Effects of Five Growing Media and Two Fertilizer Levels on Polybag—Raised Camden Whitegum (Eucalyptus benthamii Maiden & Cambage) Seedling Morphology and Drought Hardiness



Abstract
:1. Introduction
2. Materials and Methods
2.1. Nursery Phase
2.1.1. Plant Material
2.1.2. Treatments
2.1.3. Fertilization and Irrigation
2.1.4. Experimental Design
2.1.5. Measurements
2.2. Drought Test Phase
2.2.1. Experimental Design
2.2.2. Measurements
2.3. Statistical Analysis
3. Results
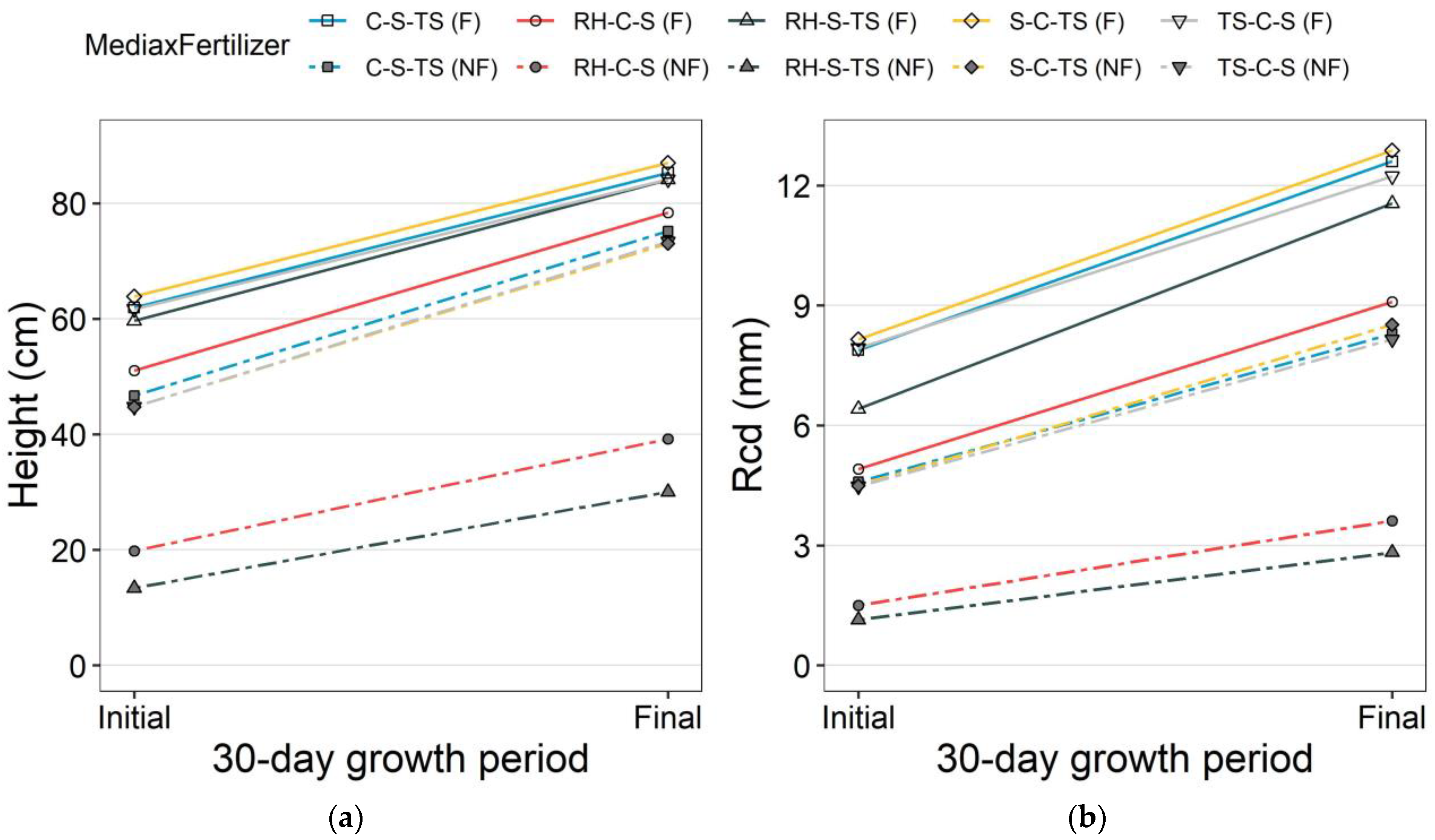
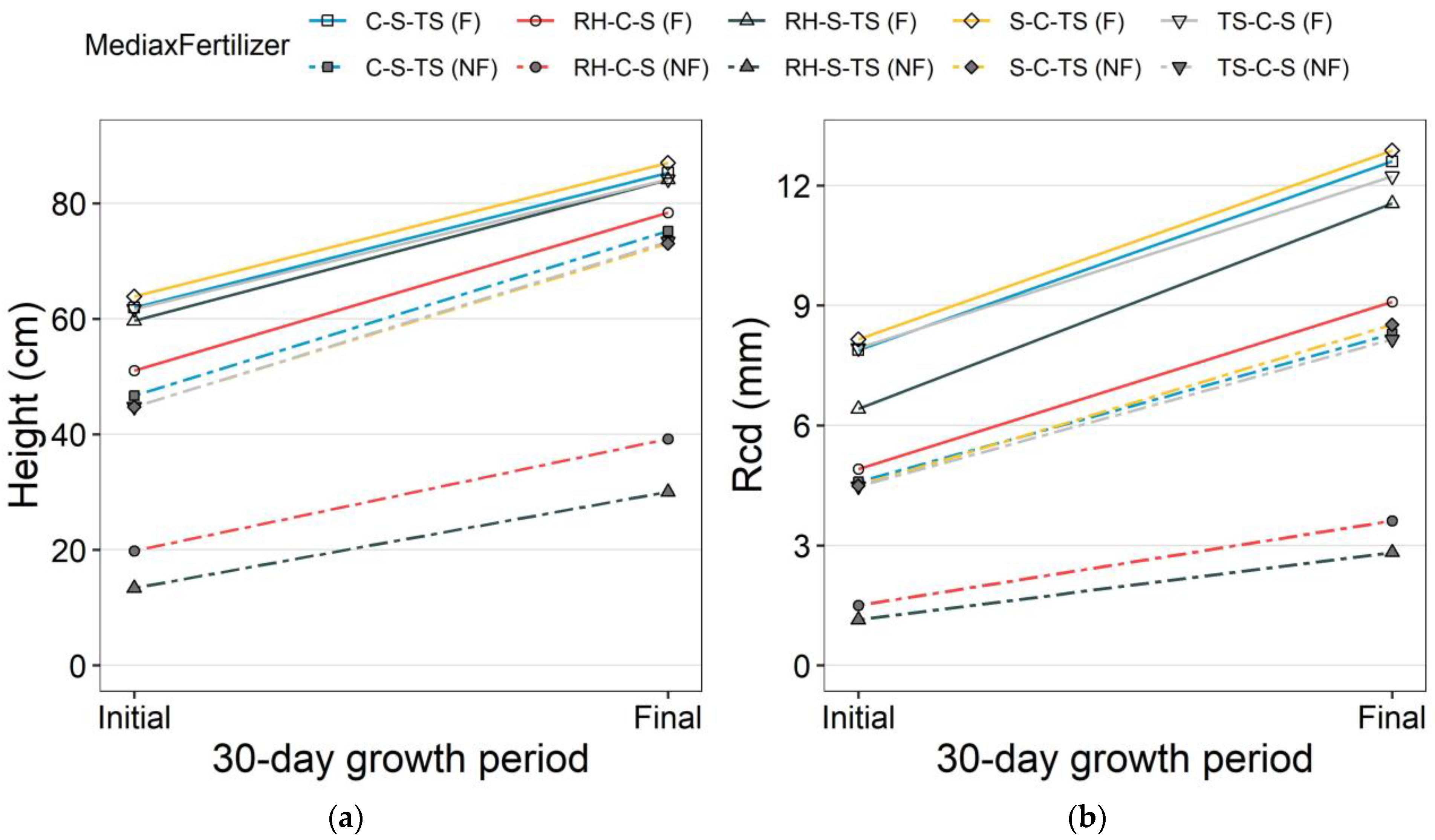
3.1. Nursery Morphological Attributes
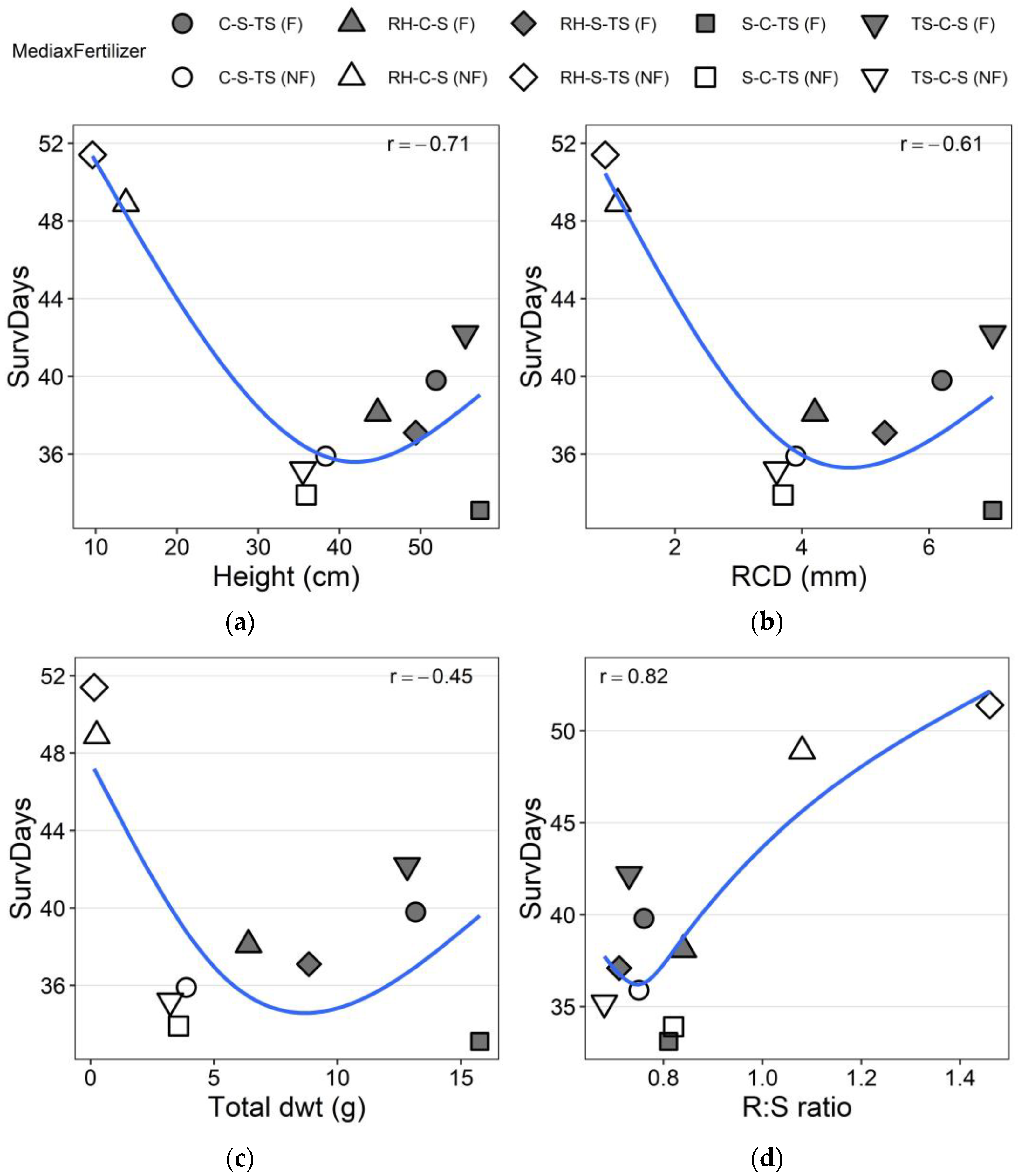
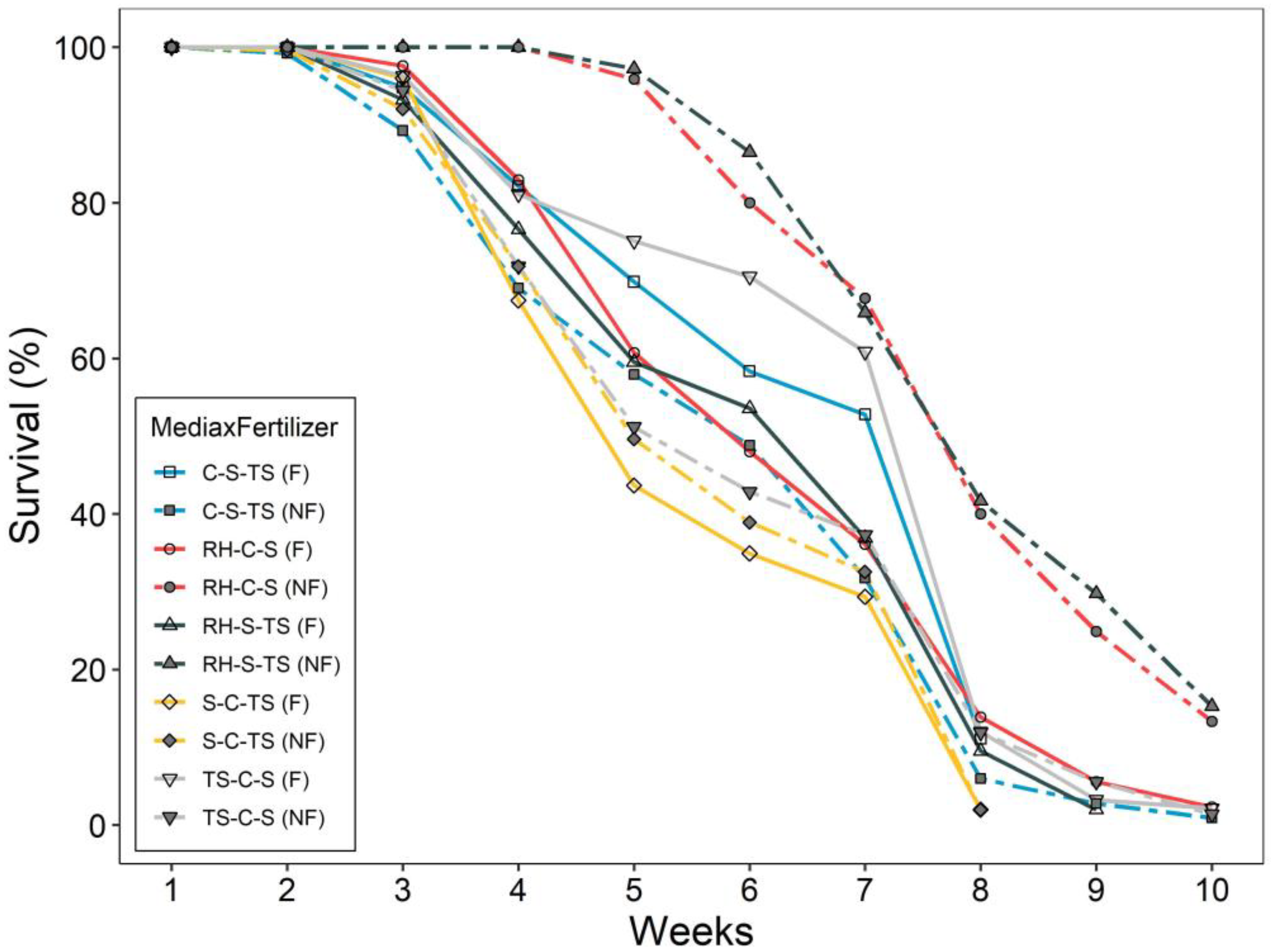
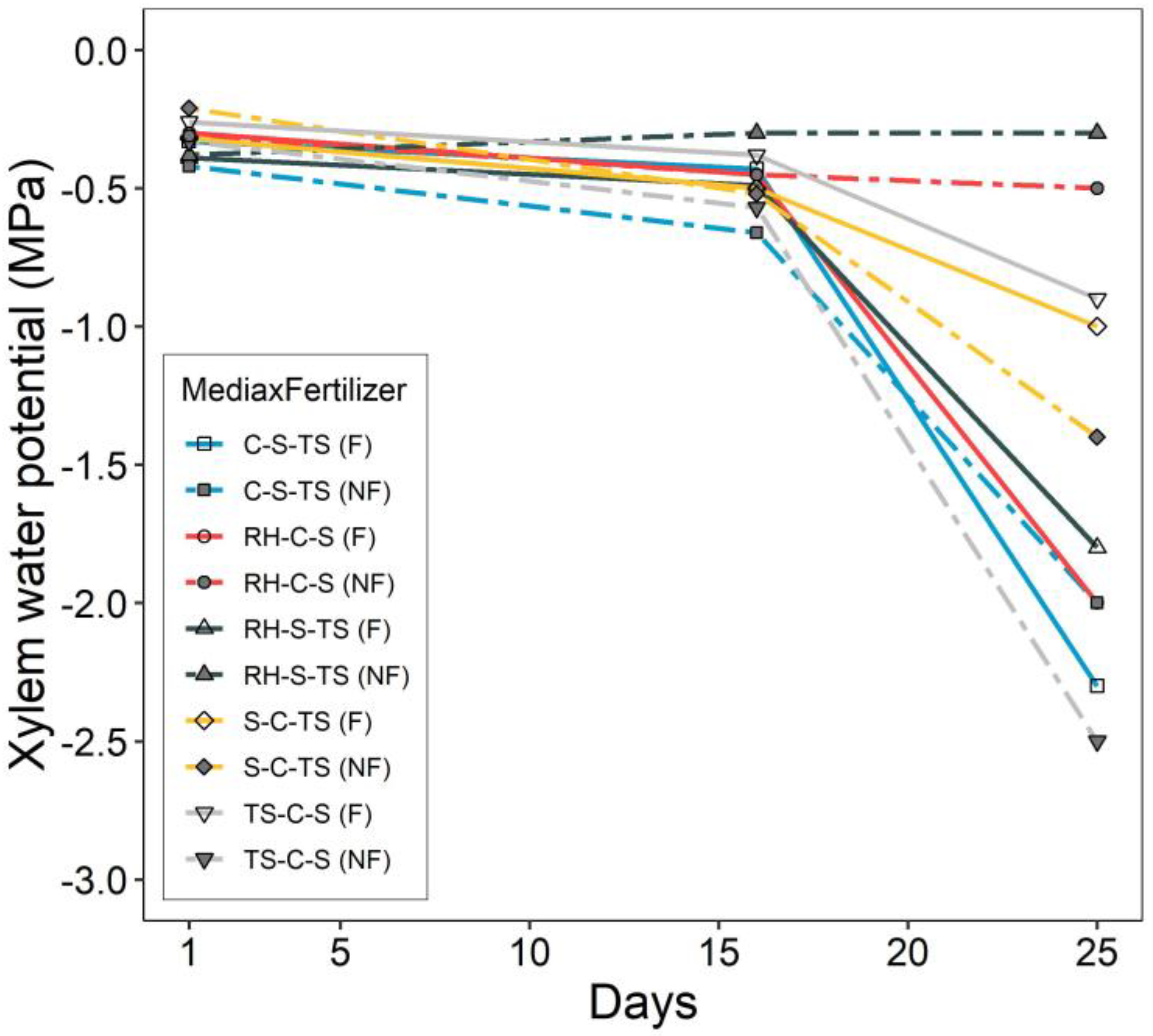
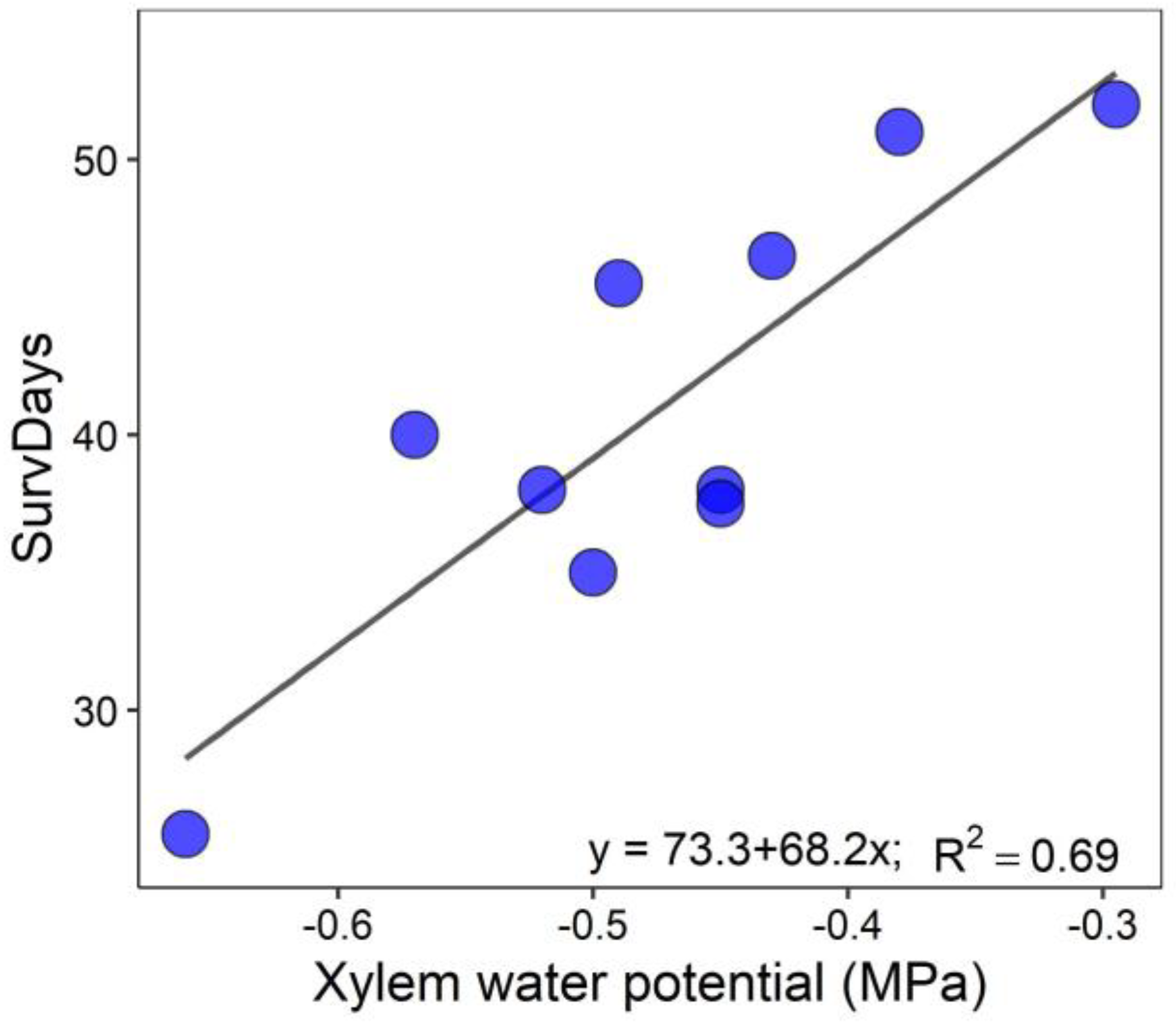
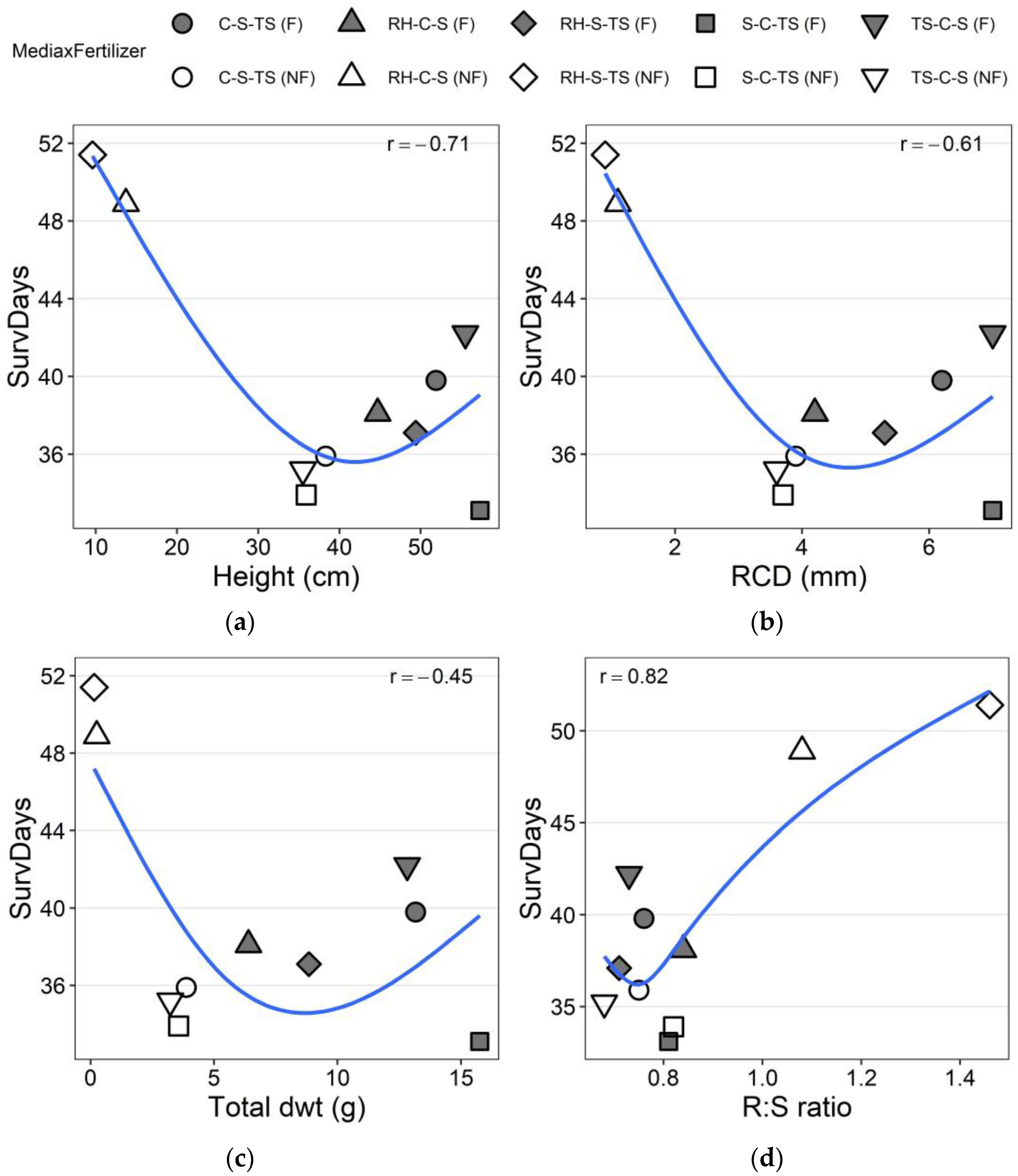
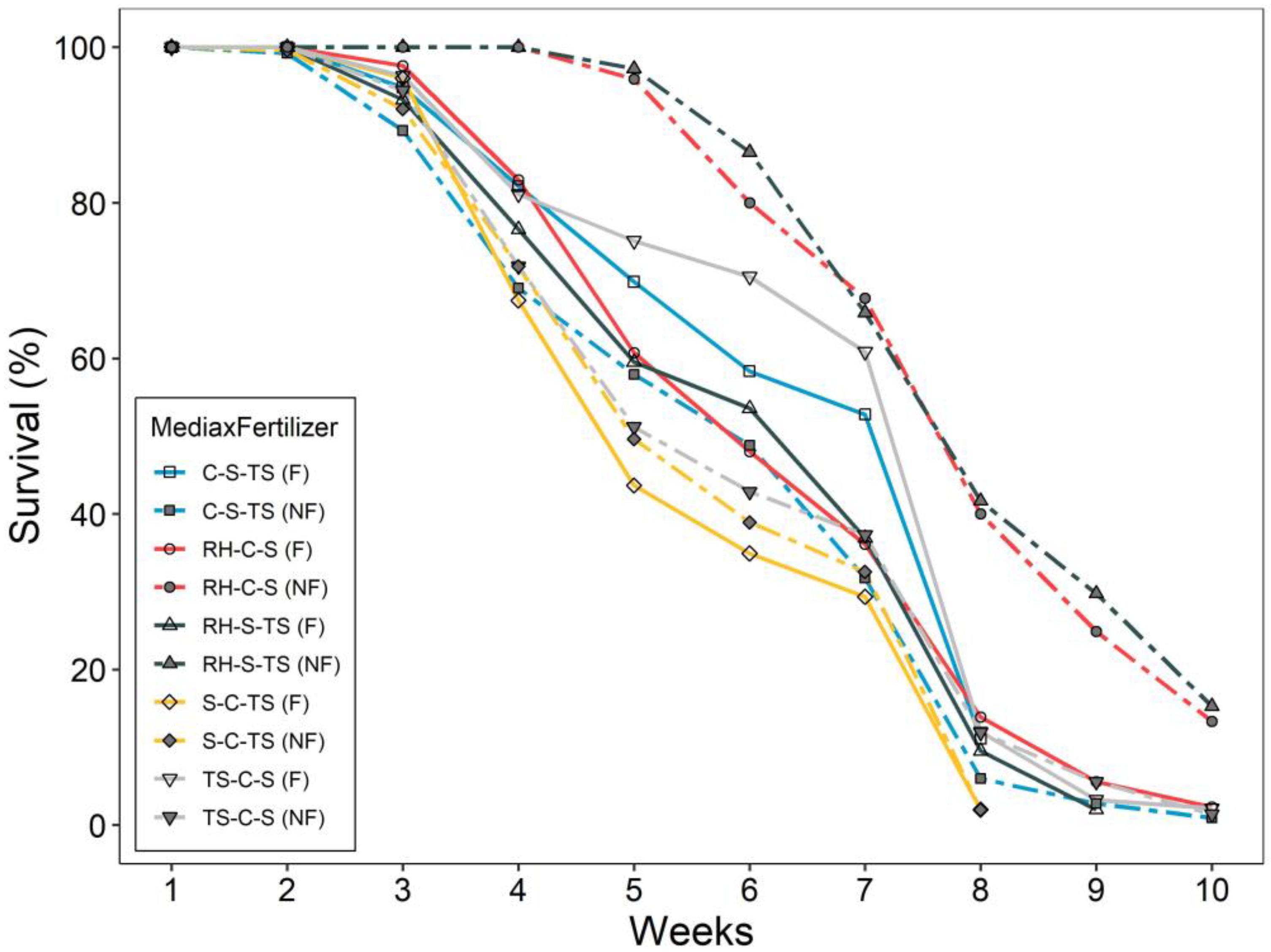
3.2. Drought Test Growth and Survival
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Landis, T.D. The target plant concept. Nurs. Man. Native Plants Guide Tribal Nurseries 2009, 1, 15–31. [Google Scholar]
- Harper, R.J.; Smettem, K.R.J.; Carter, J.O.; McGrath, J.F. Drought deaths in Eucalyptus globulus (Labill.) plantations in relation to soils, geomorphology and climate. Plant Soil 2009, 324, 199. [Google Scholar] [CrossRef]
- Burdett, A.N. Physiological processes in plantation establishment and the development of specifications for forest planting stock. Can. J. For. Res. 1990, 20, 415–427. [Google Scholar] [CrossRef]
- Slot, M.; Poorter, L. Diversity of tropical tree seedling responses to drought. Biotropica 2007, 39, 683–690. [Google Scholar] [CrossRef]
- Davis, A.S.; Jacobs, D.F. Quantifying root system quality of nursery seedlings and relationship to outplanting performance. New For. 2005, 30, 295–311. [Google Scholar] [CrossRef]
- Jacobs, D.F.; Salifu, K.F.; Seifert, J.R. Relative contribution of initial root and shoot morphology in predicting field performance of hardwood seedlings. New For. 2005, 30, 235–251. [Google Scholar] [CrossRef]
- Tsakaldimi, M.; Ganatsas, P.; Jacobs, D.F. Prediction of planted seedling survival of five Mediterranean species based on initial seedling morphology. New For. 2013, 44, 327–339. [Google Scholar] [CrossRef]
