weevil-Efficiency of a carbosulfan-based systemic insecticide. Rev. For. Fr. 2003, 55, 129–140. [Google Scholar] [CrossRef]
- Rose, D.; Matthews, G.A.; Leather, S.R. Sub-lethal responses of the large pine weevil, Hylobius abietis, to the pyrethroid insecticide lambda-cyhalothrin. Physiol. Entomol. 2006, 31, 316–327. [Google Scholar] [CrossRef]
- Glovacka, B.; Lech, A.; Wilczynski, W. Application of deltamethrin for spraying or dipping to protect Scots pine seedlings againts Hylobius abietis L. and logs against Tomicus piniperda L. Ann. Sci. For. 1991, 48, 113–117. [Google Scholar] [CrossRef]
- Torstensson, L.; Börjesson, E.; Arvidsson, B. Treatment of bare root spruce seedlings with permethrin against pine weevil before lifting. Scand. J. For. Res. 1999, 14, 408–415. [Google Scholar] [CrossRef]
- Rose, D.; Matthews, G.A.; Leather, S.R. Recognition and avoidance of insecticide-treated Scots Pine (Pinus sylvestris) by Hylobius abietis (Coleoptera: Curculionidae): Implications for pest management strategies. Agric. For. Entomol. 2005, 7, 187–191. [Google Scholar] [CrossRef]
- Ratzeburg, J.T.C. Die Forst- Insekten. Erster Teil–Die Käfer; 2 Auflage; Nicolai’sche Buchhandlung: Berlin, Germany, 1839; p. 247. [Google Scholar]
- Pfeffer, A.; Horák, E.; Kudela, M.; Muller, J.; Novakova, E.; Stolina, M. Ochrana Lesů; Státní Zemědělské Nakladatelství: Praha, Czech Republic, 1961; p. 838. [Google Scholar]
- Modlinger, R.; Knížek, M. Klikoroh borový Hylobius abietis (L.). Lesnická Pr. 2009, 88, 1–4. [Google Scholar]
- Galko, J.; Gubka, A.; Vakula, J. Praktické Skúsenosti s Využitím Lapacích Kôr na Zníženie Škôd Spôsobených Tvrdoňom Smrekovým na Mladých Výsadbách Ihličnatých Drevín. In Aktuálne Problémy v Ochrane Lesa; Kunca, A., Nový Smokovec, S., Kunca, A., Eds.; Národné Lesnícke Centrum: Zvolen, Slovakia, 2012; pp. 60–64. [Google Scholar]
- Galko, J.; Vakula, J.; Kunca, A.; Rell, S.; Gubka, A. Ochrana Lesa; Ochrana lesa proti tvrdoňom a lykokazom na sadeniciach; Úrad pre Normalizáciu, Metorologiu a Skúšobníctvo: Bratislava, Slovakia, STN 48 2714; 2016; p. 8. [Google Scholar]
- Nordenhem, H. Age, sexual development, and seasonal occurrence of the pine weevil Hylobius abietis. J. Appl. Entomol. 1989, 108, 260–270. [Google Scholar] [CrossRef]
- Skłodowski, J.J.W.; Gadziński, J. Effectiveness of beetle catches in two types of traps for Hylobius abietis L. Sylwan 2001, 6, 55–63. [Google Scholar]
- Moreira, X.; Costas, R.; Sampedro, L.; Zas, R. A simple method for trapping Hylobius abietis (L.) alive in northern Spain. For. Syst. 2008, 17, 188–192. [Google Scholar] [CrossRef]
- Kuźmiński, R.; Bilon, A. Evaluation of effectiveness of selected types of traps used in capturing of large pine weevil–Hylobius abietis (L.). Acta Sci. Pol.—Silv. Colendarum R. Ind. Lig. 2009, 8, 19–26. [Google Scholar]
- Zumr, V.; Stary, P.P. Field experiments with different attractants in baited pitfall traps for Hylobius abietis L. (Col., Curculionidae). J. Appl. Entomol. 1992, 113, 451–455. [Google Scholar] [CrossRef]
- Tilles, D.A.; Sjödin, K.; Nordlander, G.; Eidmen, H.H. Synergism between ethanol and conifer host volatiles as attractants for the pine weevil, Hylobius abietis L. (Coleoptera: Curculionidae). J. Econ. Entomol. 1986, 79, 970–973. [Google Scholar] [CrossRef]