- Thompson, B.E. Seedling Morphological Evaluation: What You Can Tell by Looking; Oregon State University: Corvallis, OR, USA, 1985. [Google Scholar]
- Sharma, R.K.; Mason, E.G.; Sorensson, C. Impact of planting stock quality on initial growth and survival of radiata pine clones and modelling initial growth and survival. N. Z. J. For. 2007, 52, 14. [Google Scholar]
- Pawsey, C.K. Survival and early development of Pinus radiata as influenced by size of planting stock. Aust. For. Res. 1972, 5, 13–24. [Google Scholar]
- Mullin, R.E.; Svaton, J. A grading study with white spruce nursery stock. Commonw. For. Rev. 1972, 51, 62–69. [Google Scholar]
- Lamhamed, M.S.; Bernier, P.Y.; Hébert, C. Effect of shoot size on the gas exchange and growth of containerized Picea mariana seedlings under different watering regimes. New For. 1997, 13, 209–223. [Google Scholar] [CrossRef]
- Haase, D.L. Understanding forest seedling quality: Measurements and interpretation. Tree Plant. Notes 2008, 52, 24–30. [Google Scholar]
- Sands, R. Transplanting stress in radiata pine. Aust. For. Res. 1984, 14, 67–72. [Google Scholar]
- Grossnickle, S.C. Why seedlings survive: Influence of plant attributes. New For. 2012, 43, 711–738. [Google Scholar] [CrossRef]
- Harrington, J.T.; Mexal, J.G.; Wagner, A.M.; Parsons, T. The state and challenges of conservation nurseries in Afghanistan. In National Proceedings: Forest and Conservation Nursery Associations—2011. Proc. RMRS-P-68; Haase, D.L., Pinto, J.R., Riley, L.E., Eds.; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2012; pp. 59–64. [Google Scholar]
- Wilkinson, K.M.; Landis, T.D.; Haase, D.L.; Daley, B.F.; Dumroese, R.K. Tropical Nursery Manual: A Guide to Starting and Operating a Nursery for Native and Traditional Plants. In Agriculture Handbook 732; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2014; 376p. [Google Scholar]
- Shrivastava, R.; Nanhorya, R.; Upadhyaya, J.K. Selection of proper potting mixture for root trainer of eucalyptus hybrid. Indian For. 1998, 124, 502–510. [Google Scholar]
- Dumroese, R.K.; Landis, T.D.; Luna, T.; Hernández, G. Simple Methods for Raising Tree and Shrub Seedlings in Afghanistan; Agency for International Development and U.S. Department of Agriculture, Foreign Agriculture Service, Office of Capacity Building and Development: Washington, DC, USA, 2008; 63p.
- Evans, M.R.; Gachukia, M. Fresh parboiled rice hulls serve as an alternative to perlite in greenhouse crop substrates. HortScience 2004, 39, 232–235. [Google Scholar] [CrossRef]
- Whitcomb, C.E. Plant Production in Containers; Lacebark Publications: Stillwater, MN, USA, 1984. [Google Scholar]
- Miller, J.H.; Jones, N. Organic and Compost-Based Growing Media for Tree Seedling Nurseries; World Bank Publications: Washington, DC, USA, 1995; Volume 264. [Google Scholar]
- Tsakaldimi, M. Kenaf (Hibiscus cannabinus L.) core and rice hulls as components of container media for growing Pinus halepensis M. seedlings. Bioresour. Technol. 2006, 97, 1631–1639. [Google Scholar] [CrossRef]
- Antônio, A.D.C.; Alves, J.D.F.; de Faria, B.R.; Almeida, J. Use of brewing biosolids as substrate for production of Eucalyptus seedlings: Analysis of early seedling growth and environmental aspects. Engenharia na Agricultura 2010, 18, 305–309. [Google Scholar] [CrossRef]
- Kratz, D.; Wendling, I. Seedlings of Eucalyptus dunnii grown in renewable substrate. Floresta 2013, 43, 125–135. [Google Scholar] [CrossRef]
- Einert, A.E.; Guidry, G. Rice hulls and hull compost in growing media for container-grown junipers. Ark. Farm Res. 1975, 24, 2. [Google Scholar]
- Van den Driessche, R. Nursery growth of conifer seedlings using fertilizers of different solubilities and application time, and their forest growth. Can. J. For. Res. 1988, 18, 172–180. [Google Scholar] [CrossRef]
- Duryea, M.L.; Landis, T.D. Forest Nursery Manual: Production of Bareroot Seedlings; Springer Science & Business Media: Berlin, Germany, 2012; Volume 11. [Google Scholar]