- Nordlander, G. A method for trapping hylobius abietis (L.) with a standardized bait and its potential for forecasting seedling damage. Scand. J. For. Res. 1987, 2, 199–213. [Google Scholar] [CrossRef]
- Erbilgin, N.; Szele, A.; Klepzig, K.D.; Raffa, K.F. Trap Type, Chirality of α-Pinene, and Geographic Region Affect Sampling Efficiency of Root and Lower Stem Insects in Pine. J. Econ. Entomol. 2001, 94, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Nordlander, G. Limonene inhibist attraction to Alpha-pinene in the pine weevils Hylobius abietis and H. pinastri. J. Chem. Ecol. 1990, 16, 1307–1320. [Google Scholar] [CrossRef] [PubMed]
- Olenici, N.; Duduman, M.L.; Teodosiu, M.; Olenici, V. Efficacy of artificial traps to prevent the damage of conifer seedlings by large pine weevil (Hylobius abietis L.)—A preliminary study. Bull. Transilv. Univ. Bras. 2016, 9, 9–20. [Google Scholar]
- Stocki, J.S. The Use of Pheromones and Pheromone Traps in Forest Protection in Poland in the Years 1980–1997; Practice oriented results on the use and production of Neem ingredients and pheromones VIII; Kleeberg, H., Zebitz, C.P.W., Eds.; Druck & Graphics: Giessen, Germany, 2000; pp. 128–133. [Google Scholar]
- Skrzecz, I. Non-target insects in the pine weevil (Hylobius abietis L.) traps with Hylodor dispenser. Folia For. Polon. Ser. A For. 2003, 35, 27–35. [Google Scholar]
- Voolma, K.; Siida, I.; Sibul, I. Forest insects attracted to ground traps baited with turpentineand ethanol on clear-cuttings. Nor. J. Entomol. 2001, 48, 103–110. [Google Scholar]
- Wheater, C.P.; Bell, J.R.; Cook, P.A. Practical Field Ecology: A Project Guide; Wiley-Blackwell: Chichester, UK, 2011; p. 388. [Google Scholar]
- Örlander, G.; Nilsson, U.; Nordlander, G. Pine weevil abundance on clear-cuttings of different ages: A 6-year study using pitfall traps. Scand. J. For. Res. 1997, 12, 225–240. [Google Scholar] [CrossRef]
- Saalas, U. Suomen Metsähyönteiset; Finnish Forest Insects; Wsoy: Porvoo, Finland, 1949; p. 719. [Google Scholar]
- Ozols, G. Prides un Egles Dedrofagien Kukaini Latvijas Mezos; (Dendrophagus insects of pine and spruce in Latvian forests); Zinātne: Riga, Latvia, 1985; p. 208. [Google Scholar]
- Ehnström, B.; Axelsson, R. Insekts Gnag I Bark Och Ved; SLU: Uppsala, Sweden, 2002. [Google Scholar]
- Längström, B. Abundance and seasonal activity of adult Hylobius-weevils in reforestation areas during the first years following final felling. Commun. Lnst. For. Fenn. 1982, 106, 1–23. [Google Scholar]
- Maavara, V.; Merihein, A.; Parmas, H.; Parmasto, E. Metsakaitse; Valgus: Tallinn, Estonia, 1961; p. 733. [Google Scholar]
- Luik, A.; Voolma, K. Some aspects of the occurrence, biology and cold-hardiness of Hylobius weevils. In Insects Affecting Reforestation: Biology and Damage; Alfaro, R.I., Glover, S.G., Eds.; Pacific and Yukon Region, Forestry Canada: Victoria, BC, Canada, 1989; pp. 28–33. [Google Scholar]
- Viiri, H.; Miettinen, O. Feeding preferences of Hylobius pinastri Gyll. Balt. For. 2013, 19, 161–164. [Google Scholar]
- Leather, S.R.; Ahmed, S.J.; Hogan, L. Adult feeding preferences of the pine weevil, Hylobius abietis (Coleoptera: Curculionidae). Eur. J. Entomol. 1994, 91, 385–389. [Google Scholar]