- Oliet, J.A.; Planelles, R.; Artero, F.; Valverde, R.; Jacobs, D.F.; Segura, M.L. Field performance of Pinus halepensis planted in Mediterranean arid conditions: Relative influence of seedling morphology and mineral nutrition. New For. 2009, 37, 313–331. [Google Scholar] [CrossRef]
- Chamshama, S.A.O.; Hall, J.B. Effects of nursery treatments on Eucalyptus camaldulensis field establishment and early growth at Mafiga, Morogoro, Tanzania. For. Ecol. Manag. 1987, 21, 91–108. [Google Scholar] [CrossRef]
- Stahl, J.; Ernani, P.R.; Gatiboni, L.C.; Chaves, D.M.; Neves, C.U. Dry matter yield and nutritional efficiency of Eucalyptus benthamii and Eucalyptus dunnii clones due to addition of phosphorus rates to the soil. Cienc. Florest. 2013, 23, 287–295. [Google Scholar]
- Luis, V.C.; Puértolas, J.; Climent, J.; Peters, J.; González-Rodríguez, Á.M.; Morales, D.; Jiménez, M.S. Nursery fertilization enhances survival and physiological status in Canary Island pine (Pinus canariensis) seedlings planted in a semiarid environment. Eur. J. For. Res. 2009, 128, 221–229. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Peñuelas, J.L.; Nicolás-Peragón, J.L.; Benito, L.F.; Domínguez-Lerena, S. Is nitrogen fertilization in the nursery a suitable tool for enhancing the performance of Mediterranean oak plantations? New For. 2013, 44, 733–751. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Planelles, R.; Enrıquez, E.; Rubira, J.P. Nursery cultivation regimes, plant functional attributes, and field performance relationships in the Mediterranean oak Quercus ilex L. For. Ecol. Manag. 2004, 196, 257–266. [Google Scholar] [CrossRef]
- Trubat, R.; Cortina, J.; Vilagrosa, A. Nursery fertilization affects seedling traits but not field performance in Quercus suber L. J. Arid Environ. 2010, 74, 491–497. [Google Scholar] [CrossRef]
- Pharis, R.P.; Kramer, P.J. The effects of nitrogen and drought on loblolly pine seedlings. For. Sci. 1964, 10, 143–150. [Google Scholar]
- Cromer, R.N.; Cameron, D.M.; Rance, S.J.; Ryan, P.A.; Brown, M. Response to nutrients in Eucalyptus grandis. 1. Biomass accumulation. For. Ecol. Manag. 1993, 62, 211–230. [Google Scholar] [CrossRef]
- Butnor, J.R.; Johnsen, K.H.; Anderson, P.H.; Hall, K.B.; Halman, J.M.; Hawley, G.J.; Maier, C.A.; Schaberg, P.G. Growth, photosynthesis, and cold tolerance of Eucalyptus benthamii planted in the piedmont of North Carolina. For. Sci. 2018, 65, 59–67. [Google Scholar] [CrossRef]
- Stanturf, J.A.; Vance, E.D.; Fox, T.R.; Kirst, M. Eucalyptus beyond its native range: Environmental issues in exotic bioenergy plantations. Int. J. For. Res. 2013, 2013. [Google Scholar] [CrossRef]
- Davidson, J. Ecological aspects of Eucalyptus plantations. In Proceedings of the Regional Expert Consultation on Eucalyptus, RAPA/FAO, Bangkok, Thailand, 4–8 October 1993; Volume 1, pp. 35–60. [Google Scholar]
- Madgwick, H.A.I.; Oliver, G.R.; Frederick, D.J.; Tew, D.T. Estimating the dry weights of Eucalyptus trees—Central North Island, New Zealand. Bioresour. Technol. 1991, 37, 111–114. [Google Scholar] [CrossRef]
- Rook, D.A. Seedling development and physiology in relation to mineral nutrition. Conifer Seedl. Miner. Nutr. 1990, 85, 86–105. [Google Scholar]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Qaisar, K.N.; Khan, P.A.; Khan, M.A. Container type and growing media for raising quality seedling stock of Cedrus deodara. Indian J. For. India 2008, 31, 383–388. [Google Scholar]
- Dickson, A.; Leaf, A.L.; Hosner, J.F. Quality appraisal of white spruce and white pine seedling stock in nurseries. For. Chron. 1960, 36, 10–13. [Google Scholar] [CrossRef]
- Isaac, R.A.; Johnson, W.C. Elemental analysis of plant tissue by plasma emission spectroscopy: Collaborative study. J. Assoc. Anal. Chem. US 1985, 68, v499–v505. [Google Scholar]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Cleary, B.D.; Zaerr, J.B. Pressure chamber techniques for monitoring and evaluating seedling water status. N. Z. J. For. Sci. 1980, 10, 133–141. [Google Scholar]
- Dueitt, S.D. Rice Hulls as a Substitute for Sphagnum Peat Moss in a Sphagnum Peat-Based Greenhouse Media. Master’s Thesis, Mississippi State University, Starkville, MS, USA, 1995. [Google Scholar]
- Tuttle, C.L.; South, D.B.; Golden, M.S.; Meldahl, R.S. Relationship between initial seedling height and survival and growth of loblolly pine seedlings planted during a droughty year. South. J. Appl. For. 1987, 11, 139–143. [Google Scholar]