- Varkonda, Š.; Florian, Š. Odparník na regulované uvoľnovanie prchavých látok; (Evaporator for controlled release of volatile substances) Utility model, SK 4570 U; Úrad Priemyselného Vlastníctva: Bratislava, Slovakia, 2006; p. 3. [Google Scholar]
- Galko, J.; Kunca, A.; Ondruš, M.; Špilda, I.; Rell, S. Zhodnotenie a porovnanie nákladov na rôznu formu ošetrenia ihličnatých sadeníc proti tvrdoňovi smrekovému. In Aktuálne Problémy v Ochrane Lesa; Kunca, A., Nový Smokovec, S., Kunca, A., Eds.; Národné Lesnícke Centrum: Zvolen, Slovakia, 2015; pp. 101–105. [Google Scholar]
- Thorin, J.; Nömmik, H. Monoterpene composition of cortical oleoresin from different clones of Pinus sylvestris. Phytochemistry 1974, 13, 1879–1881. [Google Scholar] [CrossRef]
- Yazdani, R.; Nilsson, J.E. Cortical monoterpene variation in natural populations of Pinus sylvestris in Sweden. Scand. J. For. Res. 1986, 1, 85–93. [Google Scholar] [CrossRef]
- Nordlander, G. Host finding in the pine weevil Hylobius abietis: Effects of conifer volatiles and added limonene. Entomol. Exp. Appl. 1991, 59, 229–237. [Google Scholar] [CrossRef]
- Squillace, A.E. Analyses of monoterpenes of conifers by gas-liquid chromatography. In Modem Methods in Forest Genetics; Miksche, J.P., Ed.; Springer: New York, NY, USA, 1976; pp. 139–157. [Google Scholar]
- Voolma, K.; Sibul, I. The effect of collecting fluid on the catch results of Hylobius abietis in pitfall traps. Agron. Res. 2006, 4, 457–460. [Google Scholar]
- Saintonge, F.X.; Malphettes, C.B. Un piege pour surveiller les populations d’hylobes (Hylobius abietis L.) (Coleop.: Curc.)? Etudes de Cemagref. Sér. For. 1991, 6, 138–155. [Google Scholar]
- Charitonova, N.Z. Bolšoj Sosnovyj Dolgonosik I Borba s Nim; 1. Vyd; Lesnaja Promyšlennost Progress: Moskva, Russia, 1965; p. 88. ISBN 934-0-41-595-768-24. [Google Scholar]
- Nordlander, G.; Örlander, G.; Langvall, O. Feeding by the pine weevil Hylobius abietis in relation to sun. exposure and distance to forest edges. Agric. For. Entomol. 2003, 5, 191–198. [Google Scholar] [CrossRef]
- Bejer-Petersen, B.; Juutinen, P.; Kangas, E.; Bakke, A.; Butovitsch, V.; Eidmann, H.; Heqvist, K.J.; Lekander, B. Studies on Hylobius abietis L. I. Development and life cycles in the Nordic countries. Acta Entomol. Fenn. 1962, 17, 1–107. [Google Scholar]
- Selander, J.; Kalo, P.; Kangas, E.; Pertunnen, V. Olfactory behavior of Hylobius abietis L. (Col., Curculionidae). I. Response to several terpenoid fractions isolated from Scots pine phloem. Ann. Entomol. Fenn. 1974, 39, 40–45. [Google Scholar]
- Nordenham, H.; Eidmann, H.H. Response of the pine weevil Hylobius abietis L. (Col., Curculionidae) to host volatiles in different phases of its adult life cycle. J. Appl. Entomol. 1991, 112, 353–358. [Google Scholar] [CrossRef]
- Björklund, N.; Nordlander, G.; Bylund, H. Olfactory and visual stimuli used in orientation to conifer seedlings by the pine weevil, Hylobius abietis. Physiol. Entomol. 2005, 30, 225–231. [Google Scholar] [CrossRef]
- Tilles, D.A.; Eidmann, H.H.; Solbreck, B. Mating stimulant of the pine weevil Hylobius abietis (L.). J. Chem. Ecol. 1988, 14, 1495–1503. [Google Scholar] [CrossRef] [PubMed]