- McTague, J.P.; Tinus, R.W. The effects of seedling quality and forest site weather on field survival of ponderosa pine. Tree Plant. Notes 1996, 47, 16–23. [Google Scholar]
- Grossnickle, S.C. Importance of root growth in overcoming planting stress. New For. 2005, 30, 273–294. [Google Scholar] [CrossRef]
- Ivetić, V.; Grossnickle, S.; Škorić, M. Forecasting the field performance of Austrian pine seedlings using morphological attributes. IForest-Biogeosci. For. 2016, 10, e1–e9. [Google Scholar] [CrossRef]
- Khurram, S.; Burney, O.T.; Morrissey, R.C.; Jacobs, D.F. Bottles to trees: Plastic beverage bottles as an alternative nursery growing container for reforestation in developing countries. PLoS ONE 2017, 12, e0177904. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | Substrates (%) | Fertilization | Treatment | |||
|---|---|---|---|---|---|---|
| Top Soil | Sand | Compost | Rice Hull | |||
| C-S-TS | 17 | 33 | 50 | – | F | C-S-TS(F) |
| NF | C-S-TS(NF) | |||||
| TS-C-S | 50 | 17 | 33 | – | F | TS-C-S(F) |
| NF | TS-C-S(NF) | |||||
| S-C-TS | 25 | 50 | 25 | – | F | S-C-TS(F) |
| NF | S-C-TS(NF) | |||||
| RH-S-TS | 17 | 33 | – | 50 | F | RH-S-TS(F) |
| NF | RH-S-TS(NF) | |||||
| RH-C-S | – | 17 | 33 | 50 | F | RH-C-S(F) |
| NF | RH-C-S(NF) | |||||
| Medium † | Bag Weight (g) | B.D (g/cm3) | P.D (g/cm3) | Porosity (%) | M.C (%) | Drainage (mL) |
|---|---|---|---|---|---|---|
| C-S-TS | 972 | 1.1 | 1.4 | 23.4 | 41.0 | 355.0 |
| TS-C-S | 1016 | 1.3 | 1.5 | 10.4 | 32.4 | 118.8 |
| S-C-TS | 1222 | 1.3 | 1.6 | 16.9 | 28.7 | 245.0 |
| RH-S-TS | 794 | 0.8 | 1.3 | 33.8 | 30.5 | 365.0 |
| RH-C-S | 670 | 0.5 | 0.7 | 32.5 | 73.8 | 377.5 |
| Response Variable | Media | Fertilizer | Media × Fertilizer |
|---|---|---|---|
| Height (cm) | <0.0001 | <0.0001 | <0.0001 |
| RCD (mm) | <0.0001 | <0.0001 | <0.0001 |
| Root dwt (g) | <0.0001 | <0.0001 | 0.0080 |
| Shoot dwt (g) | <0.0001 | <0.0001 | 0.0020 |
| Total dwt (g) | <0.0001 | <0.0001 | 0.0030 |
| R:S Ratio | <0.0001 | <0.0001 | <0.0001 |
| DQI | <0.0001 | <0.0001 | <0.0001 |
| FOLRs | <0.0001 | <0.0001 | 0.0030 |
| Height (cm)-RG | <0.0001 | <0.0001 | <0.0001 |
| RCD (mm)-RG | <0.0001 | <0.0001 | 0.0030 |
| Height (cm)-AG | <0.0001 | 0.9400 | <0.0001 |
| RCD (mm)-AG | <0.0001 | <0.0001 | <0.0001 |
| SurvDays | <0.0001 | 0.0030 | <0.0001 |
| Treatments | Height (cm) | RCD (mm) | Root dwt (g) | Shoot dwt (g) | Total dwt (g) | R:S Ratio | DQI | FOLRs |
|---|---|---|---|---|---|---|---|---|
| C-S-TS (F) | 51.9 (1.69) ab ‡ | 6.2 (0.30) a | 5.93 (0.89) ab | 7.22 (0.94) a | 13.16 (1.82) ab | 0.76 (0.04) c | 1.64 (0.28) ab | 68 (4.0) bc |
| TS-C-S (F) | 55.5 (1.41) a | 7.0 (0.27) a | 5.59 (0.89) ab | 7.24 (0.93) a | 12.82 (1.80) ab | 0.73 (0.04) c | 1.50 (0.27) ab | 79 (4.4) ab |
| S-C-TS (F) | 57.3 (1.85) a | 7.0 (0.33) a | 7.28 (0.94) a | 8.48 (0.89) a | 15.76 (1.80) a | 0.81 (0.04) c | 2.02 (0.31) a | 92 (5.4) a |
| RH-S-TS (F) | 49.4 (2.46) bc | 5.3 (0.26) b | 3.66 (0.34) b | 5.18 (0.45) ab | 8.84 (0.77) ab | 0.71 (0.03) c | 0.88 (0.10) bc | 69 (5.6) abc |
| RH-C-S (F) | 44.7 (1.22) c | 4.2 (0.12) c | 2.92 (0.24) bc | 3.47 (0.29) bc | 6.39 (0.52) bc | 0.84 (0.03) c | 0.59 (0.06) dc | 57 (3.7) bc |
| C-S-TS (NF) | 38.3 (1.42) d | 3.9 (0.17) c | 1.70 (0.20) cd | 2.17 (0.24) cd | 3.87 (0.43) dc | 0.75 (0.02) c | 0.34 (0.05) de | 48 (3.4) c |
| TS-C-S (NF) | 35.5 (1.50) d | 3.6 (0.12) c | 1.37 (0.13) d | 1.85 (0.15) d | 3.22 (0.28) d | 0.68 (0.03) c | 0.28 (0.03) e | 48 (2.1) c |
| S-C-TS (NF) | 35.9 (2.10) d | 3.7 (0.16) c | 1.63 (0.19) d | 1.94 (0.23) d | 3.56 (0.42) d | 0.82 (0.03) c | 0.32 (0.05) e | 55 (2.2) c |
| RH-S-TS (NF) | 9.6 (0.94) e | 0.9 (0.13) d | 0.08 (0.01) e | 0.06 (0.01) e | 0.14 (0.02) e | 1.46 (0.09) a | 0.01 (0.00) f | 15 (1.1) d |
| RH-C-S (NF) | 13.7 (1.25) e | 1.1 (0.09) d | 0.12 (0.02) e | 0.12 (0.02) f | 0.24 (0.05) e | 1.08 (0.04) b | 0.02 (0.00) f | 18 (2.5) d |
| Treatments | Height-RG (%) | RCD-RG (%) | Height-AG (cm) | RCD-AG (mm) | SurvDays |
|---|---|---|---|---|---|
| C-S-TS (F) | 38.1 (1.53) d ‡ | 65.8 (5.11) b | 23.3 (0.78) bcde | 4.8 (0.23) ab | 39.8 (2.12) bc |