- Selander, J. Evidence of pheromone-mediated behaviour in the large pine weevil, Hylobius abietis (Coleoptera, Curculionidae). Ann. Ent. Fenn. 1978, 44, 105–112. [Google Scholar]
- Mráček, Z.; Šrůtka, P. Stav znalostí a možnostech obrany proti klikorhu borovému (Hylobius abietis L.) v zemích Evropy. Zprávy Lesnického Výzk. 1984, 29, 21–25. [Google Scholar]
- Solbreck, C.; Gyldberg, B. Temporal flight pattern of the large pine weevil, Hylobius abietis L. (Coleoptera, Curculionidae), with special reference to the influence of weather. Z. Angew. Entomol. 1979, 88, 532–536. [Google Scholar] [CrossRef]

| Country/Locality | Geographical Coordinates | Area (ha) | Altitude | Aspect | Establishment of Cleared Area | Spruce Share (%) |
|---|---|---|---|---|---|---|
| CZ/Arnoštov | 48.8890528N 13.9529108E | 0.74 | 944–946 | Southeast | Felling September 2017 | 95 |
| SK/Vyšné Hágy | 49.120814N 20.102610E | 21.5 | 1216–1220 | South | Wind-throw May 2014 | 95 |
| CZ/Kašperské Hory | 49.1343394N 13.5876222E | 0.81 | 780–785 | Southwest | Felling October 2017 | 100 |
| CZ/Kostelec n. Č. L | 49.9198067N 14.840943E | 0.62 | 402–404 | South | Felling December 2017 | 90 |
| SK/Liptovská Teplička | 48.995592N 20.038704E | 10.4 | 1115–1127 | North | Bark-beetle disturbance February 2018 | 100 |
| CZ/Mariánske Lázně | 50.0246936N 12.7173419E | 0.45 | 819–820 | Plane | Bark-beetle disturbance May 2017 | 100 |
| Attractant | Daily Vapour (g) ** | Attractant Composition | Ratio of Components |
|---|---|---|---|
| A—Hylodor | 0.012 | mixture of organic and inorganic substances | - |
| B—alpha-pinene + ethanol * | 0.070 | mixture of alpha-pinene (1R)-(+)-alpha-pinene (98%) CAS7785-70-8, EINECS: 232087-8 + ethanol CAS: 64-17-5, EINECS: 200-578-6 | 1:2 *** |
| C—Turpentine oil + ethanol separated * | 1st tube: 0.023 2nd tube: 0.241 | 1st tube: ethanol CAS: 64-17-5, EINECS: 200-578-6 2nd tube: turpentine, oil CAS: 7785-70-8, EINECS: 232087-8 | 100% 100% |
| D—Turpentine oil + ethanol * | 0.093 | mixture: turpentine, oil CAS: 7785-70-8, EINECS: 232087-8 + ethanol CAS: 64-17-5, EINECS: 200-578-6 | 2:1 **** |
| PC—pine branch + ethanol in a bottle | 0.402 | ethanol CAS: 64-17-5, EINECS: 200-578-6 | - |
| - | Attractant | |||||
|---|---|---|---|---|---|---|
| Locality | Hylodor | Alpha-Pinene + Ethanol | Turpentine, Oil + Ethanol Separated | Turpentine, Oil + Ethanol Combined | Negative Control | Pine Twig + Ethanol |
| Arnoštov | 269/38 | 405/74 | 137/9 | 102/2 | 8/0 | 681/144 |
| Vyšné Hágy | 150/72 | 265/96 | 130/48 | 59/9 | 1/0 | 449/185 |
| Kašperské Hory | 34,379 | 273/54 | 205/63 | 205/76 | 23/0 | 656/202 |
| Kostelec n. Č.L | 228/72 | 286/65 | 200/51 | 139/37 | 0/0 | 367/95 |
| Liptovská Teplička | 456/88 | 416/80 | 103/18 | 137/35 | 9/0 | 1310/226 |
| Mariánské Lázně | 34/7 | 47/17 | 32/10 | 22/18 | 2/0 | 117/69 |
| Total | 1480/356 | 1692/387 | 807/199 | 664/177 | 43/0 | 3580/921 |
| Locality | Number of H. abietis Males Trapped | Number of H. abietis Females Trapped | Sex Ratio of Trapped Males:Females | Total Number of H. abietis Trapped | Number of H. abietis Trapped/Ha | Number of H. pinastri Trapped | Number of H. pinastri Trapped/Ha |
|---|---|---|---|---|---|---|---|
| Arnoštov | 480 | 1122 | 30:70 | 1602 | 16,584 | 267 | 2764 |