| TS-C-S (F) | 37.8 (2.49) d | 61.5 (6.20) b | 22.4 (1.02) de | 4.3 (0.21) abcd | 42.2 (2.35) bc |
| S-C-TS (F) | 37.6 (2.15) d | 62.8 (4.37) b | 23.1 (0.90) cde | 4.7 (0.22) abc | 33.1 (2.01) c |
| RH-S-TS (F) | 42.2 (2.07) d | 89.7 (9.69) b | 24.4 (0.88) abcde | 5.1 (0.25) a | 37.1 (2.20) c |
| RH-C-S (F) | 56.7 (4.67) cd | 87.9 (4.96) b | 27.3 (1.60) abcd | 4.2 (0.21) abcd | 38.1 (2.22) c |
| C-S-TS (NF) | 64.2 (3.77) c | 87.1 (5.92) b | 28.4 (1.15) ab | 3.7 (0.19) cd | 35.9 (2.30) c |
| TS-C-S (NF) | 67.0 (3.19) c | 84.5 (4.70) b | 28.6 (0.99) a | 3.7 (0.18) d | 35.2 (2.39) c |
| S-C-TS (NF) | 67.5 (4.31) c | 93.9 (5.55) b | 28.3 (0.99) abc | 4.0 (0.18) bcd | 33.9 (2.07) c |
| RH-S-TS (NF) | 132.0 (9.98) a | 166.3 (16.16) a | 16.7 (1.40) f | 1.7 (0.16) e | 51.4 (1.90) a |
| RH-C-S (NF) | 106.0 (8.04) b | 155.1 (15.11) a | 19.4 (1.41) ef | 2.1 (0.18) e | 48.9 (1.93) ab |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shalizi, M.N.; Goldfarb, B.; Burney, O.T.; Shear, T.H. Effects of Five Growing Media and Two Fertilizer Levels on Polybag—Raised Camden Whitegum (Eucalyptus benthamii Maiden & Cambage) Seedling Morphology and Drought Hardiness. Forests 2019, 10, 543. https://doi.org/10.3390/f10070543
Shalizi MN, Goldfarb B, Burney OT, Shear TH. Effects of Five Growing Media and Two Fertilizer Levels on Polybag—Raised Camden Whitegum (Eucalyptus benthamii Maiden & Cambage) Seedling Morphology and Drought Hardiness. Forests. 2019; 10(7):543. https://doi.org/10.3390/f10070543
Chicago/Turabian StyleShalizi, Mohammad Nasir, Barry Goldfarb, Owen Thomas Burney, and Theodore Henry Shear. 2019. "Effects of Five Growing Media and Two Fertilizer Levels on Polybag—Raised Camden Whitegum (Eucalyptus benthamii Maiden & Cambage) Seedling Morphology and Drought Hardiness" Forests 10, no. 7: 543. https://doi.org/10.3390/f10070543
APA StyleShalizi, M. N., Goldfarb, B., Burney, O. T., & Shear, T. H. (2019). Effects of Five Growing Media and Two Fertilizer Levels on Polybag—Raised Camden Whitegum (Eucalyptus benthamii Maiden & Cambage) Seedling Morphology and Drought Hardiness. Forests, 10(7), 543. https://doi.org/10.3390/f10070543