| Vyšné Hágy | 456 | 598 | 43:57 | 1054 | 10,910 | 411 | 4255 |
| Kašperské Hory | 979 | 726 | 57:43 | 1705 | 17,650 | 474 | 4907 |
| Kostelec n. Č.L. | 610 | 610 | 50:50 | 1220 | 12,630 | 320 | 3313 |
| Liptovská Teplička | 1212 | 1219 | 50:50 | 2431 | 25,165 | 447 | 4627 |
| Mariánské Lázně | 140 | 114 | 55:45 | 254 | 2630 | 121 | 1253 |
| Total or average | 3877 | 4389 | 47:53 * | 1378 | 14,261 * | 340 | 3520 * |
| Attractant Pair | Absolute Difference | Standard Error | z-Value | p-Value |
|---|---|---|---|---|
| B-A | 0.4400 | 0.1471 | 2.991 | <0.02791 * |
| C-A | −0.4595 | 0.1844 | −2.492 | 0.10974 |
| NC-A | −2.5390 | 0.4241 | −5.987 | <0.001 *** |
| D-A | −0.7198 | 0.2005 | −3.591 | <0.00368 ** |
| PC-A | 0.9268 | 0.1355 | 6.838 | <0.001 *** |
| C-B | −0.8995 | 0.1712 | −5.254 | <0.001 *** |
| NC-B | −2.9789 | 0.4185 | −7.118 | <0.001 *** |
| D-B | −1.1598 | 0.1884 | −6.155 | <0.001 *** |
| PC-B | 0.4868 | 0.1170 | 4.162 | <0.001 *** |
| NC-C | −2.0794 | 0.4330 | −4.802 | <0.001 *** |
| D-C | −0.2603 | 0.2188 | −1.190 | 0.82316 |
| PC-C | 1.3863 | 0.1614 | 8.591 | <0.001 *** |
| D-NC | 1.8192 | 0.1614 | 4.133 | <0.001 *** |
| PC-NC | 3.4657 | 0.4146 | 8.360 | <0.001 *** |
| PC-D | 1.6466 | 0.1795 | 9.171 | <0.001 *** |
| Attractant Pair | Absolute Difference | Standard Error | z-Value | p-Value |
|---|---|---|---|---|
| B-A | 0.39330 | 0.09315 | 4.222 | <0.001 *** |
| C-A | −0.77405 | 0.12813 | −6.041 | <0.001 *** |
| NC-A | −4.56954 | 0.71076 | −6.429 | <0.001 *** |
| D-A | −1.08830 | 0.14341 | −7.589 | <0.001 *** |
| PC-A | 0.92557 | 0.08506 | 10.882 | <0.001 *** |
| C-B | −1.16736 | 0.12138 | −9.618 | <0.001 *** |
| NC-B | −4.96284 | 0.70957 | −6.994 | <0.001 *** |
| D-B | −1.48160 | 0.13741 | −10.782 | <0.001 *** |
| PC-B | 0.53227 | 0.07450 | 7.145 | <0.001 *** |
| NC-C | −3.79549 | 0.71501 | −5.308 | <0.001 *** |
| D-C | −0.31425 | 0.16316 | −1.926 | 0.337 |
| PC-C | 1.69963 | 0.11528 | 14.744 | <0.001 *** |
| D-NC | 3.48124 | 0.71790 | 4.849 | <0.001 *** |
| PC-NC | 5.49512 | 0.70856 | 7.755 | <0.001 *** |
| PC-D | 2.01388 | 0.13205 | 15.251 | <0.001 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lalík, M.; Holuša, J.; Galko, J.; Resnerová, K.; Kunca, A.; Nikolov, C.; Mudrončeková, S.; Surový, P. Simple Is Best: Pine Twigs Are Better Than Artificial Lures for Trapping of Pine Weevils in Pitfall Traps. Forests 2019, 10, 642. https://doi.org/10.3390/f10080642
Lalík M, Holuša J, Galko J, Resnerová K, Kunca A, Nikolov C, Mudrončeková S, Surový P. Simple Is Best: Pine Twigs Are Better Than Artificial Lures for Trapping of Pine Weevils in Pitfall Traps. Forests. 2019; 10(8):642. https://doi.org/10.3390/f10080642
Chicago/Turabian StyleLalík, Michal, Jaroslav Holuša, Juraj Galko, Karolína Resnerová, Andrej Kunca, Christo Nikolov, Silvia Mudrončeková, and Peter Surový. 2019. "Simple Is Best: Pine Twigs Are Better Than Artificial Lures for Trapping of Pine Weevils in Pitfall Traps" Forests 10, no. 8: 642. https://doi.org/10.3390/f10080642
APA StyleLalík, M., Holuša, J., Galko, J., Resnerová, K., Kunca, A., Nikolov, C., Mudrončeková, S., & Surový, P. (2019). Simple Is Best: Pine Twigs Are Better Than Artificial Lures for Trapping of Pine Weevils in Pitfall Traps. Forests, 10(8), 642. https://doi.org/10.3390/f10080642